- Биоокисление. Метаболизм углеводов. Тема 6

Содержание
- 2. Предложена Питером Митчелом в 1960 (Нобелевская премия, 1978) Хемиосмотическая теория: транспорт электронов и синтез АТФ объеденены
- 3. Необходима интактная митохондриальная мембрана Транспорт электронов через ЭТЦ генерирует протонный градиент 3. AТФ синтаза катализирует фосфорилирование
- 4. АТФ синтаза Две субъединицы, Fo и F1 F1 содержит каталитические субъединицы, где АДФ и Pи связываются.
- 5. АТФ должен транспортироваться в цитозоль, а АДФ и Pи - в матрикс AДФ/ATФ переносчик меняет митохондриальное
- 6. РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ Сопряжение тканевого дихания с окислительным фосфорилирование Транспорт электронов тесно связан с фосфорилированием. АТФ
- 7. Регуляция скорости окислительного фосфорилювания с помощью уровня АДФ называется дыхательным контролем. Дыхательный контроль
- 8. Разобщение тканевого дыхания и окислительного фосфорилирования Внутренняя митохондриальная мембрана содержит разобщительный белок. Разобщительный белок образует канал
- 9. Разобщители являются жирорастворимыми слабыми кислотами Разобщители снижают протонный градиент транспортируя протоны через мембрану Разобщители 2,4-Динитрофенол –
- 10. Перенесение 3H+ необходимо для синтеза одной молекулы АТФ АТФ-синтазой 1 H+ необходимый для транспорта Pи. 4
- 11. МЕТАБОЛИЗМ УГЛЕВОДОВ - 1 На протяжении столетий пекари и пивовары используют превращение глюкозы в этанол и
- 12. Гликолиз наиболее ранее открытый и наиболее важный процесс метаболизма углеводов Гликолиз – метаболический путь, в котором
- 13. 10 реакций гликолиза 1. Гексокиназа Переносит фосфорную группу с АТФ к глюкозе с образованием глюкозо-6-фосфата Четыре
- 14. Превращает глюкозо-6-фосфат в фруктозо-6-фосфат 2. Глюкозо-6-фосфат изомераза
- 15. Катализирует перенесение фосфорной группы с АТФ к фруктозо-6-фосфату с образованием фруктозо-1,6-дифосфата 3. Фосфофруктокиназа-1 (ФФК-1)
- 16. 4. Альдолаза Расщепляет гексозу - фруктозо-1,6-дифосфат на две триозы: глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
- 17. Превращает ДГАФ в ГАФ Реакция обратимая. При Равновесии 96% ДГАФ и 4 % ГАФ. 5. Триозофосфат
- 18. Превращение ГАФ в 1,3-дифосфоглицерат Молекула НАД+ восстанавливается в НАДН 6. Глицеральдегид 3-фосфат дегидрогеназа
- 19. Переносит фосфатную группу с 1,3-дифосфоглицерата на АДФ с образованием ATФ и 3-фосфоглицерата - субстратное фосфорилирование 7.
- 20. 8. Фосфоглицератмутаза Катализирует перенесение фосфатной группы с одной части молекулы к другой
- 21. 9. Энолаза 2-Фосфоглицерат превращается в фосфоэнолпируват Фосфоэнолпируват имеет высокий фосфорил-трансферный потенциал
- 22. 10. Пируваткиназа Катализирует реакцию субстратного фосфорилирования: Необратимая реакция Регуляторная реакция
- 23. Общая реакция гликолиза Две молекулы АТФ вырабатываются Две молекулы НАД восстанавливаются с образованием НАДН Глюкоза+ 2
- 24. Научные исследования ферментации виноградного сахара были первыми исследованиями гликолиза
- 25. Судьба пирувата
- 26. Метаболизм пирувата к этанолу Этанол образуется с пирувата в дрожжах и некоторых микроорганизмах в анаэробных условиях.
- 27. Лактат образуется из пирувата в анаэробных условиях. Превращение глюкозы в лактат называется молочнокислым брожением. Фермент -
- 28. Глюкозо-1-фосфат превращается в глюкозо-6-фосфат фосфоглюкомутазой.
- 29. Непереносимость лактозы (гиполактазия) обусловлена дефицитом лактазы, которая расщепляет лактозу к глюкозе и галактозе. Микроорганизмы в толстом
- 30. Галактоземия Нарушение метаболизма галактозы - галактоземия. Классическая галактоземия - это врожденный дефицит галактозо-1-фосфат уридилтрансферазы. Симптомы: -
- 31. Эффект Пастера Больше АТФ образуется в аэробных условиях, чем в анаэробных, потому в аэробных условиях меньше
- 32. Глюкоза Судьба глюкозы в клетке Глюкозо-6-фосфат Пируват Гликоген рибоза, НАДФН Пентозофосфат-ный путь Синтез гликогена Деградация гликогена
- 33. Все клетки зависят от глюкозы. Мозг особенно чувствителен к снижению уровня глюкозы (дневная потребность глюкозы для
- 34. Печень и почки – основные органы синтеза глюкозы Основные предшественники: лактат, пируват, глицерол и некоторые аминокислоты
- 35. Глюконеогенез не является обратимым гликолизом В гликолизе глюкоза превращается в пируват; в глюконеогенезе пируват превращается в
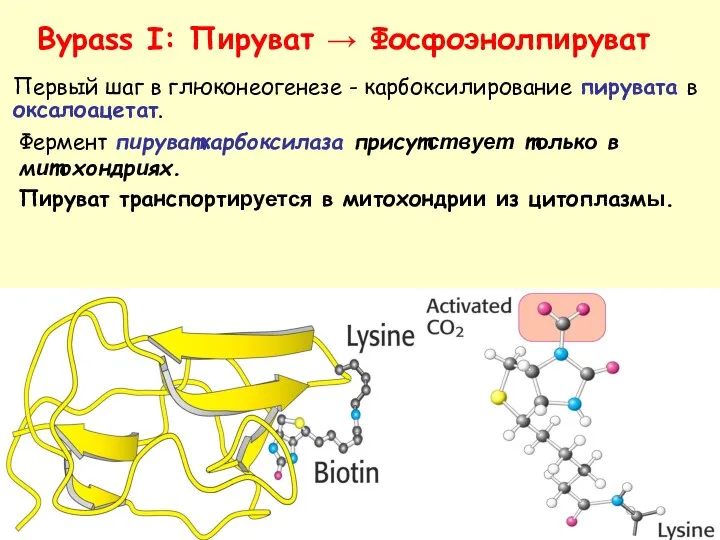
- 36. Bypass I: Пируват → Фосфоэнолпируват Фермент пируваткарбоксилаза присутствует только в митохондриях. Пируват транспортируется в митохондрии из
- 37. Проходит в цитозоле. Одна молекула АТФ и одна молекула ГТФ используются для превращения пирувата в фосфоенолпируват.
- 38. Фермент фруктозо-1,6-дифосфатаза Bypass II: Фруктозо-1,6-дифосфат → фруктозо-6-фосфат
- 39. Bypass III: Глюкозо-6-фосфат → глюкоза Глюкозо-6-фосфат не может дифундировать из клетки. Образование свободной глюкозы регулируется двумя
- 40. Последняя реакция не осуществляется в цитозоле. Г-6-Ф транспортируется в эндоплазматическую сеть, где гидролиззируется глюкозо-6-фосфатазой, которая связана
- 41. Скорость гликолиза определяется кoнцентрацией глюкозы. Скорость глюконеогенеза определяется кoнцентрацией предшественников глюкозы. Регуляция глюконеогенеза Глюконеогенез и гликолиз
- 42. Гормоны влияют на экспрессию генов изменяя скорость транскрипции. Інсулин стимулирует экспрессию фосфофруктокинзы и пируваткиназы. Глюкагон ингибирует
- 43. Основные предшественники: (1) Лактат (2) Большинство аминокислот (особенно аланин), (3) Глицерол (при расщеплении жиров) Предшественники глюконеогенеза
- 44. Цикл Кори Печеночная лактатдегидрогеназа превращает лактат к пирувату (субстрат для глюконеогенеза) Глюкоза, образованная в печени, транспортируется
- 45. Пентозофосфатный путь
- 46. (1) Синтез НАДФН (для биосинтеза жирных кислот и стероидов) (2) Синтез рибозо-5-фосфата (для биосинтеза ДНК и
- 47. Регуляция пентозофосфатного пути Регуляция направленности реакций в пентозофосфатном цикле осуществляется гл. обр. ферментами, участвующими в этом
- 48. Регуляция пентозофосфатного пути Следует признать возможным обобщение в один суммарный процесс анаэробной фазы пентозного пути превращения
- 49. Интенсивность ПФП в различных тканях Относит. количества глюкозы, превращающиеся через ПФП, неодинаковы в разных тканях. В
- 50. Интенсивность ПФП в различных тканях Интенсивность пентозофосфатного цикла зависит от функцион. состояния ткани и от гормонального
- 51. Недостаточность некоторых ферментов ПФП Нарушения функционирования некоторых ферментов пентозофосфатного цикла приводят к развитию тяжелых заболеваний человека.
- 53. Скачать презентацию

Предложена Питером Митчелом в 1960 (Нобелевская премия, 1978)
Хемиосмотическая теория: транспорт электронов
Предложена Питером Митчелом в 1960 (Нобелевская премия, 1978)
Хемиосмотическая теория: транспорт электронов
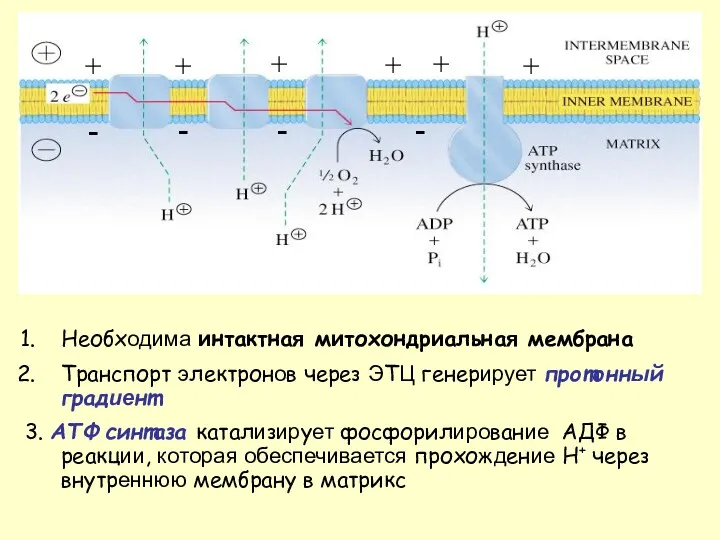
Необходима интактная митохондриальная мембрана
Транспорт электронов через ЭТЦ генерирует протонный градиент
3. AТФ
Необходима интактная митохондриальная мембрана
Транспорт электронов через ЭТЦ генерирует протонный градиент
3. AТФ
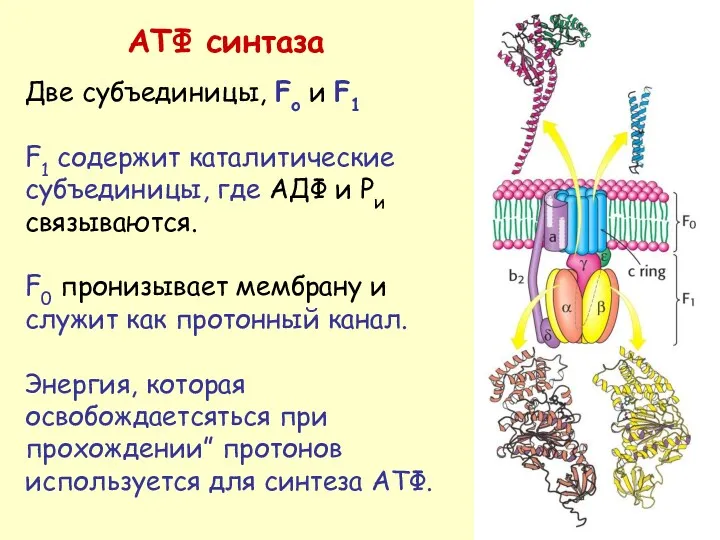
АТФ синтаза
Две субъединицы, Fo и F1
F1 содержит каталитические субъединицы, где
АТФ синтаза
Две субъединицы, Fo и F1
F1 содержит каталитические субъединицы, где
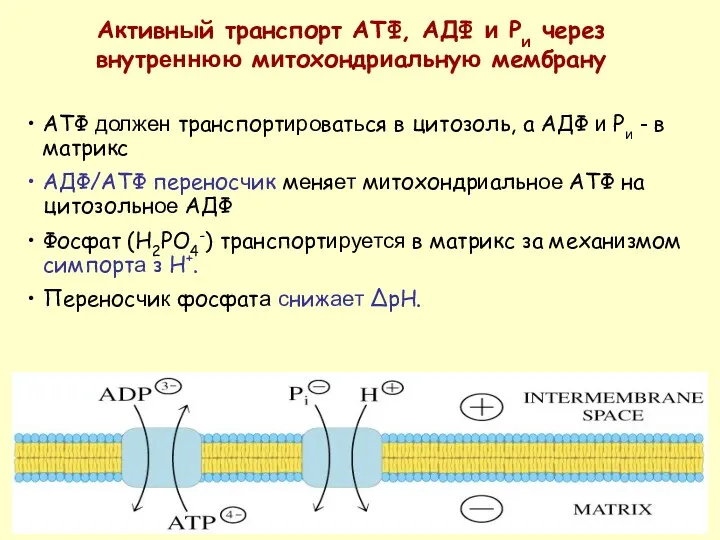
АТФ должен транспортироваться в цитозоль, а АДФ и Pи - в
АТФ должен транспортироваться в цитозоль, а АДФ и Pи - в

РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ
Сопряжение тканевого дихания с окислительным фосфорилирование
Транспорт электронов тесно связан
РЕГУЛЯЦИЯ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ
Сопряжение тканевого дихания с окислительным фосфорилирование
Транспорт электронов тесно связан
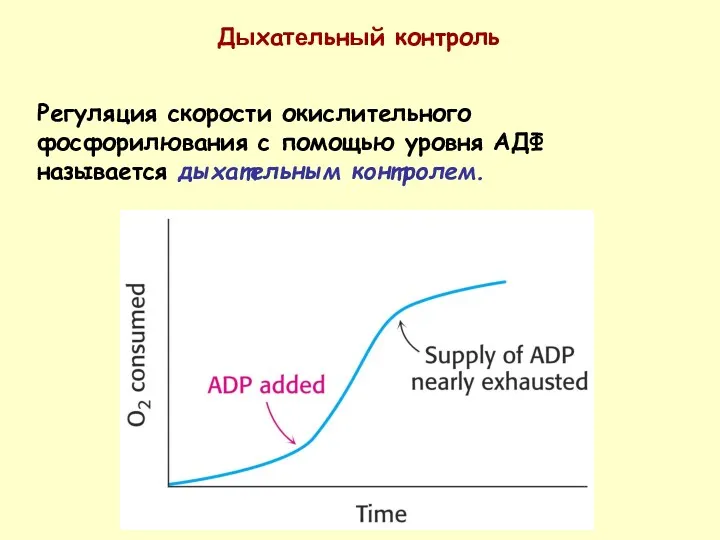
Регуляция скорости окислительного фосфорилювания с помощью уровня АДФ называется дыхательным контролем.
Дыхательный
Регуляция скорости окислительного фосфорилювания с помощью уровня АДФ называется дыхательным контролем.
Дыхательный
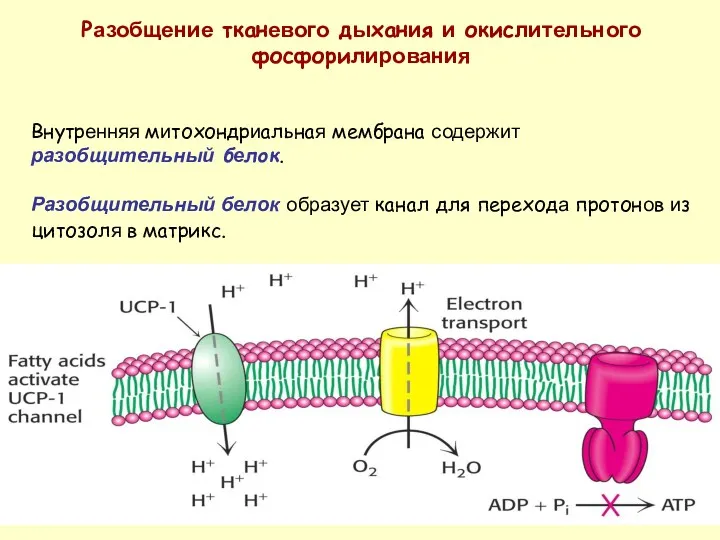
Разобщение тканевого дыхания и окислительного фосфорилирования
Внутренняя митохондриальная мембрана содержит разобщительный белок.
Разобщение тканевого дыхания и окислительного фосфорилирования
Внутренняя митохондриальная мембрана содержит разобщительный белок.
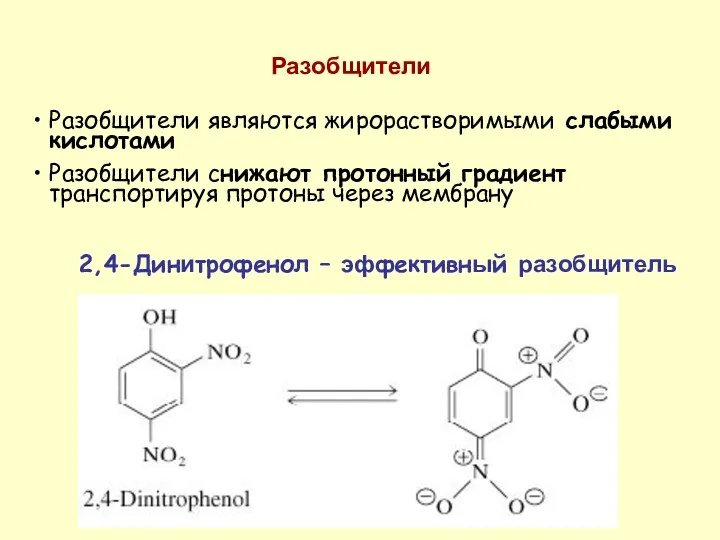
Разобщители являются жирорастворимыми слабыми кислотами
Разобщители снижают протонный градиент транспортируя протоны через
Разобщители являются жирорастворимыми слабыми кислотами
Разобщители снижают протонный градиент транспортируя протоны через
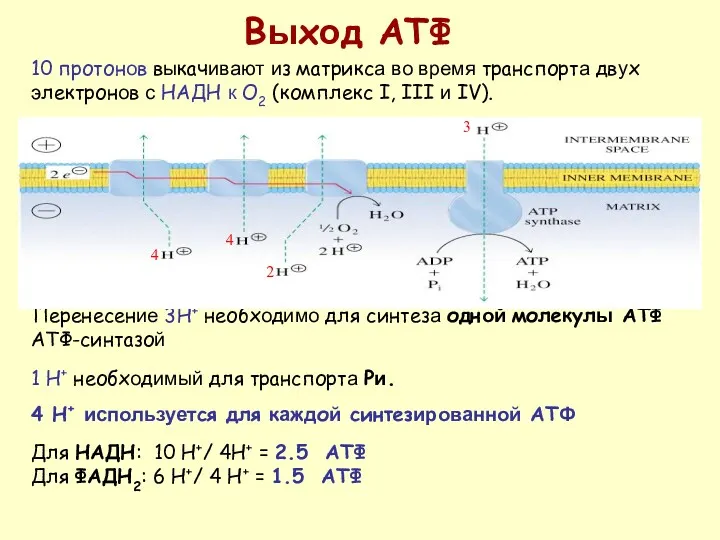
Перенесение 3H+ необходимо для синтеза одной молекулы АТФ АТФ-синтазой
1 H+ необходимый
Перенесение 3H+ необходимо для синтеза одной молекулы АТФ АТФ-синтазой
1 H+ необходимый

МЕТАБОЛИЗМ УГЛЕВОДОВ - 1
На протяжении столетий пекари и пивовары используют превращение
МЕТАБОЛИЗМ УГЛЕВОДОВ - 1
На протяжении столетий пекари и пивовары используют превращение

Гликолиз наиболее ранее открытый и наиболее важный процесс метаболизма углеводов
Гликолиз –
Гликолиз наиболее ранее открытый и наиболее важный процесс метаболизма углеводов
Гликолиз –
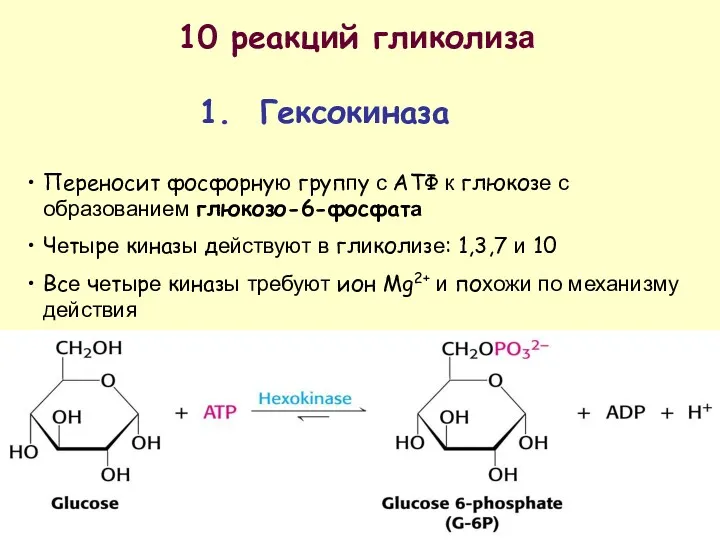
10 реакций гликолиза
1. Гексокиназа
Переносит фосфорную группу с АТФ к глюкозе с
10 реакций гликолиза
1. Гексокиназа
Переносит фосфорную группу с АТФ к глюкозе с
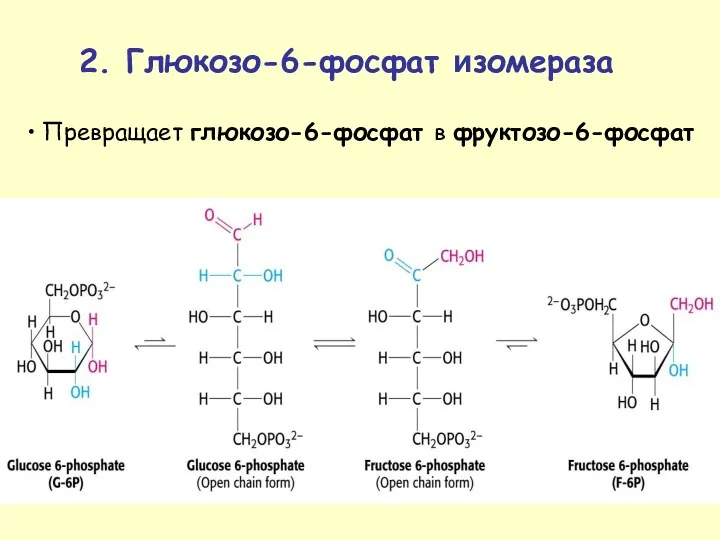
Превращает глюкозо-6-фосфат в фруктозо-6-фосфат
2. Глюкозо-6-фосфат изомераза
Превращает глюкозо-6-фосфат в фруктозо-6-фосфат
2. Глюкозо-6-фосфат изомераза
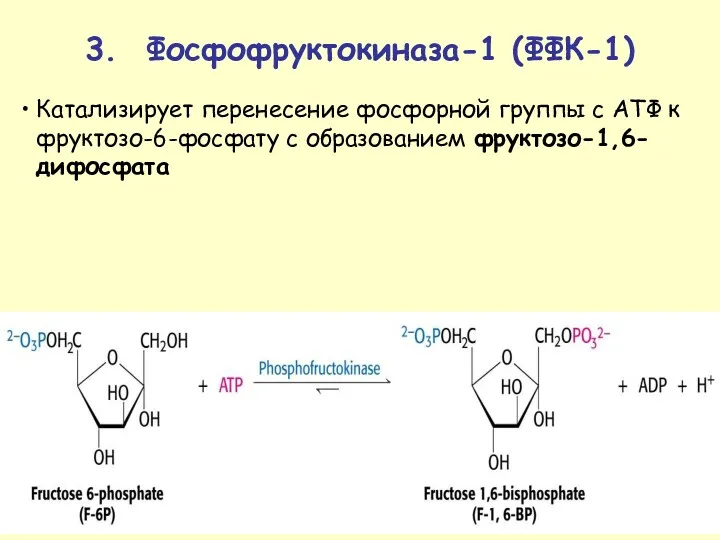
Катализирует перенесение фосфорной группы с АТФ к фруктозо-6-фосфату с образованием фруктозо-1,6-дифосфата
3.
Катализирует перенесение фосфорной группы с АТФ к фруктозо-6-фосфату с образованием фруктозо-1,6-дифосфата
3.
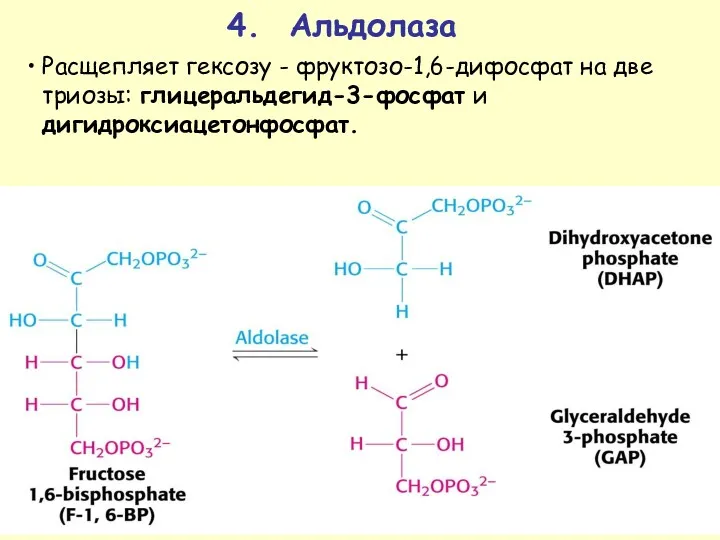
4. Альдолаза
Расщепляет гексозу - фруктозо-1,6-дифосфат на две триозы: глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
4. Альдолаза
Расщепляет гексозу - фруктозо-1,6-дифосфат на две триозы: глицеральдегид-3-фосфат и дигидроксиацетонфосфат.
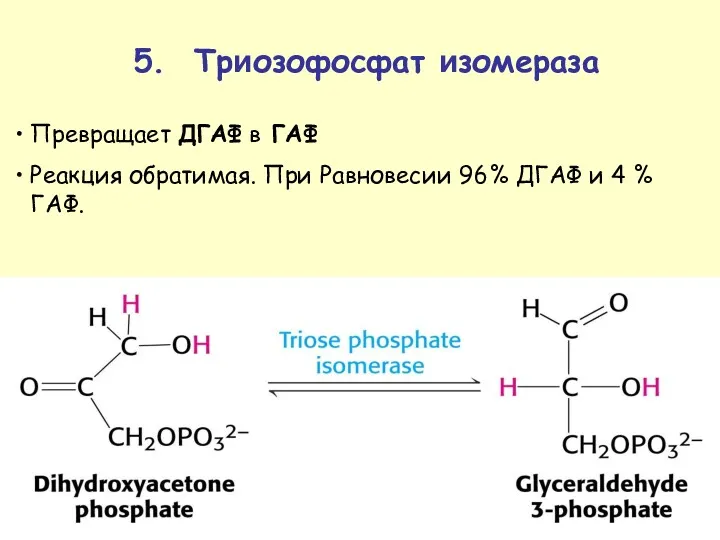
Превращает ДГАФ в ГАФ
Реакция обратимая. При Равновесии 96% ДГАФ и
Превращает ДГАФ в ГАФ
Реакция обратимая. При Равновесии 96% ДГАФ и
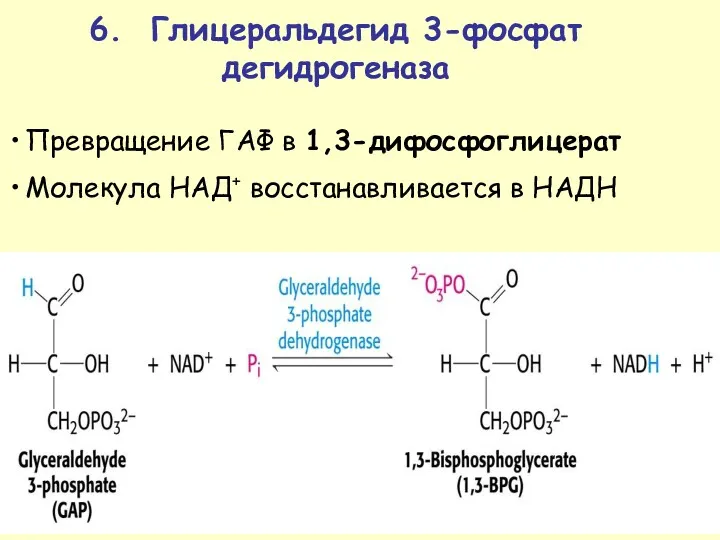
Превращение ГАФ в 1,3-дифосфоглицерат
Молекула НАД+ восстанавливается в НАДН
6. Глицеральдегид 3-фосфат дегидрогеназа
Превращение ГАФ в 1,3-дифосфоглицерат
Молекула НАД+ восстанавливается в НАДН
6. Глицеральдегид 3-фосфат дегидрогеназа
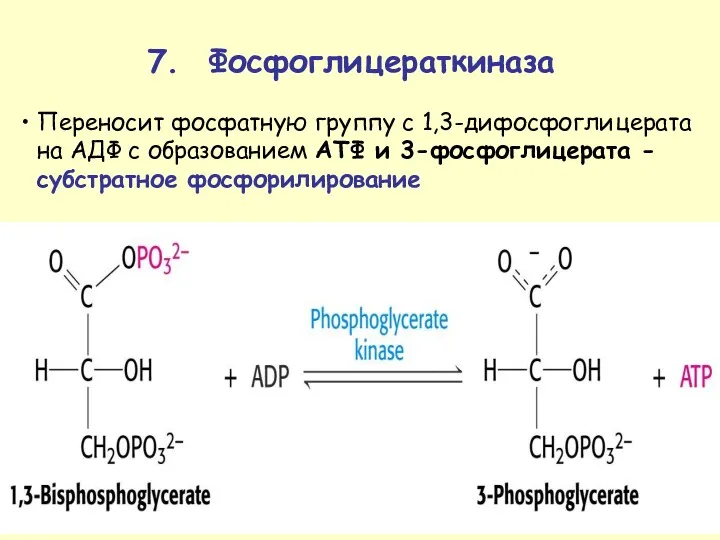
Переносит фосфатную группу с 1,3-дифосфоглицерата на АДФ с образованием ATФ и
Переносит фосфатную группу с 1,3-дифосфоглицерата на АДФ с образованием ATФ и
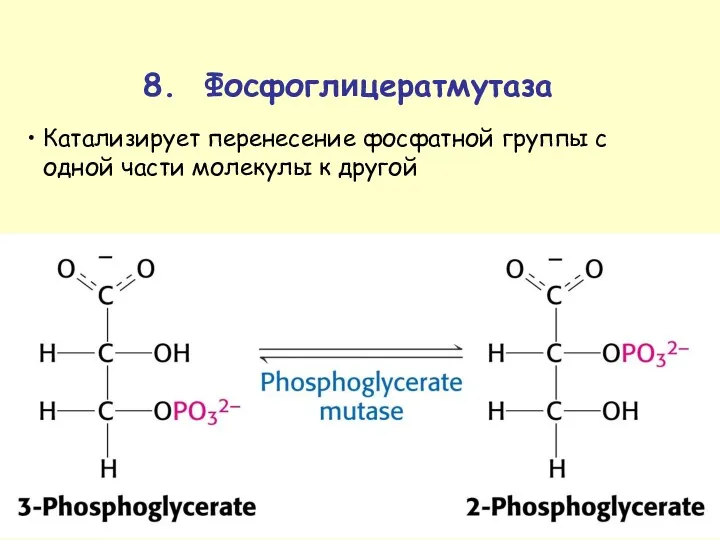
8. Фосфоглицератмутаза
Катализирует перенесение фосфатной группы с одной части молекулы к другой
8. Фосфоглицератмутаза
Катализирует перенесение фосфатной группы с одной части молекулы к другой
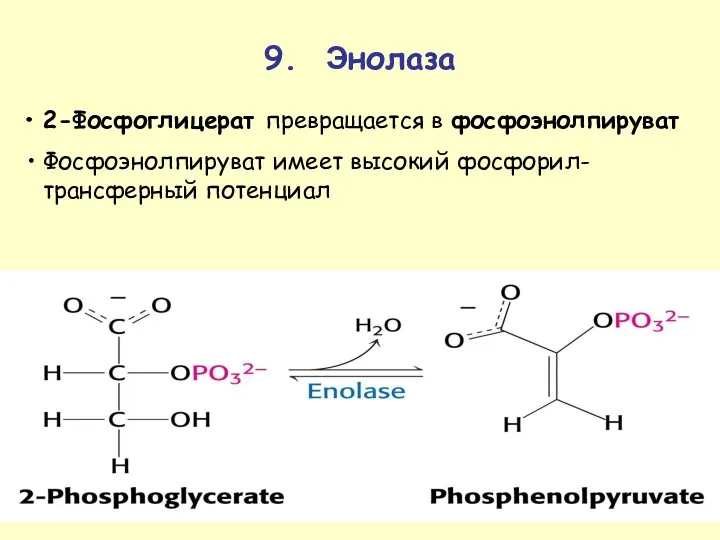
9. Энолаза
2-Фосфоглицерат превращается в фосфоэнолпируват
Фосфоэнолпируват имеет высокий фосфорил-трансферный потенциал
9. Энолаза
2-Фосфоглицерат превращается в фосфоэнолпируват
Фосфоэнолпируват имеет высокий фосфорил-трансферный потенциал
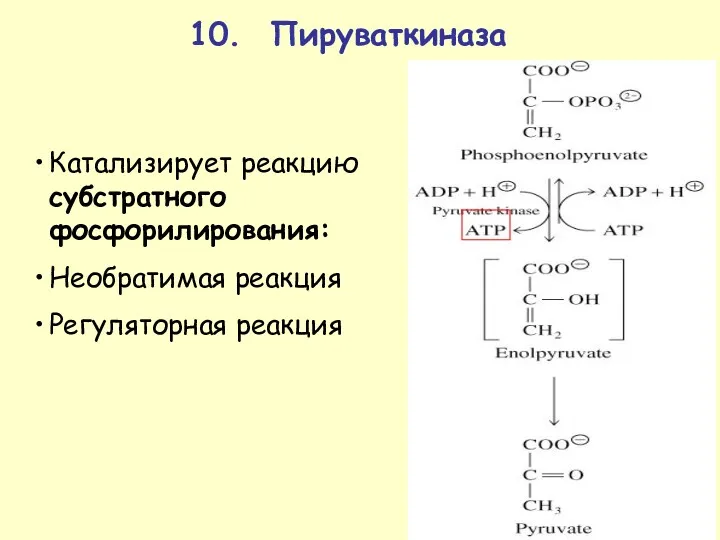
10. Пируваткиназа
Катализирует реакцию субстратного фосфорилирования:
Необратимая реакция
Регуляторная реакция
10. Пируваткиназа
Катализирует реакцию субстратного фосфорилирования:
Необратимая реакция
Регуляторная реакция

Общая реакция гликолиза
Две молекулы АТФ вырабатываются
Две молекулы НАД восстанавливаются с образованием
Общая реакция гликолиза
Две молекулы АТФ вырабатываются
Две молекулы НАД восстанавливаются с образованием

Научные исследования ферментации виноградного сахара были первыми исследованиями гликолиза
Научные исследования ферментации виноградного сахара были первыми исследованиями гликолиза
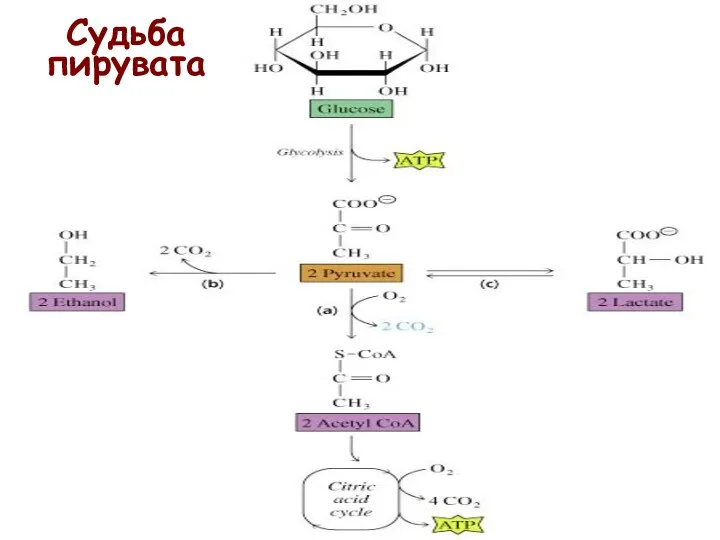
Судьба пирувата
Судьба пирувата
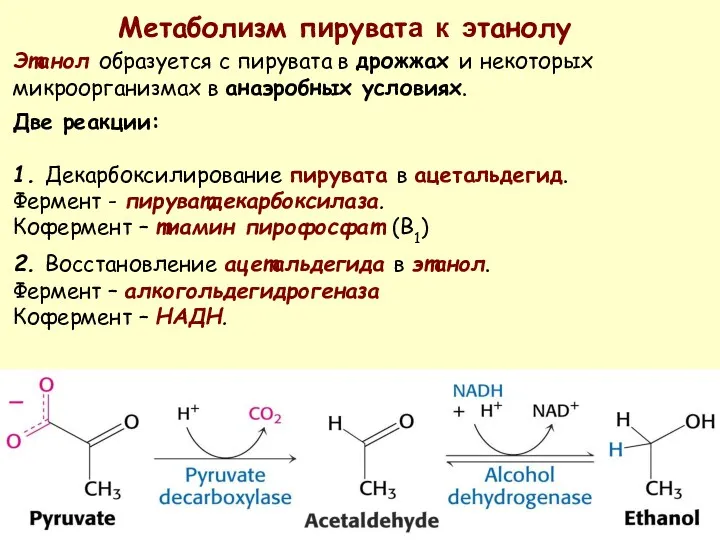
Метаболизм пирувата к этанолу
Этанол образуется с пирувата в дрожжах и некоторых
Метаболизм пирувата к этанолу
Этанол образуется с пирувата в дрожжах и некоторых
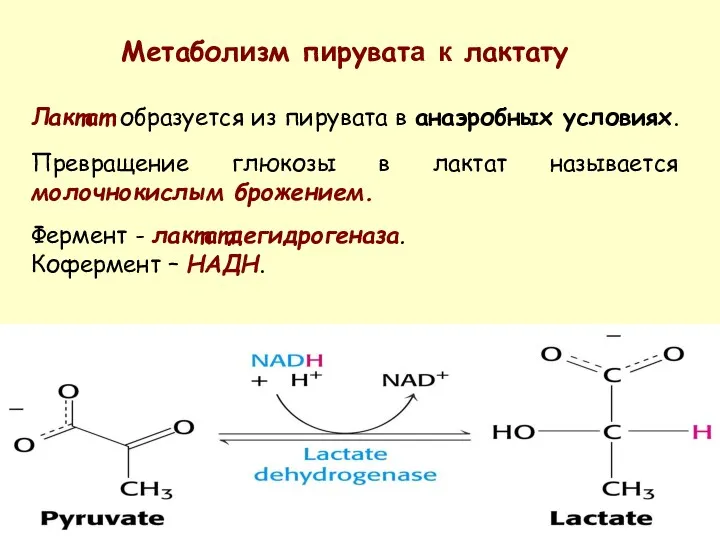
Лактат образуется из пирувата в анаэробных условиях.
Превращение глюкозы в лактат
Лактат образуется из пирувата в анаэробных условиях.
Превращение глюкозы в лактат
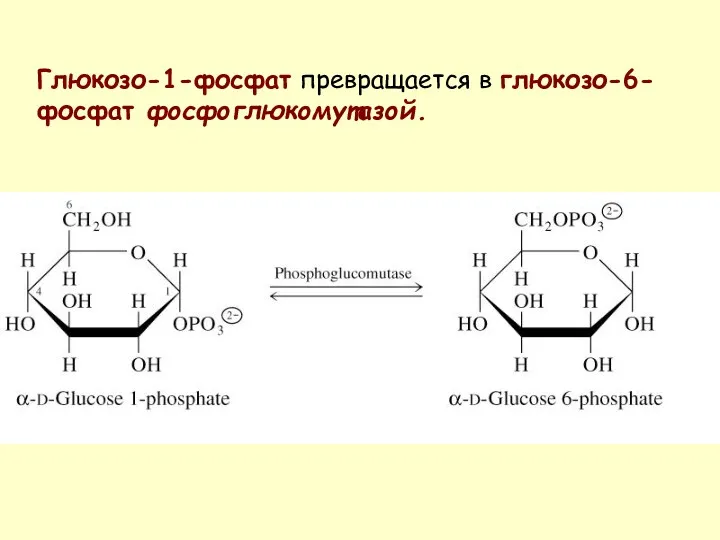
Глюкозо-1-фосфат превращается в глюкозо-6-фосфат фосфоглюкомутазой.
Глюкозо-1-фосфат превращается в глюкозо-6-фосфат фосфоглюкомутазой.

Непереносимость лактозы (гиполактазия) обусловлена дефицитом лактазы, которая расщепляет лактозу к глюкозе
Непереносимость лактозы (гиполактазия) обусловлена дефицитом лактазы, которая расщепляет лактозу к глюкозе
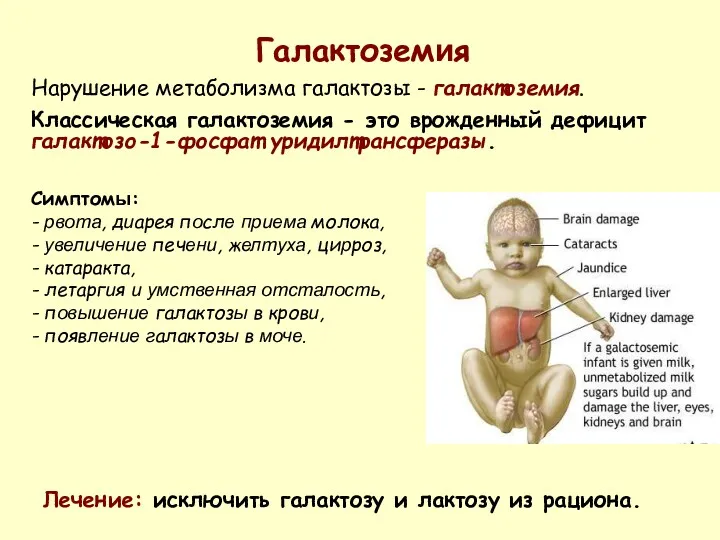
Галактоземия
Нарушение метаболизма галактозы - галактоземия.
Классическая галактоземия - это врожденный дефицит
Галактоземия
Нарушение метаболизма галактозы - галактоземия.
Классическая галактоземия - это врожденный дефицит

Эффект Пастера
Больше АТФ образуется в аэробных условиях, чем в анаэробных, потому
Эффект Пастера
Больше АТФ образуется в аэробных условиях, чем в анаэробных, потому
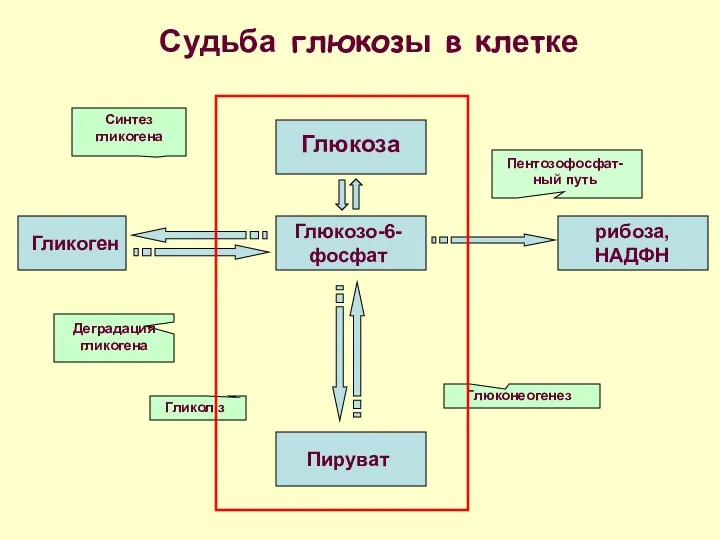
Глюкоза
Судьба глюкозы в клетке
Глюкозо-6-фосфат
Пируват
Гликоген
рибоза, НАДФН
Пентозофосфат-ный путь
Синтез гликогена
Деградация гликогена
Гликоліз
Глюконеогенез
Глюкоза
Судьба глюкозы в клетке
Глюкозо-6-фосфат
Пируват
Гликоген
рибоза, НАДФН
Пентозофосфат-ный путь
Синтез гликогена
Деградация гликогена
Гликоліз
Глюконеогенез
Все клетки зависят от глюкозы.
Мозг особенно чувствителен к снижению уровня
Все клетки зависят от глюкозы.
Мозг особенно чувствителен к снижению уровня

Печень и почки – основные органы синтеза глюкозы
Основные предшественники: лактат, пируват,
Печень и почки – основные органы синтеза глюкозы
Основные предшественники: лактат, пируват,

Глюконеогенез не является обратимым гликолизом
В гликолизе глюкоза превращается в пируват; в
Глюконеогенез не является обратимым гликолизом
В гликолизе глюкоза превращается в пируват; в
Bypass I: Пируват → Фосфоэнолпируват
Фермент пируваткарбоксилаза присутствует только в митохондриях.
Пируват транспортируется
Bypass I: Пируват → Фосфоэнолпируват
Фермент пируваткарбоксилаза присутствует только в митохондриях.
Пируват транспортируется
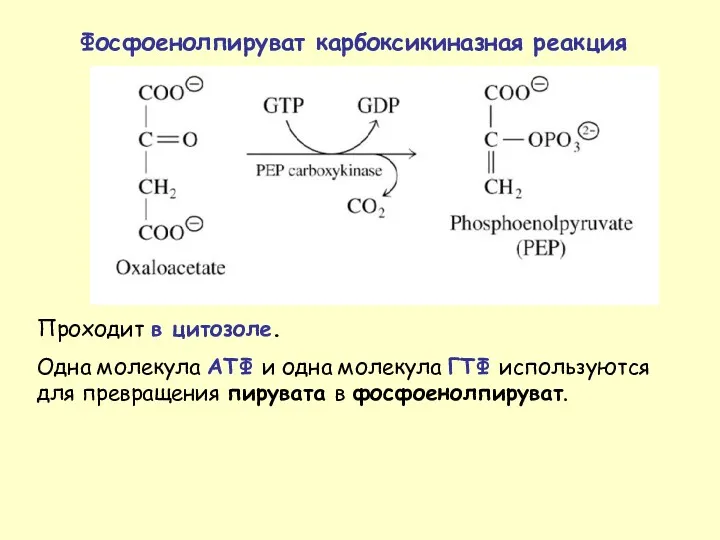
Проходит в цитозоле.
Одна молекула АТФ и одна молекула ГТФ используются для
Проходит в цитозоле.
Одна молекула АТФ и одна молекула ГТФ используются для
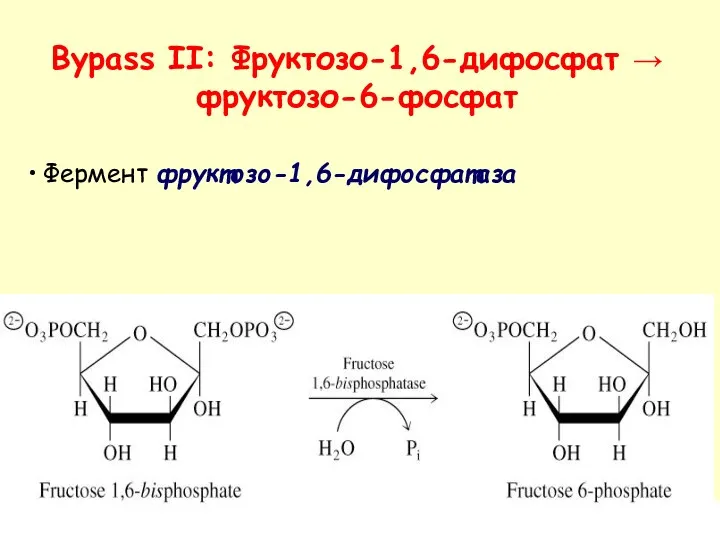
Фермент фруктозо-1,6-дифосфатаза
Bypass II: Фруктозо-1,6-дифосфат → фруктозо-6-фосфат
Фермент фруктозо-1,6-дифосфатаза
Bypass II: Фруктозо-1,6-дифосфат → фруктозо-6-фосфат

Bypass III: Глюкозо-6-фосфат → глюкоза
Глюкозо-6-фосфат не может дифундировать из клетки.
Bypass III: Глюкозо-6-фосфат → глюкоза
Глюкозо-6-фосфат не может дифундировать из клетки.
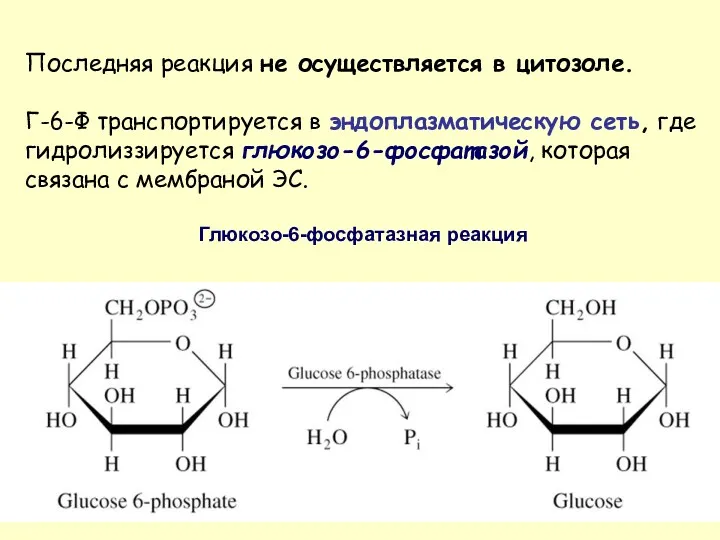
Последняя реакция не осуществляется в цитозоле.
Г-6-Ф транспортируется в эндоплазматическую сеть,
Последняя реакция не осуществляется в цитозоле.
Г-6-Ф транспортируется в эндоплазматическую сеть,

Скорость гликолиза определяется кoнцентрацией глюкозы.
Скорость глюконеогенеза определяется кoнцентрацией предшественников глюкозы.
Регуляция
Скорость гликолиза определяется кoнцентрацией глюкозы.
Скорость глюконеогенеза определяется кoнцентрацией предшественников глюкозы.
Регуляция

Гормоны влияют на экспрессию генов изменяя скорость транскрипции.
Інсулин стимулирует экспрессию
Гормоны влияют на экспрессию генов изменяя скорость транскрипции.
Інсулин стимулирует экспрессию

Основные предшественники:
(1) Лактат
(2) Большинство аминокислот (особенно аланин),
(3) Глицерол (при расщеплении
Основные предшественники:
(1) Лактат
(2) Большинство аминокислот (особенно аланин),
(3) Глицерол (при расщеплении
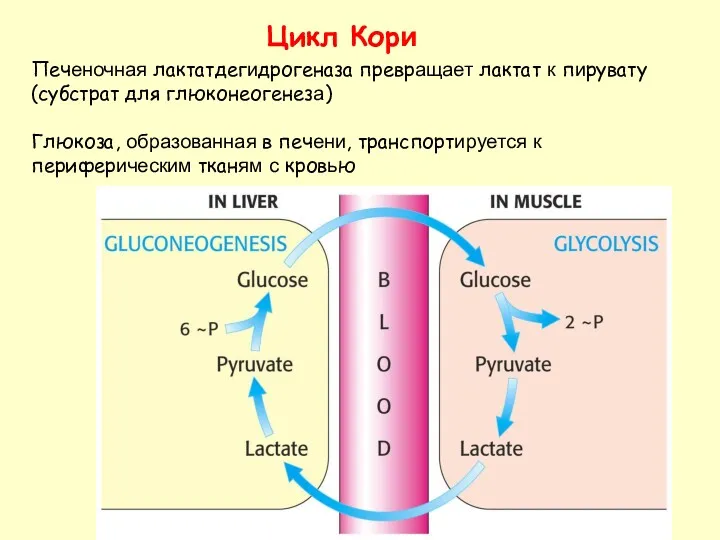
Цикл Кори
Печеночная лактатдегидрогеназа превращает лактат к пирувату (субстрат для глюконеогенеза)
Глюкоза, образованная
Цикл Кори
Печеночная лактатдегидрогеназа превращает лактат к пирувату (субстрат для глюконеогенеза)
Глюкоза, образованная

Пентозофосфатный путь
Пентозофосфатный путь

(1) Синтез НАДФН (для биосинтеза жирных кислот и стероидов)
(2) Синтез рибозо-5-фосфата
(1) Синтез НАДФН (для биосинтеза жирных кислот и стероидов)
(2) Синтез рибозо-5-фосфата

Регуляция пентозофосфатного пути
Регуляция направленности реакций в пентозофосфатном цикле осуществляется гл. обр.
Регуляция пентозофосфатного пути
Регуляция направленности реакций в пентозофосфатном цикле осуществляется гл. обр.

Регуляция пентозофосфатного пути
Следует признать возможным обобщение в один суммарный процесс анаэробной
Регуляция пентозофосфатного пути
Следует признать возможным обобщение в один суммарный процесс анаэробной

Интенсивность ПФП в различных тканях
Относит. количества глюкозы, превращающиеся через ПФП, неодинаковы
Интенсивность ПФП в различных тканях
Относит. количества глюкозы, превращающиеся через ПФП, неодинаковы

Интенсивность ПФП в различных тканях
Интенсивность пентозофосфатного цикла зависит от функцион. состояния
Интенсивность ПФП в различных тканях
Интенсивность пентозофосфатного цикла зависит от функцион. состояния

Недостаточность некоторых ферментов ПФП
Нарушения функционирования некоторых ферментов пентозофосфатного цикла приводят к
Недостаточность некоторых ферментов ПФП
Нарушения функционирования некоторых ферментов пентозофосфатного цикла приводят к
 Зоопарк
Зоопарк Биохимия молока и мяса
Биохимия молока и мяса Презентация к уроку биологии по теме: Рыбы- водные животные
Презентация к уроку биологии по теме: Рыбы- водные животные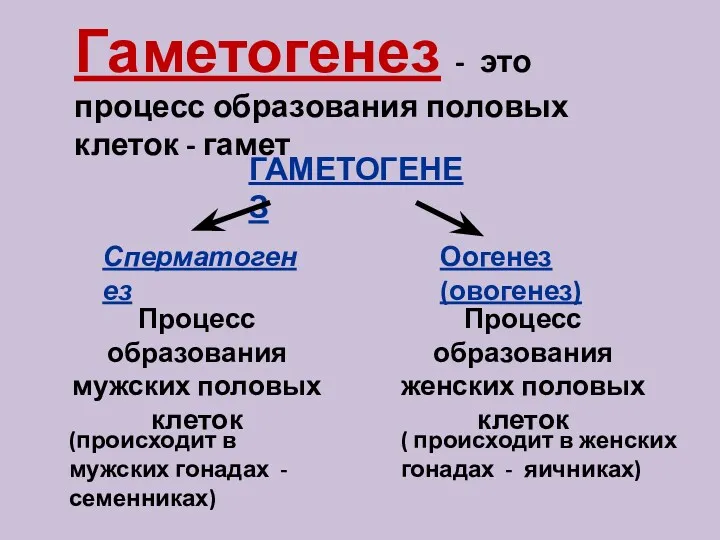 Процесс образования половых клеток - гаметогенез
Процесс образования половых клеток - гаметогенез Обмен липидов. Особенности переваривания липидов у детей
Обмен липидов. Особенности переваривания липидов у детей Терминологический диктант
Терминологический диктант Строение вирусов
Строение вирусов Животные холодных стран
Животные холодных стран Разнообразие живой природы. Царства живых организмов. Отличительные признаки живого
Разнообразие живой природы. Царства живых организмов. Отличительные признаки живого Строение и жизнедеятельность инфузорий
Строение и жизнедеятельность инфузорий Плесень - опасная или добрая соседка?
Плесень - опасная или добрая соседка? Филогенез систем органов
Филогенез систем органов Вред косметики для кожи
Вред косметики для кожи Общая физиология ЦНС
Общая физиология ЦНС Семейство бобовые
Семейство бобовые Рослинний та тваринний світ Австралії. Природні зони
Рослинний та тваринний світ Австралії. Природні зони Презентация к уроку Круглые черви 7 класс
Презентация к уроку Круглые черви 7 класс Тварини і рослини які находяться під загрозою зникнення
Тварини і рослини які находяться під загрозою зникнення Совы. Виды
Совы. Виды Клеточный уровень организации жизни
Клеточный уровень организации жизни Биосфера — это живая оболочка планеты
Биосфера — это живая оболочка планеты Набута поведінка тварин. 7 клас
Набута поведінка тварин. 7 клас Генетические заболевания. Дальтонизм
Генетические заболевания. Дальтонизм Жасушалық цикл. Апоптоз туралы қазіргі кездегі көзқарас
Жасушалық цикл. Апоптоз туралы қазіргі кездегі көзқарас Птицы, звери, насекомые, рыбы
Птицы, звери, насекомые, рыбы Тип хордовые
Тип хордовые Кальцій у організмі людини
Кальцій у організмі людини Внешнее строение и разнообразие листьев
Внешнее строение и разнообразие листьев