- Biochimia. Enzimele

Содержание
- 2. OBIECTIVELE: Noţiune despre enzime şi rolul lor biologic. Asemănările și deosebirile dintre acţiunea enzimelor şi a
- 3. NOȚIUNE DE ENZIMĂ ENZIMĂ – de la grecescul “EN ZYME”- în drojdii Enzime – biocatalizatori de
- 4. ROLUL BIOLOGIC Digestiv (enzimele digestiei) Metabolic MEDICAL ENZIMODIAGNOSTICUL ENZIMOTERAPIA
- 5. Natura chimică a E E- sunt proteine şi posedă toate proprietăţile fizico-chimice specifice acestor molecule (solubilitate,
- 6. Asemănările E cu catalizatorii neorganici catalizează numai reacţiile posibile din punct de vedere energetic nu modifică
- 7. Deosebirile enzimelor (E) de catalizatorii neorganici Viteza catalizei enzimatice este cu mult mai mare decât a
- 8. Deosebirile E de catalizatorii neorganici 4. E catalizează reacţiile fără formarea produselor intermediare – randamentul este
- 9. Structura enzimelor (E) Masa moleculară a E e de mii de ori mai mare decât masa
- 12. Particularităţile CA este o structură tridimensională unicală, formată din radicali ai aminoacizilor distanţaţi în catena primară
- 15. Particularităţile CA 3. Are formă de adâncitură sau cavitate 4. Ocupă o parte relativ mică din
- 16. Organizarea funcţională a enzimelor Centrul alosteric Este centrul reglator Fixează modulatorul alosteric Adiţionarea modulatorului modifică conformaţia
- 17. Reglarea alosterică a activităţii enzimatice
- 18. S COO- NH3+ COO- E Centru Activ Modulator Reglarea alosterică OH NH3 -OOC Centru Alosteric
- 19. Enzime alosterice Moleculele enzimelor alosterice sunt mai mari, mai complexe şi sunt oligomere pare Au cinetica
- 21. Structura enzimelor Din punct de vedere structural deosebim: E simple – alcătuite numai din AA (proteazele,
- 22. PARTEA NEPROTEICĂ A E A: Când componenta neproteică este un ion metalic – este denumită cofactor
- 23. PARTEA NEPROTEICĂ A E B: Când componenta neproteică este o moleculă organică de mici dimensiuni –
- 24. Coenzimele Coenzima strâns legată în structura E – grupare prostetică (FMN; FAD, biotina, acidul lipoic) Coenzima
- 25. Rolul metalelor in cataliza enzimatica Sunt componente esențiale ale centrului catalitic (activ); Participă la legarea S
- 26. Ionii de metale –cofactori ai E După modul de legare şi rolul ionului metalic E sunt:
- 27. EXEMPLE DE METALOENZIME - Fe-enzime: hem (citocromi, catalaze, peroxidaze); - Cu-enzime: citocromoxidaza, superoxid-dismutaza, tiroxin-hidroxilaza; - Mn-enzime:
- 28. Coenzimele (CoE) Sunt parte componentă a centrului activ Contribuie la stabilizarea conformaţiei enzimei Contribuie la fixarea
- 29. Clasificarea CoE CoE vitaminice Tiaminice Flavinice Nicotinamidice Piridoxinice Folice Cobamidice Biotinice lipoice CoE nevitaminice hemurile de
- 31. Coenzimele tiaminice Derivaţii vitaminei B1 (tiamina) CoE - TMP, TDP (TPP)cocarboxilaza, TTP Rolul: Decarboxilarea oxidativă a
- 32. Vitamin B1: Tiamina Coenzima - Tiamin pirofosfat – TPP
- 33. Reacţia sumară a decarboxilării oxidative a piruvatului cu participarea TPP (deriv. Vit. B1)
- 34. Coenzimele flavinice Derivaţi ai vitaminei B2 - riboflavina structură heterociclică numita izoaloxazină
- 35. Coenzimele flavinice CoE- Flavin mononucleotidul (FMN) Flavinadenindinucleotidul (FAD) Rolul: Participă în reacţiile de oxido-reducere: Dezaminarea AA
- 36. Vitamina B2: riboflavina Coenzim - Flavin adenin dinucleotidul (FAD) Coenzima - Flavin mononucleotidul (FMN)
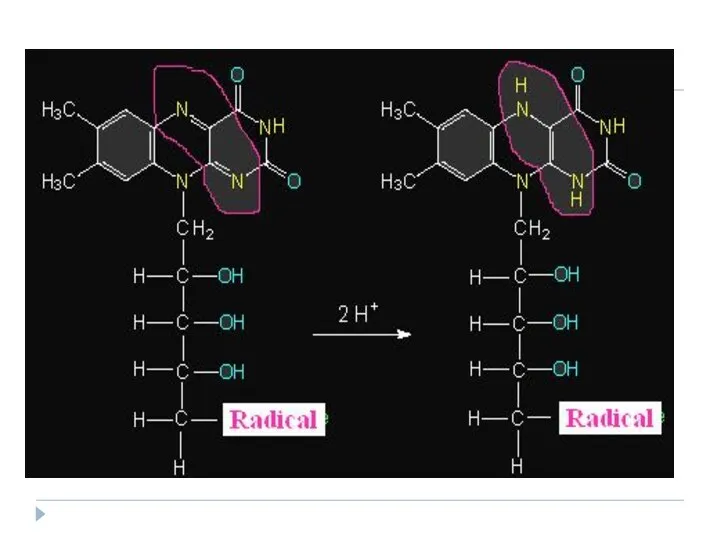
- 37. Coenzimele flavinice
- 39. Reacţia de dehidrogenare a succinatului cu participarea FAD – derivatul vit. B2
- 40. Coenzimele nicotinamidice Sunt derivaţi ai vitaminei PP
- 41. Coenzimele nicotinamidice CoE - NAD şi NADP Rolul Participă în reacţiile de oxido-reducere (dehidrogenarea S –transferul
- 42. Vitamina PP: nicotinamida Coenzimele - nicotinamid adenin dinudleotidul (NAD+) nicotinamid adenin dinudleotid fosfat (NADP+)
- 43. Coenzimele nicotinamidice
- 44. Reacţia de dehidrogenare a malatului cu participarea NAD – derivatul vit. PP
- 45. Coenzimele piridoxinice Derivaţi a vitaminei B6 Co – piridoxalfosfat şi piridoxaminfosfat Rolul: Transaminarea AA Decarboxilarea AA
- 46. Vitamina B6 : Piridoxal fosfat Piridoxamin fosfatul Piridoxina Piridoxal Piridoxamina Coenzimele:
- 47. Reacţia de transaminare a aminoacizilor cu participarea PALP şi PAMP – derivaţii vit. B6
- 48. ACIDUL PANTOTENIC (vitamina B5)
- 49. Coenzimele acidului pantotenic: Coenzima A Fosfopantoteina Rolul metabolic: Coenzima A - la activarea acizilor grași Fosfopantoteina
- 51. Co pantotenice (B5)- HS-CoA biosinteza AG biosinteza Col sinteza corpilor cetonici Oxidarea AG Ciclul Krebs Sinteza
- 52. Co biotinice – vitamina H
- 53. Co biotinice – vitamina H ROL: Participă în reacțiile de carboxilare a piruvatului, acetil Co A
- 54. Coenzimele folice – derivat al acidului folic (vitamina Bc sau vitamina B9) Acidul folic
- 55. Co folice acidul tetrahidrofolic
- 56. ROLUL THF transportator al unor fragmente cu un atom de carbon (grupări monocarbonice): -metil (-CH3), -metilen
- 57. Co cobamidice – B12 -ciancobalamina Vitamină antipernicioasă pentru om şi factor de creştere pentru microorganisme În
- 59. MECANISMUL DE ACŢIUNE AL ENZIMELOR. CLASIFICAREA ŞI NOMENCLATURA ENZIMELOR. CINETICA REACŢIILOR ENZIMATICE
- 60. OBIECTIVE Nomenclatura (denumirea) şi clasificarea enzimelor. Caracteristica generală a claselor şi subclaselor principale de enzime. Numărul
- 61. CLASIFICAREA ENZIMELOR
- 62. NOMENCLATURA ENZIMELOR DENUMIREA COMUNĂ: Numele substratului (S) + sufixul «aza» (glucozidaza, ureaza, lipaza, nucleaza) sau denumirea
- 63. Clasificarea actuală a enzimelor Toate E se împart în: şapte clase, clasele în subclase, subclasele în
- 64. Clasificarea actuală a enzimelor Ex: LDH - 1.1.1.27 Clasa reprezintă tipul de reacţie, catalizat de enzime
- 65. Clasificarea actuală a enzimelor Oxidoreductaze Transferaze Hidrolaze Liaze Izomeraze Ligaze (sintetaze) Translocaze
- 66. Clasificarea enzimelor. - catalizează formarea de legături între carbon şi O, S, N, cuplate cu hidroliza
- 68. 7 clase de Enzime 1. Oxidoreductaze catalizează reacţiii de oxido-reducere SUBCLASELE: Dehidrogenaze Reductaze Oxidaze, Peroxidaze, Hidroxilaze,
- 69. 1. Oxidoreductaze
- 70. 1. Oxidoreductaze
- 71. 2. Transferaze catalizează transferuri grupelor funcţionale de la un substrat la altul (metil, amino, acil) SUBCLASELE:
- 72. 2. Transferaze
- 73. TRANSFERAZE:
- 74. 3. Hidrolaze catalizează scindări de legături covalente cu adiţionarea apei SUBCLASELE: Esteraze Glicozidaze Peptidaze Fosfataze Tiolaze
- 75. H2O H3PO4 Glucoza -6- fosfat Glucoză 3. Hidrolaze
- 76. 3. Hidrolaze
- 77. 4. Liaze catalizează ruperea leg. C-C, C-S şi C-N; fără adiţionarea apei, adiţia la legături duble
- 78. 4. Liaze
- 79. 4. Liaze
- 80. 4. Liaze
- 81. 5. Izomeraze catalizează toate tipurile de transformări în cadrul uneia şi aceleiaşi moleculă SUBCLASELE: Racemaze Epimeraze
- 82. 5. Izomeraze
- 83. 5. Izomeraze
- 84. 5. Izomeraze
- 85. 6. Ligaze (sintetaze) catalizează formarea de legături între carbon şi O, S, N, cuplate cu hidroliza
- 86. 6. Ligaze (sintetaze)
- 87. 6. Ligaze (sintetaze) COOH I C=O I CH3 COOH I C=О I CН2 I CООН ADP
- 88. 7. Translocaze (2018) Catalizează transferul ionilor sau moleculelor prin membrane sau separarea lor in (interiorul; între)
- 89. Specificitatea E S1 S2 S4 este condiţionată de complimentaritatea conformaţională şi electrostatică între CA al E
- 90. Deci fiecare E catalizează un anumit tip de reactii sau transformarea unui anumit S
- 91. SPECIFICITATEA De reacție De substrat: A. Absolută B. Relativă C. stereospecificitate
- 92. Specificitatea: Specificitatea de reacţie: enzimele catalizează un anumit tip de reacţie ce stă la baza clasificării
- 93. 2. Specificitatea de substrat a enzimelor I. Specificitatea absolută de substrat – enzima catalizează transformarea doar
- 94. Specificitatea de substrat a enzimelor II. Specificitatea relativă de substrat – enzima catalizează transformarea unei grup
- 95. Specificitatea de substrat a enzimelor specificitate relativa de substrat - asigura transformarea unui grup de substante
- 96. Specificitatea relativă de substrat a enzimelor Specificitatea absolută de grup – E catalizează transformarea unui grup
- 97. Specificitatea relativă de substrat a enzimelor Specificitatea relativă de grup – enzima catalizează transformarea unei anumite
- 98. Specificitatea relativă de grupă
- 99. Specificitate stereochimică E catalizează transformarea numai a unuia din stereoizomerii posibili (D sau L; sau numai
- 100. Specificitate stereochimică Hexokinaza – specificitate la D- glucoză Oxidazele – specificitate la L-AA Fumaraza – specificitate
- 101. MECANISMUL DE ACȚIUNE AL ENZIMELOR
- 102. Mecanismul de acţiune al E Pentru decurgerea unei reacţii este necesar ca molecula de S şi
- 103. Mecanismul de acţiune al E E - micşorează energia de activare ale reacţiilor chimice. Cu cât
- 104. S + E ↔ E-S → E + P Enzimele reduc energia de activare fara sa
- 105. Enzymes Lower a Reaction’s Activation Energy
- 106. Etapele acţiunii enzimatice E + S ES ES* EP E + P I et. III et.
- 107. Mecanismul de acţiune al enzimelor Prima etapă: Difuzia S spre E şi legarea cu CA al
- 108. Mecanismul de acţiune al E 2. Transformarea complexului primar ES în unul sau cîteva complexe activate
- 109. Ipoteza “lacăt-cheie” (Fischer) şi “coincidenţa forţată” (Koshland) Modelul clasic (Emil Fischer) consideră că potrivirea S cu
- 110. Teoria “cheie-lacăt” - Fisher
- 111. Teoria “coincidenţei induse” - Koshland
- 112. S P1 P2 Conceptul clasic "lacăt- cheie E+S-- ES- E+P
- 113. S P1 P2 Conceptul coencidenţei inductive “coincidenţa forţată” (Kochland)
- 114. Mecanismul de acţiune al E La nivel molecular acţiunea E poate fi lămurită prin următoarele efecte:
- 115. PROPRIETĂŢILE ENZIMELOR
- 116. Obiectivele: Cinetica enzimatică. Influenţa concentraţiei enzimei şi a substratului, a pH-ului şi a temperaturii asupra activităţii
- 117. CINETICA ENZIMATICĂ Studiază viteza reacției enzimatice, luând în considerație factorii fizico-chimici ce o influențează
- 118. VITEZA REACȚIEI ENZIMATICE reprezintă numărul moleculelor de substrat transformate în produs final pe unitatea de timp
- 119. Factorii care influenţiază viteza reacției enzimatice Concentraţia S Concentraţia E Temperatura pH
- 120. Influența [S] asupta vitezei reacției enzimatice Grafic se reprezintă sub formă de o curbă de tip
- 121. Influența [S] asupta vitezei reacției enzimatice Analiza curbei arată 3 zone: Zona “a”- v creste proportional
- 122. Această curbă este numită curba lui Michaelis-Menten şi se exprimă prin ecuaţia: [S] V0=Vmax x _________
- 124. Semnificația lui Km și V max Km-constanta lui Michaelis Menten - este acea concentraţie de S
- 125. Ecuaţia lui Lineweawer- Burk: Din curba lui Michaelis Menten nu poate fi determinată V max (deoarece
- 126. Diagrama lui Lineweawer- Burk: Reciproca ecuatiei Michaelis-Menten
- 127. Influența [E] asupta vitezei reacției în condiţii standard 2 mol de E într-o anumită perioadă de
- 128. Influența t° asupta vitezei reacției
- 129. Termolabilitatea (t°) Unele E a microorganismelor termofile sunt active la t de 80°C La t joase
- 130. Influența pH asupta vitezei reacției Fiecare E are pH optim propriu (manifestă activitate max). Majoritatea E
- 131. Acţiunea pH asupra activităţii enzimatice La E digestive pH optim este cel al sediului lor de
- 132. Acţiunea pH asupra activităţii enzimatice
- 133. Acţiunea pH asupra activităţii enzimatice pH optim e dependent de: gradul de ionizare a grupelor funcţionale,
- 134. Acţiunea pH asupra activităţii enzimatice Centrul activ al enzimelor contine grupe ionizabile, acide sau bazice, deci
- 135. Influenta efectorilor enzimatici Efectorii enzimatici sunt substante cu structuri chimice variate care aduse in mediul de
- 136. Inhibitorii enzimatici Inhibitorii enzimatici sunt compusi care influenteaza negativ activitatea enzimelor pe care o pot anula
- 137. Inhibiţia enzimatică Deosebim: inhibiţie nespecifică (T, pH, agenţii denaturării ) inhibiţie specifică Inhibiţia poate fi reversibilă
- 138. EXEMPLE DE INHIBITORI IREVERSIBILI Compușii organofosforici (Diizopropilfluorfosfatul) (toxina neuroparalitica) se fixează de OH-Ser în CA a
- 139. Inhibiţie ireversibilă
- 140. INHIBIȚIE IREVERSIBILĂ Exemple: Diizopropilfluorfosfatul se fixează de OH-serinei în CA a proteinazelor
- 141. INHIBIȚIE IREVERSIBILĂ
- 142. Tipuri de inhibitie revesibilă Deosebim: Inhibiţie competitivă Inhibiţie necompetitivă (noncompetitivă) Inhibiţie uncompetetivă Inhibiţie alosterică
- 143. Inhibiţia competitivă I se aseamănă după structură cu S. Apare o competiţie dintre I şi S
- 144. Inhibiţia competitivă
- 145. Inhibiţia competitivă Viteza de reactie atinge Vmax ca si cum I nu ar fi prezent, daca
- 146. Exemple de I competitivi: inhibiţia SDH cu malonat (SDH -oxideaza succinatul in fumarat). Malonatul inhibă aceasta
- 147. Exemple de I competitivi: Sulfamidele –substituie acidul p-amino-benzoic din a. folic, indispensabil pentru creşterea microorganismelor, împedicând
- 148. Exemple de I competitivi: Sulfamidele –substituie acidul p-amino-benzoic din a. folic, indispensabil pentru creşterea microorganismelor, împedicând
- 149. Exemple de I competitivi: Alopurinol – administrat ăn tratamentul gutei analog structural al hipoxantinei – inhibă
- 150. Exemple de I competitivi: 5 fluoruracilul - Inhibitorul timidilat sintazei Metotrexatul - inhibitorul dihidrofolatreductazei –Astfel se
- 151. Inhibiţia necompetitivă Inhibitorul nu se aseamănă ca structură cu S I şi S se leagă simultan
- 152. Inhibiţie necompetitiva
- 153. Inhibiţie necompetitiva In cazul inhibitiei necompetitive, I chiar daca nu se leaga la centru activ al
- 154. Inhibiţia necompetitivă
- 155. Tipuri de inhibitori necompetitivi: cianurile, CO se fixează cu Fe 3+ din citocromoxidază ---se întrerupe LR
- 156. Inhibiţia necompetitivă I poate fi înlăturat de substanţe care îl leagă – numite reactivatori
- 157. TABEL COMPARATIV INHIBITORI REVERSIBILI !
- 158. Efectul inhibitorilor asupra vitezei de reactie
- 159. Inhibiţia uncompetetivă – E+S----ES +I-----ESI I se combina cu complexul ES formand un complex ESI ce
- 160. Inhibiţia uncompetetivă –
- 161. Inhibiţia uncompetitivă Lineweawer- Burk
- 162. Alte tipuri de inhibiție inhibiţia prin modificarea covalentă a moleculei E - prin fosforilare pe baza
- 163. REGLAREA ACTIVITATII ENZIMELOR
- 164. Mecanismele de activare a E Sunt : 1. nespecifice: temperatura , iradierea 2. specifice Se activează
- 165. I. REGLAREA CANTITATIVA Functie de dinamica SINTEZA/DEGRADARE Enzime inductibile Ks > Kd Enzime represibile Kd >
- 166. II. REGLAREA CALITATIVA Mecanismele de activare a E Deosebim următoarele tipuri de reglare a activităţii enzimatice:
- 167. II. REGLAREA CALITATIVA II.1.Reglarea alosterica (necovalenta)
- 168. Enzime alosterice. Efectori alosterici Sunt proteine oligomere alcatuite din mai multe subunitati identice sau diferite, in
- 169. Enzime alosterice. Efectori alosterici Pe langa centri activi, monomerii prezinta si centri alosterici de care se
- 170. II. REGLAREA CALITATIVA II.1.Reglarea alosterica de tip heterotrop
- 171. Cinetica enzimelor alosterice („ ne-Michaeliene") Dependenţa sigmoidală a V0 în funcţie de [S] Enzimele alosterice, spre
- 172. Tipuri de reglare alosterică în funcţie de poziţia relativă a efectorului faţă de E în calea
- 173. Reglarea activitatii enzimatice
- 174. Retroinhibiţie A B C D P E E 1 E 2 3 E 4
- 175. Reglarea activitatii enzimatice
- 176. Reglarea covalentă - Proteoliză limitată Unele enzime (proteine) se sintetizează în forma neactivă de precursor –
- 177. Proteoliză limitată -este scindarea unui sector al catenei în rezultatul căreia E se restructurează şi se
- 178. Importanța biologică a prezenţei formelor neactive Zimogenii sunt produse la locurile de sinteză (mucoasa gastrică pentru
- 179. Reglarea covalentă - fosforilare-defosforilare unele E sunt active în forma fosforilată, iar altele în forma defosforilată.
- 180. Reglarea covalentă - fosforilare-defosforilare Reacţiile de fosforilare sunt catalizate de kinaze specifice. E-OH + ATP --------→
- 181. Autostructurarea cuaternară Este caracteristică E ce posedă structură cuaternară Fiecare protomer în parte nu e activ
- 182. E CA Allos S M CA Reglarea alosterică Modulator pozitiv
- 183. Izoenzimele- izoE forme moleculare multiple ale E, ce catalizează aceeaşi reacţie chimică, dar diferă prin structură,
- 184. IzoE diferă între ele prin: sarcina electrică (ce permite separarea lor prin electroforeză); V max de
- 185. STRUCTURA IZOE Sunt E oligomere, cu structură cuaternară, alcătuite din cel puţin 2 protomeri diferiţi Ex.
- 187. Izoformele lactat dehidrogenazei (LDH) LDH-1 (4H) - in inima LDH-2 (3H1M) - in sistemul reticuloendotelial LDH-3
- 188. ROLUL izoenzimelor în controlul metabolic ( faciliteaza adaptarea metabolismului in diferite ţesuturi.) Ex: in miocard predomina
- 189. LDH Norma- 100-190U/L Creşterea de LDH 1 şi LDH2: infarctul miocardic anemia hemolitică/ megaloblastică LDH5 crescut
- 190. Exemple de izoenzime: Creatinfosfokinaza -2 tipuri de monomeri: M-Muscle şi B –brain LDH MDH Aldolaza Fosfataza
- 191. Creatinfosfokinaza (CPK)
- 194. Sistemele polienzimatice Fiecare celulă a organismului conţine setul său specific de E. Unele se găsesc în
- 195. Tipurile de organizare a sistemelor polienzimatice Se cunosc urmatoarele tipuri de organizare a sistemelor polienzimatice: -
- 196. Organizarea funcţională enzimele sunt asociate în sistemul polienzimatic cu ajutorul metaboliţilor, care difuzează de la o
- 197. Organizarea structural-funcţională E sunt fixate prin legături slabe pe o proteină “centrală”, care poate fi chiar
- 198. Organizarea structural-funcţională Ex.- complexul polienzimatic piruvatdehidrogenazic, constituit din 3 E şi 5 Co sintetaza acizilor graşi
- 199. Tipul mixt de organizare reprezintă o îmbinare a ambelor tipuri de organizare, adică o parte din
- 200. Unităţile de măsurare a activităţii E 1 UI – cantitatea de E care catalizează transformarea unui
- 201. 1 cat = 6 · 107 UI 1 UI = 16.67 · 10-9 cat
- 202. Unităţile de măsurare a activităţii E Activitatea specifică – reprezintă numărul de unităţi enzimatice per mg
- 203. Metodele de separare şi purificare ale E Dializă Salifiere Cromatografie Gel-filtrare Electroforeză Cea mai eficientă –
- 204. Metodele de determinare a activităţii E Viteza reacţiei este proporţională cu Viteza consumului substratului Viteza formării
- 205. Deosebirea privind componenţa enzimatică a organelor şi ţesuturilor. Enzimele organospecifice. Enzimele indicatorii – sunt localizate intracelular:
- 208. Utilizarea E în practica medicală Preparatele farmaceutice contemporane sunt asociate şi conţin ca regulă următoarele enzime:
- 209. Utilizarea E în practica medicală Efectele exercitate de preparatele enzimatice De substituţie (E digestive) Fibrinolitică şi
- 210. Terapia cu enzime Enzimele sunt agenţi terapeutici unici ce produc efecte importante şi specifice. Motivele care
- 211. Tehnici propuse pentru optimizarea proprietăţilor terapeutice ale enzimelor. - prin N-acilare a fost crescută semiviata asparaginazei,
- 213. Скачать презентацию

OBIECTIVELE:
Noţiune despre enzime şi rolul lor biologic. Asemănările și deosebirile dintre
OBIECTIVELE:
Noţiune despre enzime şi rolul lor biologic. Asemănările și deosebirile dintre

NOȚIUNE DE ENZIMĂ
ENZIMĂ – de la grecescul
“EN ZYME”- în drojdii
Enzime –
NOȚIUNE DE ENZIMĂ
ENZIMĂ – de la grecescul
“EN ZYME”- în drojdii
Enzime –

ROLUL BIOLOGIC
Digestiv (enzimele digestiei)
Metabolic
MEDICAL
ENZIMODIAGNOSTICUL
ENZIMOTERAPIA
ROLUL BIOLOGIC
Digestiv (enzimele digestiei)
Metabolic
MEDICAL
ENZIMODIAGNOSTICUL
ENZIMOTERAPIA

Natura chimică a E
E- sunt proteine şi posedă toate proprietăţile fizico-chimice
Natura chimică a E
E- sunt proteine şi posedă toate proprietăţile fizico-chimice

Asemănările E cu catalizatorii neorganici
catalizează numai reacţiile posibile din punct de
Asemănările E cu catalizatorii neorganici
catalizează numai reacţiile posibile din punct de

Deosebirile enzimelor (E) de catalizatorii neorganici
Viteza catalizei enzimatice este cu mult
Deosebirile enzimelor (E) de catalizatorii neorganici
Viteza catalizei enzimatice este cu mult

Deosebirile E de catalizatorii neorganici
4. E catalizează reacţiile fără formarea produselor
Deosebirile E de catalizatorii neorganici
4. E catalizează reacţiile fără formarea produselor

Structura enzimelor (E)
Masa moleculară a E e de mii de ori
Structura enzimelor (E)
Masa moleculară a E e de mii de ori
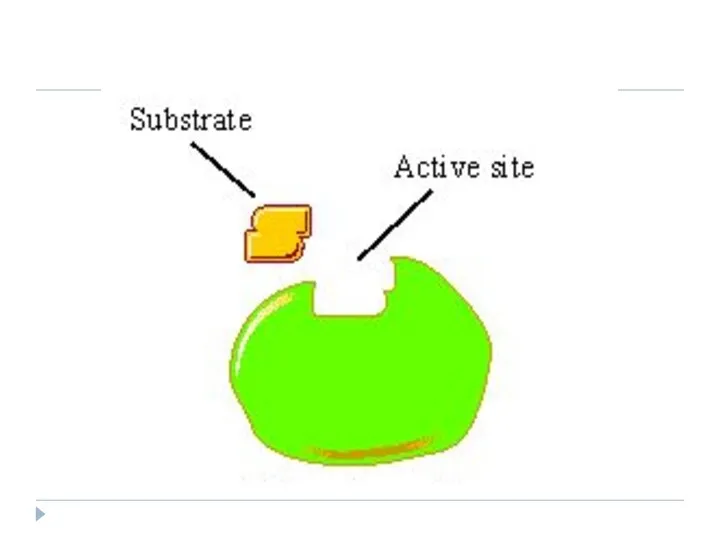
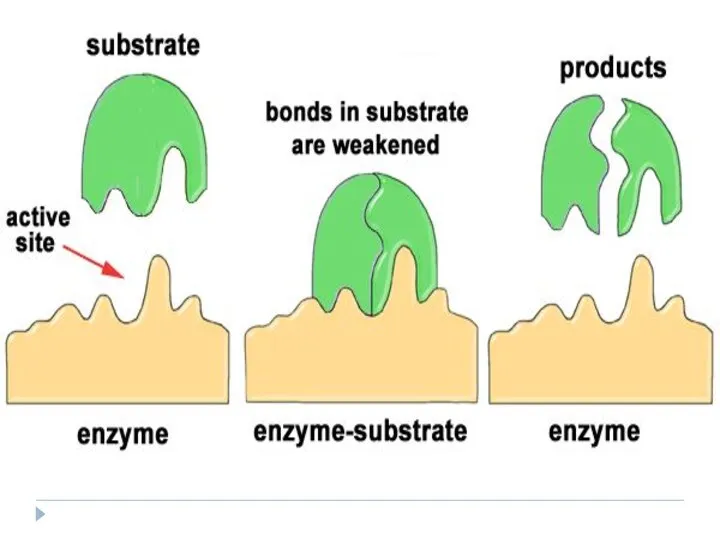

Particularităţile CA
este o structură tridimensională unicală, formată din radicali ai aminoacizilor
Particularităţile CA
este o structură tridimensională unicală, formată din radicali ai aminoacizilor

Particularităţile CA
3. Are formă de adâncitură sau cavitate
4. Ocupă o
Particularităţile CA
3. Are formă de adâncitură sau cavitate
4. Ocupă o

Organizarea funcţională a enzimelor
Centrul alosteric
Este centrul reglator
Fixează modulatorul alosteric
Adiţionarea modulatorului
Organizarea funcţională a enzimelor
Centrul alosteric
Este centrul reglator
Fixează modulatorul alosteric
Adiţionarea modulatorului
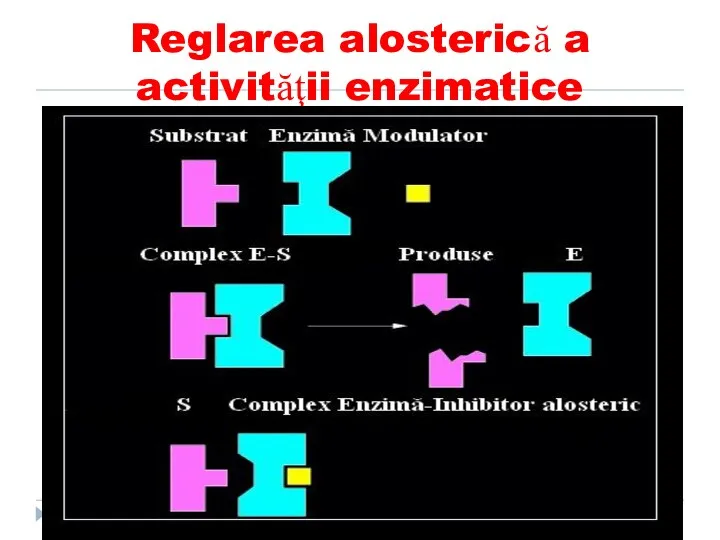
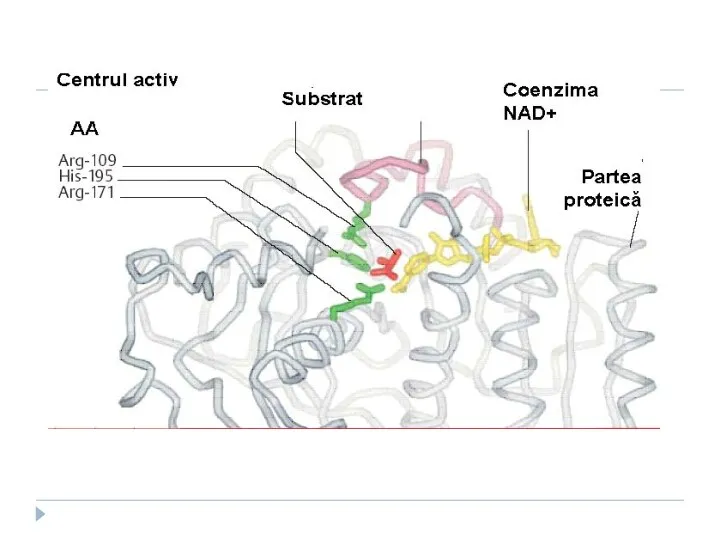
Reglarea alosterică a activităţii enzimatice
Reglarea alosterică a activităţii enzimatice
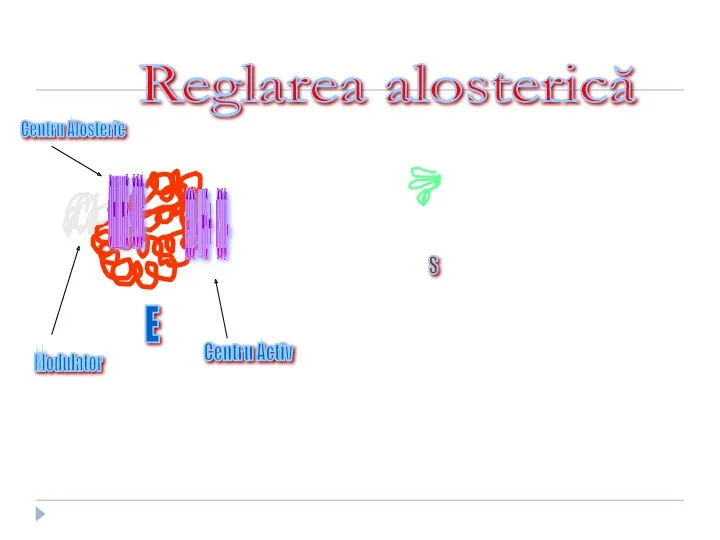
S
COO- NH3+ COO-
E
Centru Activ
Modulator
Reglarea alosterică
S
COO- NH3+ COO-
E
Centru Activ
Modulator
Reglarea alosterică

Enzime alosterice
Moleculele enzimelor alosterice sunt mai mari, mai complexe şi sunt
Enzime alosterice
Moleculele enzimelor alosterice sunt mai mari, mai complexe şi sunt

Structura enzimelor
Din punct de vedere structural deosebim:
E simple – alcătuite
Structura enzimelor
Din punct de vedere structural deosebim:
E simple – alcătuite

PARTEA NEPROTEICĂ A E
A: Când componenta neproteică este un ion metalic
PARTEA NEPROTEICĂ A E
A: Când componenta neproteică este un ion metalic

PARTEA NEPROTEICĂ A E
B: Când componenta neproteică este o moleculă organică
PARTEA NEPROTEICĂ A E
B: Când componenta neproteică este o moleculă organică

Coenzimele
Coenzima strâns legată în structura E – grupare prostetică (FMN; FAD,
Coenzimele
Coenzima strâns legată în structura E – grupare prostetică (FMN; FAD,
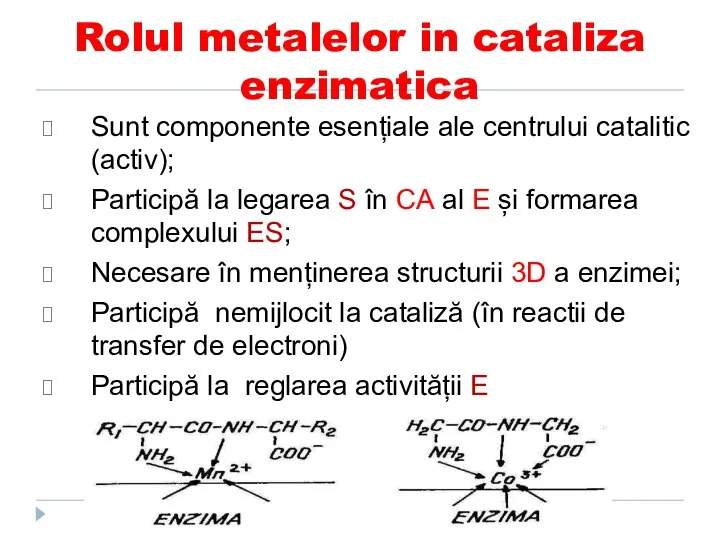
Rolul metalelor in cataliza enzimatica
Sunt componente esențiale ale centrului catalitic (activ);
Participă
Rolul metalelor in cataliza enzimatica
Sunt componente esențiale ale centrului catalitic (activ);
Participă

Ionii de metale –cofactori ai E
După modul de legare şi rolul
Ionii de metale –cofactori ai E
După modul de legare şi rolul

EXEMPLE DE METALOENZIME
- Fe-enzime: hem (citocromi, catalaze, peroxidaze);
- Cu-enzime: citocromoxidaza, superoxid-dismutaza,
EXEMPLE DE METALOENZIME
- Fe-enzime: hem (citocromi, catalaze, peroxidaze);
- Cu-enzime: citocromoxidaza, superoxid-dismutaza,

Coenzimele (CoE)
Sunt parte componentă a centrului activ
Contribuie la stabilizarea conformaţiei enzimei
Contribuie
Coenzimele (CoE)
Sunt parte componentă a centrului activ
Contribuie la stabilizarea conformaţiei enzimei
Contribuie

Clasificarea CoE
CoE vitaminice
Tiaminice
Flavinice
Nicotinamidice
Piridoxinice
Folice
Cobamidice
Biotinice
lipoice
CoE nevitaminice
hemurile de diversă natură
Nucleotidele
Fosfaţii monozaharidelor
Clasificarea CoE
CoE vitaminice
Tiaminice
Flavinice
Nicotinamidice
Piridoxinice
Folice
Cobamidice
Biotinice
lipoice
CoE nevitaminice
hemurile de diversă natură
Nucleotidele
Fosfaţii monozaharidelor
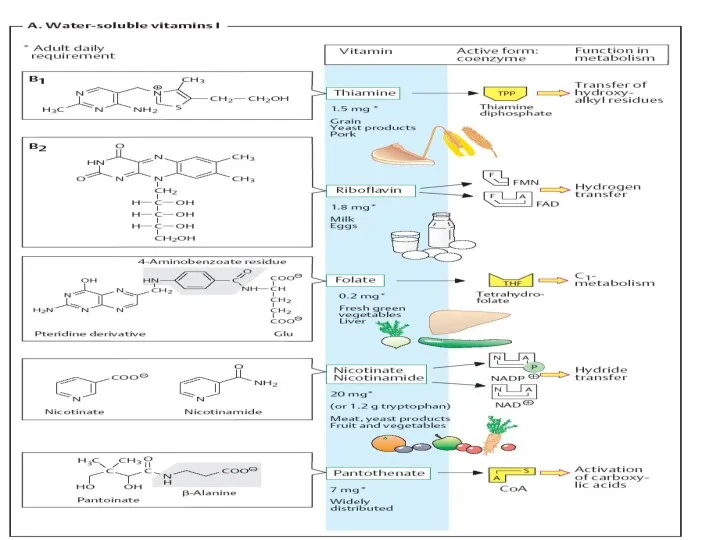

Coenzimele tiaminice
Derivaţii vitaminei B1 (tiamina)
CoE - TMP, TDP (TPP)cocarboxilaza, TTP
Rolul:
Decarboxilarea oxidativă
Coenzimele tiaminice
Derivaţii vitaminei B1 (tiamina)
CoE - TMP, TDP (TPP)cocarboxilaza, TTP
Rolul:
Decarboxilarea oxidativă
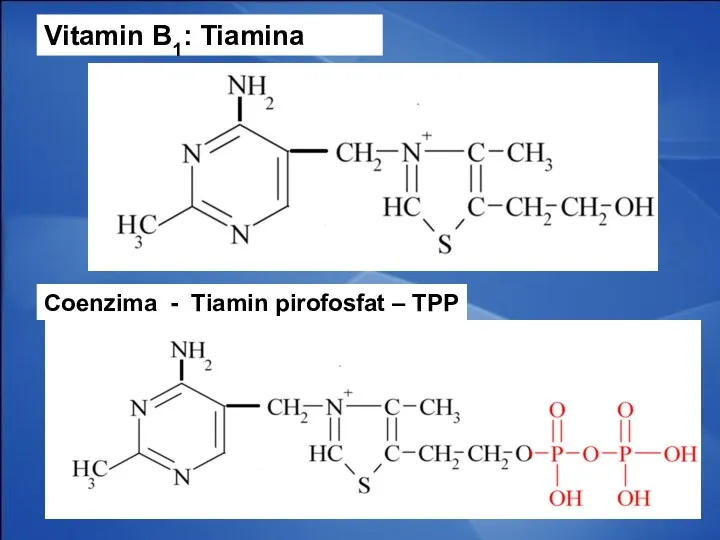
Vitamin B1: Tiamina
Coenzima - Tiamin pirofosfat – TPP
Vitamin B1: Tiamina
Coenzima - Tiamin pirofosfat – TPP
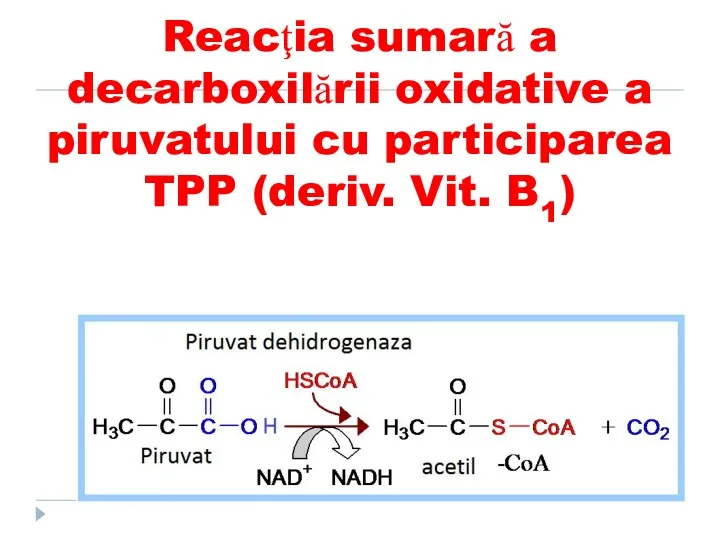
Reacţia sumară a decarboxilării oxidative a piruvatului cu participarea TPP (deriv.
Reacţia sumară a decarboxilării oxidative a piruvatului cu participarea TPP (deriv.

Coenzimele flavinice
Derivaţi ai vitaminei B2 - riboflavina
structură heterociclică numita izoaloxazină
Coenzimele flavinice
Derivaţi ai vitaminei B2 - riboflavina
structură heterociclică numita izoaloxazină

Coenzimele flavinice
CoE- Flavin mononucleotidul (FMN)
Flavinadenindinucleotidul (FAD)
Rolul:
Participă în reacţiile de oxido-reducere:
Dezaminarea
Coenzimele flavinice
CoE- Flavin mononucleotidul (FMN)
Flavinadenindinucleotidul (FAD)
Rolul:
Participă în reacţiile de oxido-reducere:
Dezaminarea
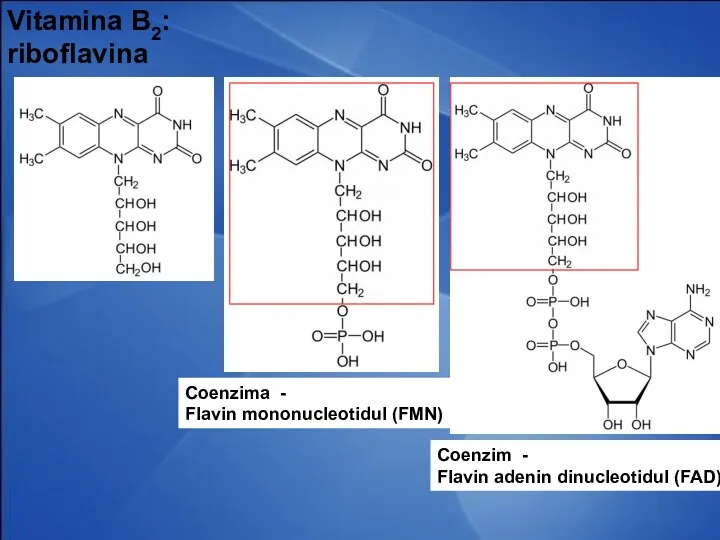
Vitamina B2:
riboflavina
Coenzim -
Flavin adenin dinucleotidul (FAD)
Coenzima -
Flavin
Vitamina B2:
riboflavina
Coenzim -
Flavin adenin dinucleotidul (FAD)
Coenzima -
Flavin
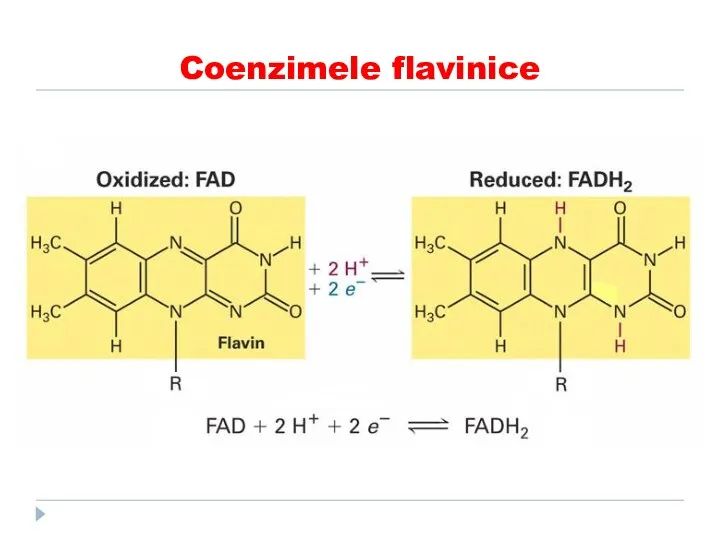
Coenzimele flavinice
Coenzimele flavinice
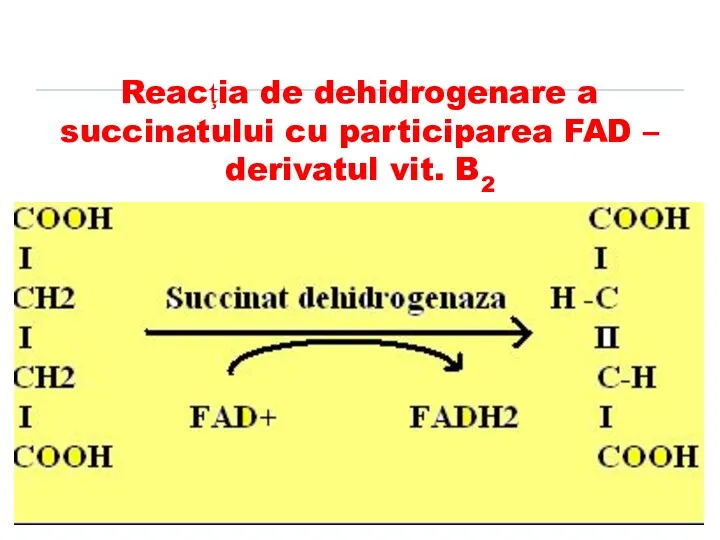
Reacţia de dehidrogenare a succinatului cu participarea FAD – derivatul vit.
Reacţia de dehidrogenare a succinatului cu participarea FAD – derivatul vit.
Coenzimele nicotinamidice
Sunt derivaţi ai vitaminei PP
Coenzimele nicotinamidice
Sunt derivaţi ai vitaminei PP
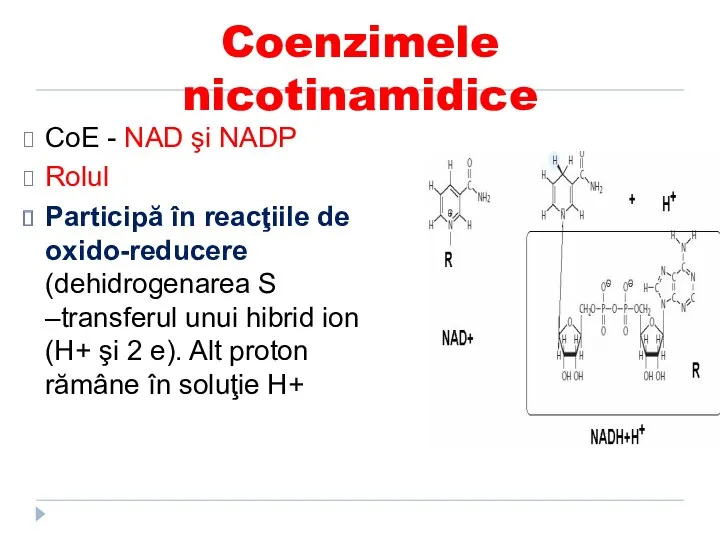
Coenzimele nicotinamidice
CoE - NAD şi NADP
Rolul
Participă în reacţiile de oxido-reducere (dehidrogenarea
Coenzimele nicotinamidice
CoE - NAD şi NADP
Rolul
Participă în reacţiile de oxido-reducere (dehidrogenarea
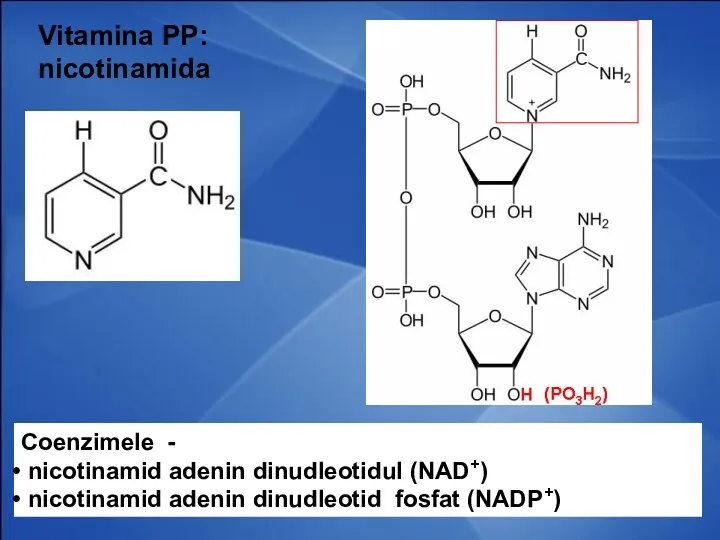
Vitamina PP:
nicotinamida
Coenzimele -
nicotinamid adenin dinudleotidul (NAD+)
nicotinamid adenin
Vitamina PP:
nicotinamida
Coenzimele -
nicotinamid adenin dinudleotidul (NAD+)
nicotinamid adenin

Coenzimele nicotinamidice
Coenzimele nicotinamidice
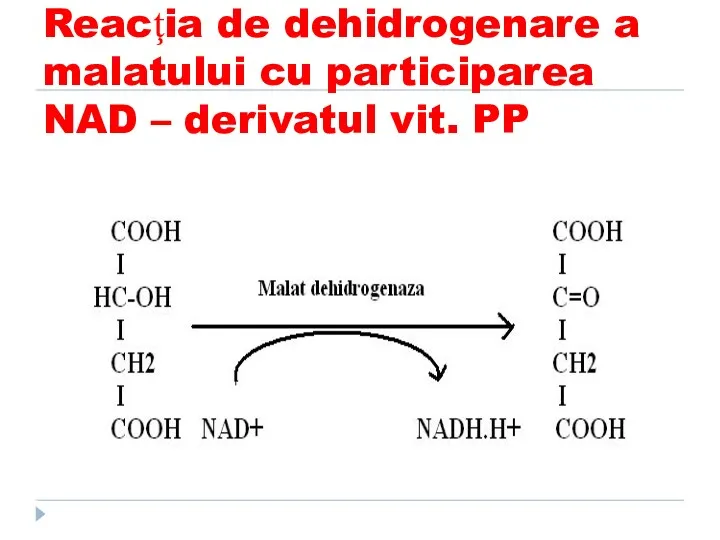
Reacţia de dehidrogenare a malatului cu participarea NAD – derivatul vit.
Reacţia de dehidrogenare a malatului cu participarea NAD – derivatul vit.
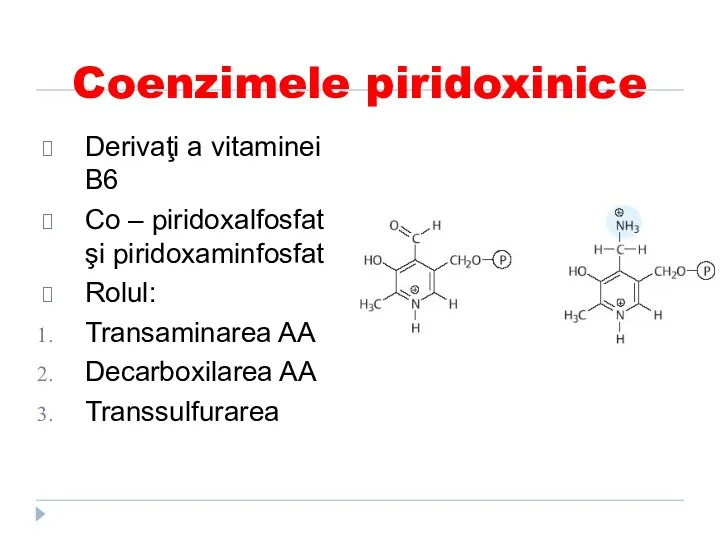
Coenzimele piridoxinice
Derivaţi a vitaminei B6
Co – piridoxalfosfat şi piridoxaminfosfat
Rolul:
Transaminarea AA
Decarboxilarea AA
Transsulfurarea
Coenzimele piridoxinice
Derivaţi a vitaminei B6
Co – piridoxalfosfat şi piridoxaminfosfat
Rolul:
Transaminarea AA
Decarboxilarea AA
Transsulfurarea
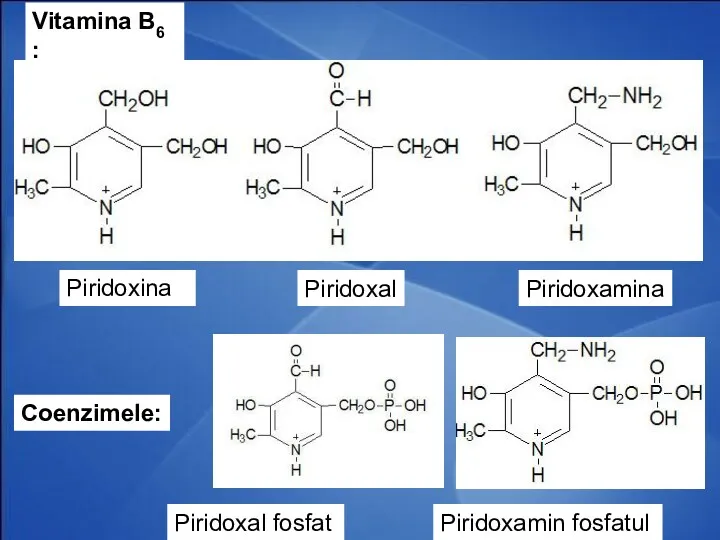
Vitamina B6 :
Piridoxal fosfat
Piridoxamin fosfatul
Piridoxina
Piridoxal
Piridoxamina
Coenzimele:
Vitamina B6 :
Piridoxal fosfat
Piridoxamin fosfatul
Piridoxina
Piridoxal
Piridoxamina
Coenzimele:
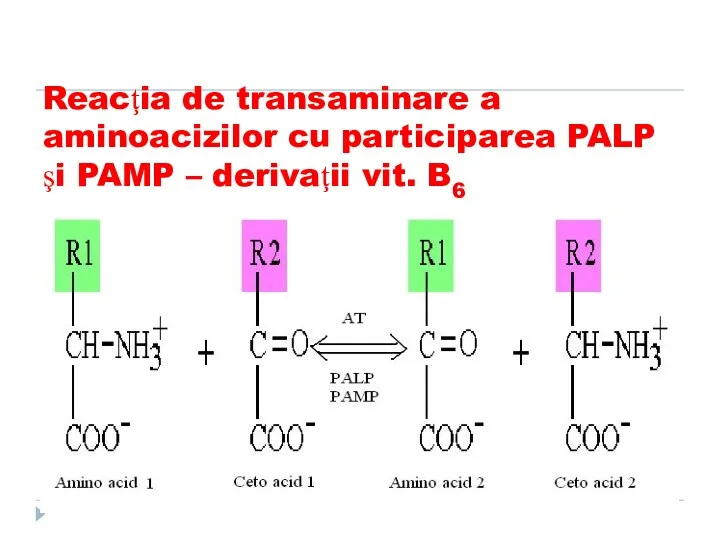
Reacţia de transaminare a aminoacizilor cu participarea PALP şi PAMP –
Reacţia de transaminare a aminoacizilor cu participarea PALP şi PAMP –
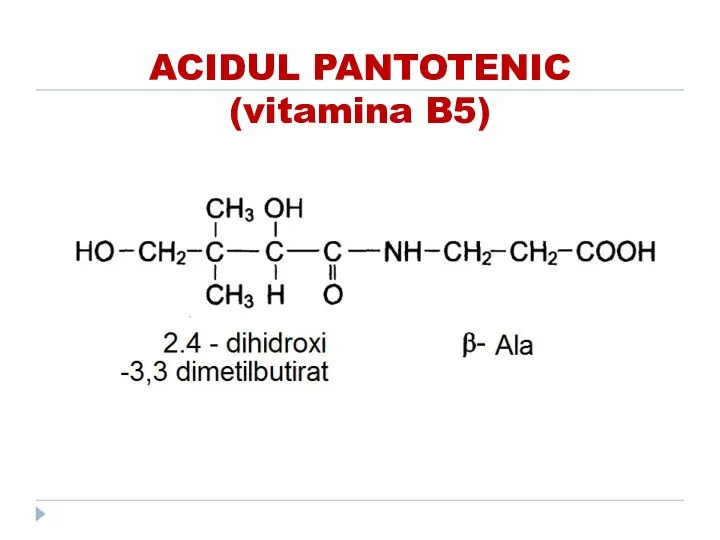
ACIDUL PANTOTENIC
(vitamina B5)
ACIDUL PANTOTENIC
(vitamina B5)

Coenzimele acidului pantotenic:
Coenzima A
Fosfopantoteina
Rolul metabolic:
Coenzima A - la activarea
Coenzimele acidului pantotenic:
Coenzima A
Fosfopantoteina
Rolul metabolic:
Coenzima A - la activarea
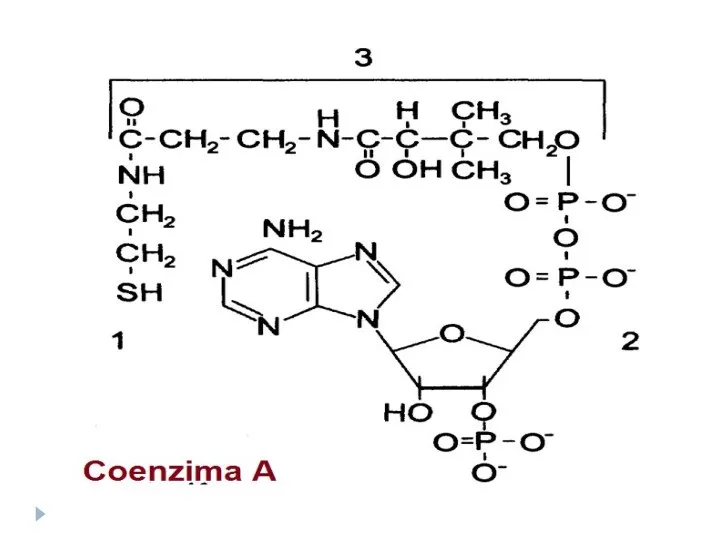

Co pantotenice (B5)- HS-CoA
biosinteza AG
biosinteza Col
sinteza corpilor cetonici
Oxidarea AG
Ciclul Krebs
Sinteza aminolevulinatului
DOP
DO
Co pantotenice (B5)- HS-CoA
biosinteza AG
biosinteza Col
sinteza corpilor cetonici
Oxidarea AG
Ciclul Krebs
Sinteza aminolevulinatului
DOP
DO
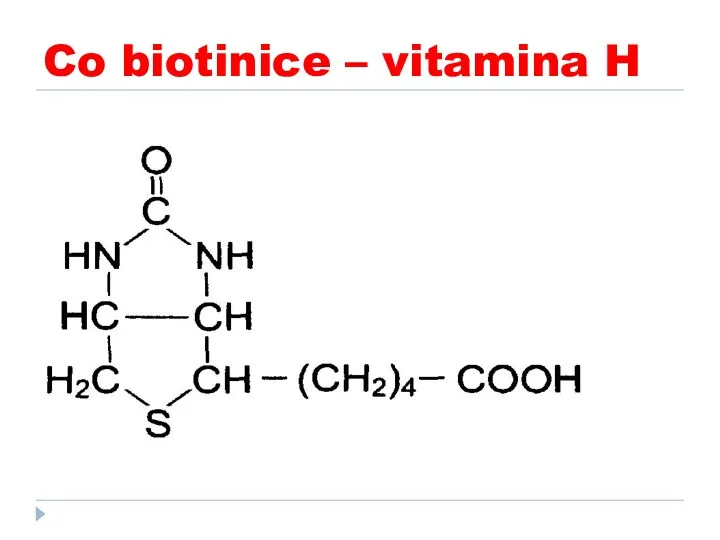
Co biotinice – vitamina H
Co biotinice – vitamina H

Co biotinice – vitamina H
ROL:
Participă în reacțiile de carboxilare a piruvatului,
Co biotinice – vitamina H
ROL:
Participă în reacțiile de carboxilare a piruvatului,
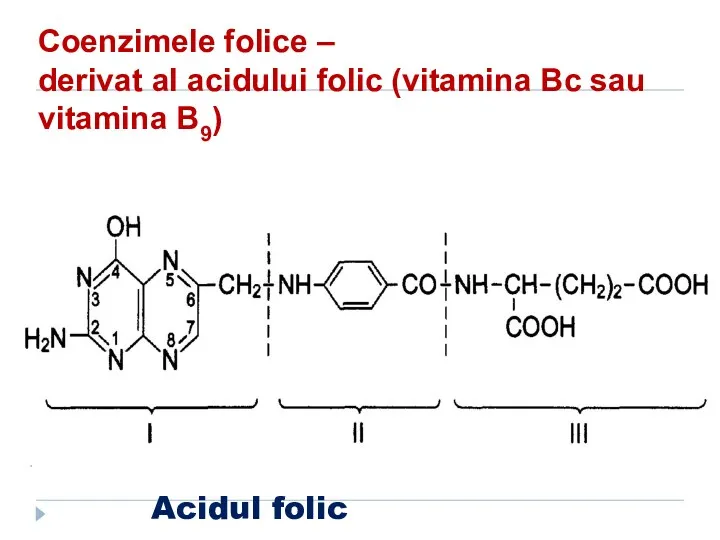
Coenzimele folice –
derivat al acidului folic (vitamina Bc sau vitamina
Coenzimele folice – derivat al acidului folic (vitamina Bc sau vitamina

Co folice
acidul tetrahidrofolic
Co folice
acidul tetrahidrofolic

ROLUL THF
transportator al unor fragmente cu un atom de carbon
ROLUL THF
transportator al unor fragmente cu un atom de carbon

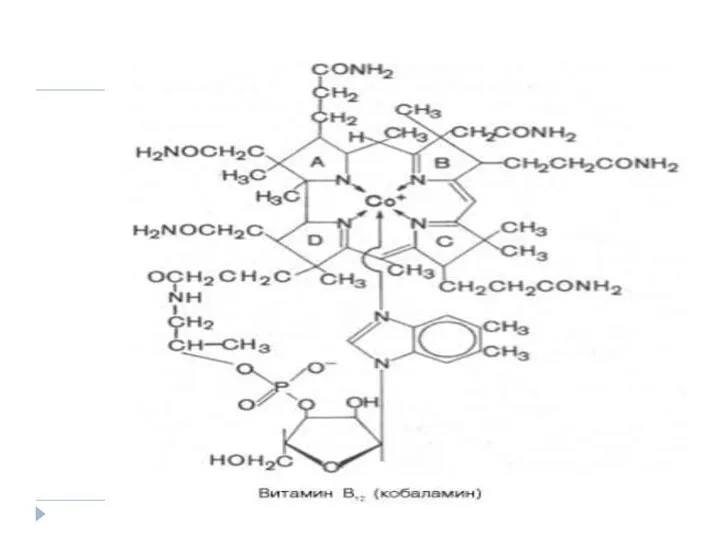
Co cobamidice – B12 -ciancobalamina
Vitamină antipernicioasă pentru om şi factor de
Co cobamidice – B12 -ciancobalamina
Vitamină antipernicioasă pentru om şi factor de

MECANISMUL DE ACŢIUNE AL ENZIMELOR.
CLASIFICAREA ŞI NOMENCLATURA ENZIMELOR.
CINETICA REACŢIILOR
MECANISMUL DE ACŢIUNE AL ENZIMELOR.
CLASIFICAREA ŞI NOMENCLATURA ENZIMELOR.
CINETICA REACŢIILOR

OBIECTIVE
Nomenclatura (denumirea) şi clasificarea enzimelor. Caracteristica generală a claselor şi subclaselor
OBIECTIVE
Nomenclatura (denumirea) şi clasificarea enzimelor. Caracteristica generală a claselor şi subclaselor

CLASIFICAREA ENZIMELOR
CLASIFICAREA ENZIMELOR

NOMENCLATURA ENZIMELOR
DENUMIREA COMUNĂ:
Numele substratului (S) + sufixul «aza» (glucozidaza, ureaza, lipaza,
NOMENCLATURA ENZIMELOR
DENUMIREA COMUNĂ:
Numele substratului (S) + sufixul «aza» (glucozidaza, ureaza, lipaza,

Clasificarea actuală a enzimelor
Toate E se împart în:
şapte clase,
Clasificarea actuală a enzimelor
Toate E se împart în:
şapte clase,

Clasificarea actuală a enzimelor
Ex: LDH - 1.1.1.27
Clasa reprezintă tipul de
Clasificarea actuală a enzimelor
Ex: LDH - 1.1.1.27
Clasa reprezintă tipul de

Clasificarea actuală a enzimelor
Oxidoreductaze
Transferaze
Hidrolaze
Liaze
Izomeraze
Ligaze (sintetaze)
Translocaze
Clasificarea actuală a enzimelor
Oxidoreductaze
Transferaze
Hidrolaze
Liaze
Izomeraze
Ligaze (sintetaze)
Translocaze
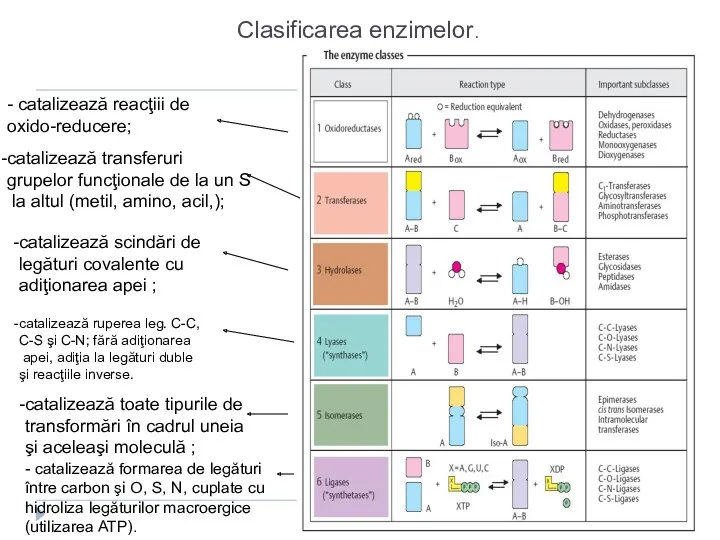
Clasificarea enzimelor.
- catalizează formarea de legături între carbon şi
Clasificarea enzimelor.
- catalizează formarea de legături între carbon şi
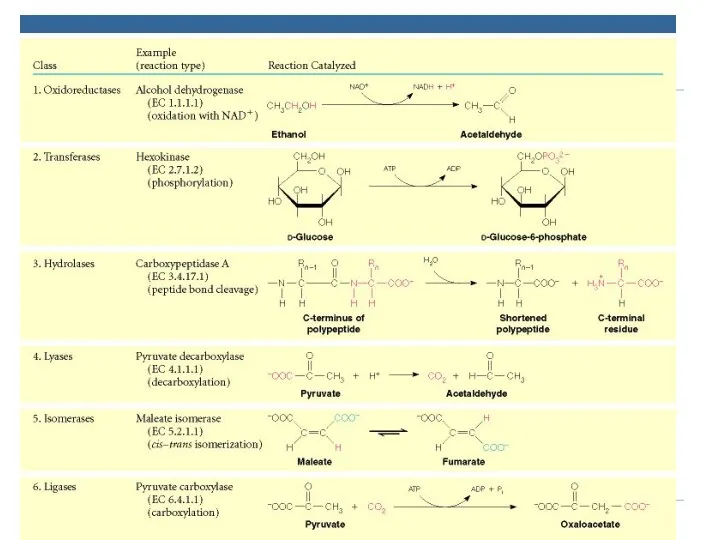

7 clase de Enzime
1. Oxidoreductaze
catalizează reacţiii de oxido-reducere
SUBCLASELE:
Dehidrogenaze
Reductaze
Oxidaze,
Peroxidaze,
Hidroxilaze,
7 clase de Enzime
1. Oxidoreductaze
catalizează reacţiii de oxido-reducere
SUBCLASELE:
Dehidrogenaze
Reductaze
Oxidaze,
Peroxidaze,
Hidroxilaze,
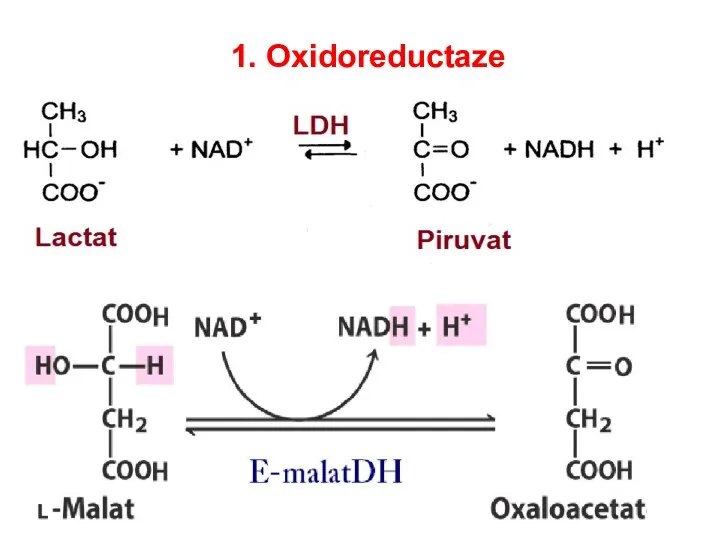
1. Oxidoreductaze
1. Oxidoreductaze
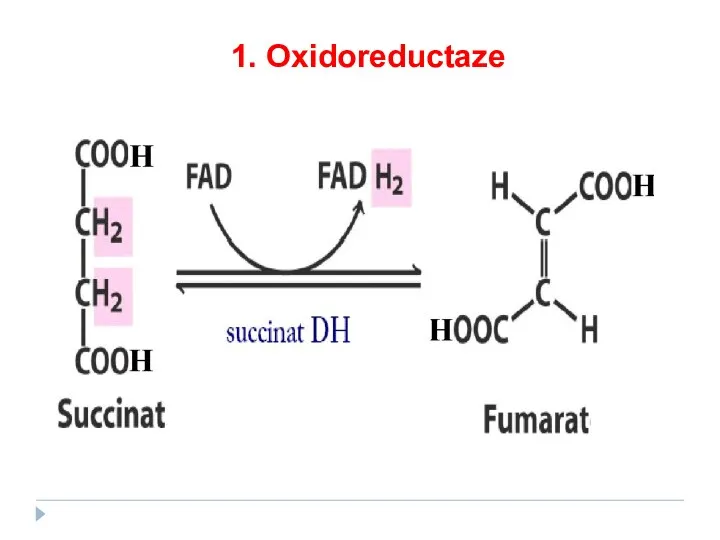
1. Oxidoreductaze
1. Oxidoreductaze

2. Transferaze
catalizează transferuri grupelor funcţionale de la un substrat la altul
2. Transferaze
catalizează transferuri grupelor funcţionale de la un substrat la altul
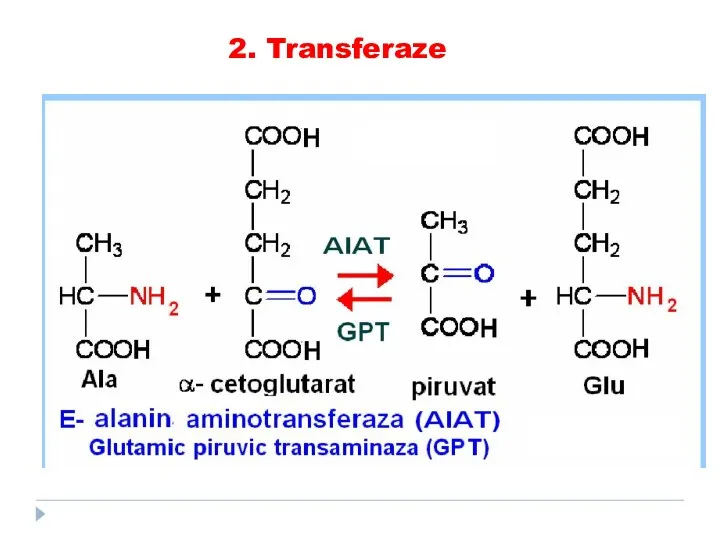
2. Transferaze
2. Transferaze
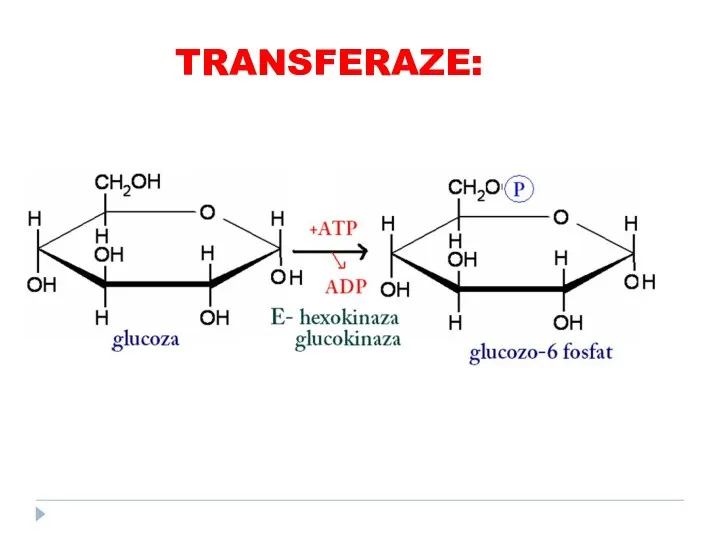
TRANSFERAZE:
TRANSFERAZE:

3. Hidrolaze
catalizează scindări de legături covalente cu adiţionarea apei
SUBCLASELE:
Esteraze
3. Hidrolaze
catalizează scindări de legături covalente cu adiţionarea apei
SUBCLASELE:
Esteraze
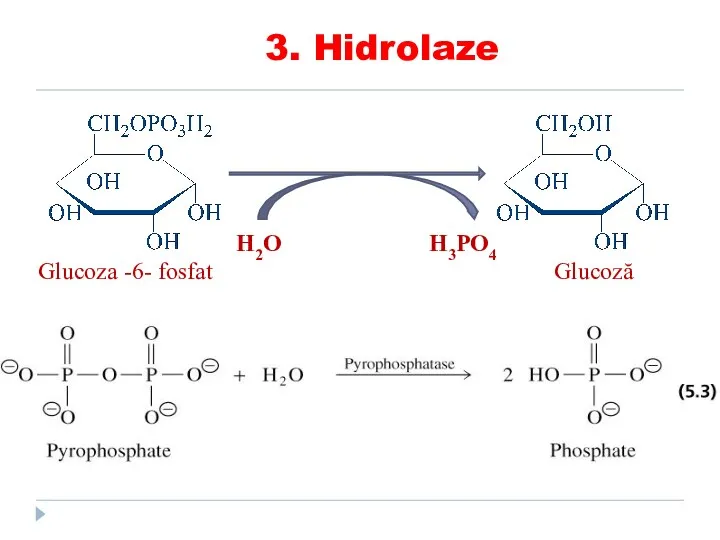
H2O
H3PO4
Glucoza -6- fosfat
Glucoză
3. Hidrolaze
H2O
H3PO4
Glucoza -6- fosfat
Glucoză
3. Hidrolaze
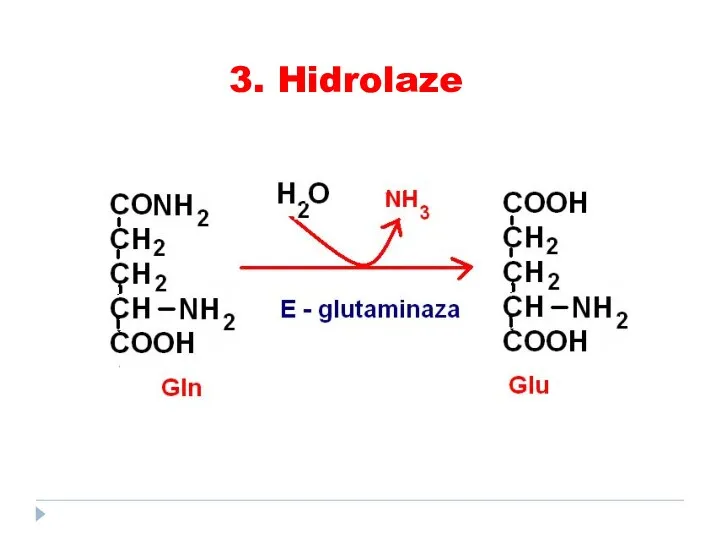
3. Hidrolaze
3. Hidrolaze

4. Liaze
catalizează ruperea leg. C-C, C-S şi C-N; fără adiţionarea
4. Liaze
catalizează ruperea leg. C-C, C-S şi C-N; fără adiţionarea
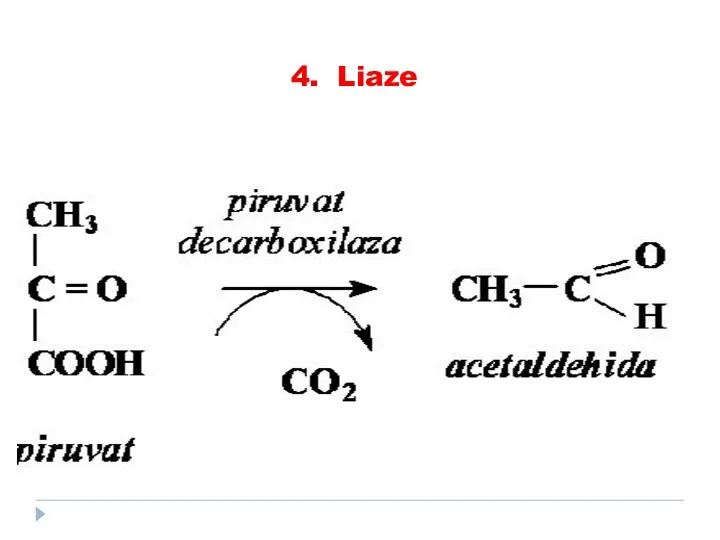
4. Liaze
4. Liaze
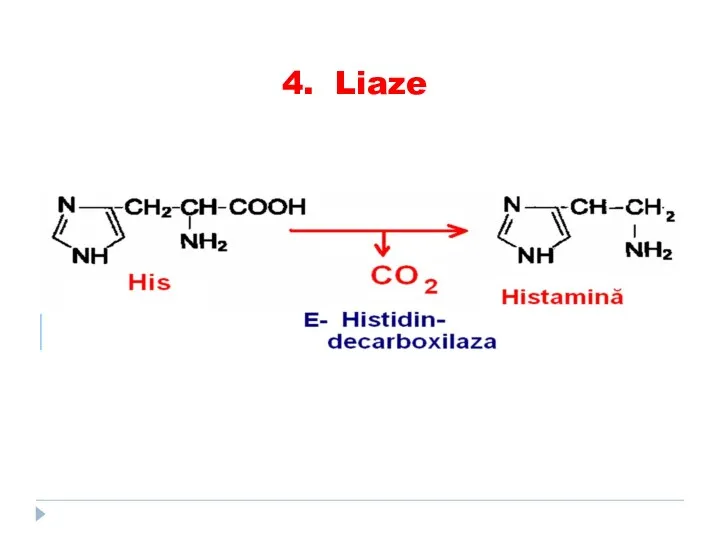
4. Liaze
4. Liaze

4. Liaze
4. Liaze

5. Izomeraze
catalizează toate tipurile de transformări în cadrul uneia şi
5. Izomeraze
catalizează toate tipurile de transformări în cadrul uneia şi
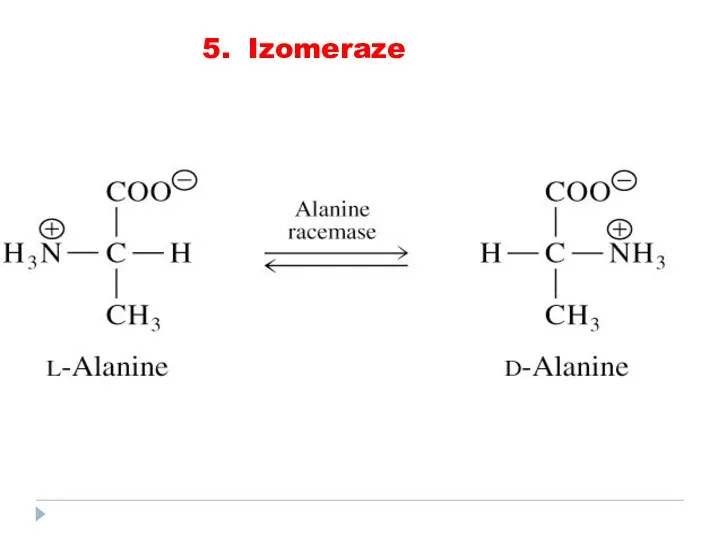
5. Izomeraze
5. Izomeraze
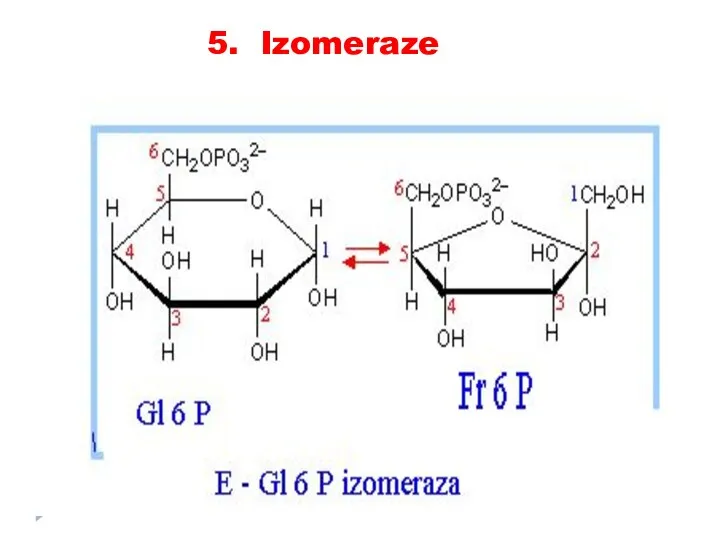
5. Izomeraze
5. Izomeraze
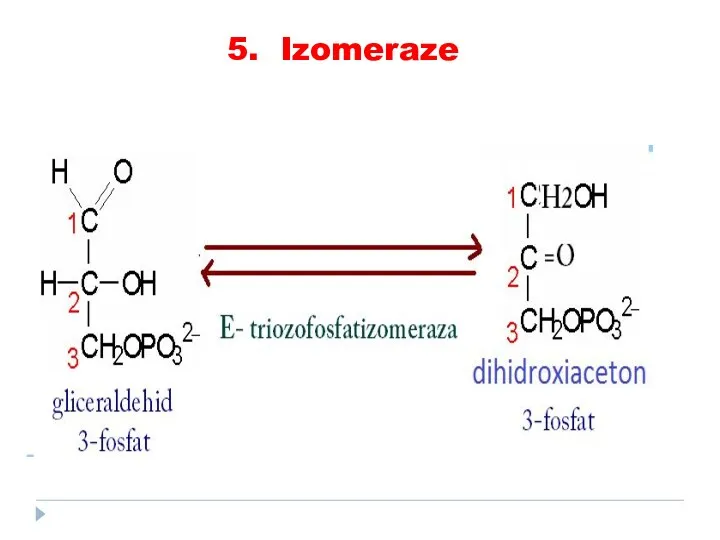
5. Izomeraze
5. Izomeraze

6. Ligaze (sintetaze)
catalizează formarea de legături între carbon şi O, S,
6. Ligaze (sintetaze)
catalizează formarea de legături între carbon şi O, S,
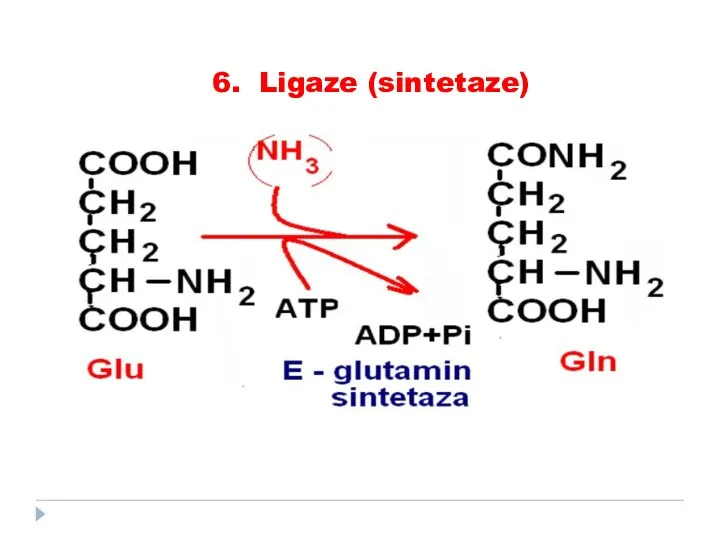
6. Ligaze (sintetaze)
6. Ligaze (sintetaze)
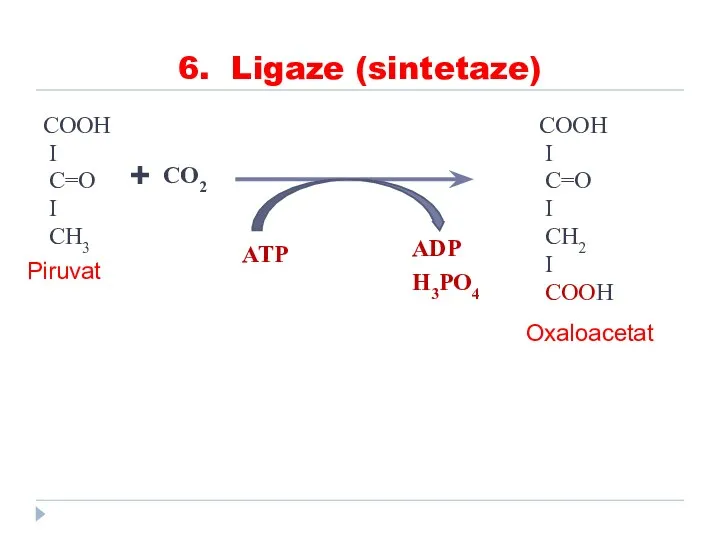
6. Ligaze (sintetaze)
COOH
I
C=O
I
CH3
COOH
I
6. Ligaze (sintetaze)
COOH
I
C=O
I
CH3
COOH
I
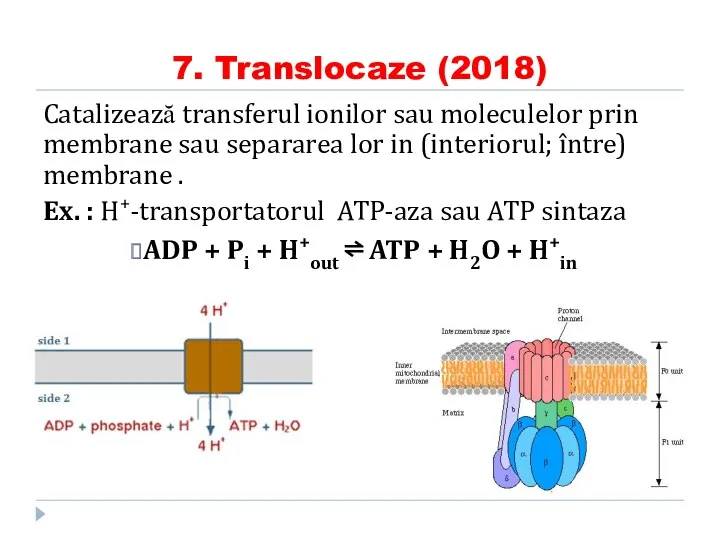
7. Translocaze (2018)
Catalizează transferul ionilor sau moleculelor prin membrane sau separarea
7. Translocaze (2018)
Catalizează transferul ionilor sau moleculelor prin membrane sau separarea
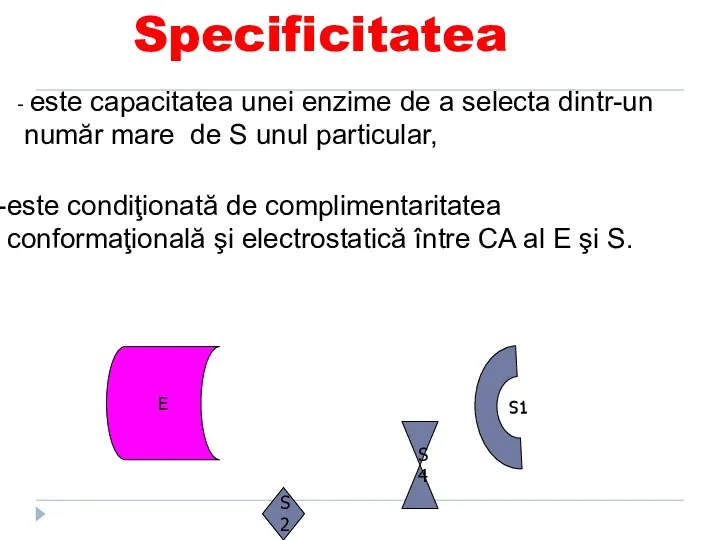
Specificitatea
E
S1
S2
S4
este condiţionată de complimentaritatea conformaţională şi electrostatică între CA al E
Specificitatea
E
S1
S2
S4
este condiţionată de complimentaritatea conformaţională şi electrostatică între CA al E
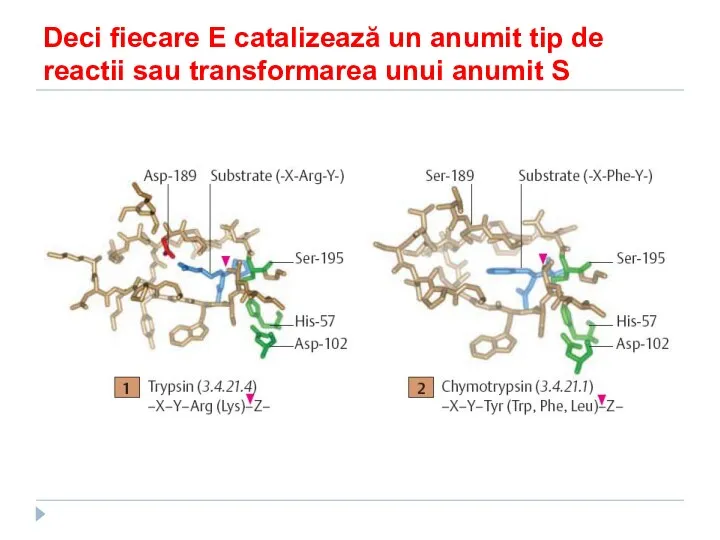
Deci fiecare E catalizează un anumit tip de reactii sau transformarea
Deci fiecare E catalizează un anumit tip de reactii sau transformarea

SPECIFICITATEA
De reacție
De substrat:
A. Absolută
B. Relativă
C. stereospecificitate
SPECIFICITATEA
De reacție
De substrat:
A. Absolută
B. Relativă
C. stereospecificitate

Specificitatea:
Specificitatea de reacţie:
enzimele catalizează un anumit tip de reacţie
Specificitatea:
Specificitatea de reacţie:
enzimele catalizează un anumit tip de reacţie
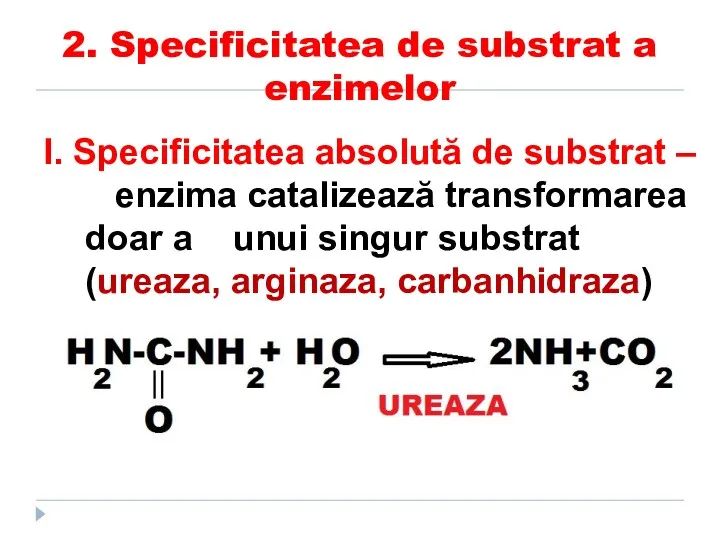
2. Specificitatea de substrat a enzimelor
I. Specificitatea absolută de substrat –
2. Specificitatea de substrat a enzimelor
I. Specificitatea absolută de substrat –

Specificitatea de substrat a enzimelor
II. Specificitatea relativă de substrat –
enzima catalizează
Specificitatea de substrat a enzimelor
II. Specificitatea relativă de substrat –
enzima catalizează

Specificitatea de substrat a enzimelor
specificitate relativa de substrat - asigura transformarea unui
Specificitatea de substrat a enzimelor
specificitate relativa de substrat - asigura transformarea unui
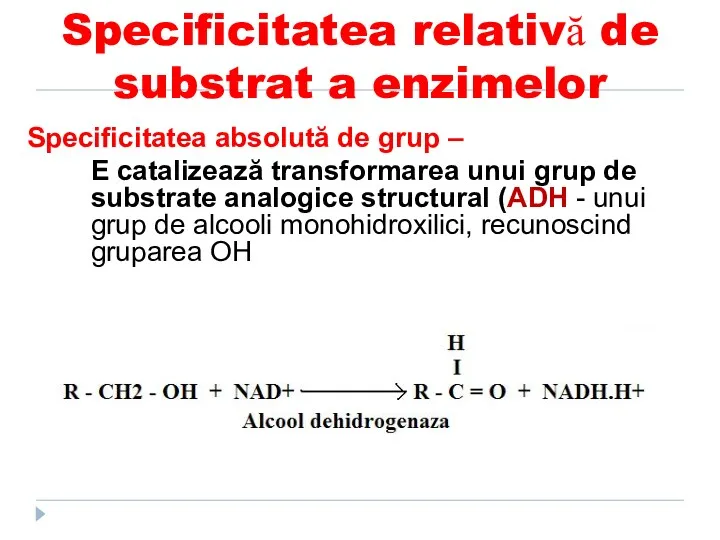
Specificitatea relativă de substrat a enzimelor
Specificitatea absolută de grup –
E catalizează
Specificitatea relativă de substrat a enzimelor
Specificitatea absolută de grup –
E catalizează

Specificitatea relativă de substrat a enzimelor
Specificitatea relativă de grup –
enzima
Specificitatea relativă de substrat a enzimelor
Specificitatea relativă de grup –
enzima

Specificitatea relativă de grupă
Specificitatea relativă de grupă

Specificitate stereochimică
E catalizează transformarea numai a unuia din stereoizomerii posibili (D
Specificitate stereochimică
E catalizează transformarea numai a unuia din stereoizomerii posibili (D

Specificitate stereochimică
Hexokinaza – specificitate la D- glucoză
Oxidazele – specificitate la
Specificitate stereochimică
Hexokinaza – specificitate la D- glucoză
Oxidazele – specificitate la

MECANISMUL DE ACȚIUNE AL ENZIMELOR

Mecanismul de acţiune al E
Pentru decurgerea unei reacţii este necesar ca
Mecanismul de acţiune al E
Pentru decurgerea unei reacţii este necesar ca

Mecanismul de acţiune al E
E - micşorează energia de activare ale
Mecanismul de acţiune al E
E - micşorează energia de activare ale

S + E ↔ E-S → E + P
Enzimele reduc energia
S + E ↔ E-S → E + P
Enzimele reduc energia
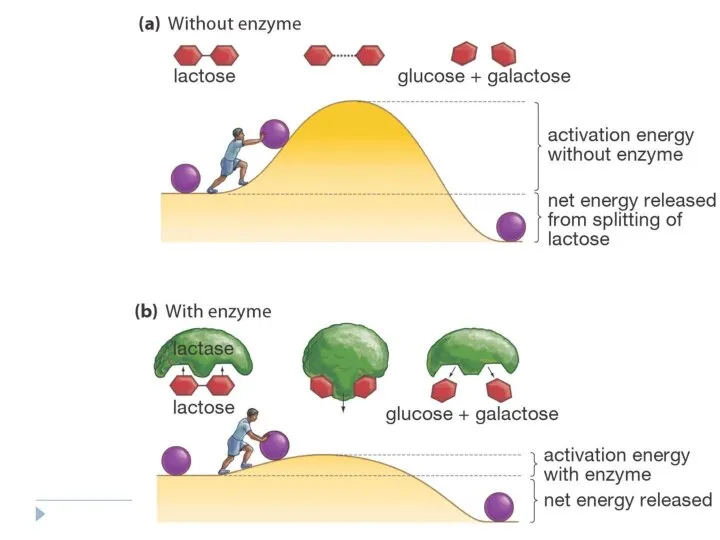
Enzymes Lower a Reaction’s Activation Energy
Enzymes Lower a Reaction’s Activation Energy
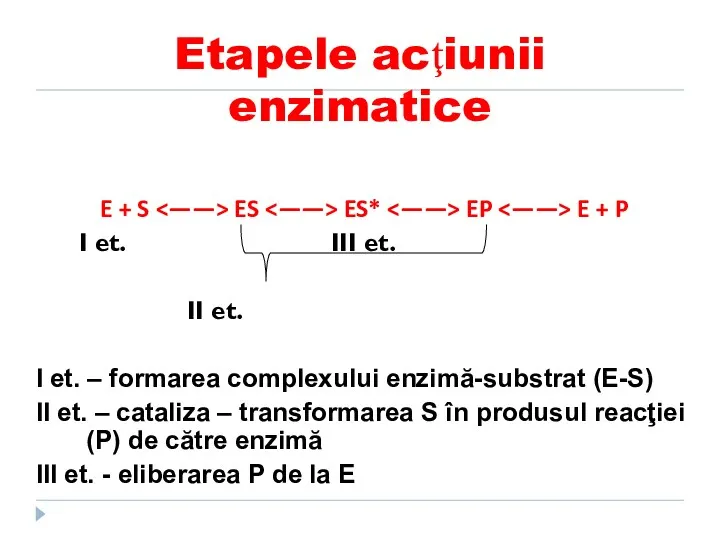
Etapele acţiunii enzimatice
E + S <——> ES <——> ES* <——> EP
Etapele acţiunii enzimatice
E + S <——> ES <——> ES* <——> EP

Mecanismul de acţiune al enzimelor
Prima etapă:
Difuzia S spre E şi
Mecanismul de acţiune al enzimelor
Prima etapă:
Difuzia S spre E şi
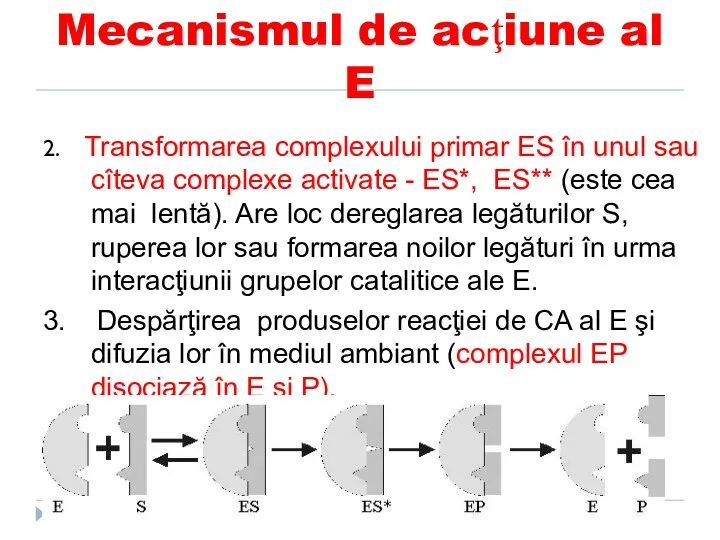
Mecanismul de acţiune al E
2. Transformarea complexului primar ES în unul
Mecanismul de acţiune al E
2. Transformarea complexului primar ES în unul

Ipoteza “lacăt-cheie” (Fischer) şi “coincidenţa forţată” (Koshland)
Modelul clasic (Emil Fischer) consideră
Ipoteza “lacăt-cheie” (Fischer) şi “coincidenţa forţată” (Koshland)
Modelul clasic (Emil Fischer) consideră

Teoria “cheie-lacăt” - Fisher
Teoria “cheie-lacăt” - Fisher
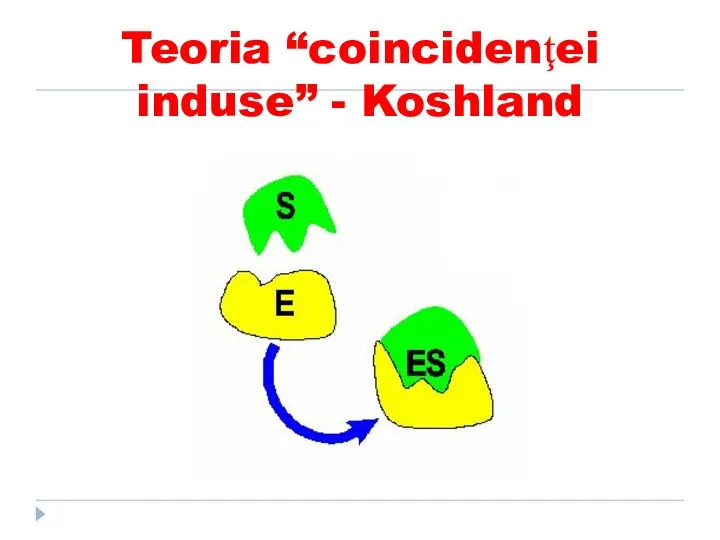
Teoria “coincidenţei induse” - Koshland
Teoria “coincidenţei induse” - Koshland
S
P1
P2
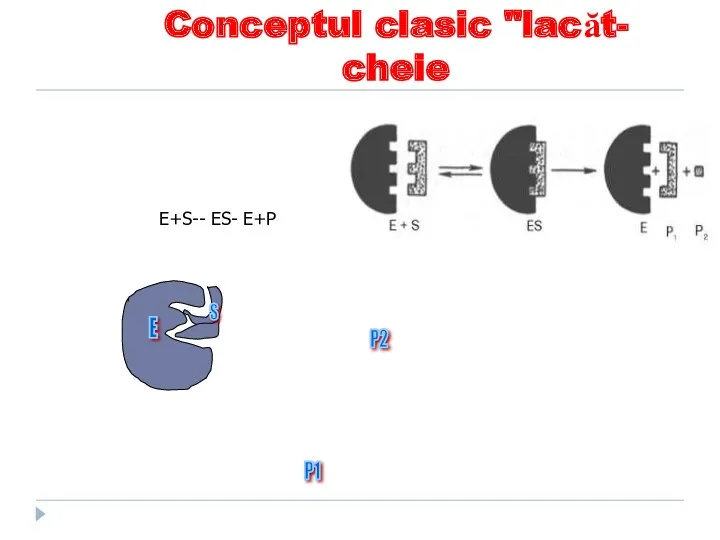
Conceptul clasic "lacăt- cheie
E+S-- ES- E+P
S
P1
P2
Conceptul clasic "lacăt- cheie
E+S-- ES- E+P

S
P1
P2
Conceptul coencidenţei inductive
“coincidenţa forţată” (Kochland)
S
P1
P2
Conceptul coencidenţei inductive
“coincidenţa forţată” (Kochland)

Mecanismul de acţiune al E
La nivel molecular acţiunea E poate fi
Mecanismul de acţiune al E
La nivel molecular acţiunea E poate fi

PROPRIETĂŢILE ENZIMELOR
PROPRIETĂŢILE ENZIMELOR

Obiectivele:
Cinetica enzimatică. Influenţa concentraţiei enzimei şi a substratului, a pH-ului şi
Obiectivele:
Cinetica enzimatică. Influenţa concentraţiei enzimei şi a substratului, a pH-ului şi

CINETICA ENZIMATICĂ
Studiază viteza reacției enzimatice, luând în considerație factorii fizico-chimici ce
CINETICA ENZIMATICĂ
Studiază viteza reacției enzimatice, luând în considerație factorii fizico-chimici ce

VITEZA REACȚIEI ENZIMATICE
reprezintă numărul moleculelor de substrat transformate în produs final
VITEZA REACȚIEI ENZIMATICE
reprezintă numărul moleculelor de substrat transformate în produs final

Factorii care influenţiază viteza reacției enzimatice
Concentraţia S
Concentraţia E
Temperatura
pH
Factorii care influenţiază viteza reacției enzimatice
Concentraţia S
Concentraţia E
Temperatura
pH
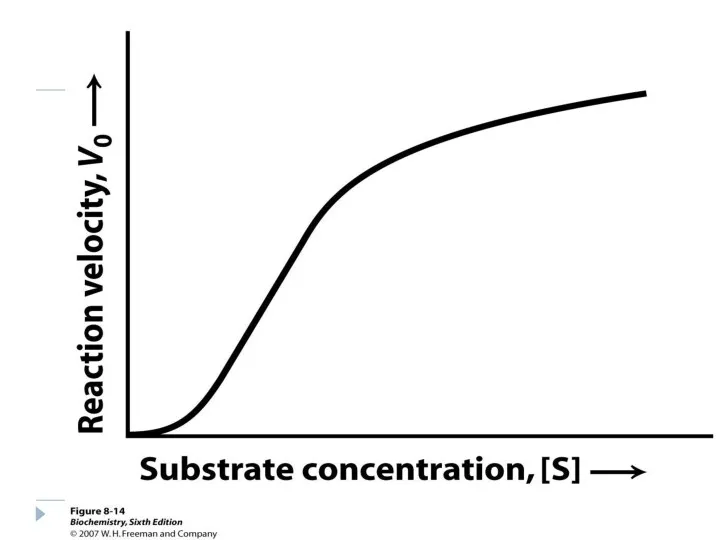
![Influența [S] asupta vitezei reacției enzimatice Grafic se reprezintă sub](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/378235/slide-119.jpg)
Influența [S] asupta vitezei reacției enzimatice
Grafic se reprezintă sub formă
Influența [S] asupta vitezei reacției enzimatice
Grafic se reprezintă sub formă
![Influența [S] asupta vitezei reacției enzimatice Analiza curbei arată 3](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/378235/slide-120.jpg)
Influența [S] asupta vitezei reacției enzimatice
Analiza curbei arată 3 zone:
Zona “a”-
Influența [S] asupta vitezei reacției enzimatice
Analiza curbei arată 3 zone:
Zona “a”-

Această curbă este numită curba lui Michaelis-Menten şi se exprimă prin
Această curbă este numită curba lui Michaelis-Menten şi se exprimă prin


Semnificația lui Km și
V max
Km-constanta lui Michaelis Menten
- este
Semnificația lui Km și
V max
Km-constanta lui Michaelis Menten - este
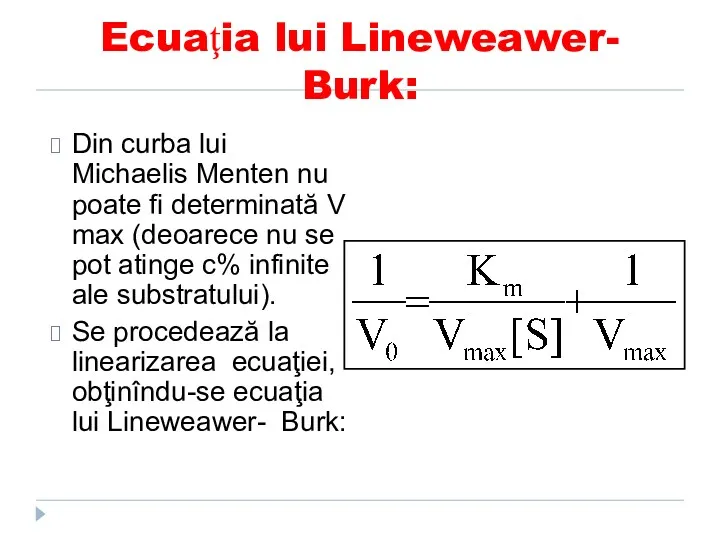
Ecuaţia lui Lineweawer- Burk:
Din curba lui Michaelis Menten nu poate fi
Ecuaţia lui Lineweawer- Burk:
Din curba lui Michaelis Menten nu poate fi

Diagrama lui Lineweawer- Burk:
Reciproca ecuatiei
Michaelis-Menten
Diagrama lui Lineweawer- Burk:
Reciproca ecuatiei
Michaelis-Menten
![Influența [E] asupta vitezei reacției în condiţii standard 2 mol](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/378235/slide-126.jpg)
Influența [E] asupta vitezei reacției
în condiţii standard 2 mol de
Influența [E] asupta vitezei reacției
în condiţii standard 2 mol de
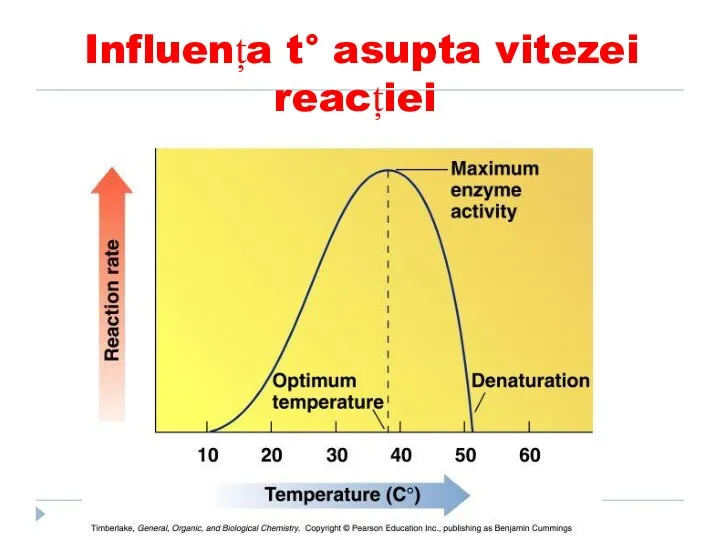
Influența t° asupta vitezei reacției
Influența t° asupta vitezei reacției

Termolabilitatea (t°)
Unele E a microorganismelor termofile sunt active la t de
Termolabilitatea (t°)
Unele E a microorganismelor termofile sunt active la t de
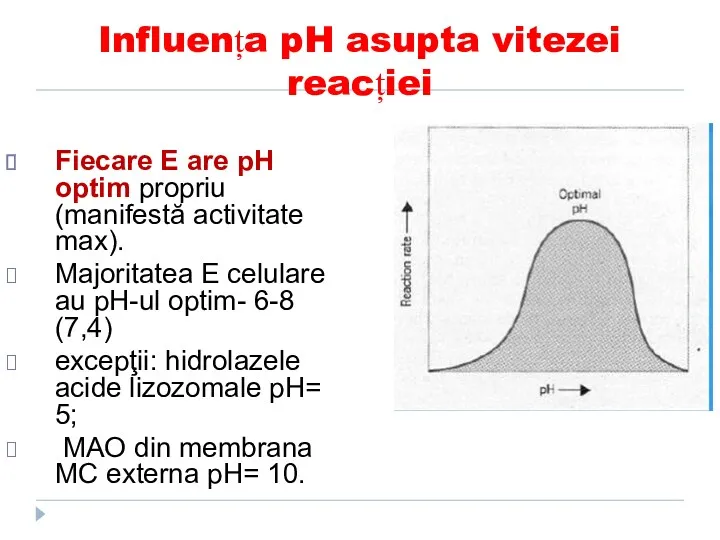
Influența pH asupta vitezei reacției
Fiecare E are pH optim propriu (manifestă
Influența pH asupta vitezei reacției
Fiecare E are pH optim propriu (manifestă
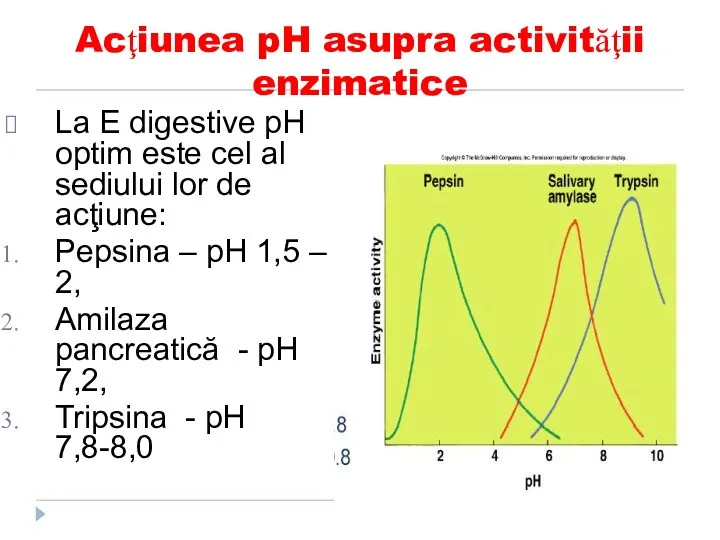
Acţiunea pH asupra activităţii enzimatice
La E digestive pH optim este cel
Acţiunea pH asupra activităţii enzimatice
La E digestive pH optim este cel
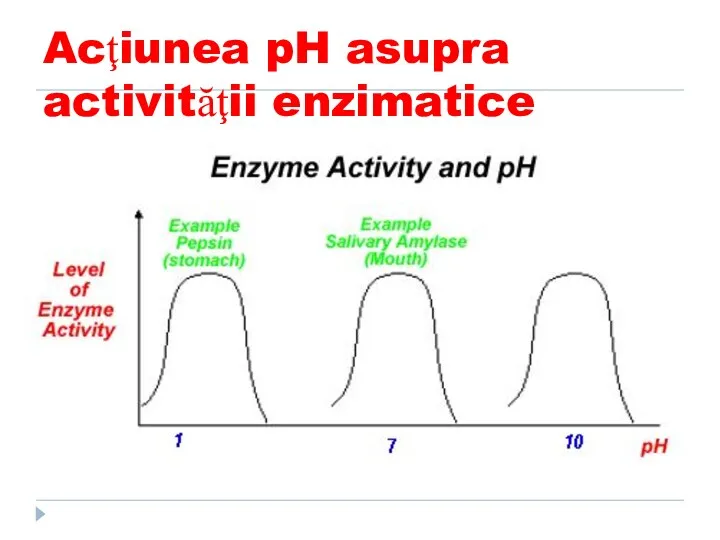
Acţiunea pH asupra activităţii enzimatice
Acţiunea pH asupra activităţii enzimatice
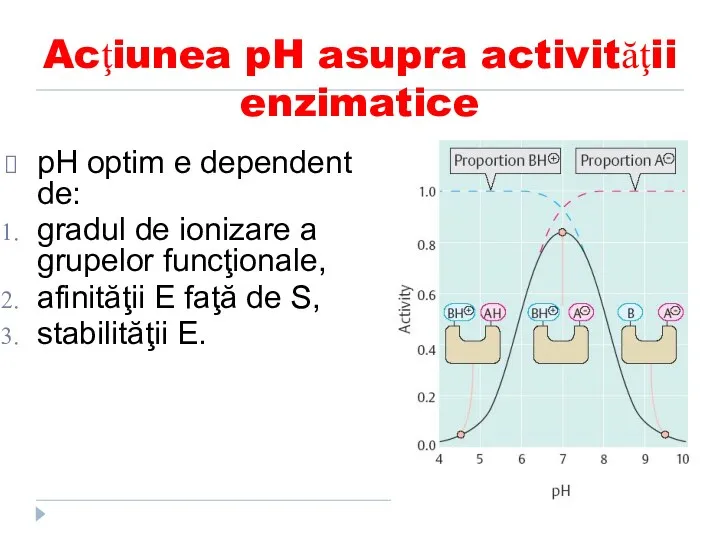
Acţiunea pH asupra activităţii enzimatice
pH optim e dependent de:
gradul de
Acţiunea pH asupra activităţii enzimatice
pH optim e dependent de:
gradul de

Acţiunea pH asupra activităţii enzimatice
Centrul activ al enzimelor contine grupe ionizabile,
Acţiunea pH asupra activităţii enzimatice
Centrul activ al enzimelor contine grupe ionizabile,

Influenta efectorilor enzimatici
Efectorii enzimatici sunt substante cu structuri chimice variate care
Influenta efectorilor enzimatici
Efectorii enzimatici sunt substante cu structuri chimice variate care

Inhibitorii enzimatici
Inhibitorii enzimatici sunt compusi care influenteaza negativ activitatea enzimelor pe
Inhibitorii enzimatici
Inhibitorii enzimatici sunt compusi care influenteaza negativ activitatea enzimelor pe

Inhibiţia enzimatică
Deosebim:
inhibiţie nespecifică (T, pH, agenţii denaturării )
inhibiţie specifică
Inhibiţia
Inhibiţia enzimatică
Deosebim:
inhibiţie nespecifică (T, pH, agenţii denaturării )
inhibiţie specifică
Inhibiţia

EXEMPLE DE INHIBITORI IREVERSIBILI
Compușii organofosforici (Diizopropilfluorfosfatul) (toxina neuroparalitica) se fixează de
EXEMPLE DE INHIBITORI IREVERSIBILI
Compușii organofosforici (Diizopropilfluorfosfatul) (toxina neuroparalitica) se fixează de

Inhibiţie ireversibilă
Inhibiţie ireversibilă
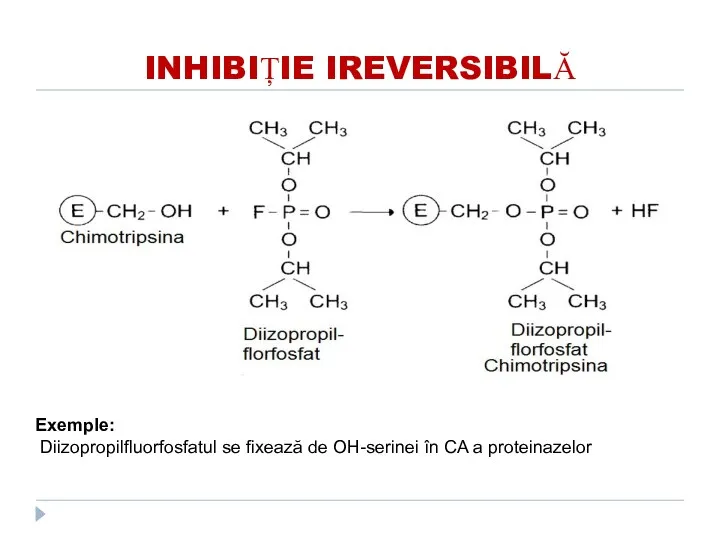
INHIBIȚIE IREVERSIBILĂ
Exemple:
Diizopropilfluorfosfatul se fixează de OH-serinei în CA a proteinazelor
INHIBIȚIE IREVERSIBILĂ
Exemple:
Diizopropilfluorfosfatul se fixează de OH-serinei în CA a proteinazelor
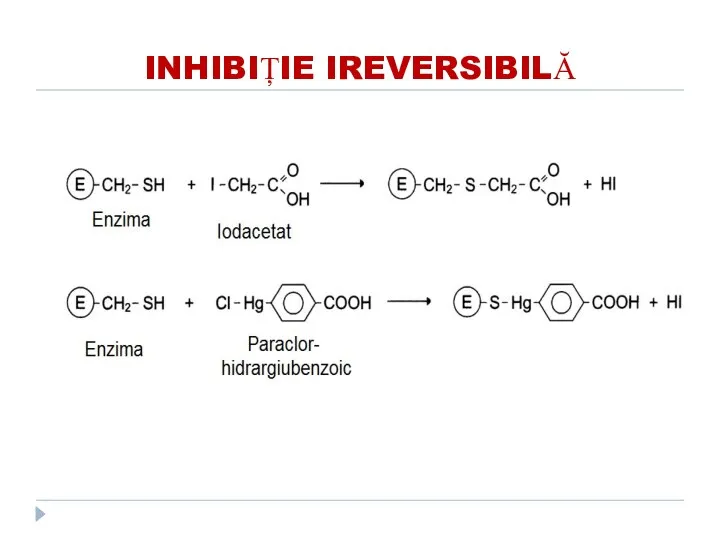
INHIBIȚIE IREVERSIBILĂ
INHIBIȚIE IREVERSIBILĂ

Tipuri de inhibitie revesibilă
Deosebim:
Inhibiţie competitivă
Inhibiţie necompetitivă (noncompetitivă)
Inhibiţie uncompetetivă
Inhibiţie alosterică
Tipuri de inhibitie revesibilă
Deosebim:
Inhibiţie competitivă
Inhibiţie necompetitivă (noncompetitivă)
Inhibiţie uncompetetivă
Inhibiţie alosterică

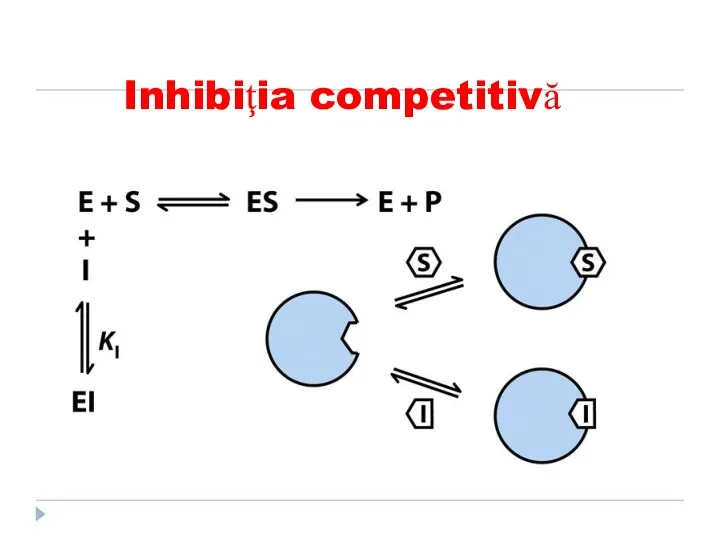
Inhibiţia competitivă
I se aseamănă după structură cu S.
Apare o competiţie
Inhibiţia competitivă
I se aseamănă după structură cu S.
Apare o competiţie
Inhibiţia competitivă
Inhibiţia competitivă
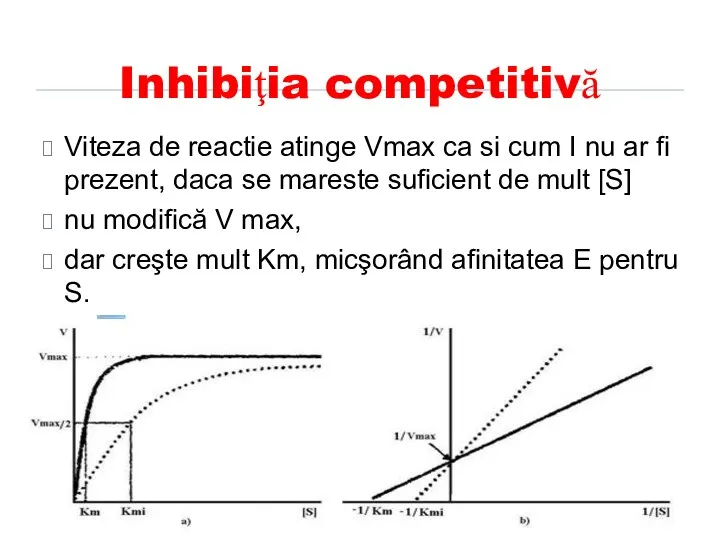
Inhibiţia competitivă
Viteza de reactie atinge Vmax ca si cum I nu
Inhibiţia competitivă
Viteza de reactie atinge Vmax ca si cum I nu

Exemple de I competitivi:
inhibiţia SDH cu malonat (SDH -oxideaza succinatul in
Exemple de I competitivi:
inhibiţia SDH cu malonat (SDH -oxideaza succinatul in
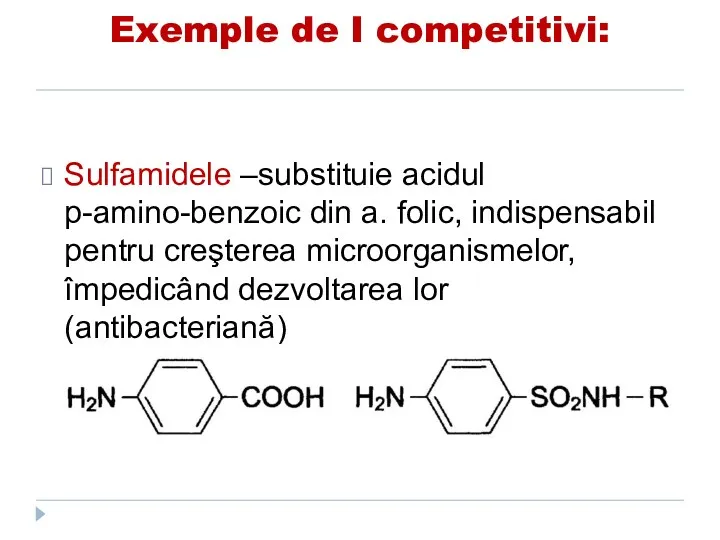
Exemple de I competitivi:
Sulfamidele –substituie acidul p-amino-benzoic din a. folic, indispensabil
Exemple de I competitivi:
Sulfamidele –substituie acidul p-amino-benzoic din a. folic, indispensabil
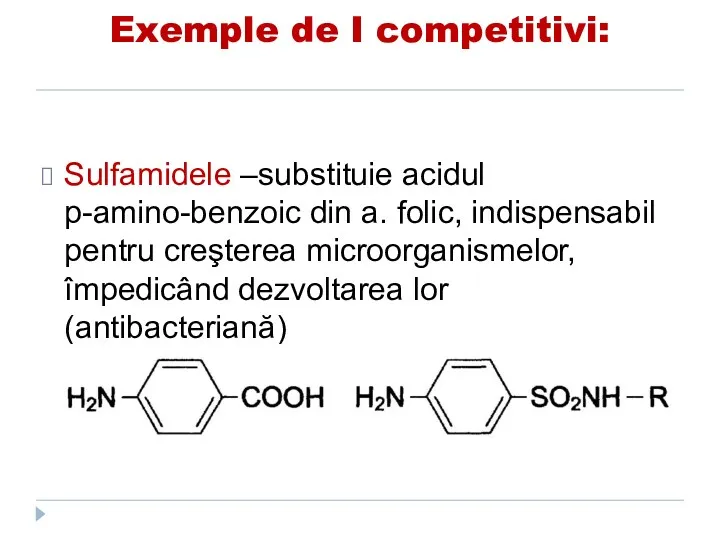
Exemple de I competitivi:
Sulfamidele –substituie acidul p-amino-benzoic din a. folic, indispensabil
Exemple de I competitivi:
Sulfamidele –substituie acidul p-amino-benzoic din a. folic, indispensabil
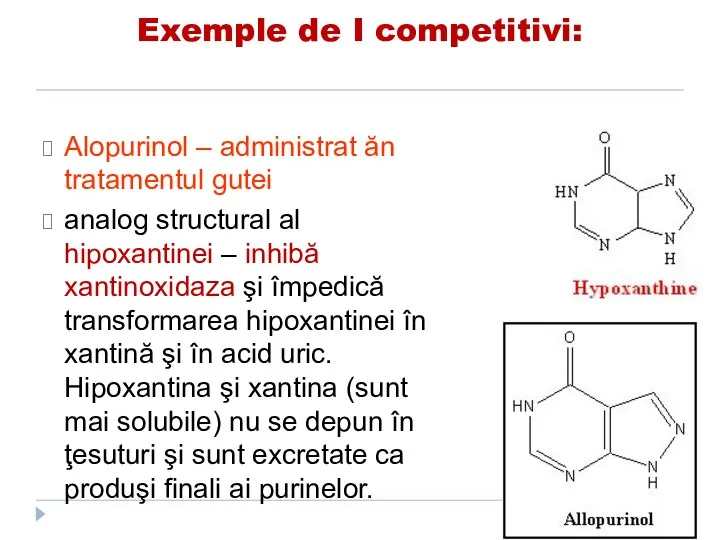
Exemple de I competitivi:
Alopurinol – administrat ăn tratamentul gutei
analog structural al
Exemple de I competitivi:
Alopurinol – administrat ăn tratamentul gutei
analog structural al
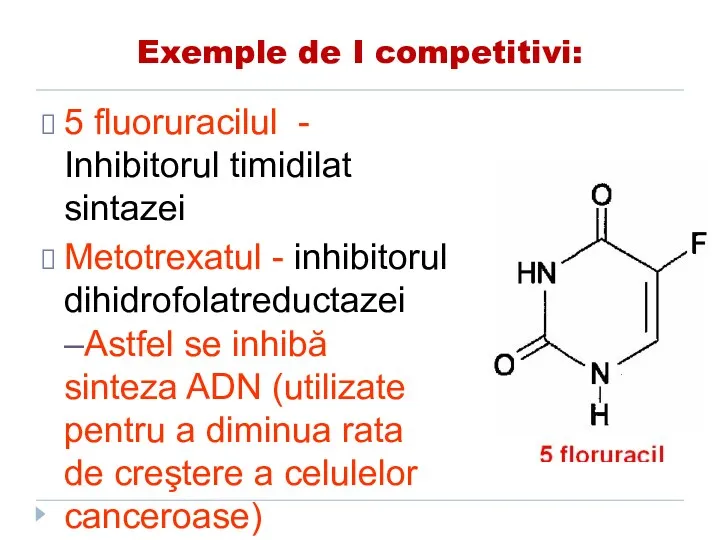
Exemple de I competitivi:
5 fluoruracilul - Inhibitorul timidilat sintazei
Metotrexatul -
Exemple de I competitivi:
5 fluoruracilul - Inhibitorul timidilat sintazei
Metotrexatul -

Inhibiţia necompetitivă
Inhibitorul nu se aseamănă ca structură cu S
I şi S
Inhibiţia necompetitivă
Inhibitorul nu se aseamănă ca structură cu S
I şi S
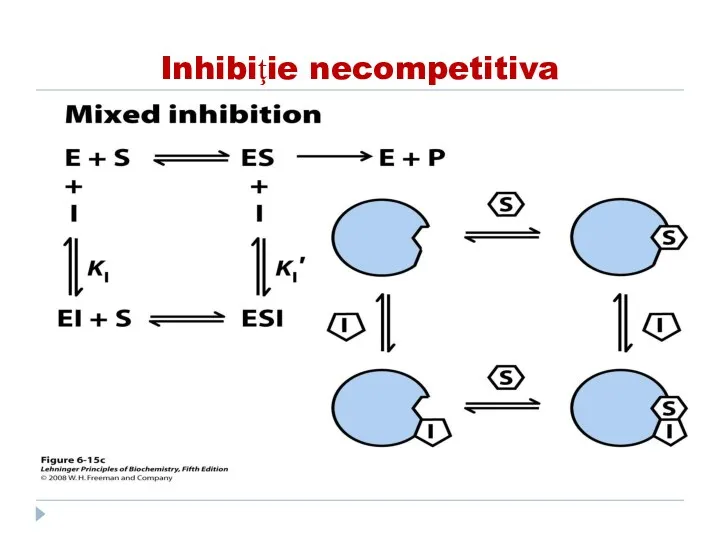
Inhibiţie necompetitiva
Inhibiţie necompetitiva

Inhibiţie necompetitiva
In cazul inhibitiei necompetitive, I chiar daca nu se leaga
Inhibiţie necompetitiva
In cazul inhibitiei necompetitive, I chiar daca nu se leaga
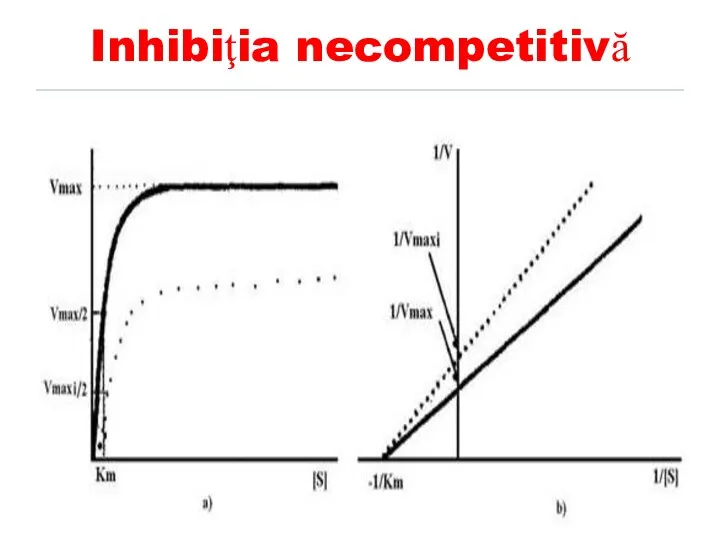
Inhibiţia necompetitivă
Inhibiţia necompetitivă

Tipuri de inhibitori necompetitivi:
cianurile, CO se fixează cu Fe 3+ din
Tipuri de inhibitori necompetitivi:
cianurile, CO se fixează cu Fe 3+ din

Inhibiţia necompetitivă
I poate fi înlăturat de substanţe care îl leagă –
Inhibiţia necompetitivă
I poate fi înlăturat de substanţe care îl leagă –
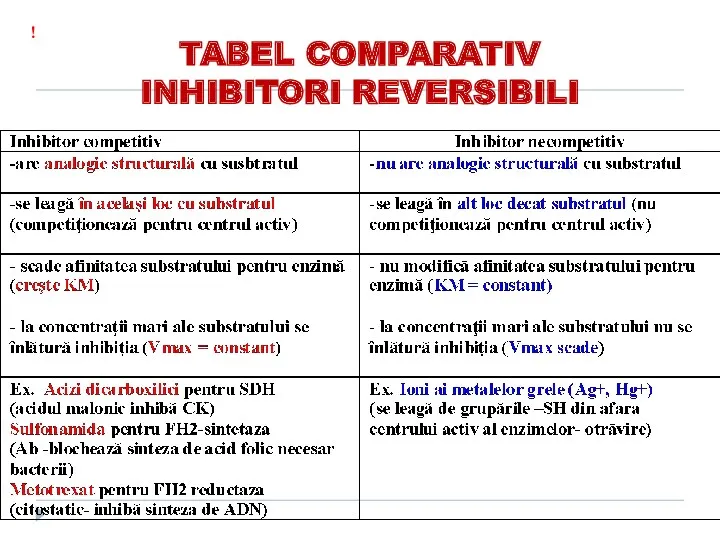
TABEL COMPARATIV
INHIBITORI REVERSIBILI
!
TABEL COMPARATIV
INHIBITORI REVERSIBILI
!
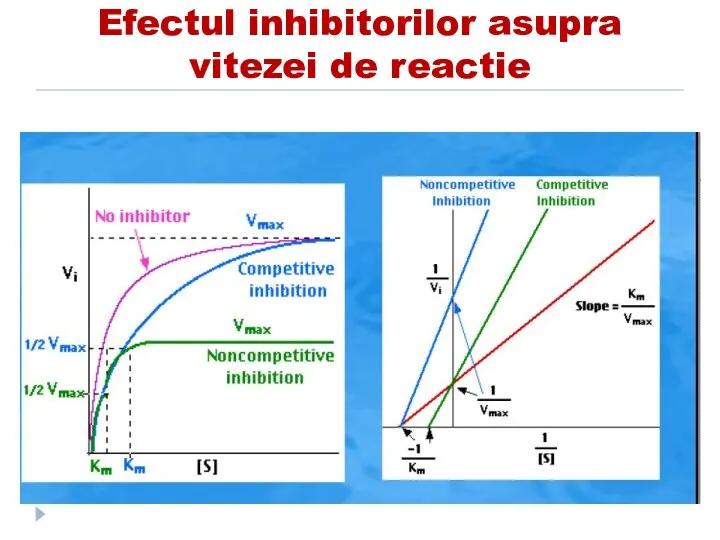
Efectul inhibitorilor asupra vitezei de reactie
Efectul inhibitorilor asupra vitezei de reactie

Inhibiţia uncompetetivă –
E+S----ES +I-----ESI
I se combina cu complexul ES formand
Inhibiţia uncompetetivă –
E+S----ES +I-----ESI
I se combina cu complexul ES formand
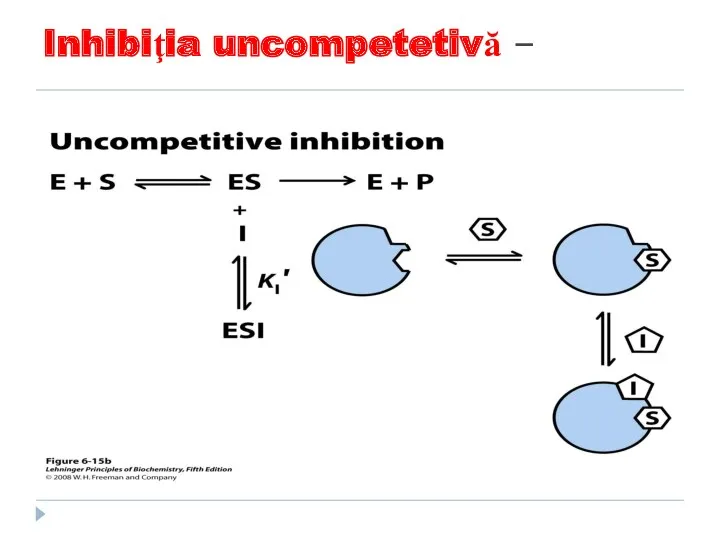
Inhibiţia uncompetetivă –
Inhibiţia uncompetetivă –
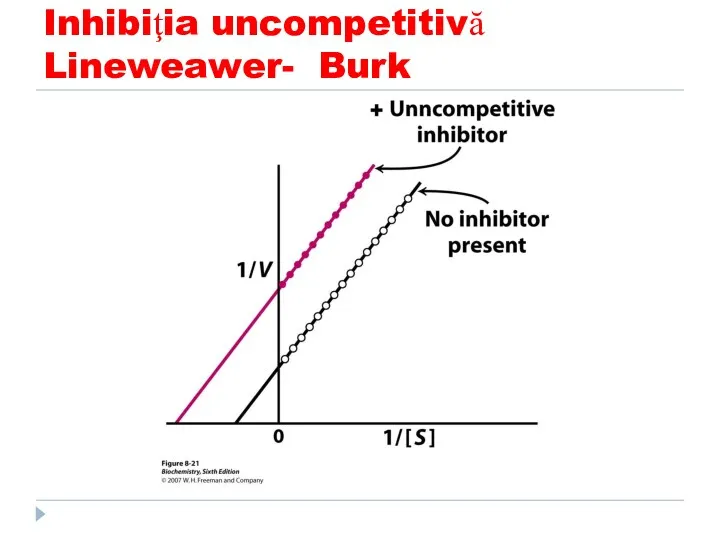
Inhibiţia uncompetitivă
Lineweawer- Burk
Inhibiţia uncompetitivă
Lineweawer- Burk

Alte tipuri de inhibiție
inhibiţia prin modificarea covalentă a moleculei E -
Alte tipuri de inhibiție
inhibiţia prin modificarea covalentă a moleculei E -

REGLAREA ACTIVITATII ENZIMELOR
REGLAREA ACTIVITATII ENZIMELOR

Mecanismele de activare a E
Sunt : 1. nespecifice: temperatura , iradierea
2.
Mecanismele de activare a E
Sunt : 1. nespecifice: temperatura , iradierea
2.

I. REGLAREA CANTITATIVA
Functie de dinamica SINTEZA/DEGRADARE
Enzime inductibile Ks > Kd
Enzime
I. REGLAREA CANTITATIVA
Functie de dinamica SINTEZA/DEGRADARE
Enzime inductibile Ks > Kd
Enzime

II. REGLAREA CALITATIVA
Mecanismele de activare a E
Deosebim următoarele tipuri de reglare
II. REGLAREA CALITATIVA
Mecanismele de activare a E
Deosebim următoarele tipuri de reglare

II. REGLAREA CALITATIVA
II.1.Reglarea alosterica (necovalenta)
II. REGLAREA CALITATIVA
II.1.Reglarea alosterica (necovalenta)

Enzime alosterice. Efectori alosterici
Sunt proteine oligomere alcatuite din mai multe subunitati
Enzime alosterice. Efectori alosterici
Sunt proteine oligomere alcatuite din mai multe subunitati

Enzime alosterice. Efectori alosterici
Pe langa centri activi, monomerii prezinta si centri
Enzime alosterice. Efectori alosterici
Pe langa centri activi, monomerii prezinta si centri

II. REGLAREA CALITATIVA
II.1.Reglarea alosterica de tip heterotrop
II. REGLAREA CALITATIVA
II.1.Reglarea alosterica de tip heterotrop
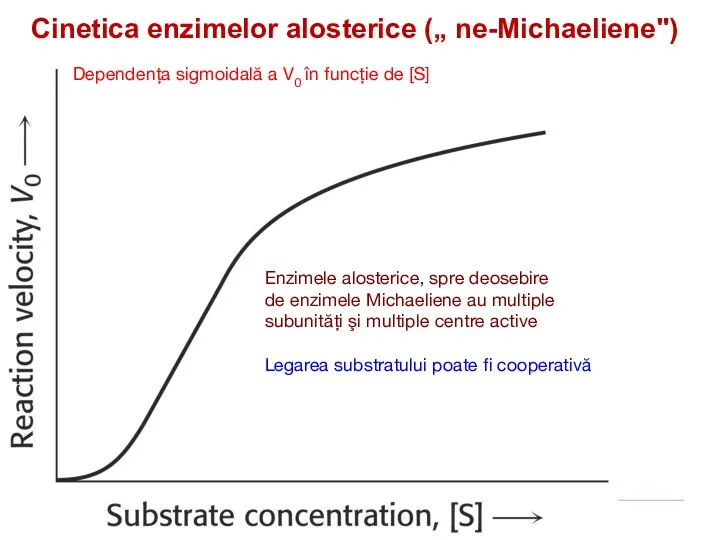
Cinetica enzimelor alosterice („ ne-Michaeliene")
Dependenţa sigmoidală a V0 în funcţie de
Cinetica enzimelor alosterice („ ne-Michaeliene")
Dependenţa sigmoidală a V0 în funcţie de
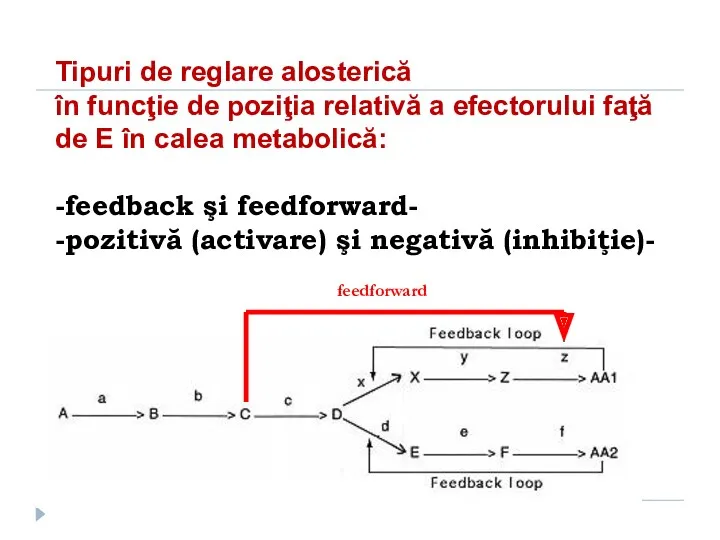
Tipuri de reglare alosterică
în funcţie de poziţia relativă a efectorului
Tipuri de reglare alosterică în funcţie de poziţia relativă a efectorului
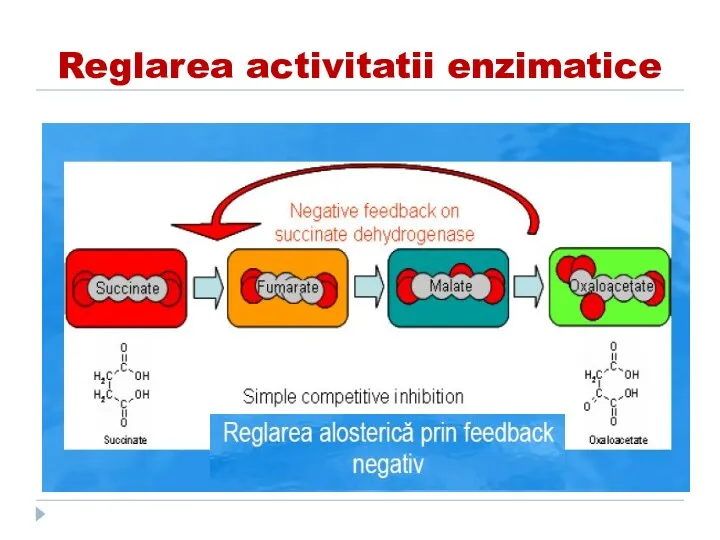
Reglarea activitatii enzimatice
Reglarea activitatii enzimatice
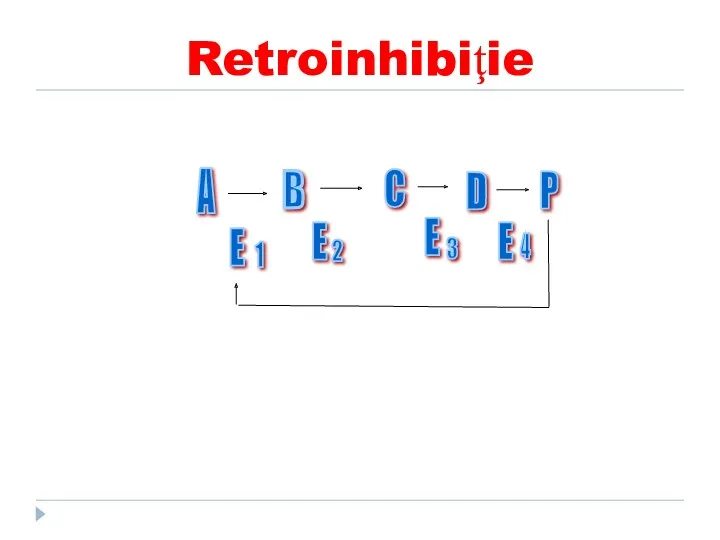
Retroinhibiţie
A
B
C
D
P
E
E
1
E
2
3
Retroinhibiţie
A
B
C
D
P
E
E
1
E
2
3

Reglarea activitatii enzimatice
Reglarea activitatii enzimatice

Reglarea covalentă - Proteoliză limitată
Unele enzime (proteine) se sintetizează în forma
Reglarea covalentă - Proteoliză limitată
Unele enzime (proteine) se sintetizează în forma

Proteoliză limitată
-este scindarea unui sector al catenei în rezultatul căreia E
Proteoliză limitată
-este scindarea unui sector al catenei în rezultatul căreia E

Importanța biologică a prezenţei formelor neactive
Zimogenii sunt produse la locurile
Importanța biologică a prezenţei formelor neactive
Zimogenii sunt produse la locurile

Reglarea covalentă - fosforilare-defosforilare
unele E sunt active în forma fosforilată, iar
Reglarea covalentă - fosforilare-defosforilare
unele E sunt active în forma fosforilată, iar
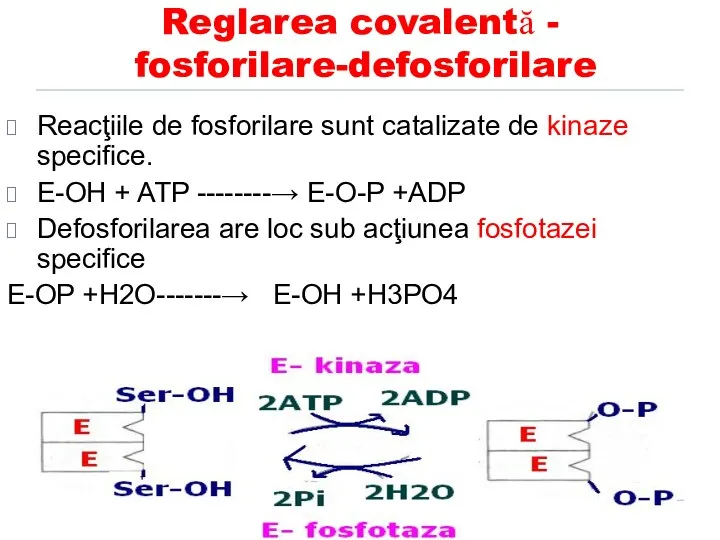
Reglarea covalentă -
fosforilare-defosforilare
Reacţiile de fosforilare sunt catalizate de kinaze specifice.
Reglarea covalentă -
fosforilare-defosforilare
Reacţiile de fosforilare sunt catalizate de kinaze specifice.

Autostructurarea cuaternară
Este caracteristică E ce posedă structură cuaternară
Fiecare protomer în parte
Autostructurarea cuaternară
Este caracteristică E ce posedă structură cuaternară
Fiecare protomer în parte

E
CA
Allos
S
M
CA
Reglarea alosterică
Modulator pozitiv
E
CA
Allos
S
M
CA
Reglarea alosterică
Modulator pozitiv

Izoenzimele- izoE
forme moleculare multiple ale E, ce catalizează aceeaşi reacţie chimică,
Izoenzimele- izoE
forme moleculare multiple ale E, ce catalizează aceeaşi reacţie chimică,

IzoE diferă între ele prin:
sarcina electrică (ce permite separarea lor prin
IzoE diferă între ele prin:
sarcina electrică (ce permite separarea lor prin

STRUCTURA IZOE
Sunt E oligomere, cu structură cuaternară, alcătuite din cel puţin
STRUCTURA IZOE
Sunt E oligomere, cu structură cuaternară, alcătuite din cel puţin


Izoformele lactat dehidrogenazei (LDH)
LDH-1 (4H) - in inima
LDH-2 (3H1M) -
Izoformele lactat dehidrogenazei (LDH)
LDH-1 (4H) - in inima
LDH-2 (3H1M) -

ROLUL izoenzimelor
în controlul metabolic ( faciliteaza adaptarea metabolismului in diferite ţesuturi.)
ROLUL izoenzimelor
în controlul metabolic ( faciliteaza adaptarea metabolismului in diferite ţesuturi.)

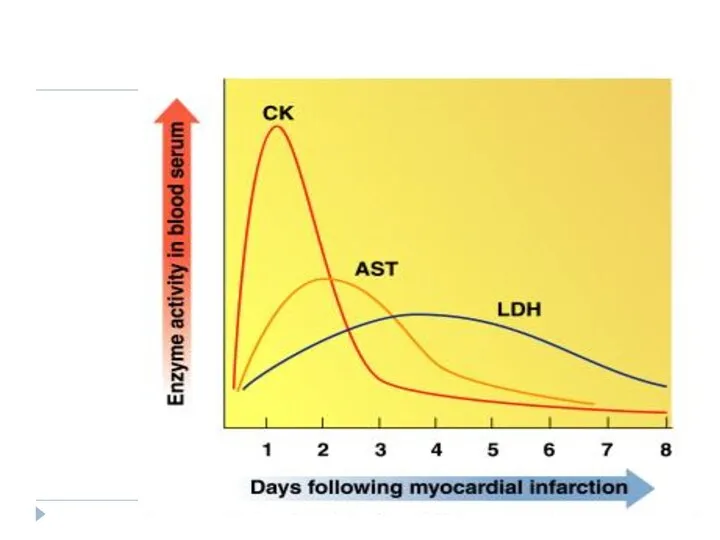
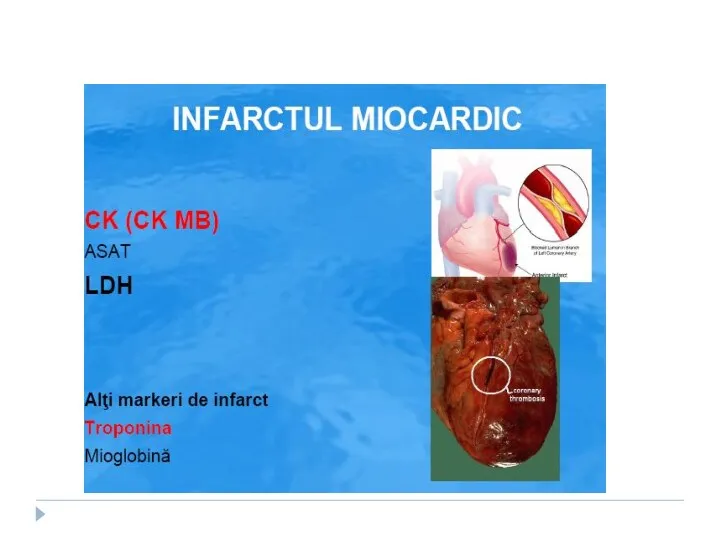
LDH
Norma- 100-190U/L
Creşterea de LDH 1 şi LDH2:
infarctul miocardic
anemia hemolitică/
LDH
Norma- 100-190U/L
Creşterea de LDH 1 şi LDH2:
infarctul miocardic
anemia hemolitică/

Exemple de izoenzime:
Creatinfosfokinaza -2 tipuri de monomeri: M-Muscle şi B –brain
LDH
MDH
Aldolaza
Fosfataza
Exemple de izoenzime:
Creatinfosfokinaza -2 tipuri de monomeri: M-Muscle şi B –brain
LDH
MDH
Aldolaza
Fosfataza

Creatinfosfokinaza (CPK)
Creatinfosfokinaza (CPK)


Sistemele polienzimatice
Fiecare celulă a organismului conţine setul său specific de E.
Sistemele polienzimatice
Fiecare celulă a organismului conţine setul său specific de E.

Tipurile de organizare a sistemelor polienzimatice
Se cunosc urmatoarele tipuri de organizare
Tipurile de organizare a sistemelor polienzimatice
Se cunosc urmatoarele tipuri de organizare

Organizarea funcţională
enzimele sunt asociate în sistemul polienzimatic cu ajutorul metaboliţilor,
Organizarea funcţională
enzimele sunt asociate în sistemul polienzimatic cu ajutorul metaboliţilor,

Organizarea structural-funcţională
E sunt fixate prin legături slabe pe o proteină
Organizarea structural-funcţională
E sunt fixate prin legături slabe pe o proteină

Organizarea structural-funcţională
Ex.- complexul polienzimatic piruvatdehidrogenazic, constituit din 3 E şi 5
Organizarea structural-funcţională
Ex.- complexul polienzimatic piruvatdehidrogenazic, constituit din 3 E şi 5

Tipul mixt de organizare
reprezintă o îmbinare a ambelor tipuri de organizare,
Tipul mixt de organizare
reprezintă o îmbinare a ambelor tipuri de organizare,

Unităţile de măsurare a activităţii E
1 UI – cantitatea de E
Unităţile de măsurare a activităţii E
1 UI – cantitatea de E

1 cat = 6 · 107 UI
1 UI = 16.67
1 cat = 6 · 107 UI
1 UI = 16.67

Unităţile de măsurare a activităţii E
Activitatea specifică – reprezintă numărul de
Unităţile de măsurare a activităţii E
Activitatea specifică – reprezintă numărul de

Metodele de separare şi purificare ale E
Dializă
Salifiere
Cromatografie
Gel-filtrare
Electroforeză
Cea mai eficientă –
cromatografia
Metodele de separare şi purificare ale E
Dializă
Salifiere
Cromatografie
Gel-filtrare
Electroforeză
Cea mai eficientă –
cromatografia

Metodele de determinare a activităţii E
Viteza reacţiei este proporţională cu
Viteza
Metodele de determinare a activităţii E
Viteza reacţiei este proporţională cu
Viteza

Deosebirea privind componenţa enzimatică a organelor şi ţesuturilor. Enzimele organospecifice.
Enzimele
Deosebirea privind componenţa enzimatică a organelor şi ţesuturilor. Enzimele organospecifice.
Enzimele


Utilizarea E în practica medicală
Preparatele farmaceutice contemporane sunt asociate şi conţin
Utilizarea E în practica medicală
Preparatele farmaceutice contemporane sunt asociate şi conţin

Utilizarea E în practica medicală
Efectele
exercitate de preparatele enzimatice
De substituţie (E
Utilizarea E în practica medicală
Efectele
exercitate de preparatele enzimatice
De substituţie (E

Terapia cu enzime
Enzimele sunt agenţi terapeutici unici ce produc efecte importante
Terapia cu enzime
Enzimele sunt agenţi terapeutici unici ce produc efecte importante

Tehnici propuse pentru optimizarea proprietăţilor terapeutice ale enzimelor.
- prin N-acilare a
Tehnici propuse pentru optimizarea proprietăţilor terapeutice ale enzimelor.
- prin N-acilare a
 Отряд Парнокопытные
Отряд Парнокопытные Синапсы
Синапсы Строение, свойства, биологическая роль нуклеотидов и нуклеиновых кислот. Катаболизм нуклеиновых кислот
Строение, свойства, биологическая роль нуклеотидов и нуклеиновых кислот. Катаболизм нуклеиновых кислот Endangered species
Endangered species Задания ЕГЭ по теме Антропогенез
Задания ЕГЭ по теме Антропогенез Regnul plante
Regnul plante Airfow and Ventilation
Airfow and Ventilation Семейство растений луковые
Семейство растений луковые Сообщество. Экосистема. Биогеоценоз
Сообщество. Экосистема. Биогеоценоз Жизнь в аквариуме
Жизнь в аквариуме Презентация Бактерии 5 класс
Презентация Бактерии 5 класс Кроссворд по биологии по теме: Антропогенез
Кроссворд по биологии по теме: Антропогенез Основные экологические группы растений
Основные экологические группы растений Животные Африки. (ДОУ)
Животные Африки. (ДОУ) Выращивание картофеля на даче
Выращивание картофеля на даче Клетка
Клетка Класс Пресмыкающиеся
Класс Пресмыкающиеся Жабы и лягушки. 2 класс
Жабы и лягушки. 2 класс Полимеразная цепная реакция
Полимеразная цепная реакция Мезозойская эра
Мезозойская эра Класс Насекомые. Внешнее строение майского жука
Класс Насекомые. Внешнее строение майского жука Методы исследования генетики человека
Методы исследования генетики человека Молекулярные основы наследственности
Молекулярные основы наследственности Тип Кишечнополостные. Гидра. на основе современных информационных технологий.
Тип Кишечнополостные. Гидра. на основе современных информационных технологий. Террариум и его обитатели
Террариум и его обитатели Млекопитающие белки. Виды белок
Млекопитающие белки. Виды белок Травлення у Кишечнику. Функції печінки
Травлення у Кишечнику. Функції печінки Мхи и лишайники
Мхи и лишайники