- Биоинженерные эпохи

Содержание
- 2. Секвенирование Сегодня, ребятки, у нас будет переходная лекция! Мы переходим из одной биоинженерной эпохи в другую.
- 3. Вторая биоинженерная эпоха: работа с геномами (продолжается по настоящее время) Что к сегодняшнему дню научились делать
- 4. Секвенирование по Максаму-Гилберту Первый метод определения последовательности нуклеотидов ДНК, 1970-е годы Под действием различных химических агентов
- 5. Как пометить радиоактивностью двуцепочечную ДНК? Выступающий 5’-конец – можно пометить ДНК-полимеразой, добавив меченные нуклеотиды (матричный синтез).
- 6. Метод Сэнгера Появился в конце 1970-х годов и мгновенно вытеснил метод Максама-Гилберта. Основа данного метода –
- 7. Пометить олигонуклеотид – ваще не вопрос, пацаны! 1. Обработайте олигонуклеотид щелочной фосфатазой (АР). Она откусит пирофосфат
- 8. Метод Сэнгера меченный!!! Проведите 4 независимых реакции удлинения праймера с каждым из ddNTP, нанесите результат на
- 9. Результат метода Сэнгера За один прогон на геле можно было прочесть не больше 100 нуклеотидов.
- 10. Сравнение методов секвенирования по Максаму-Гилберту и по Сэнгеру Неудивительно, что метод Максама-Гилберта был очень быстро вытеснен
- 11. Автоматические секвенаторы, использующие метод Сэнгера Четыре разных флюоресцентных красителя => одна реакция в одной пробирке Капиллярный
- 12. Набор флюоресцентных красителей BigDye для мечения ddNTPs
- 13. Автоматический секвенатор дает на выходе набор пиков и сам же пишет соответствующую нуклеотидную последовательность. Очень удобно!
- 14. Высокопроизводительное секвенирование (секвенирование следующего поколения, Next Generation Sequencing, NGS) Общее название нескольких методов секвенирования, объединенных следующими
- 15. Пиросеквенирование (Sequencing By Synthesis) Метод основан на детекции пирофосфата, высвобождающегося при присоединении каждого нового нуклеотида к
- 16. Пиросеквенирование ДНК фрагментируется, к фрагментам пришиваются олигонуклеотиды-«адаптеры»; полученные двуцепочечные молекулы ДНК разделяются на две комплементарные цепи.
- 17. Пиросеквенирование. Схема эПЦР. Бусинка покрыта адаптерными олигонуклеотидами. В каплю попадает один фрагмент ДНК… и амплифицируется!
- 18. Пиросеквенирование Эмульсия разбивается, и цепи ДНК-фрагментов, образовавшиеся в результате эПЦР, разделяются. Бусинки, несущие на своей поверхности
- 19. Пиросеквенирование ДНК– полимераза катализирует добавление dNTP к праймеру для секвенирования, если он комплементарен к последовательности матрицы
- 20. Результат пиросеквенирования Отсутствие пика – что-то пошло не так. Пик в два раза выше обычного –
- 21. Ионное полупроводниковое секвенирование (ion semiconductor sequencing) В целом, этот метод тоже можно назвать Sequencing by Synthesis.
- 22. Ионное полупроводниковое секвенирование В ячейку на каждом этапе синтеза добавляются разные dNTP по очереди. Когда какой-то
- 23. Ионное полупроводниковое секвенирование Вот у вас и получается нуклеотидная последовательность!
- 24. Циклическое лигазное секвенирование (Sequencing By Ligation) Метод основан на детекции флюорофора при лигировании олигонуклеотида к секвенируемой
- 25. Циклическое лигазное секвенирование Первые этапы технологии аналогичны таковым для пиросеквенирования: Фрагментация ДНК Пришивание адаптеров Распределение фрагментов
- 26. Циклическое лигазное секвенирование … и посредством этой модификации закрепляются в проточных ячейках. В ячейки сначала добавляется
- 27. Циклическое лигазное секвенирование Универсальный олигонуклеотид отжегся на секвенируемую ДНК. Среди хитрых нуклеотидов предпочтительно будут отжигаться те,
- 28. Циклическое лигазное секвенирование Итак, X-Y-n-n-n у вас теперь лигированы к универсальному олигонуклеотиду и находятся в составе
- 29. Циклическое лигазное секвенирование Когда вы дошли до конца секвенируемой молекулы, уберите с нее все, что там
- 30. Циклическое лигазное секвенирование В результате этой невероятно красивой по своей идее процедуры у вас за несколько
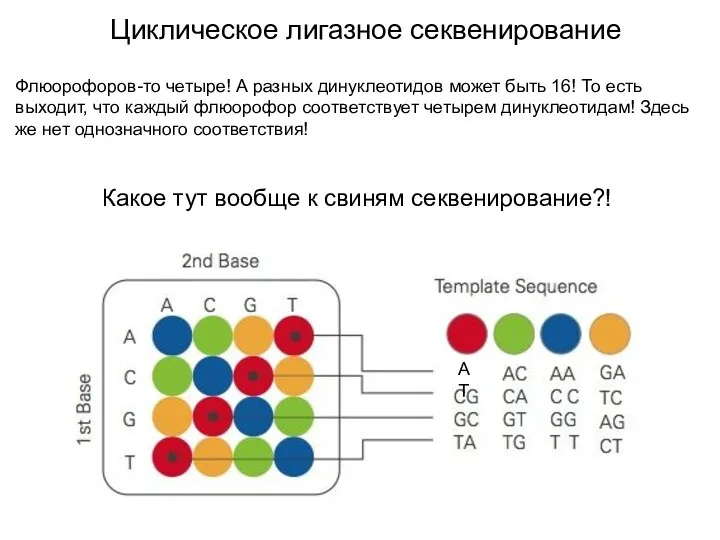
- 31. Циклическое лигазное секвенирование Флюорофоров-то четыре! А разных динуклеотидов может быть 16! То есть выходит, что каждый
- 33. Скачать презентацию

Секвенирование
Сегодня, ребятки, у нас будет переходная лекция! Мы переходим из одной
Секвенирование
Сегодня, ребятки, у нас будет переходная лекция! Мы переходим из одной

Вторая биоинженерная эпоха: работа с геномами
(продолжается по настоящее время)
Что к сегодняшнему
Вторая биоинженерная эпоха: работа с геномами
(продолжается по настоящее время)
Что к сегодняшнему

Секвенирование по Максаму-Гилберту
Первый метод определения последовательности нуклеотидов ДНК, 1970-е годы
Под действием
Секвенирование по Максаму-Гилберту
Первый метод определения последовательности нуклеотидов ДНК, 1970-е годы
Под действием
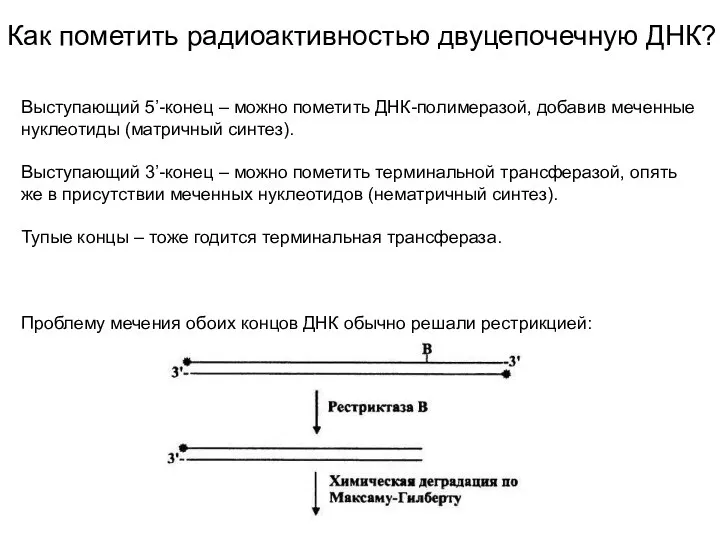
Как пометить радиоактивностью двуцепочечную ДНК?
Выступающий 5’-конец – можно пометить ДНК-полимеразой, добавив
Как пометить радиоактивностью двуцепочечную ДНК?
Выступающий 5’-конец – можно пометить ДНК-полимеразой, добавив
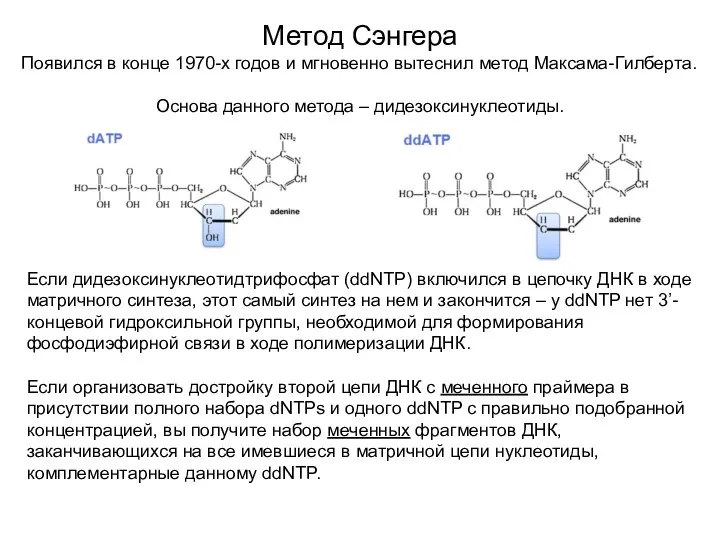
Метод Сэнгера
Появился в конце 1970-х годов и мгновенно вытеснил метод Максама-Гилберта.
Основа
Метод Сэнгера
Появился в конце 1970-х годов и мгновенно вытеснил метод Максама-Гилберта.
Основа

Пометить олигонуклеотид – ваще не вопрос, пацаны!
1. Обработайте олигонуклеотид щелочной фосфатазой
Пометить олигонуклеотид – ваще не вопрос, пацаны!
1. Обработайте олигонуклеотид щелочной фосфатазой
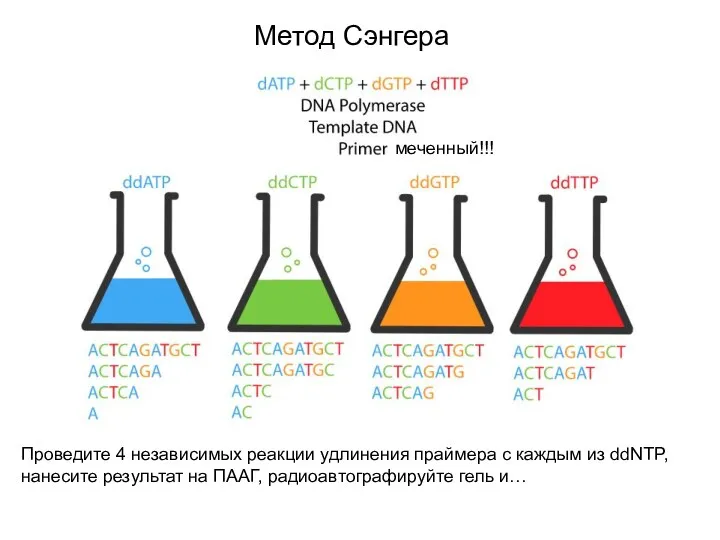
Метод Сэнгера
меченный!!!
Проведите 4 независимых реакции удлинения праймера с каждым из ddNTP,
Метод Сэнгера
меченный!!!
Проведите 4 независимых реакции удлинения праймера с каждым из ddNTP,
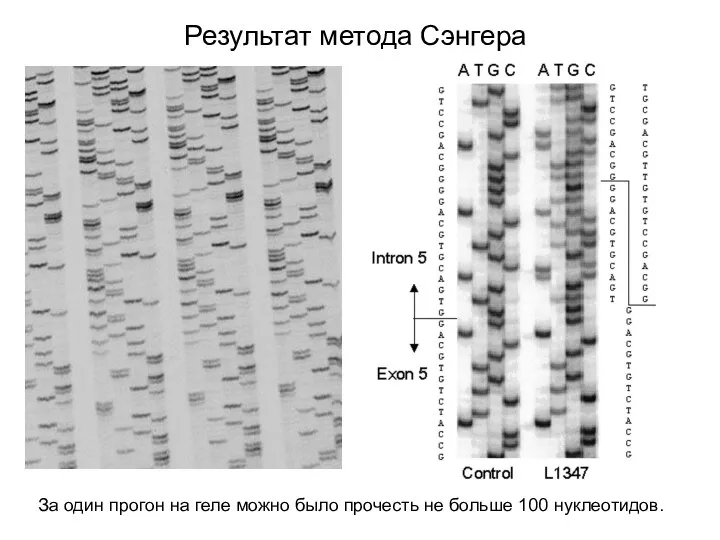
Результат метода Сэнгера
За один прогон на геле можно было прочесть не
Результат метода Сэнгера
За один прогон на геле можно было прочесть не

Сравнение методов секвенирования по Максаму-Гилберту и по Сэнгеру
Неудивительно, что метод Максама-Гилберта
Сравнение методов секвенирования по Максаму-Гилберту и по Сэнгеру
Неудивительно, что метод Максама-Гилберта

Автоматические секвенаторы, использующие метод Сэнгера
Четыре разных флюоресцентных красителя => одна реакция
Автоматические секвенаторы, использующие метод Сэнгера
Четыре разных флюоресцентных красителя => одна реакция
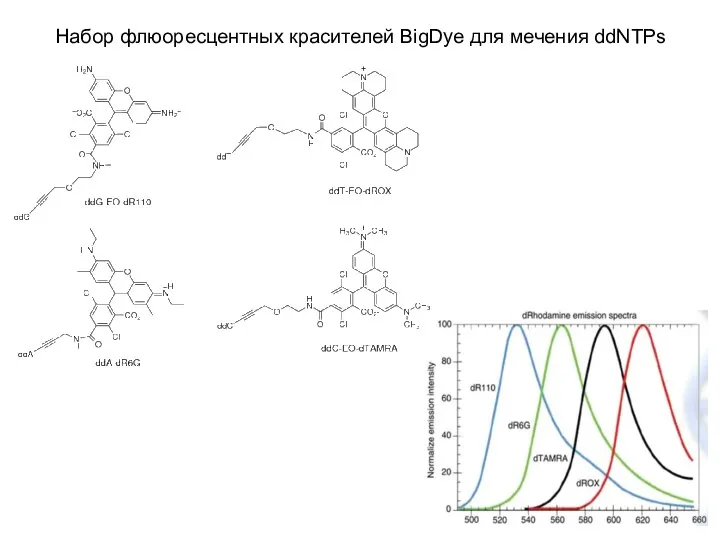
Набор флюоресцентных красителей BigDye для мечения ddNTPs
Набор флюоресцентных красителей BigDye для мечения ddNTPs

Автоматический секвенатор дает на выходе набор пиков и сам же пишет
Автоматический секвенатор дает на выходе набор пиков и сам же пишет

Высокопроизводительное секвенирование
(секвенирование следующего поколения,
Next Generation Sequencing, NGS)
Общее название нескольких методов
Высокопроизводительное секвенирование
(секвенирование следующего поколения,
Next Generation Sequencing, NGS)
Общее название нескольких методов

Пиросеквенирование (Sequencing By Synthesis)
Метод основан на детекции пирофосфата, высвобождающегося при присоединении
Пиросеквенирование (Sequencing By Synthesis)
Метод основан на детекции пирофосфата, высвобождающегося при присоединении
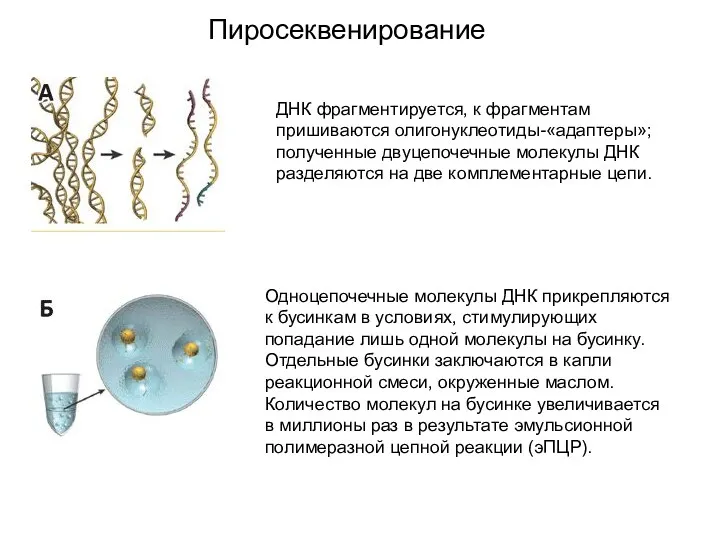
Пиросеквенирование
ДНК фрагментируется, к фрагментам пришиваются олигонуклеотиды-«адаптеры»; полученные двуцепочечные молекулы ДНК разделяются на две
Пиросеквенирование
ДНК фрагментируется, к фрагментам пришиваются олигонуклеотиды-«адаптеры»; полученные двуцепочечные молекулы ДНК разделяются на две

Пиросеквенирование. Схема эПЦР.
Бусинка покрыта адаптерными олигонуклеотидами.
В каплю попадает один фрагмент ДНК…
Пиросеквенирование. Схема эПЦР.
Бусинка покрыта адаптерными олигонуклеотидами.
В каплю попадает один фрагмент ДНК…
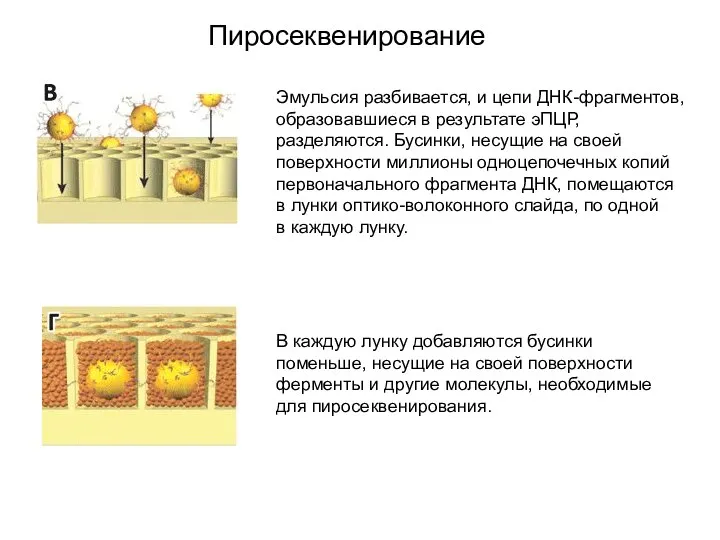
Пиросеквенирование
Эмульсия разбивается, и цепи ДНК-фрагментов, образовавшиеся в результате эПЦР, разделяются. Бусинки, несущие на своей
Пиросеквенирование
Эмульсия разбивается, и цепи ДНК-фрагментов, образовавшиеся в результате эПЦР, разделяются. Бусинки, несущие на своей
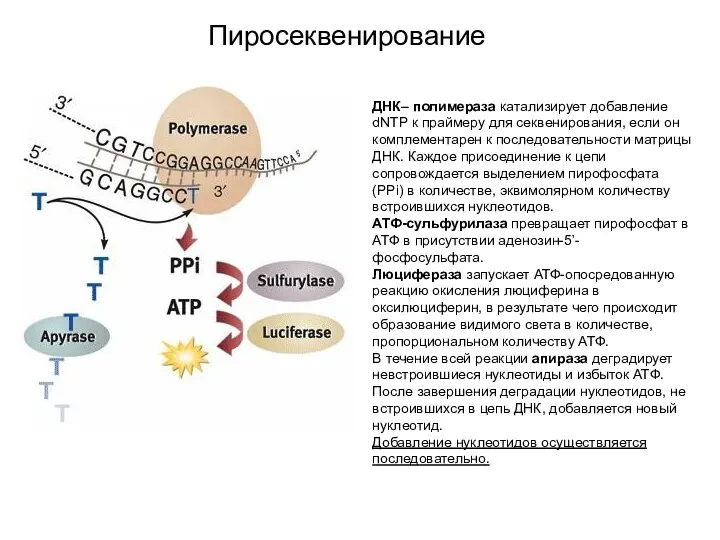
Пиросеквенирование
ДНК– полимераза катализирует добавление dNTP к праймеру для секвенирования, если он
Пиросеквенирование
ДНК– полимераза катализирует добавление dNTP к праймеру для секвенирования, если он
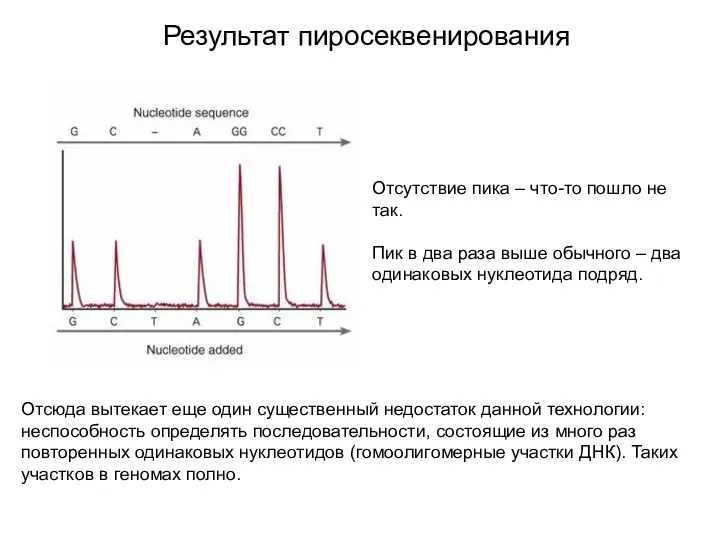
Результат пиросеквенирования
Отсутствие пика – что-то пошло не так.
Пик в два раза
Результат пиросеквенирования
Отсутствие пика – что-то пошло не так.
Пик в два раза

Ионное полупроводниковое секвенирование
(ion semiconductor sequencing)
В целом, этот метод тоже можно назвать
Ионное полупроводниковое секвенирование
(ion semiconductor sequencing)
В целом, этот метод тоже можно назвать
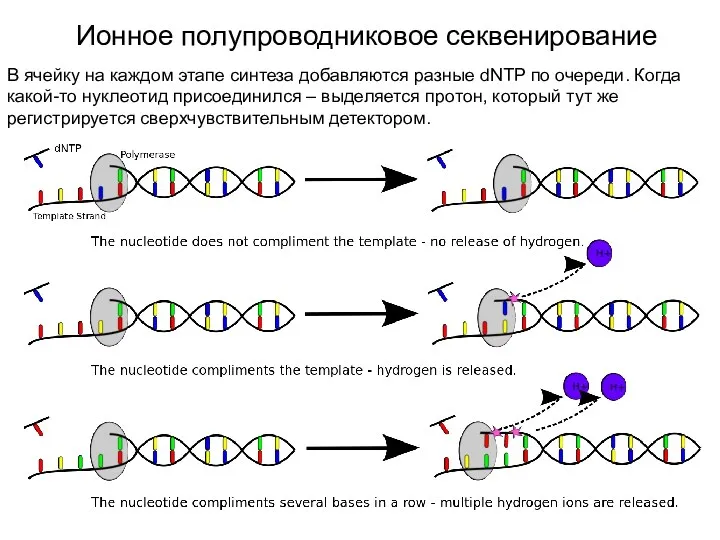
Ионное полупроводниковое секвенирование
В ячейку на каждом этапе синтеза добавляются разные dNTP
Ионное полупроводниковое секвенирование
В ячейку на каждом этапе синтеза добавляются разные dNTP
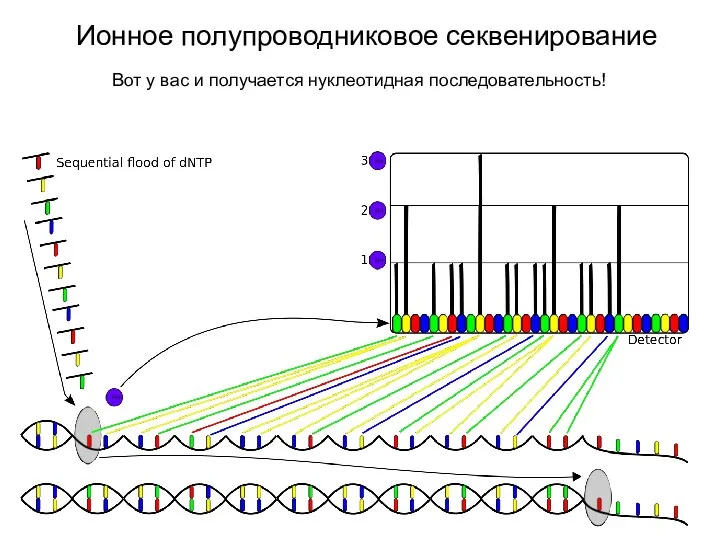
Ионное полупроводниковое секвенирование
Вот у вас и получается нуклеотидная последовательность!
Ионное полупроводниковое секвенирование
Вот у вас и получается нуклеотидная последовательность!

Циклическое лигазное секвенирование
(Sequencing By Ligation)
Метод основан на детекции флюорофора при
Циклическое лигазное секвенирование
(Sequencing By Ligation)
Метод основан на детекции флюорофора при
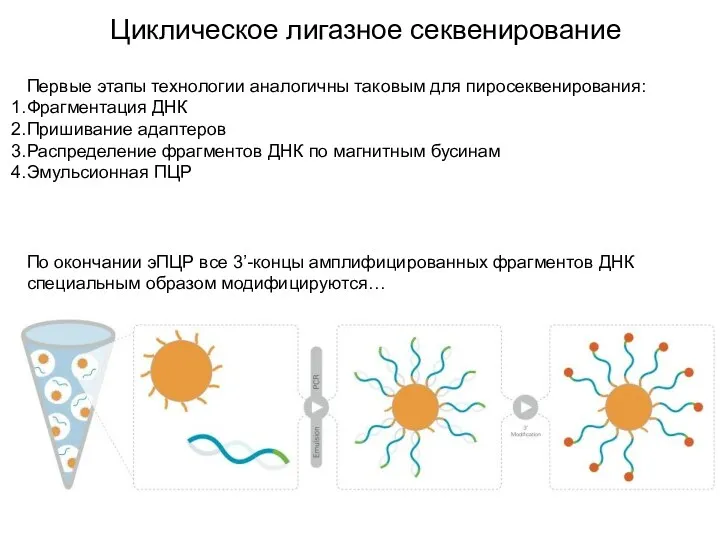
Циклическое лигазное секвенирование
Первые этапы технологии аналогичны таковым для пиросеквенирования:
Фрагментация ДНК
Пришивание адаптеров
Распределение
Циклическое лигазное секвенирование
Первые этапы технологии аналогичны таковым для пиросеквенирования:
Фрагментация ДНК
Пришивание адаптеров
Распределение

Циклическое лигазное секвенирование
… и посредством этой модификации закрепляются в проточных ячейках.
В
Циклическое лигазное секвенирование
… и посредством этой модификации закрепляются в проточных ячейках.
В
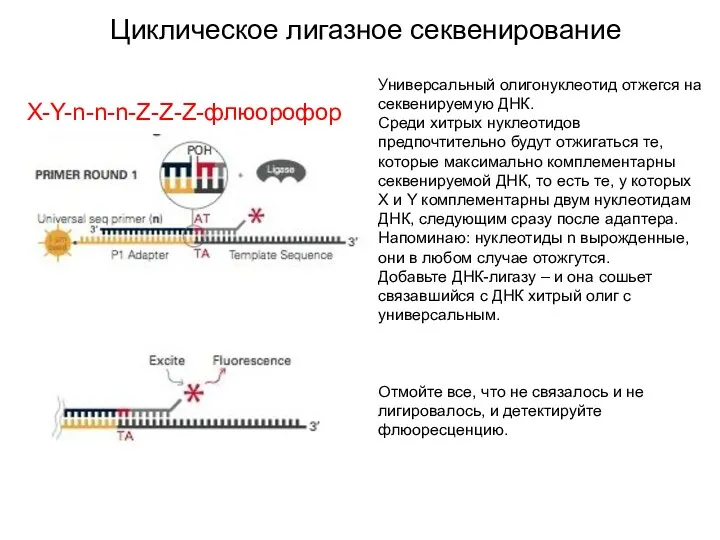
Циклическое лигазное секвенирование
Универсальный олигонуклеотид отжегся на секвенируемую ДНК.
Среди хитрых нуклеотидов предпочтительно
Циклическое лигазное секвенирование
Универсальный олигонуклеотид отжегся на секвенируемую ДНК.
Среди хитрых нуклеотидов предпочтительно
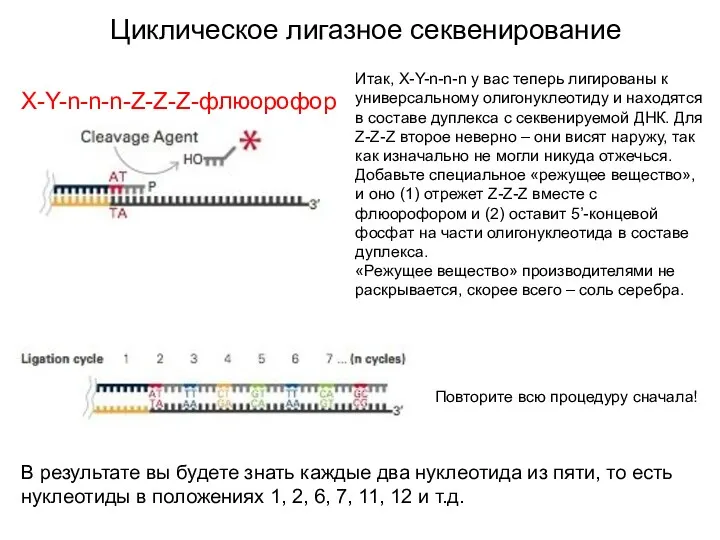
Циклическое лигазное секвенирование
Итак, X-Y-n-n-n у вас теперь лигированы к универсальному олигонуклеотиду
Циклическое лигазное секвенирование
Итак, X-Y-n-n-n у вас теперь лигированы к универсальному олигонуклеотиду
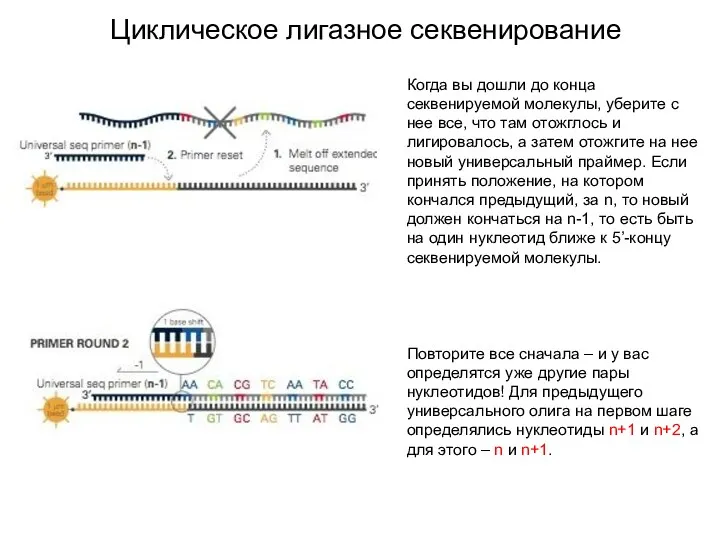
Циклическое лигазное секвенирование
Когда вы дошли до конца секвенируемой молекулы, уберите с
Циклическое лигазное секвенирование
Когда вы дошли до конца секвенируемой молекулы, уберите с
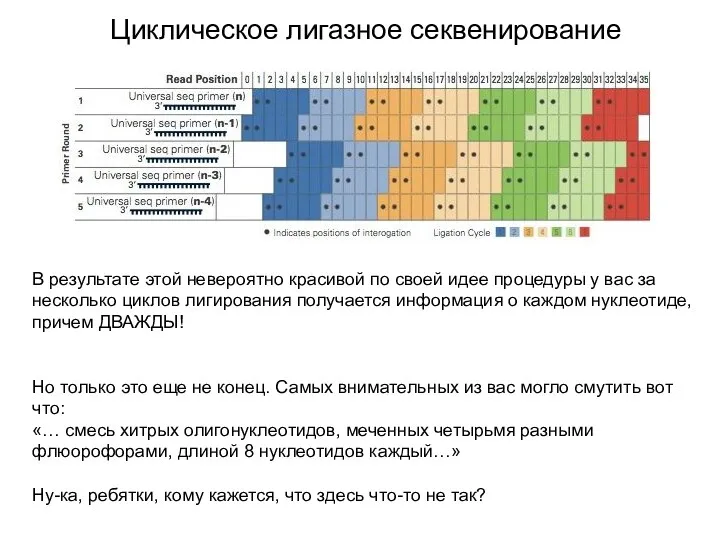
Циклическое лигазное секвенирование
В результате этой невероятно красивой по своей идее процедуры
Циклическое лигазное секвенирование
В результате этой невероятно красивой по своей идее процедуры
Циклическое лигазное секвенирование
Флюорофоров-то четыре! А разных динуклеотидов может быть 16! То
Циклическое лигазное секвенирование
Флюорофоров-то четыре! А разных динуклеотидов может быть 16! То
 Цианобактерии. Отличия от бактерий
Цианобактерии. Отличия от бактерий Презентация к уроку 5 класса ФГОС
Презентация к уроку 5 класса ФГОС Серологические реакции
Серологические реакции Физиология анализаторов
Физиология анализаторов Полиморфизм человеческих популяций. Генетический груз. Популяционная структура человечества
Полиморфизм человеческих популяций. Генетический груз. Популяционная структура человечества Врожденные и приобретенные формы поведения
Врожденные и приобретенные формы поведения Целительная сила лекарственныхрастений
Целительная сила лекарственныхрастений Овощи: капуста, пасленовые, тыквенные
Овощи: капуста, пасленовые, тыквенные Дыхание. Значение дыхания в живых организмах
Дыхание. Значение дыхания в живых организмах Кровеносная система
Кровеносная система Геохронология Земли
Геохронология Земли Модификационная изменчивость
Модификационная изменчивость Строение волос
Строение волос Периферическая нервная система
Периферическая нервная система Биотехнология туралы түсінік
Биотехнология туралы түсінік Насекомые с полным превращением
Насекомые с полным превращением Закономерности преобразования функций органов в филогенезе. Примеры изменений функций органов у человека
Закономерности преобразования функций органов в филогенезе. Примеры изменений функций органов у человека Биология. Лекция 1. Вводная. Неорганические элементы клеток. Органические вещества
Биология. Лекция 1. Вводная. Неорганические элементы клеток. Органические вещества Взаємозв΄язок метаболічних процесів в рослинних клітинах
Взаємозв΄язок метаболічних процесів в рослинних клітинах Человек как результат биологической и социокультурной эволюции
Человек как результат биологической и социокультурной эволюции Екологічні фактори. Форми біотичних зв’язків
Екологічні фактори. Форми біотичних зв’язків Контейнерное озеленение
Контейнерное озеленение Физиологическая характеристика процессов восстановления после мышечной деятельности
Физиологическая характеристика процессов восстановления после мышечной деятельности Ovqatlanish biokimyosi
Ovqatlanish biokimyosi Zebras are wild animals from Africa
Zebras are wild animals from Africa Среды обитания организмов
Среды обитания организмов Заповедник Брянский лес
Заповедник Брянский лес Анатомия, физиология сердечно-сосудистой системы
Анатомия, физиология сердечно-сосудистой системы