- Антропогенез эволюция человека

Содержание
- 2. Царство: Животные Тип: Хордовые Класс: Млекопитающие Отряд: Приматы Семейство: Гоминиды
- 3. подсемейство Понгины Ponginae орангутаны, Pongo подсемейство Гоминиды Homininae триба Гоминини Hominini гориллы, Gorilla шимпанзе, Pan обыкновенный
- 4. семейство гоминид исторически были выделены три важнейших системы, считающиеся истинно гоминидными. Эти системы были названы гоминидной
- 5. Эволюция мозга Мозг грацильных австралопитеков, 4 - 2,5 м. назад, отличался от мозга шимпанзе большей длиной,
- 6. В сравнении же с массивными австралопитеками, грацильные, вероятно, имели более развитый зрительный анализатор, что, возможно, в
- 7. массивные австралопитеки, в сравнении с грацильными, обладали лучшей координацией движений, но меньшим контролем за эмоциями и
- 8. Архантроп Homo erectus Объём мозга достигал значений современного человека, В увеличении мозга -резко возросшая ширина мозга.
- 9. В теменной доле бурно расширялась надкраевая часть. Височная доля расширялась в задней части и удлинялась по
- 10. У архантропов в сравнении с "ранними Homo" наверняка был усилен сознательный контроль эмоций и, по всей
- 11. Homo heidelbergensis Форма мозга H. Heidelber-gensis свидетельствуе о резком прогрессе в области контроля за движениями, в
- 12. Все указанные прогрессивные изменения развития мозга проявились в культуре и особенностях образа жизни пре-палеоантропов. Они заселили
- 13. Homo neanderthalensis или палеоантропы У неандертальцев ширина мозга крайне велика, максимальна для всех групп гоминид. Очень
- 14. Мозг H. neanderthalensis отличался от мозга современного человека, вероятно, большим развитием подкорковых центров подсознательного контроля за
- 15. По-видимому, можно констатировать существенное отставание палеоантропов от современного человека в плане возможностей контроля и координации движений.
- 16. пропорции древнейших прямоходящих существ – Ardipithecus ramidus из Арамиса – ещё настолько напоминают обезьяньи, что закономерности
- 18. Sahelanthropus tchadensis Сахелантроп 7.0 млн.л.н. - 6.0 млн.л.н. 2002 Чад
- 19. Останки найдены в Республике Чад в Торос-Меналла. Шесть находок гоминид включили полный череп, получивший имя Тумай
- 21. Orrorin tugenensis Оррорин 6.0 млн.л.н. 2001 Кения
- 23. В 2001 г. Б. Сеню и М. Пикфорд обнаружила в Туген Хиллс 12 костей гоминид, в
- 24. Ardipithecus kadabba Ардипитек кадабба 5.5 млн.л.н. 2001 Эфиопия
- 25. Ардипитек кадабба (Ardipithecus ramidus kadabba) представляет ещё один род поздномиоценовых гоминид, населявших тогда нашу Землю. Его
- 26. Ardipithecus kadabba одно из древнейших прямоходящих существ, от которого сохранились кости ног и рук. Находки включают
- 27. Ardipithecus ramidus Ардипитек рамидус 4 млн.л.н. 1995 Эфиопия
- 28. Ардипитеки (лат. Ardipithecus) — древний род гоминид, который жил в раннем плиоцене. Поскольку род ардипитеки имеет
- 29. Найденные в Арамисе в 1995 гг. остатки гоминид включили фрагменты черепов, нижней челюсти детеныша, большое количество
- 30. Australopithecus anamensis Австралопитек анамский 4.2 млн.л.н. - 3.9 млн.л.н. 1994 Кения
- 31. Австралопитек анамский (лат. Australopithecus anamensis) — ископаемый вид рода австралопитеков, обитавший в Восточной Африке около 4
- 32. Australopithecus afarensis Австралопитек афарский 4.0 млн.л.н. - 2.5 млн.л.н. 1978 Кения, Эфиопия, Танзания
- 33. Австралопитек афарский (лат. Australopithecus afarensis) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид, живший
- 34. Морфологические особенности хадарских австралопитеков, благодаря представительности материала, изучены очень хорошо и могут служить своеобразным эталоном для
- 35. Из особенностей строения зубов замечательны значительные размеры резцов и большая разница размеров резцов I1 и I2,
- 36. Морфология стопы A. afarensis также свидетельствует о разнообразии видов локомоции, использовавшихся этим видом; в этом отношении
- 37. Kenyanthropus platyops 3.5 млн.л.н. - 3.2 млн.л.н. 2001 Кения
- 38. Кениантропы (лат. Kenyanthropus) — древний род гоминид) — древний род гоминид, живший в плиоцене около 3,5-3,2
- 39. Australopithecus africanu Австралопитек африканский 3.5 млн.л.н. - 2.4 млн.л.н. 1925 ЮАР
- 40. Австралопитек африканский (лат. Australopithecus africanus) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид, живший
- 41. Пропорции, видимо, были промежуточными между пропорциями шимпанзе и современного человека. Общие размеры тела A. africanus варьировали
- 42. Череп A. africanus в среднем больше, чем у A. afarensis; наклон лба сравнительно небольшой, затылок тоже
- 43. Australopithecus bahrelghazali Австралопитек бахр-эль-газальский 3.5 млн.л.н. - 3.0 млн.л.н 1996 Чад
- 44. Австралопитек бахр-эль-газальский (лат. Australopithecus bahrelghazali) — предположительно, самостоятельный вид австралопитека) — предположительно, самостоятельный вид австралопитека, семейство
- 45. Всего обнаружены один фрагмент верхней и один – нижней челюстей, а также изолированный зуб. В целом
- 46. Australopithecus garhi Австралопитек гари 2.5 млн.л.н. 1999 Эфиопия
- 47. Австралопитек гари (лат. Australopithecus garhi) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид) —
- 48. «Garhi» в переводе с афарского означает «удивительный» или «сюрприз». Это поздний грацильный австралопитек с датировкой около
- 49. Australopithecus sediba Австралопитек седиба 2.0 млн.л.н. - 1.8 млн.л.н. 2010 ЮАР
- 50. Австралопитек седиба (Australopithecus sediba) — вид австралопитеков) — вид австралопитеков, известный по останкам, возраст которых оценивается
- 51. Интересно, что находка останков Australopithecus sediba сделана с использованием популярного сервиса Google Earth, позволившего выявить местонахождения
- 52. Данный вид отнесли всё-таки к роду Australopithecus, однако некоторые исследователи не согласны с таким решением, и
- 53. Однако, древнейшие "ранние Homo" и в Южной, и в Восточной Африке имеют гораздо большие – как
- 54. Paranthropus aethiopicus Парантроп эфиопский 2.5 млн.л.н. 1968 Кения, Эфиопия
- 55. Парантроп эфиопский (лат. Paranthropus aethiopicus) — ископаемый вид рода парантропов) — ископаемый вид рода парантропов («массивных
- 56. (Если быть точным, история данного вида довольно запутанная. Первоначально род и видParaustralopithecus aethiopicus Arambourg et Coppens,
- 57. Paranthropus boisei Парантроп Бойса 2.5 млн.л.н. - 1.0 млн.л.н. 1959 Кения, Танзания
- 58. Парантроп Бойса (Paranthropus boisei Mary Leakey, 1959), прежде также зинджантроп (Zinjanthropus boisei), — самый массивный вид
- 59. Череп P. boisei хорошо известен по множеству находок хорошей сохранности, с датировками в пределах 1,7-1,9 млн.л.
- 60. Строение мозга P. boisei довольно примитивное.Эндокраны характеризуются сильно суженной лобной долей даже по сравнению с более
- 61. Относительные размеры передних и заклыковых зубов P. boisei резко отличаются от пропорций, характерных для грацильных австралопитеков.
- 62. Paranthropus robustus Парантроп массивный 2.5 млн.л.н. - 900 тыс.л.н. 1939 ЮАР
- 63. В 1938 г. Роберт Брум проводил раскопки в пещерах Стеркфонтейна. 8 июня 1938 г. школьник Герт
- 64. Эндокран P. robustus имеет примитивную форму, мало отличающуюся от формы человекообразных обезьян. Верхние конечности P. robustus,
- 65. Homo habilis Человек умелый 2.3 млн.л.н. - 1.5 млн.л.н. 1964 Кения, ЮАР
- 66. Челове́к уме́лый (лат. Homo habilis) — высокоразвитый австралопитек или первый представитель рода Homo. Обнаружен археологами Лики
- 67. Человек умелый был первым из наших предков, который стал регулярно изготавливать каменные орудия труда и перешёл
- 68. Лицевой скелет H. habilis более прогрессивный, чем у грацильных австралопитеков. Горизонтальная уплощенность лица в большинстве случаев
- 69. Homo rudolfensis Человек рудольфийский 2.3 млн.л.н. - 1.5 млн.л.н. 1978 Кения
- 70. Человек рудольфский (лат. Homo rudolfensis) — вымерший «старейший» вид людей, представляющий (согласно традиционным представлениям) переходную ступень
- 71. Наиболее характерным отличием от Homo habilis является большая массивность и большие размеры зубов у Homo rudolfensis
- 72. Homo ergaster Человек работающий 1.8 млн.л.н. - 1.4 млн.л.н. 1975 Кения, Танзания, Грузия
- 73. Homo ergaster (Человек работающий) — ископаемый вид людей, появившийся в Африке) — ископаемый вид людей, появившийся
- 74. Обычно под термином Homo ergaster понимаются ранние сравнительно массивные восточноафриканские (точнее – из области озера Туркана)
- 75. Черепа H.ergaster низкие и массивные, кости свода толстые. Лобная кость убегающая назад, слабо изогнутая. Надглазничные дуги
- 76. Homo erectus Человек прямоходящий 1.5 млн.л.н. - 400 тыс.л.н. 1894 Ява, Китай, Греция, Испания, и т.д.
- 77. Эректус (лат. Homo erectus — человек прямоходящий; устар. архантропы) — ископаемый вид людей, который рассматривают как
- 78. Наиболее массивные представители древнейших людей. Являясь уже несомненными людьми, Homo erectus всё же сильно отличались от
- 79. Homo heidelbergensis Человек гейдельбергский 800 тыс.л.н. - 130 тыс.л.н. 1908 Кения, Эфиопия, Германия, и т.д.
- 80. Гейдельбергский человек (лат. Homo heidelbergensis) — ископаемый вид людей, европейская разновидность человека прямоходящего (родственный восточноазиатскому синантропу
- 81. Относительно поздняя форма, близкая Homo erectus. От архантропов отличались некоторыми прогрессивными (и иногда специализированно-неандертальскими) признаками, в
- 82. Homo helmei 500 тыс.л.н. – 130 тыс.л.н. 1935 Африка
- 83. Африканские формы палеоантропов, не имеющие специализированных признаков европейских неандертальцев. Часто их относят в "ранним архаичным Homo
- 84. Homo neanderthalensis Неандерталец 130 тыс.л.н. - 28 тыс.л.н. 1864 Африка, Европа, Ближний Восток
- 85. Неандерта́лец, человек неандертальский (лат. Homo neanderthalensis или Homo sapiens neanderthalensis; в советской литературе также именовался палеоантропом)
- 86. Специализированный к хищничеству вариант человека. Обладали множеством вполне человеческих черт строения и поведения, но всё же
- 87. Homo floresiensis Человек флоресский 95 тыс.л.н. - 12 тыс.л.н. 2004 Флорес
- 88. Человек флоресский (Homo floresiensis BROWN ET AL., 2004) — возможно, карликовый ископаемый вид людей. Именуются также
- 89. Наилучшим образом сохранился почти полный скелет Лянг Буа 1 (LB1), найденный в сентябре 2003 г. и
- 90. Homo sapiens sapiens Человек разумный 100 тыс.л.н. 1758 Планета Земля
- 91. Челове́к разу́мный (лат. Homo sapiens; в русскоязычных текстах встречается также написание Хомо Сапиенс илиГомо Сапиенс —
- 92. Эволюционное дерево человека Следующая схема была подготовлена нашим Научным редактором Станиславом ДробышевскимСледующая схема была подготовлена нашим
- 94. Скачать презентацию

Царство: Животные
Тип: Хордовые
Класс: Млекопитающие
Отряд: Приматы
Семейство: Гоминиды
Царство: Животные
Тип: Хордовые
Класс: Млекопитающие
Отряд: Приматы
Семейство: Гоминиды

подсемейство Понгины Ponginae
орангутаны, Pongo
подсемейство Гоминиды Homininae
триба Гоминини Hominini
гориллы, Gorilla
шимпанзе, Pan
обыкновенный шимпанзе, Pan troglodytes
бонобо или карликовый шимпанзе, Pan paniscus
люди, Homo
человек разумный, Homo sapiens.
подсемейство Понгины Ponginae
орангутаны, Pongo
подсемейство Гоминиды Homininae
триба Гоминини Hominini
гориллы, Gorilla
шимпанзе, Pan
обыкновенный шимпанзе, Pan troglodytes
бонобо или карликовый шимпанзе, Pan paniscus
люди, Homo
человек разумный, Homo sapiens.

семейство гоминид
исторически были выделены три важнейших системы, считающиеся истинно гоминидными.
семейство гоминид
исторически были выделены три важнейших системы, считающиеся истинно гоминидными.

Эволюция мозга
Мозг грацильных австралопитеков, 4 - 2,5 м. назад, отличался от
Эволюция мозга
Мозг грацильных австралопитеков, 4 - 2,5 м. назад, отличался от

В сравнении же с массивными австралопитеками, грацильные, вероятно, имели более развитый
В сравнении же с массивными австралопитеками, грацильные, вероятно, имели более развитый

массивные австралопитеки, в сравнении с грацильными, обладали лучшей координацией движений, но
массивные австралопитеки, в сравнении с грацильными, обладали лучшей координацией движений, но

Архантроп Homo erectus
Объём мозга достигал значений современного человека, В увеличении мозга
Архантроп Homo erectus
Объём мозга достигал значений современного человека, В увеличении мозга

В теменной доле бурно расширялась надкраевая часть. Височная доля расширялась в
В теменной доле бурно расширялась надкраевая часть. Височная доля расширялась в

У архантропов в сравнении с "ранними Homo" наверняка был усилен сознательный контроль
У архантропов в сравнении с "ранними Homo" наверняка был усилен сознательный контроль

Homo heidelbergensis
Форма мозга H. Heidelber-gensis свидетельствуе о резком прогрессе в области контроля за движениями,
Homo heidelbergensis
Форма мозга H. Heidelber-gensis свидетельствуе о резком прогрессе в области контроля за движениями,

Все указанные прогрессивные изменения развития мозга проявились в культуре и особенностях
Все указанные прогрессивные изменения развития мозга проявились в культуре и особенностях

Homo neanderthalensis или палеоантропы
У неандертальцев ширина мозга крайне велика, максимальна для всех
Homo neanderthalensis или палеоантропы
У неандертальцев ширина мозга крайне велика, максимальна для всех

Мозг H. neanderthalensis отличался от мозга современного человека, вероятно, большим развитием подкорковых центров
Мозг H. neanderthalensis отличался от мозга современного человека, вероятно, большим развитием подкорковых центров

По-видимому, можно констатировать существенное отставание палеоантропов от современного человека в плане
По-видимому, можно констатировать существенное отставание палеоантропов от современного человека в плане

пропорции древнейших прямоходящих существ – Ardipithecus ramidus из Арамиса – ещё настолько напоминают обезьяньи, что
пропорции древнейших прямоходящих существ – Ardipithecus ramidus из Арамиса – ещё настолько напоминают обезьяньи, что

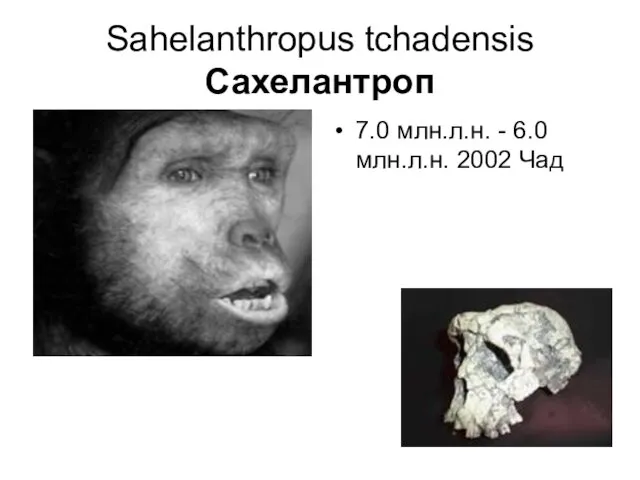
Sahelanthropus tchadensis
Сахелантроп
7.0 млн.л.н. - 6.0 млн.л.н. 2002 Чад
Sahelanthropus tchadensis
Сахелантроп
7.0 млн.л.н. - 6.0 млн.л.н. 2002 Чад

Останки найдены в Республике Чад в Торос-Меналла. Шесть находок гоминид включили
Останки найдены в Республике Чад в Торос-Меналла. Шесть находок гоминид включили

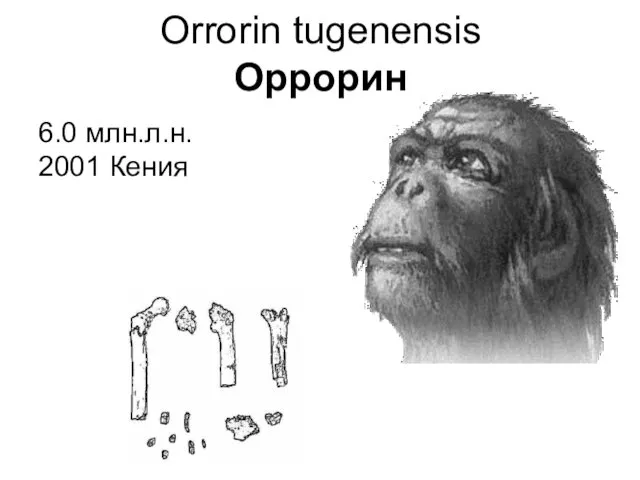
Orrorin tugenensis
Оррорин
6.0 млн.л.н.
2001 Кения
Orrorin tugenensis
Оррорин
6.0 млн.л.н.
2001 Кения


В 2001 г. Б. Сеню и М. Пикфорд обнаружила в Туген
В 2001 г. Б. Сеню и М. Пикфорд обнаружила в Туген

Ardipithecus kadabba
Ардипитек кадабба
5.5 млн.л.н.
2001 Эфиопия
Ardipithecus kadabba
Ардипитек кадабба
5.5 млн.л.н.
2001 Эфиопия

Ардипитек кадабба (Ardipithecus ramidus kadabba) представляет ещё один род поздномиоценовых гоминид,
Ардипитек кадабба (Ardipithecus ramidus kadabba) представляет ещё один род поздномиоценовых гоминид,

Ardipithecus kadabba одно из древнейших прямоходящих существ, от которого сохранились кости ног
Ardipithecus kadabba одно из древнейших прямоходящих существ, от которого сохранились кости ног

Ardipithecus ramidus
Ардипитек рамидус
4 млн.л.н.
1995 Эфиопия
Ardipithecus ramidus
Ардипитек рамидус
4 млн.л.н.
1995 Эфиопия

Ардипитеки (лат. Ardipithecus) — древний род гоминид, который жил в раннем плиоцене. Поскольку род ардипитеки имеет
Ардипитеки (лат. Ardipithecus) — древний род гоминид, который жил в раннем плиоцене. Поскольку род ардипитеки имеет

Найденные в Арамисе в 1995 гг. остатки гоминид включили фрагменты черепов, нижней челюсти
Найденные в Арамисе в 1995 гг. остатки гоминид включили фрагменты черепов, нижней челюсти

Australopithecus anamensis
Австралопитек анамский
4.2 млн.л.н. - 3.9 млн.л.н.
1994 Кения
Australopithecus anamensis
Австралопитек анамский
4.2 млн.л.н. - 3.9 млн.л.н.
1994 Кения

Австралопитек анамский (лат. Australopithecus anamensis) — ископаемый вид рода австралопитеков, обитавший в Восточной Африке около
Австралопитек анамский (лат. Australopithecus anamensis) — ископаемый вид рода австралопитеков, обитавший в Восточной Африке около

Australopithecus afarensis
Австралопитек афарский
4.0 млн.л.н. - 2.5 млн.л.н.
1978 Кения, Эфиопия, Танзания
Australopithecus afarensis
Австралопитек афарский
4.0 млн.л.н. - 2.5 млн.л.н.
1978 Кения, Эфиопия, Танзания

Австралопитек афарский (лат. Australopithecus afarensis) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид, живший
Австралопитек афарский (лат. Australopithecus afarensis) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид, живший

Морфологические особенности хадарских австралопитеков, благодаря представительности материала, изучены очень хорошо и
Морфологические особенности хадарских австралопитеков, благодаря представительности материала, изучены очень хорошо и

Из особенностей строения зубов замечательны значительные размеры резцов и большая разница
Из особенностей строения зубов замечательны значительные размеры резцов и большая разница

Морфология стопы A. afarensis также свидетельствует о разнообразии видов локомоции, использовавшихся этим видом;
Морфология стопы A. afarensis также свидетельствует о разнообразии видов локомоции, использовавшихся этим видом;

Kenyanthropus platyops
3.5 млн.л.н. - 3.2 млн.л.н.
2001 Кения
Kenyanthropus platyops
3.5 млн.л.н. - 3.2 млн.л.н.
2001 Кения

Кениантропы (лат. Kenyanthropus) — древний род гоминид) — древний род гоминид, живший в плиоцене около 3,5-3,2 млн лет назад.
Найден череп
Кениантропы (лат. Kenyanthropus) — древний род гоминид) — древний род гоминид, живший в плиоцене около 3,5-3,2 млн лет назад.
Найден череп

Australopithecus africanu
Австралопитек африканский
3.5 млн.л.н. - 2.4 млн.л.н.
1925 ЮАР
Australopithecus africanu
Австралопитек африканский
3.5 млн.л.н. - 2.4 млн.л.н.
1925 ЮАР

Австралопитек африканский (лат. Australopithecus africanus) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид, живший
Австралопитек африканский (лат. Australopithecus africanus) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид, живший

Пропорции, видимо, были промежуточными между пропорциями шимпанзе и современного человека.
Общие размеры
Пропорции, видимо, были промежуточными между пропорциями шимпанзе и современного человека.
Общие размеры

Череп A. africanus в среднем больше, чем у A. afarensis; наклон лба сравнительно небольшой,
Череп A. africanus в среднем больше, чем у A. afarensis; наклон лба сравнительно небольшой,

Australopithecus bahrelghazali
Австралопитек бахр-эль-газальский
3.5 млн.л.н. - 3.0 млн.л.н
1996 Чад
Australopithecus bahrelghazali
Австралопитек бахр-эль-газальский
3.5 млн.л.н. - 3.0 млн.л.н
1996 Чад

Австралопитек бахр-эль-газальский (лат. Australopithecus bahrelghazali) — предположительно, самостоятельный вид австралопитека) — предположительно, самостоятельный вид австралопитека, семейство гоминид, живший
Австралопитек бахр-эль-газальский (лат. Australopithecus bahrelghazali) — предположительно, самостоятельный вид австралопитека) — предположительно, самостоятельный вид австралопитека, семейство гоминид, живший

Всего обнаружены один фрагмент верхней и один – нижней челюстей, а
Всего обнаружены один фрагмент верхней и один – нижней челюстей, а

Australopithecus garhi
Австралопитек гари
2.5 млн.л.н.
1999 Эфиопия
Australopithecus garhi
Австралопитек гари
2.5 млн.л.н.
1999 Эфиопия

Австралопитек гари (лат. Australopithecus garhi) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид) — вымерший вид австралопитека, семейство гоминид,
Австралопитек гари (лат. Australopithecus garhi) — вымерший вид австралопитека) — вымерший вид австралопитека, семейство гоминид) — вымерший вид австралопитека, семейство гоминид,

«Garhi» в переводе с афарского означает «удивительный» или «сюрприз».
Это поздний грацильный
«Garhi» в переводе с афарского означает «удивительный» или «сюрприз».
Это поздний грацильный

Australopithecus sediba
Австралопитек седиба
2.0 млн.л.н. - 1.8 млн.л.н.
2010 ЮАР
Australopithecus sediba
Австралопитек седиба
2.0 млн.л.н. - 1.8 млн.л.н.
2010 ЮАР

Австралопитек седиба (Australopithecus sediba) — вид австралопитеков) — вид австралопитеков, известный по останкам, возраст
Австралопитек седиба (Australopithecus sediba) — вид австралопитеков) — вид австралопитеков, известный по останкам, возраст

Интересно, что находка останков Australopithecus sediba сделана с использованием популярного сервиса Google Earth, позволившего
Интересно, что находка останков Australopithecus sediba сделана с использованием популярного сервиса Google Earth, позволившего
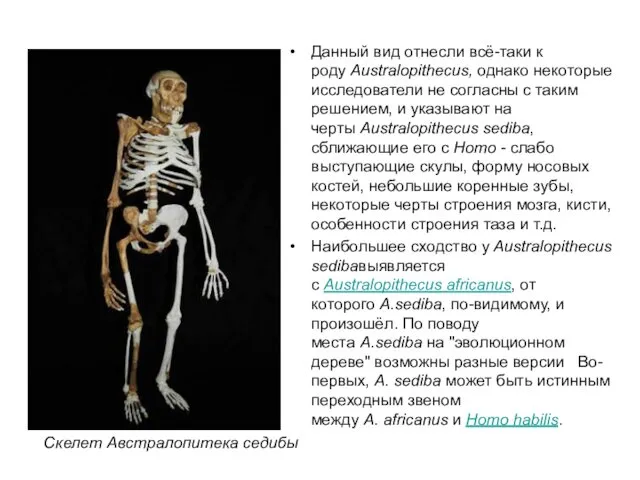
Данный вид отнесли всё-таки к роду Australopithecus, однако некоторые исследователи не согласны с
Данный вид отнесли всё-таки к роду Australopithecus, однако некоторые исследователи не согласны с

Однако, древнейшие "ранние Homo" и в Южной, и в Восточной Африке имеют
Однако, древнейшие "ранние Homo" и в Южной, и в Восточной Африке имеют

Paranthropus aethiopicus
Парантроп эфиопский
2.5 млн.л.н.
1968 Кения, Эфиопия
Paranthropus aethiopicus
Парантроп эфиопский
2.5 млн.л.н.
1968 Кения, Эфиопия

Парантроп эфиопский (лат. Paranthropus aethiopicus) — ископаемый вид рода парантропов) — ископаемый вид рода парантропов («массивных
Парантроп эфиопский (лат. Paranthropus aethiopicus) — ископаемый вид рода парантропов) — ископаемый вид рода парантропов («массивных

(Если быть точным, история данного вида довольно запутанная. Первоначально род и
(Если быть точным, история данного вида довольно запутанная. Первоначально род и

Paranthropus boisei
Парантроп Бойса
2.5 млн.л.н. - 1.0 млн.л.н.
1959 Кения, Танзания
Paranthropus boisei
Парантроп Бойса
2.5 млн.л.н. - 1.0 млн.л.н.
1959 Кения, Танзания

Парантроп Бойса (Paranthropus boisei Mary Leakey, 1959), прежде также зинджантроп (Zinjanthropus boisei), — самый массивный вид парантропов), — самый
Парантроп Бойса (Paranthropus boisei Mary Leakey, 1959), прежде также зинджантроп (Zinjanthropus boisei), — самый массивный вид парантропов), — самый

Череп P. boisei хорошо известен по множеству находок хорошей сохранности, с датировками в
Череп P. boisei хорошо известен по множеству находок хорошей сохранности, с датировками в

Строение мозга P. boisei довольно примитивное.Эндокраны характеризуются сильно суженной лобной долей даже по сравнению
Строение мозга P. boisei довольно примитивное.Эндокраны характеризуются сильно суженной лобной долей даже по сравнению

Относительные размеры передних и заклыковых зубов P. boisei резко отличаются от пропорций, характерных
Относительные размеры передних и заклыковых зубов P. boisei резко отличаются от пропорций, характерных

Paranthropus robustus
Парантроп массивный
2.5 млн.л.н. - 900 тыс.л.н.
1939 ЮАР
Paranthropus robustus
Парантроп массивный
2.5 млн.л.н. - 900 тыс.л.н.
1939 ЮАР

В 1938 г. Роберт Брум проводил раскопки в пещерах Стеркфонтейна. 8
В 1938 г. Роберт Брум проводил раскопки в пещерах Стеркфонтейна. 8

Эндокран P. robustus имеет примитивную форму, мало отличающуюся от формы человекообразных обезьян.
Верхние конечности P.
Эндокран P. robustus имеет примитивную форму, мало отличающуюся от формы человекообразных обезьян.
Верхние конечности P.

Homo habilis
Человек умелый
2.3 млн.л.н. - 1.5 млн.л.н.
1964 Кения, ЮАР
Homo habilis
Человек умелый
2.3 млн.л.н. - 1.5 млн.л.н.
1964 Кения, ЮАР

Челове́к уме́лый (лат. Homo habilis) — высокоразвитый австралопитек или первый представитель рода Homo.
Обнаружен археологами Лики (МэриОбнаружен
Челове́к уме́лый (лат. Homo habilis) — высокоразвитый австралопитек или первый представитель рода Homo.
Обнаружен археологами Лики (МэриОбнаружен

Человек умелый был первым из наших предков, который стал регулярно изготавливать
Человек умелый был первым из наших предков, который стал регулярно изготавливать

Лицевой скелет H. habilis более прогрессивный, чем у грацильных австралопитеков. Горизонтальная уплощенность лица
Лицевой скелет H. habilis более прогрессивный, чем у грацильных австралопитеков. Горизонтальная уплощенность лица
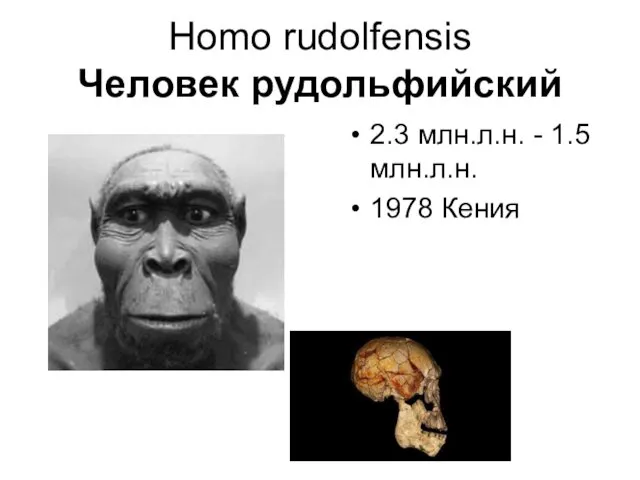
Homo rudolfensis
Человек рудольфийский
2.3 млн.л.н. - 1.5 млн.л.н.
1978 Кения
Homo rudolfensis
Человек рудольфийский
2.3 млн.л.н. - 1.5 млн.л.н.
1978 Кения

Человек рудольфский (лат. Homo rudolfensis) — вымерший «старейший» вид людей, представляющий (согласно традиционным представлениям) переходную
Человек рудольфский (лат. Homo rudolfensis) — вымерший «старейший» вид людей, представляющий (согласно традиционным представлениям) переходную

Наиболее характерным отличием от Homo habilis является большая массивность и большие размеры зубов
Наиболее характерным отличием от Homo habilis является большая массивность и большие размеры зубов

Homo ergaster
Человек работающий
1.8 млн.л.н. - 1.4 млн.л.н.
1975 Кения, Танзания, Грузия
Homo ergaster
Человек работающий
1.8 млн.л.н. - 1.4 млн.л.н.
1975 Кения, Танзания, Грузия

Homo ergaster (Человек работающий) — ископаемый вид людей, появившийся в Африке) — ископаемый вид людей,
Homo ergaster (Человек работающий) — ископаемый вид людей, появившийся в Африке) — ископаемый вид людей,

Обычно под термином Homo ergaster понимаются ранние сравнительно массивные восточноафриканские (точнее – из
Обычно под термином Homo ergaster понимаются ранние сравнительно массивные восточноафриканские (точнее – из

Черепа H.ergaster низкие и массивные, кости свода толстые. Лобная кость убегающая
Черепа H.ergaster низкие и массивные, кости свода толстые. Лобная кость убегающая

Homo erectus
Человек прямоходящий
1.5 млн.л.н. - 400 тыс.л.н.
1894 Ява, Китай, Греция, Испания,
Homo erectus
Человек прямоходящий
1.5 млн.л.н. - 400 тыс.л.н.
1894 Ява, Китай, Греция, Испания,

Эректус (лат. Homo erectus — человек прямоходящий; устар. архантропы) — ископаемый вид людей, который рассматривают как непосредственного предшественника современных
Эректус (лат. Homo erectus — человек прямоходящий; устар. архантропы) — ископаемый вид людей, который рассматривают как непосредственного предшественника современных

Наиболее массивные представители древнейших людей. Являясь уже несомненными людьми, Homo erectus всё
Наиболее массивные представители древнейших людей. Являясь уже несомненными людьми, Homo erectus всё

Homo heidelbergensis
Человек гейдельбергский
800 тыс.л.н. - 130 тыс.л.н.
1908 Кения, Эфиопия, Германия, и
Homo heidelbergensis
Человек гейдельбергский
800 тыс.л.н. - 130 тыс.л.н.
1908 Кения, Эфиопия, Германия, и

Гейдельбергский человек (лат. Homo heidelbergensis) — ископаемый вид людей, европейская разновидность человека прямоходящего (родственный восточноазиатскому синантропу (родственный восточноазиатскому синантропу и
Гейдельбергский человек (лат. Homo heidelbergensis) — ископаемый вид людей, европейская разновидность человека прямоходящего (родственный восточноазиатскому синантропу (родственный восточноазиатскому синантропу и

Относительно поздняя форма, близкая Homo erectus. От архантропов отличались некоторыми прогрессивными (и
Относительно поздняя форма, близкая Homo erectus. От архантропов отличались некоторыми прогрессивными (и

Homo helmei
500 тыс.л.н. – 130 тыс.л.н.
1935 Африка
Homo helmei
500 тыс.л.н. – 130 тыс.л.н.
1935 Африка

Африканские формы палеоантропов, не имеющие специализированных признаков европейских неандертальцев. Часто их относят
Африканские формы палеоантропов, не имеющие специализированных признаков европейских неандертальцев. Часто их относят

Homo neanderthalensis
Неандерталец
130 тыс.л.н. - 28 тыс.л.н.
1864 Африка, Европа, Ближний Восток
Homo neanderthalensis
Неандерталец
130 тыс.л.н. - 28 тыс.л.н.
1864 Африка, Европа, Ближний Восток

Неандерта́лец, человек неандертальский (лат. Homo neanderthalensis или Homo sapiens neanderthalensis; в советской литературе также именовался палеоантропом) — ископаемый
Неандерта́лец, человек неандертальский (лат. Homo neanderthalensis или Homo sapiens neanderthalensis; в советской литературе также именовался палеоантропом) — ископаемый

Специализированный к хищничеству вариант человека. Обладали множеством вполне человеческих черт строения
Специализированный к хищничеству вариант человека. Обладали множеством вполне человеческих черт строения

Homo floresiensis
Человек флоресский
95 тыс.л.н. - 12 тыс.л.н.
2004 Флорес
Homo floresiensis
Человек флоресский
95 тыс.л.н. - 12 тыс.л.н.
2004 Флорес

Человек флоресский (Homo floresiensis BROWN ET AL., 2004) — возможно, карликовый ископаемый вид людей.
Человек флоресский (Homo floresiensis BROWN ET AL., 2004) — возможно, карликовый ископаемый вид людей.

Наилучшим образом сохранился почти полный скелет Лянг Буа 1 (LB1), найденный
Наилучшим образом сохранился почти полный скелет Лянг Буа 1 (LB1), найденный
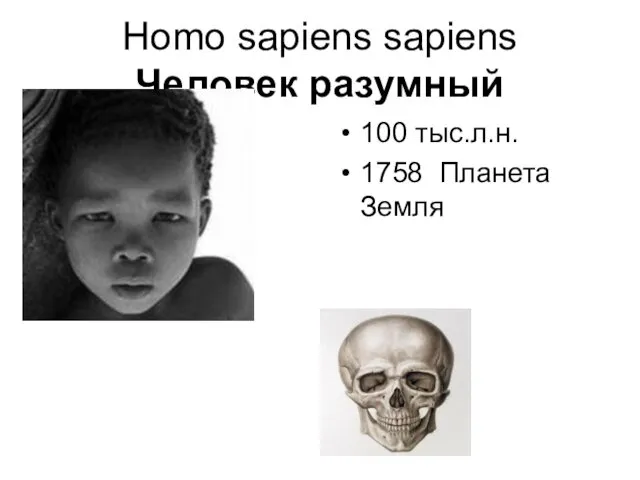
Homo sapiens sapiens
Человек разумный
100 тыс.л.н.
1758 Планета Земля
Homo sapiens sapiens
Человек разумный
100 тыс.л.н.
1758 Планета Земля

Челове́к разу́мный (лат. Homo sapiens; в русскоязычных текстах встречается также написание Хомо Сапиенс илиГомо Сапиенс — вид — вид рода — вид рода Люди (Homo) из
Челове́к разу́мный (лат. Homo sapiens; в русскоязычных текстах встречается также написание Хомо Сапиенс илиГомо Сапиенс — вид — вид рода — вид рода Люди (Homo) из

Эволюционное дерево человека
Следующая схема была подготовлена нашим Научным редактором Станиславом ДробышевскимСледующая схема
Эволюционное дерево человека
Следующая схема была подготовлена нашим Научным редактором Станиславом ДробышевскимСледующая схема
 Место человека в системе органического мира
Место человека в системе органического мира Значение бактерий в природе и жизни человека
Значение бактерий в природе и жизни человека Таємниці мітозу
Таємниці мітозу Закономерности изменчивости. Наследственная и ненаследственная изменчивость. Норма реакции
Закономерности изменчивости. Наследственная и ненаследственная изменчивость. Норма реакции Почему нужно есть много овощей и фруктов
Почему нужно есть много овощей и фруктов Итальянский стиль садов
Итальянский стиль садов Строение организмов. Животные: морфология и анатомия. Хордовые
Строение организмов. Животные: морфология и анатомия. Хордовые Грибы
Грибы Видільна система тварин
Видільна система тварин Пищеварение. Обмен веществ у человека. (ОГЭ. Тест 4)
Пищеварение. Обмен веществ у человека. (ОГЭ. Тест 4) Грибы - гетеротрофы
Грибы - гетеротрофы Вегетативтік жүйке жүйесі
Вегетативтік жүйке жүйесі Основные элементы биотехнологических процессов
Основные элементы биотехнологических процессов Витамины в жизни человека. Понятия авитаминоз, гиповитаминоз
Витамины в жизни человека. Понятия авитаминоз, гиповитаминоз Класс пресмыкающиеся
Класс пресмыкающиеся Тип Моллюски
Тип Моллюски Строение нефрона. Функции почек
Строение нефрона. Функции почек Товароведная характеристика бананов
Товароведная характеристика бананов История развития генетики как науки
История развития генетики как науки Основные направления эволюции
Основные направления эволюции презентация к уроку в 7 классе Отдел Голосеменные
презентация к уроку в 7 классе Отдел Голосеменные Ретинол
Ретинол Проверочная работа Зрительный анализатор 8 класс
Проверочная работа Зрительный анализатор 8 класс Кондиции сельскохозяйственных животных
Кондиции сельскохозяйственных животных Пример составления рабочей программы по фгос
Пример составления рабочей программы по фгос Проектирование агротехнологий. Понятие об агротехнологиях, принципы их формирования и освоения
Проектирование агротехнологий. Понятие об агротехнологиях, принципы их формирования и освоения Закономерность для биологических процессов роста и развития растений. Вид биологических кривых
Закономерность для биологических процессов роста и развития растений. Вид биологических кривых Презентация по теме: Высшая нервная деятельность
Презентация по теме: Высшая нервная деятельность