- Белки

Содержание
- 2. Белки (протеины) – важнейший класс БАВ Жизнь есть способ существования белковых тел Ф. Энгельс Белки –
- 3. Особенности белков Разнообразие структур и высокая видовая специфичность Способность к различным внутримолекулярным взаимодействиям ⇒ динамичность структур
- 4. Молекулярная масса белков. Форма белковых молекул (самостоятельно)
- 5. Аминокислотный состав белков (см. лабораторные и семинарские занятия)
- 6. Структурные элементы в белках – α-аминокислоты, отличающиеся друг от друга строением боковых групп (боковых цепей, радикалов)
- 7. Две категории аминокислот Постоянно встречающиеся – 18 аминокислот, 2 амида (аспарагин и глутамин) Иногда встречающиеся Орнитин
- 8. Классификации α-аминокислот I. По химическому составу Алифатические Моноаминомонокарбоновые (нейтральные) – гли, ала, сер, цис, лей, иле,
- 9. Свойства аминокислот Важная особенность – оптическая активность (кроме гли) 7 аминокислот характеризуются правым (+) 10 –
- 10. Обозначения оптических изомеров (+), (–) – обозначение направления вращения плоскости поляризации (+) – правовращающий изомер (–)
- 11. Обозначения оптических изомеров D, L – обозначают «фамильные» признаки (выражают конфигурацию, относительную к опорному соединению) D
- 12. Обозначения оптических изомеров R, S – выражают абсолютную конфигурацию R – атомные номера заместителей (кроме заместителя
- 13. Радикалы аминокислот (длина, объем, взаимное расположение, химические свойства) определяют объем, форму, рельеф поверхности белковой частицы, степень
- 14. Общее число аминокислотных остатков в молекулах белков изменяется в широких пределах Мr,ср.(аминокислотного остатка) = 115 Коэффициент
- 15. Пептиды
- 16. Образование пептидов Аланилфенилаланин (дипептид) Аланин Фенилаланин
- 17. Образование пептидных связей в воде термодинамически невыгодно ⇒ необходимость предварительной активации взаимодействующих групп при химическом синтезе
- 18. Природные пептиды
- 19. Глутатион Трипептид Ф. Хопкинс (1930), из дрожжей γ-глутамилцистеинилглицин легко окисляется: 2R–SН ⇄ R–S–S–R + 2Н восстановленная
- 20. Пептидные антибиотики Многие циклопептиды Грамицидин Грамицидин S – циклодекапептид, молекула которого состоит из 2-х симметричных пентапептидов,
- 21. Грамицидин S
- 22. Циклоспорин
- 23. Нейропептиды Обнаружены в мозге и способны влиять на функции цнс энкефалины и эндорфины (опиоидные пептиды) способны
- 24. Пептидные гормоны Окситоцин (В. дю Виньо, 1953) Нонапептид, 1-я и 6-я АК – цис HS-группы связаны
- 25. Пептидные токсины токсины бледной поганки – фаллоидин и α-аманитин – циклопептиды необычной структуры – бициклические системы
- 26. Пептиды с вкусовыми качествами Аспартам – метиловый эфир L-аспартил-L-фенилаланина – в 200 раз слаще сахарозы Гептапептид
- 27. Структура белковой молекулы
- 28. Полипептидная теория строения белка 1902 г., Э. Фишер Белки – сложные полипептиды, в которых отдельные аминокислоты
- 29. Структурные особенности пептидной цепи
- 30. Структурные особенности пептидной цепи Пептидная связь примерно на 10% короче связи –С–N– и имеет характер «частично
- 31. Структурные особенности пептидной цепи Транс-конфигурация Выгоднее цис-конфигурации на 2,6 ккал/моль (10,878 кДж/моль), т. к. сближение α-углеродных
- 32. Уровни организации белковой молекулы
- 33. К. У. Линдерстрем-Ланг 4 уровня организации белковых молекул Первичная Вторичная Третичная Четвертичная с т р у
- 34. Первичная структура белка
- 35. Первичная структура белка последовательность аминокислотных остатков в полипептидной цепи кодируется структурным геном данного белка содержит все
- 36. определение вторичной и третичной структур выяснение расположения функциональных групп в активном центре, механизма его функционирования выяснение
- 37. Серповидноклеточная анемия
- 38. определение вторичной и третичной структур выяснение расположения функциональных групп в активном центре, механизма его функционирования выяснение
- 39. Невалентные взаимодействия в пептидной цепи
- 40. Водородные связи В неполярном окружении энергия водородной связи –СО⋅⋅⋅⋅НN– составляет около 16,7 кДж/моль, а повышение полярности
- 41. Гидрофобные взаимодействия Энтропийная природа Неполярные заместители «выталкиваются» из воды и стремятся ограничить свой контакт с ней
- 43. Ван-дер-ваальсовы взаимодействия Дисперсионные силы притяжения атомов и Силы взаимного отталкивания их электронных оболочек Энергетический вклад каждого
- 44. Ионные (электростатические) взаимодействия Взаимодействия ионогенных групп, образующих солевые связи Энергия солевых связей в гидрофобном окружении может
- 45. Торсионные взаимодействия Характеризуют «скрученность» одинарной связи Относительно слабы, но при анализе поворотов вокруг связей С–С, С–N
- 46. Вторичная структура белков
- 47. Вторичная структура белка… пространственное расположение отдельных участков полипептидной цепи без учета типа и конформации боковых радикалов
- 48. Стабилизация вторичной структуры за счет водородной связи Влияние окружающей белок воды Влияние воды снижается при формировании
- 49. α-Спираль 50-е годы ХХ в. Л. Полинг и Р. Кори Лайнус Карл Полинг (1901 – 1994)
- 50. α-Спираль Радикалы аминокислотных остатков – на периферии образованного спиралью цилиндра и могут обеспечивать гидрофобную или гидрофильную
- 51. Геометрические параметры α-спирали r = 2,3 Å (0,23 нм) высота спирали (смещение) на 1 остаток d
- 52. α-Спираль Длина α-спиральных участков в глобулярных белках относительно невелика (5–15 аминокислотных остатков, 3–4 витка спирали) в
- 53. β-Структура У. Т. Астбери, 1941 г. 1951 г., Л. Полинг и Р. Кори установили, что β-структура,
- 55. β-Структура Число аминокислотных остатков в отрезке пептидной цепи, образующем β-структуру, обычно 3 – 8 Протяженная β-структура
- 56. β-Изгиб Петли, позволяющие изменить направление пептидной цепи Наиболее экономно – β-изгиб Стабилизируется одной водородной связью Практически
- 57. β-Изгиб
- 58. Вторичная структура белков Зависимость от аминокислотной последовательности
- 59. Способ укладки молекулы белка определяется его аминокислотной последовательностью α-спираль – ала, лей, глу β-структура – мет,
- 60. Сверхвторичная (надвторичная) структура белков
- 61. Сверхвторичная (надвторичная) структура белков Пространственное строение ансамблей взаимодействующих между собой вторичных структур
- 62. Сверхвторичная (надвторичная) структура белков Типы сверхвторичных структур
- 63. Суперспирализованная α-спираль (α-кератин) αα’ ββ’ βxβ βcβ βαβ (греческий орнамент) βββ (β-зигзаг) βββ (греческий ключ)
- 64. Домены
- 65. Домены… (от фр. domaine – владение) – структурно и функционально обособленные области (субобласти) молекулы, соединенные друг
- 66. Домены Классификация структурных доменов и белков по организации пространственной структуры полипептидной цепи
- 67. 1. α-Белки Миоглобин Гемоглобин
- 68. 2. β-Белки построены в основном из антипараллельных β-слоев Порин Рубредоксин
- 69. Лизоцим, рибонуклеаза 3. α+β-Белки
- 70. Триозофосфатизомераза 4. α/β-Белки
- 71. 5. Домены и белки без выраженной вторичной структуры
- 72. Третичная структура белка
- 73. Третичная структура белка… характеризует пространственное расположение упорядоченных и аморфных участков в полипептидной цепи в целом, которое
- 74. Нитевидные, или фибриллярные белки фиброин шелка кератин волос, рогов, копыт коллаген и др.
- 75. Шаровидные, или глобулярные белки
- 76. Третичная структура белка основа функциональности белка, которая требует точной пространственной организации больших ансамблей, построенных из множества
- 77. Четвертичная структура белка
- 78. Четвертичная структура белка… Олигомерные белки Четвертичная структура – размещение в пространстве взаимодействующих между собой субъединиц, образованных
- 79. Ферритин 24 субъединицы 3500 группировок FеО⋅ОН
- 80. Четвертичная структура белка гомомерные белки гетеромерные белки Объединение в одной структуре нескольких взаимосвязанных функций, создание полифункциональной
- 81. Четвертичная структура белка Межсубъединичные контакты – система нековалентных взаимодействий гидрофобные взаимодействия (контактные площадки) водородные связи электростатические
- 82. Четвертичная структура белка Функциональное значение
- 83. Функции четвертичной структуры Объединение нескольких взаимосвязанных функций в единой структуре Архитектурная функция Формирование пространственных образований весьма
- 84. Функции четвертичной структуры Обеспечение множественных взаимодействий белка с протяженными структурами ДНК-связывающие белки – димеры (?) Регуляторная
- 85. Четвертичная структура белка. Гемоглобин
- 86. Каждый индивидуальный белок характеризуется уникальной структурой, обеспечивающей уникальность его функций Поэтому выяснение структуры разнообразных белков может
- 87. Свойства белков (см. лабораторные занятия)
- 88. Классификация белков
- 89. По степени сложности простые белки (протеины) дают при гидролизе только аминокислоты Альбумины Глобулины Проламины Глютелины Гистоны
- 90. По степени сложности сложные белки (протеиды) = протеин + добавочная группа: Хромопротеины (гемоглобин, цитохромы, каталаза, хлорофилл)
- 91. По форме частиц фибриллярные (волокнистые) белки (фиброин шелка, кератин волос, коллаген кожи) глобулярные (корпускулярные) белки
- 92. По растворимости протеиноиды (склеропротеины) – нерастворимы в обычных растворителях – почти все фибриллярные белки альбумины –
- 93. По аминокислотному составу протамины – содержат 80–90% арг, простейшие белки, растворяются в слабых кислотах – белки
- 94. По выполняемым функциям структурные белки – компоненты клеточных мембран, органелл; коллаген соединительной ткани; кератин волос, ногтей;
- 95. По выполняемым функциям защитные белки: антитела (иммуноглобулины); белки системы свертывания крови (фибриноген, тромбин, фибрин, факторы свертывания);
- 96. По выполняемым функциям резервные белки (овальбумины яиц, белки молока – казеин) рецепторные белки: рецептор ацетилхолина; фоторецепторный
- 97. По выполняемым функциям Все белки выполняют энергетическую функцию: при окислении 1 г белка выделяется 17,2 кДж
- 99. Скачать презентацию

Белки (протеины) – важнейший класс БАВ
Жизнь есть способ существования белковых тел
Белки (протеины) – важнейший класс БАВ
Жизнь есть способ существования белковых тел

Особенности белков
Разнообразие структур и высокая видовая специфичность
Способность к различным внутримолекулярным взаимодействиям
Особенности белков
Разнообразие структур и высокая видовая специфичность
Способность к различным внутримолекулярным взаимодействиям

Молекулярная масса белков. Форма белковых молекул
(самостоятельно)
Молекулярная масса белков. Форма белковых молекул
(самостоятельно)

Аминокислотный состав белков (см. лабораторные и семинарские занятия)
Аминокислотный состав белков (см. лабораторные и семинарские занятия)

Структурные элементы в белках – α-аминокислоты, отличающиеся друг от друга строением
Структурные элементы в белках – α-аминокислоты, отличающиеся друг от друга строением
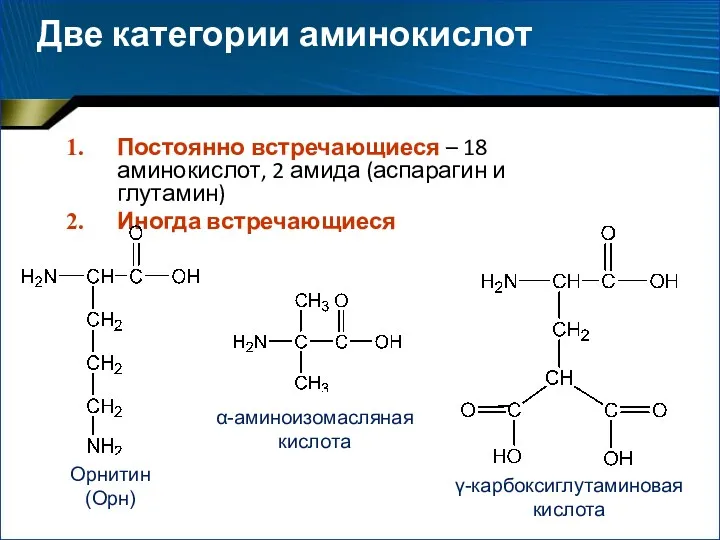
Две категории аминокислот
Постоянно встречающиеся – 18 аминокислот, 2 амида (аспарагин и
Две категории аминокислот
Постоянно встречающиеся – 18 аминокислот, 2 амида (аспарагин и

Классификации α-аминокислот
I. По химическому составу
Алифатические
Моноаминомонокарбоновые (нейтральные) – гли, ала, сер, цис,
Классификации α-аминокислот
I. По химическому составу
Алифатические
Моноаминомонокарбоновые (нейтральные) – гли, ала, сер, цис,

Свойства аминокислот
Важная особенность – оптическая активность (кроме гли)
7 аминокислот характеризуются
Свойства аминокислот
Важная особенность – оптическая активность (кроме гли)
7 аминокислот характеризуются
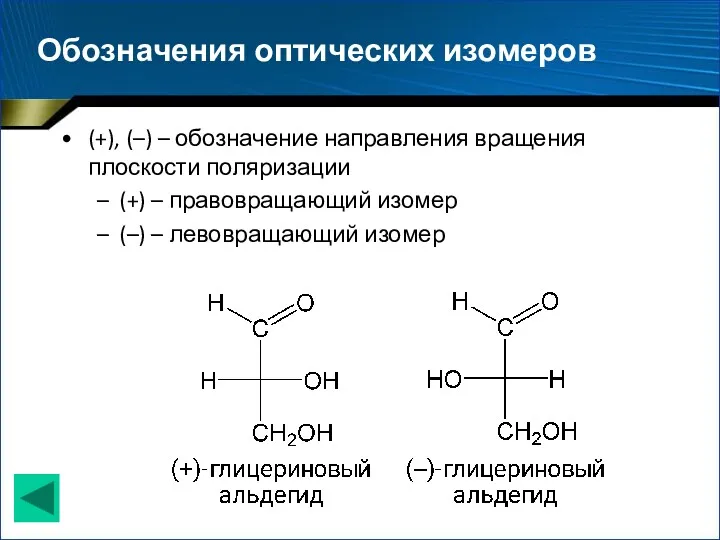
Обозначения оптических изомеров
(+), (–) – обозначение направления вращения плоскости поляризации
(+) –
Обозначения оптических изомеров
(+), (–) – обозначение направления вращения плоскости поляризации
(+) –
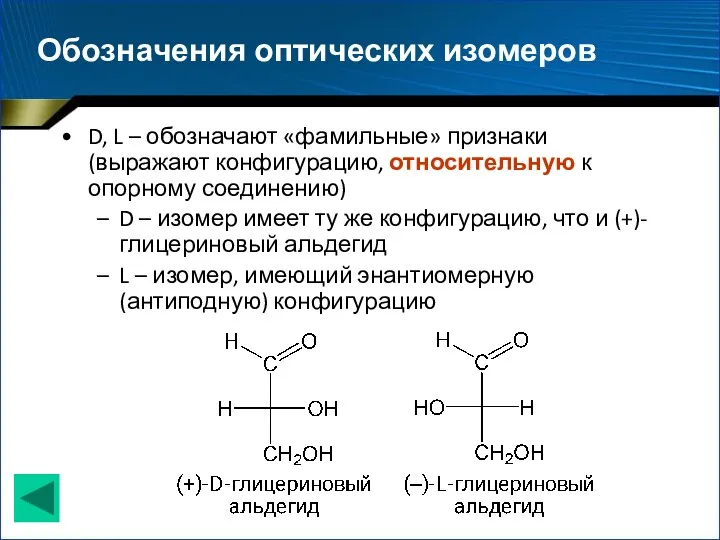
Обозначения оптических изомеров
D, L – обозначают «фамильные» признаки (выражают конфигурацию, относительную
Обозначения оптических изомеров
D, L – обозначают «фамильные» признаки (выражают конфигурацию, относительную
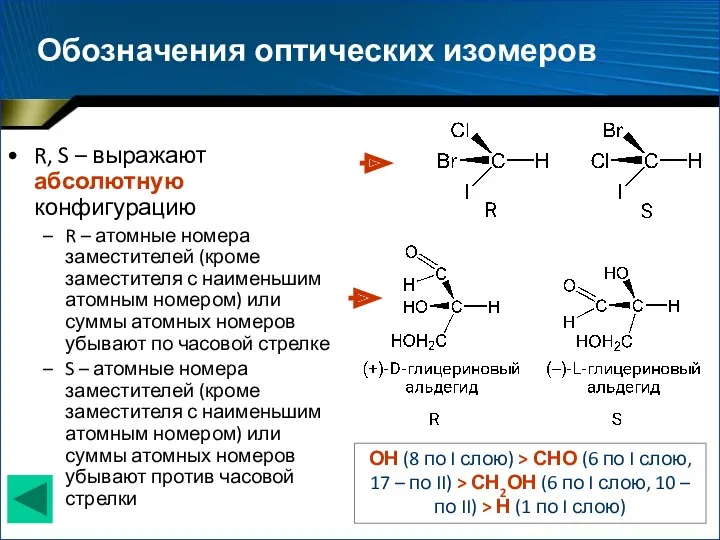
Обозначения оптических изомеров
R, S – выражают абсолютную конфигурацию
R – атомные номера
Обозначения оптических изомеров
R, S – выражают абсолютную конфигурацию
R – атомные номера

Радикалы аминокислот (длина, объем, взаимное расположение, химические свойства) определяют объем, форму,
Радикалы аминокислот (длина, объем, взаимное расположение, химические свойства) определяют объем, форму,

Общее число аминокислотных остатков в молекулах белков изменяется в широких пределах
Мr,ср.(аминокислотного
Общее число аминокислотных остатков в молекулах белков изменяется в широких пределах
Мr,ср.(аминокислотного

Пептиды
Пептиды
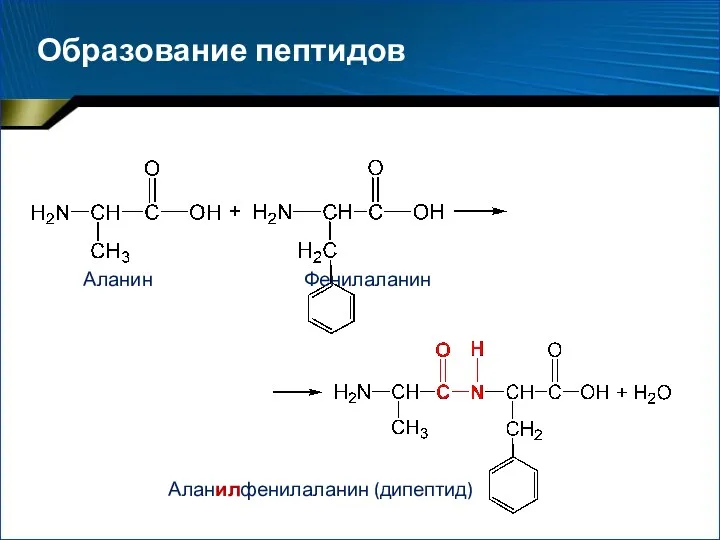
Образование пептидов
Аланилфенилаланин (дипептид)
Аланин Фенилаланин
Образование пептидов
Аланилфенилаланин (дипептид)
Аланин Фенилаланин

Образование пептидных связей в воде термодинамически невыгодно ⇒ необходимость предварительной активации
Образование пептидных связей в воде термодинамически невыгодно ⇒ необходимость предварительной активации

Природные пептиды
Природные пептиды
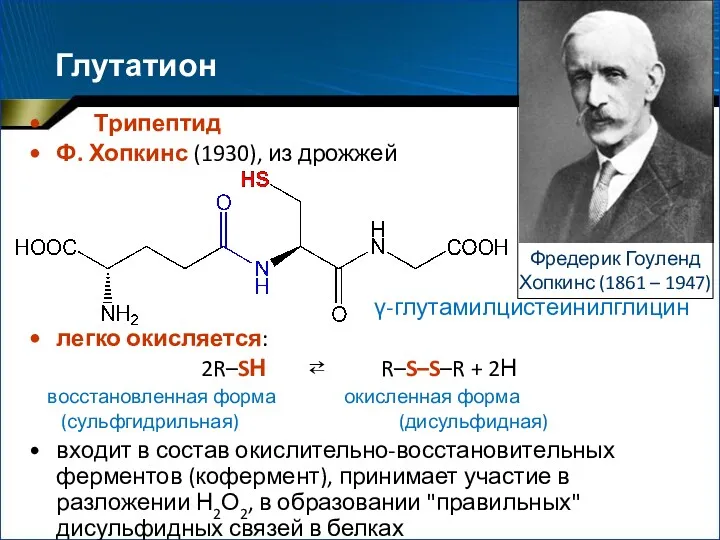
Глутатион
Трипептид
Ф. Хопкинс (1930), из дрожжей
γ-глутамилцистеинилглицин
легко окисляется:
2R–SН ⇄ R–S–S–R +
Глутатион
Трипептид
Ф. Хопкинс (1930), из дрожжей
γ-глутамилцистеинилглицин
легко окисляется:
2R–SН ⇄ R–S–S–R +

Пептидные антибиотики
Многие циклопептиды
Грамицидин Грамицидин S – циклодекапептид, молекула которого состоит
Пептидные антибиотики
Многие циклопептиды
Грамицидин Грамицидин S – циклодекапептид, молекула которого состоит
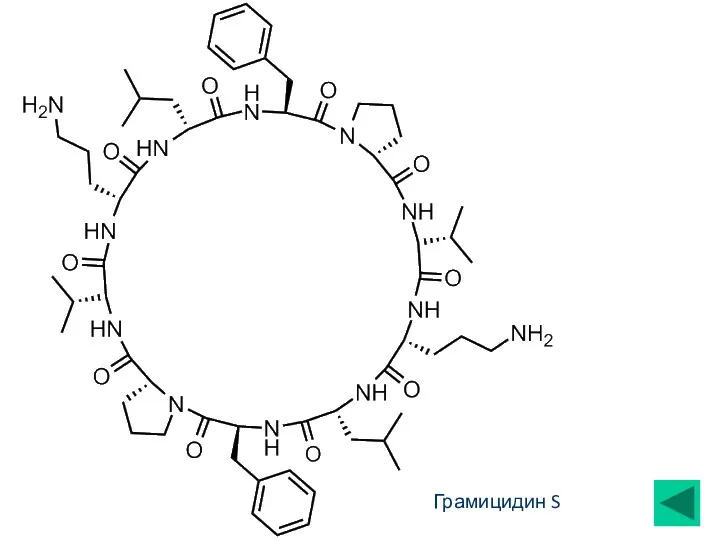
Грамицидин S
Грамицидин S
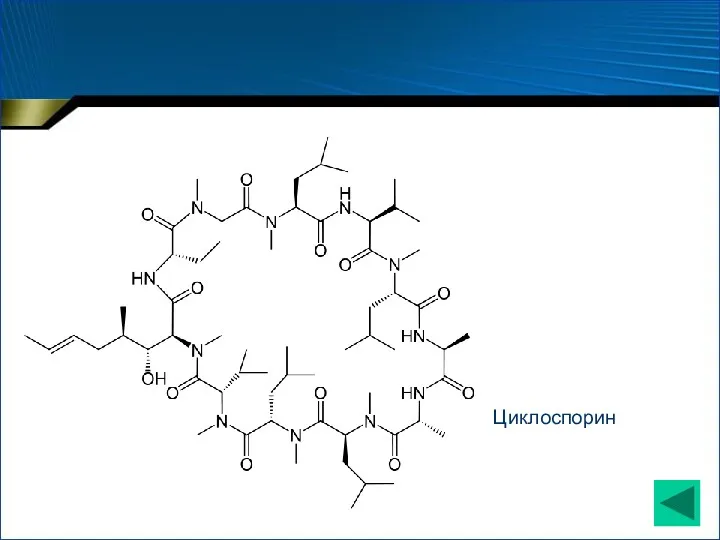
Циклоспорин
Циклоспорин

Нейропептиды
Обнаружены в мозге и способны влиять на функции цнс
энкефалины и
Нейропептиды
Обнаружены в мозге и способны влиять на функции цнс
энкефалины и
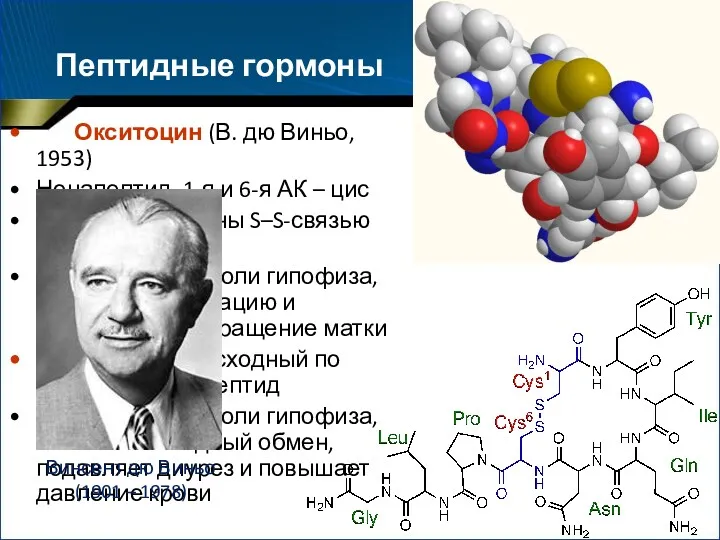
Пептидные гормоны
Окситоцин (В. дю Виньо, 1953)
Нонапептид, 1-я и 6-я
Пептидные гормоны
Окситоцин (В. дю Виньо, 1953)
Нонапептид, 1-я и 6-я
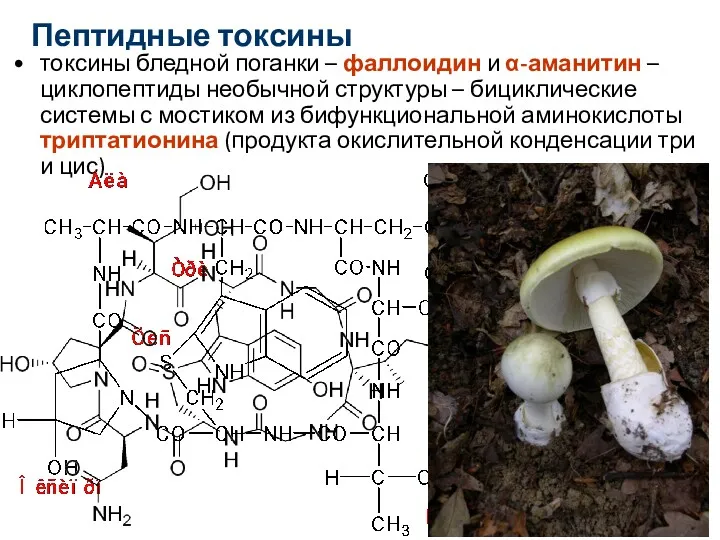
Пептидные токсины
токсины бледной поганки – фаллоидин и α-аманитин – циклопептиды
Пептидные токсины
токсины бледной поганки – фаллоидин и α-аманитин – циклопептиды
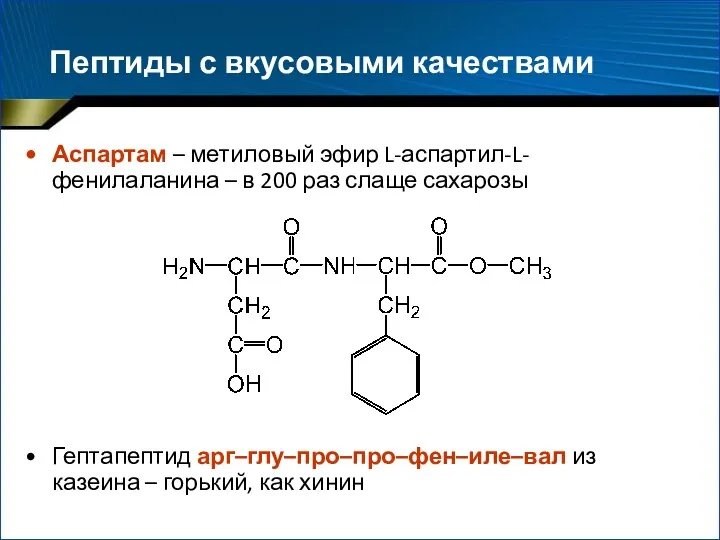
Пептиды с вкусовыми качествами
Аспартам – метиловый эфир L-аспартил-L-фенилаланина – в
Пептиды с вкусовыми качествами
Аспартам – метиловый эфир L-аспартил-L-фенилаланина – в

Структура белковой молекулы
Структура белковой молекулы

Полипептидная теория строения белка
1902 г., Э. Фишер
Белки – сложные полипептиды,
Полипептидная теория строения белка
1902 г., Э. Фишер
Белки – сложные полипептиды,

Структурные особенности пептидной цепи
Структурные особенности пептидной цепи

Структурные особенности пептидной цепи
Пептидная связь примерно на 10% короче связи
Структурные особенности пептидной цепи
Пептидная связь примерно на 10% короче связи
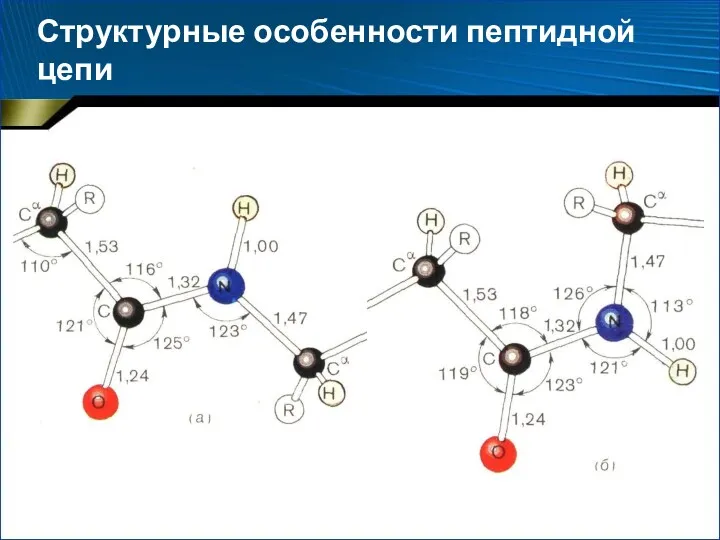
Структурные особенности пептидной цепи
Транс-конфигурация
Выгоднее цис-конфигурации на 2,6 ккал/моль (10,878 кДж/моль),
Структурные особенности пептидной цепи
Транс-конфигурация
Выгоднее цис-конфигурации на 2,6 ккал/моль (10,878 кДж/моль),

Уровни организации белковой молекулы
Уровни организации белковой молекулы

К. У. Линдерстрем-Ланг
4 уровня организации белковых молекул
Первичная
Вторичная
Третичная
Четвертичная
с
т
р
у
к
т
у
р
ы
Кай Ульрик Линдерстрём-Ланг (1896
К. У. Линдерстрем-Ланг
4 уровня организации белковых молекул
Первичная
Вторичная
Третичная
Четвертичная
с
т
р
у
к
т
у
р
ы
Кай Ульрик Линдерстрём-Ланг (1896

Первичная структура белка
Первичная структура белка

Первичная структура белка
последовательность аминокислотных остатков в полипептидной цепи
кодируется структурным геном
Первичная структура белка
последовательность аминокислотных остатков в полипептидной цепи
кодируется структурным геном

определение вторичной и третичной структур
выяснение расположения функциональных групп в активном центре,
определение вторичной и третичной структур
выяснение расположения функциональных групп в активном центре,
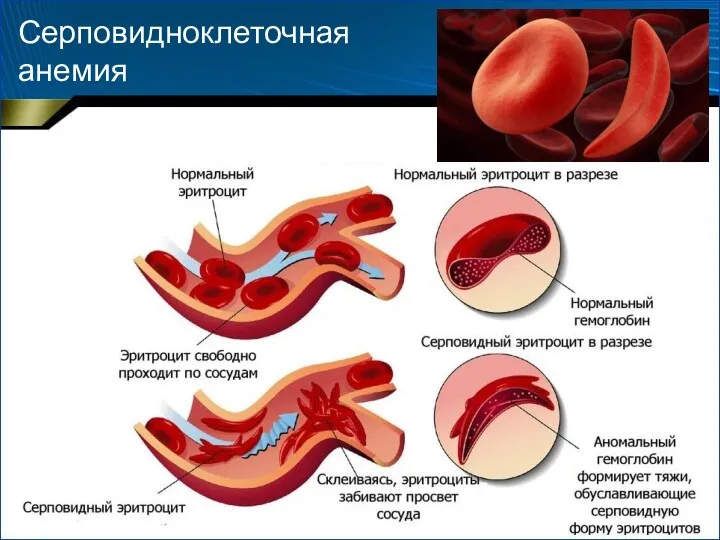
Серповидноклеточная анемия
Серповидноклеточная анемия

определение вторичной и третичной структур
выяснение расположения функциональных групп в активном центре,
определение вторичной и третичной структур
выяснение расположения функциональных групп в активном центре,

Невалентные взаимодействия в пептидной цепи
Невалентные взаимодействия в пептидной цепи
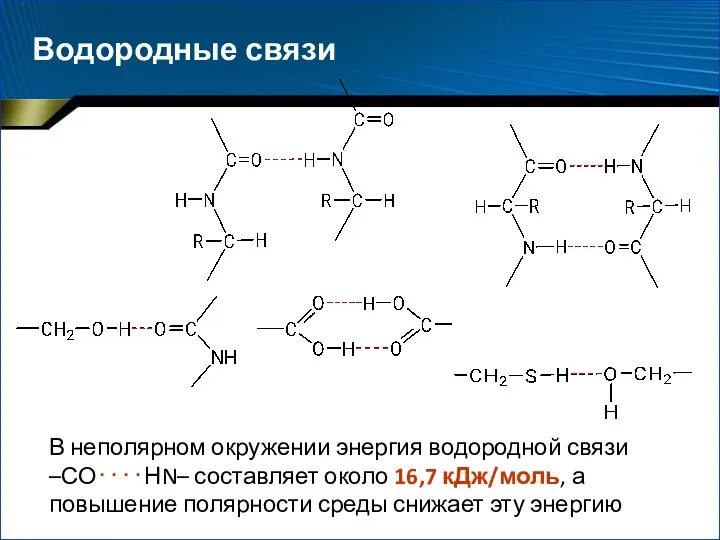
Водородные связи
В неполярном окружении энергия водородной связи –СО⋅⋅⋅⋅НN– составляет около 16,7
Водородные связи
В неполярном окружении энергия водородной связи –СО⋅⋅⋅⋅НN– составляет около 16,7

Гидрофобные взаимодействия
Энтропийная природа
Неполярные заместители «выталкиваются» из воды и стремятся ограничить свой
Гидрофобные взаимодействия
Энтропийная природа
Неполярные заместители «выталкиваются» из воды и стремятся ограничить свой

Ван-дер-ваальсовы взаимодействия
Дисперсионные силы притяжения атомов и
Силы взаимного отталкивания их
Ван-дер-ваальсовы взаимодействия
Дисперсионные силы притяжения атомов и
Силы взаимного отталкивания их
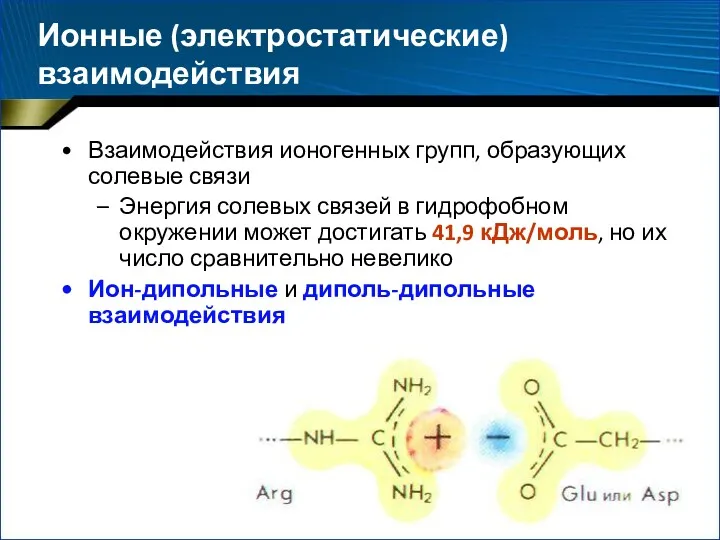
Ионные (электростатические) взаимодействия
Взаимодействия ионогенных групп, образующих солевые связи
Энергия солевых связей
Ионные (электростатические) взаимодействия
Взаимодействия ионогенных групп, образующих солевые связи
Энергия солевых связей
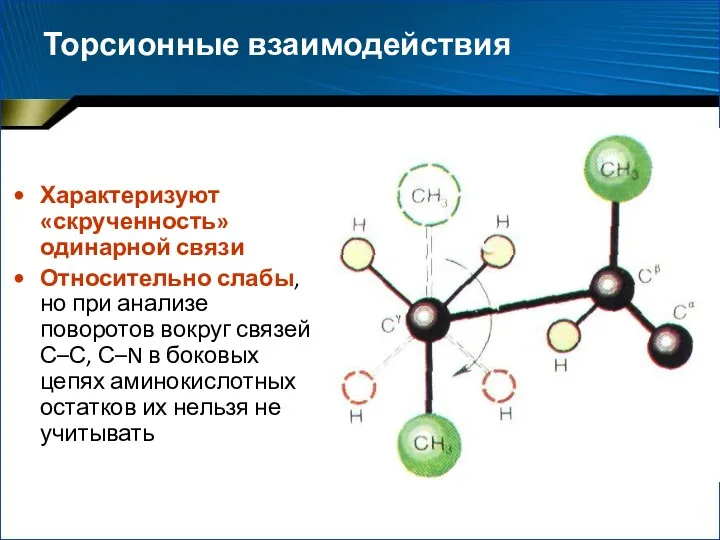
Торсионные взаимодействия
Характеризуют «скрученность» одинарной связи
Относительно слабы, но при анализе поворотов
Торсионные взаимодействия
Характеризуют «скрученность» одинарной связи
Относительно слабы, но при анализе поворотов

Вторичная структура белков
Вторичная структура белков

Вторичная структура белка…
пространственное расположение отдельных участков полипептидной цепи без учета типа
Вторичная структура белка…
пространственное расположение отдельных участков полипептидной цепи без учета типа

Стабилизация вторичной структуры за счет водородной связи
Влияние окружающей белок воды
Влияние воды
Стабилизация вторичной структуры за счет водородной связи
Влияние окружающей белок воды
Влияние воды

α-Спираль
50-е годы ХХ в.
Л. Полинг и Р. Кори
Лайнус Карл Полинг (1901
α-Спираль
50-е годы ХХ в.
Л. Полинг и Р. Кори
Лайнус Карл Полинг (1901

α-Спираль
Радикалы аминокислотных остатков – на периферии образованного спиралью цилиндра и могут
α-Спираль
Радикалы аминокислотных остатков – на периферии образованного спиралью цилиндра и могут
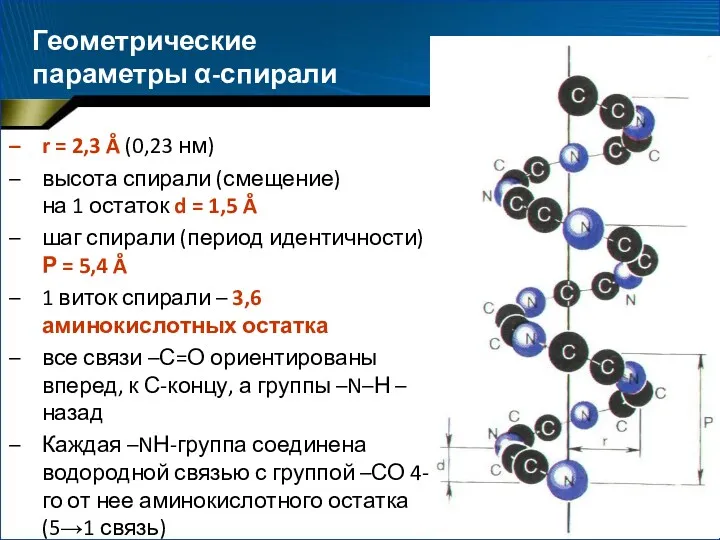
Геометрические параметры α-спирали
r = 2,3 Å (0,23 нм)
высота спирали (смещение) на
Геометрические параметры α-спирали
r = 2,3 Å (0,23 нм)
высота спирали (смещение) на

α-Спираль
Длина α-спиральных участков
в глобулярных белках относительно невелика (5–15 аминокислотных остатков, 3–4
α-Спираль
Длина α-спиральных участков
в глобулярных белках относительно невелика (5–15 аминокислотных остатков, 3–4

β-Структура
У. Т. Астбери, 1941 г.
1951 г., Л. Полинг и Р.
β-Структура
У. Т. Астбери, 1941 г.
1951 г., Л. Полинг и Р.
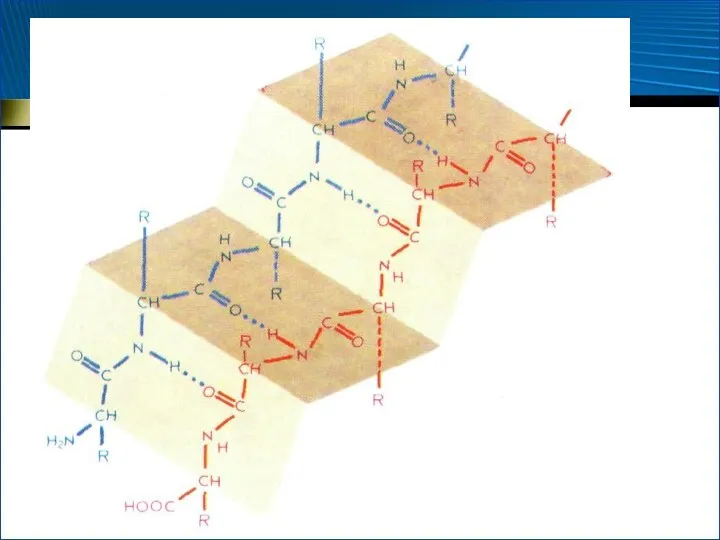

β-Структура
Число аминокислотных остатков в отрезке пептидной цепи, образующем β-структуру, обычно
β-Структура
Число аминокислотных остатков в отрезке пептидной цепи, образующем β-структуру, обычно

β-Изгиб
Петли, позволяющие изменить направление пептидной цепи
Наиболее экономно – β-изгиб
Стабилизируется одной водородной
β-Изгиб
Петли, позволяющие изменить направление пептидной цепи
Наиболее экономно – β-изгиб
Стабилизируется одной водородной
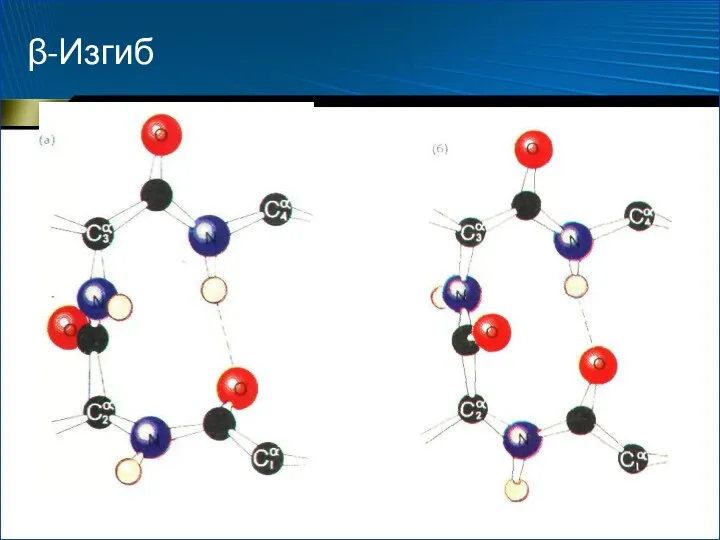
β-Изгиб
β-Изгиб

Вторичная структура белков
Зависимость от аминокислотной последовательности
Вторичная структура белков
Зависимость от аминокислотной последовательности

Способ укладки молекулы белка определяется его аминокислотной последовательностью
α-спираль – ала, лей,
Способ укладки молекулы белка определяется его аминокислотной последовательностью
α-спираль – ала, лей,

Сверхвторичная (надвторичная) структура белков
Сверхвторичная (надвторичная) структура белков

Сверхвторичная (надвторичная) структура белков
Пространственное строение ансамблей взаимодействующих между собой вторичных структур
Сверхвторичная (надвторичная) структура белков
Пространственное строение ансамблей взаимодействующих между собой вторичных структур

Сверхвторичная (надвторичная) структура белков
Типы сверхвторичных структур
Сверхвторичная (надвторичная) структура белков
Типы сверхвторичных структур

Суперспирализованная α-спираль (α-кератин)
αα’
ββ’
βxβ
βcβ
βαβ
(греческий орнамент)
βββ
(β-зигзаг)
βββ
(греческий ключ)
Суперспирализованная α-спираль (α-кератин)
αα’
ββ’
βxβ
βcβ
βαβ
(греческий орнамент)
βββ
(β-зигзаг)
βββ
(греческий ключ)

Домены
Домены
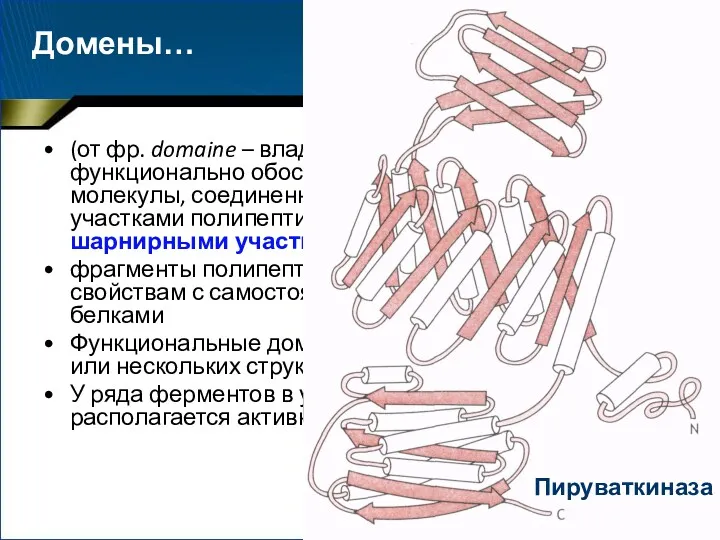
Домены…
(от фр. domaine – владение) – структурно и функционально обособленные области
Домены…
(от фр. domaine – владение) – структурно и функционально обособленные области

Домены
Классификация структурных доменов и белков по организации пространственной структуры полипептидной цепи
Домены
Классификация структурных доменов и белков по организации пространственной структуры полипептидной цепи
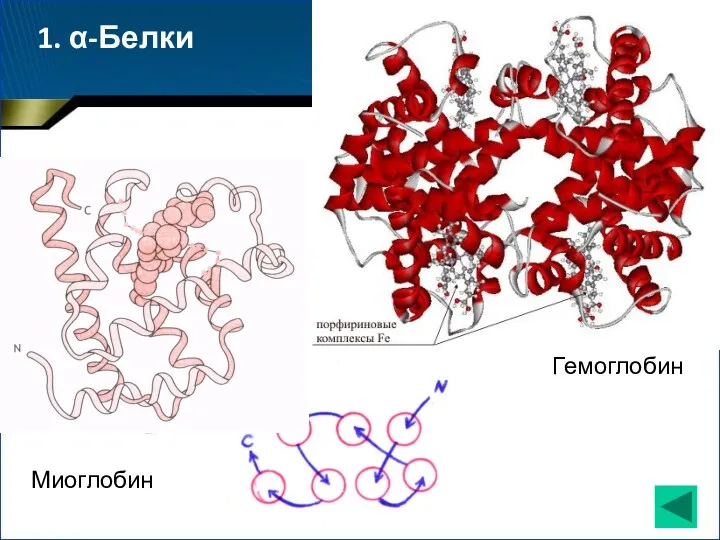
1. α-Белки
Миоглобин
Гемоглобин
1. α-Белки
Миоглобин
Гемоглобин
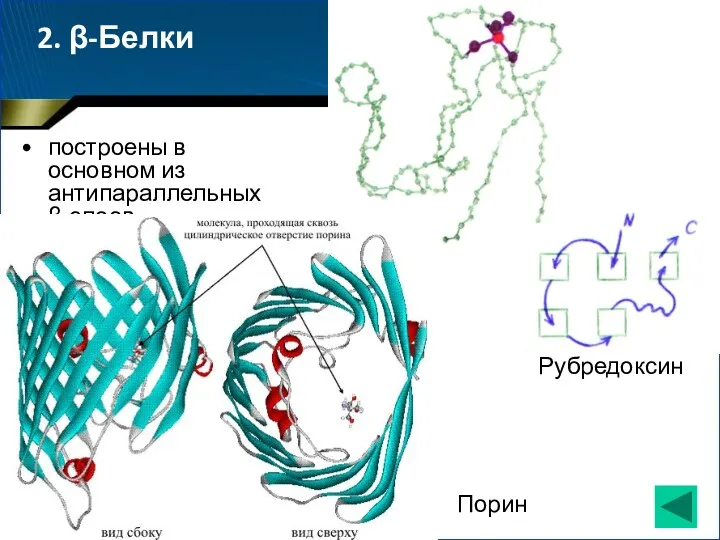
2. β-Белки
построены в основном из антипараллельных β-слоев
Порин
Рубредоксин
2. β-Белки
построены в основном из антипараллельных β-слоев
Порин
Рубредоксин
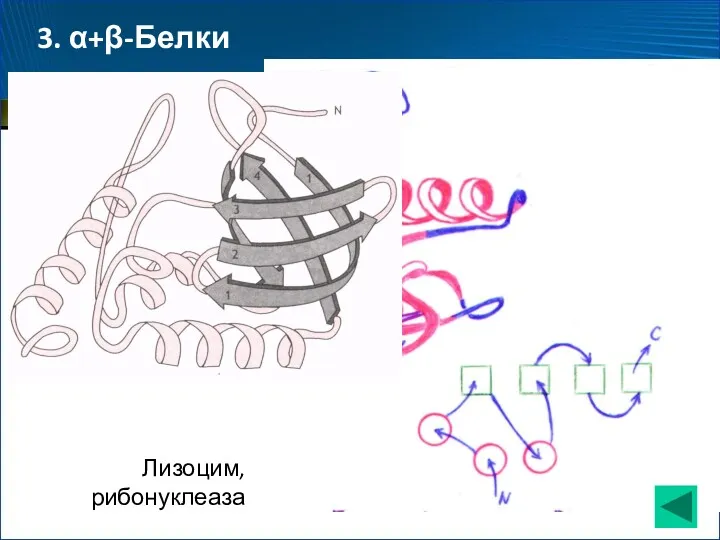
Лизоцим, рибонуклеаза
3. α+β-Белки
Лизоцим, рибонуклеаза
3. α+β-Белки
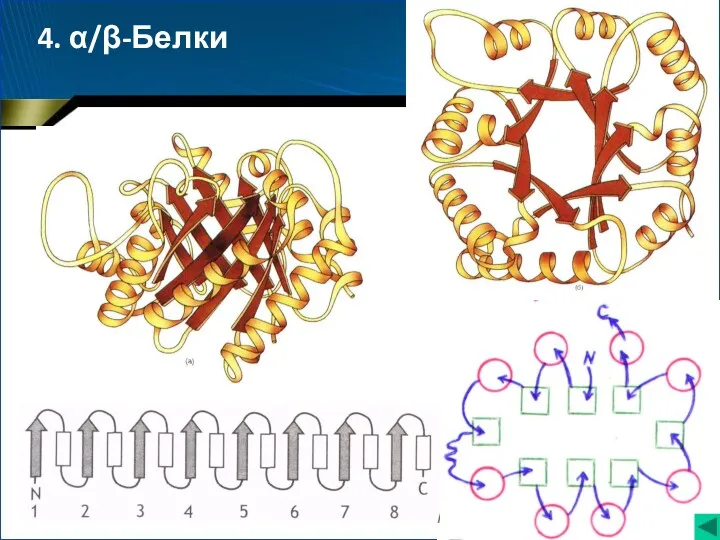
Триозофосфатизомераза
4. α/β-Белки
Триозофосфатизомераза
4. α/β-Белки

5. Домены и белки без выраженной вторичной структуры
5. Домены и белки без выраженной вторичной структуры

Третичная структура белка
Третичная структура белка

Третичная структура белка…
характеризует пространственное расположение упорядоченных и аморфных участков в
Третичная структура белка…
характеризует пространственное расположение упорядоченных и аморфных участков в
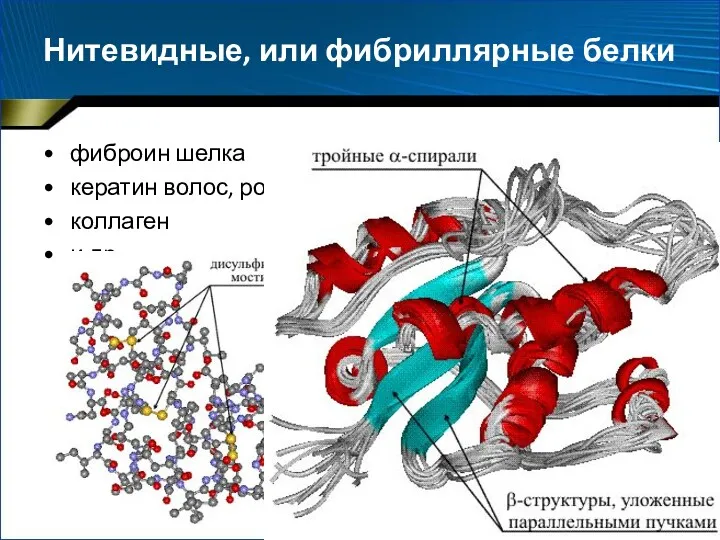
Нитевидные, или фибриллярные белки
фиброин шелка
кератин волос, рогов, копыт
коллаген
и др.
Нитевидные, или фибриллярные белки
фиброин шелка
кератин волос, рогов, копыт
коллаген
и др.

Шаровидные, или глобулярные белки
Шаровидные, или глобулярные белки

Третичная структура белка
основа функциональности белка, которая требует точной пространственной организации
Третичная структура белка
основа функциональности белка, которая требует точной пространственной организации

Четвертичная структура белка
Четвертичная структура белка

Четвертичная структура белка…
Олигомерные белки
Четвертичная структура – размещение в пространстве взаимодействующих
Четвертичная структура белка…
Олигомерные белки
Четвертичная структура – размещение в пространстве взаимодействующих
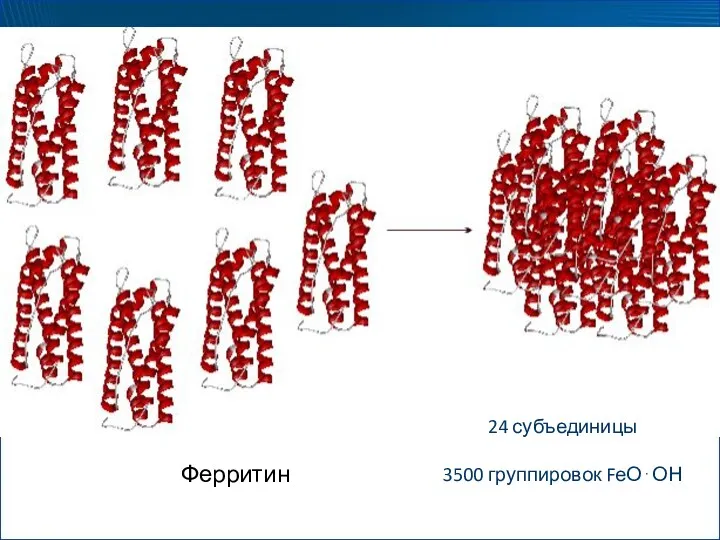
Ферритин
24 субъединицы
3500 группировок FеО⋅ОН
Ферритин
24 субъединицы
3500 группировок FеО⋅ОН

Четвертичная структура белка
гомомерные белки
гетеромерные белки
Объединение в одной структуре нескольких взаимосвязанных
Четвертичная структура белка
гомомерные белки
гетеромерные белки
Объединение в одной структуре нескольких взаимосвязанных

Четвертичная структура белка
Межсубъединичные контакты – система нековалентных взаимодействий
гидрофобные взаимодействия (контактные
Четвертичная структура белка
Межсубъединичные контакты – система нековалентных взаимодействий
гидрофобные взаимодействия (контактные

Четвертичная структура белка
Функциональное значение
Четвертичная структура белка
Функциональное значение

Функции четвертичной структуры
Объединение нескольких взаимосвязанных функций в единой структуре
Архитектурная функция
Формирование
Функции четвертичной структуры
Объединение нескольких взаимосвязанных функций в единой структуре
Архитектурная функция
Формирование

Функции четвертичной структуры
Обеспечение множественных взаимодействий белка с протяженными структурами
ДНК-связывающие белки
Функции четвертичной структуры
Обеспечение множественных взаимодействий белка с протяженными структурами
ДНК-связывающие белки
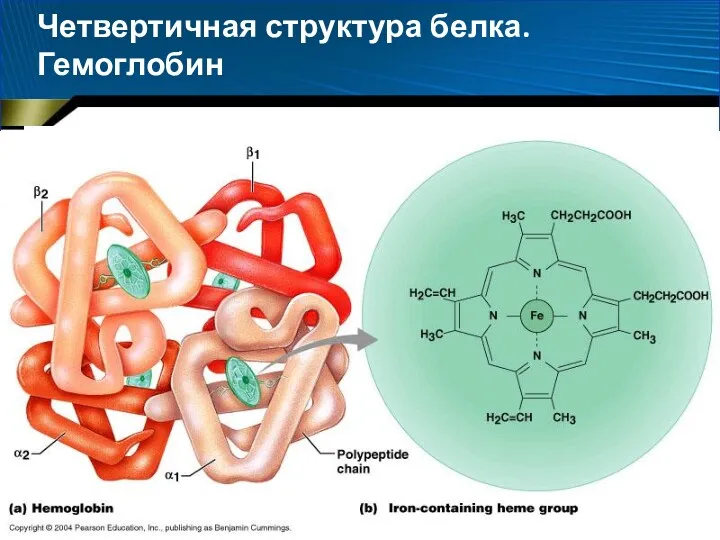
Четвертичная структура белка. Гемоглобин
Четвертичная структура белка. Гемоглобин

Каждый индивидуальный белок характеризуется уникальной структурой, обеспечивающей уникальность его функций
Поэтому выяснение
Каждый индивидуальный белок характеризуется уникальной структурой, обеспечивающей уникальность его функций
Поэтому выяснение

Свойства белков
(см. лабораторные занятия)
Свойства белков
(см. лабораторные занятия)

Классификация белков
Классификация белков

По степени сложности
простые белки (протеины) дают при гидролизе только аминокислоты
По степени сложности
простые белки (протеины) дают при гидролизе только аминокислоты

По степени сложности
сложные белки (протеиды) = протеин + добавочная группа:
Хромопротеины
По степени сложности
сложные белки (протеиды) = протеин + добавочная группа:
Хромопротеины

По форме частиц
фибриллярные (волокнистые) белки (фиброин шелка, кератин волос, коллаген
По форме частиц
фибриллярные (волокнистые) белки (фиброин шелка, кератин волос, коллаген

По растворимости
протеиноиды (склеропротеины) – нерастворимы в обычных растворителях – почти
По растворимости
протеиноиды (склеропротеины) – нерастворимы в обычных растворителях – почти

По аминокислотному составу
протамины – содержат 80–90% арг, простейшие белки, растворяются
По аминокислотному составу
протамины – содержат 80–90% арг, простейшие белки, растворяются

По выполняемым функциям
структурные белки – компоненты клеточных мембран, органелл; коллаген
По выполняемым функциям
структурные белки – компоненты клеточных мембран, органелл; коллаген

По выполняемым функциям
защитные белки: антитела (иммуноглобулины); белки системы свертывания крови
По выполняемым функциям
защитные белки: антитела (иммуноглобулины); белки системы свертывания крови

По выполняемым функциям
резервные белки (овальбумины яиц, белки молока – казеин)
рецепторные
По выполняемым функциям
резервные белки (овальбумины яиц, белки молока – казеин)
рецепторные

По выполняемым функциям
Все белки выполняют энергетическую функцию:
при окислении 1 г белка
По выполняемым функциям
Все белки выполняют энергетическую функцию:
при окислении 1 г белка
 Зайчишка-трусишка, или заяц - хитрец, удалой молодец?
Зайчишка-трусишка, или заяц - хитрец, удалой молодец? Видоизменения корней
Видоизменения корней Отравление животных гербицидами
Отравление животных гербицидами Неклеточные формы жизни. Вирусы и бактериофаги.
Неклеточные формы жизни. Вирусы и бактериофаги. Движение крови в организме
Движение крови в организме Абиотические факторы среды
Абиотические факторы среды Презентация к уроку биологии в 9 классе Формы размножения организмов
Презентация к уроку биологии в 9 классе Формы размножения организмов Ферменты. Тема 2
Ферменты. Тема 2 Топографічна анатомія шиї
Топографічна анатомія шиї Выращивание из семян растения мушмула
Выращивание из семян растения мушмула Синэкология, биоценоз, биогеоценоз
Синэкология, биоценоз, биогеоценоз Птичьи посиделки
Птичьи посиделки Кастрация поросят
Кастрация поросят Суцвіття. Процеси запилення та запліднення квіткових рослин
Суцвіття. Процеси запилення та запліднення квіткових рослин Дикие животные леса. Дидактическая игра
Дикие животные леса. Дидактическая игра Молекулалық генетика негіздері. Ген негіздері. Гендердің интроиды, экзонды орналасуы. Прокориоттардың тұқым қуалауының
Молекулалық генетика негіздері. Ген негіздері. Гендердің интроиды, экзонды орналасуы. Прокориоттардың тұқым қуалауының презентация к уроку биологии 7 класс
презентация к уроку биологии 7 класс презентация к уроку Ветер
презентация к уроку Ветер Анатомия крысы
Анатомия крысы растения огэ
растения огэ Съедобные и ядовитые грибы Красноярского края
Съедобные и ядовитые грибы Красноярского края Анатомия черепа
Анатомия черепа Биотические взаимоотношения организмов
Биотические взаимоотношения организмов Дуб - царь деревьев
Дуб - царь деревьев Функции тонкого и толстого кишечника. Всасывание. Барьерная роль печени. Аппендицит
Функции тонкого и толстого кишечника. Всасывание. Барьерная роль печени. Аппендицит Приспособленность растений к условиям жизни в лесном биогеоценозе.Лабораторная работа.
Приспособленность растений к условиям жизни в лесном биогеоценозе.Лабораторная работа. Муравьиная ферма - Формикарий в домашних условиях
Муравьиная ферма - Формикарий в домашних условиях Механизмы половой дифференцировки
Механизмы половой дифференцировки