- Биосинтез углеводов в организме. Метаболизм гексоз. (Лекция 11)

Содержание
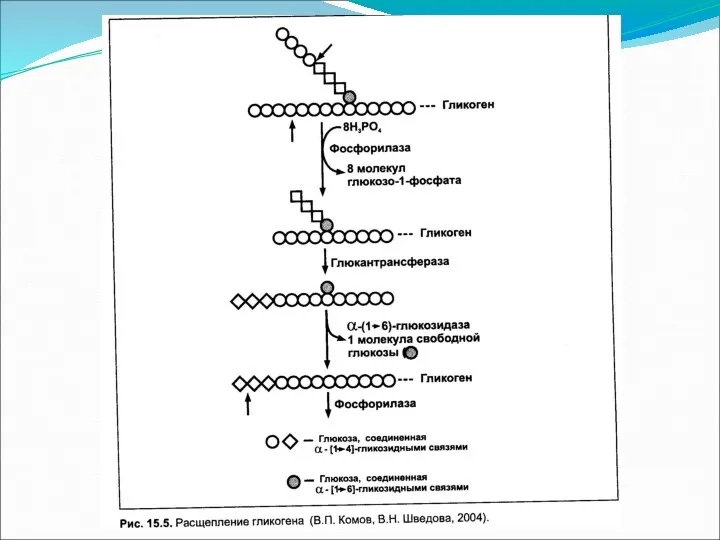
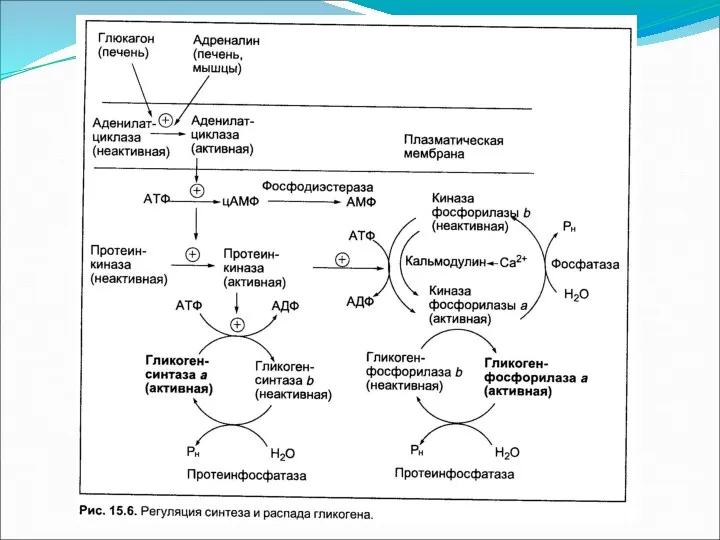
- 2. Гликоген является запасной формой углеводов в организме животных. Синтезируется главным образом в печени и составляет до
- 3. Синтез гликогена начинается с затраты молекулы АТФ для фосфорилирования глюкозы с образованием глюкозо-6-фосфата. Фосфорилирование свободной глюкозы
- 4. Изомеризация глюкозо-6-фосфата в глюкозо-1-фосфат Следующая реакция изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат катализируется ферментом фосфоглюкомутазой. Именно в таком
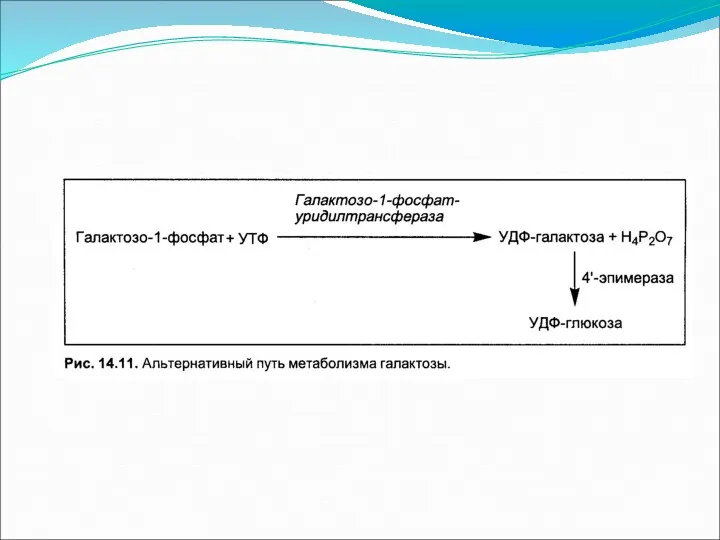
- 5. Образование УДФ-глюкозы В силу обратимости фосфоглюкомутазной реакции и чтобы синтез гликогена была термодинамически необратим, необходима дополнительная
- 6. Схема синтеза гликогена Перенос глюкозидного остатка, входящего в состав УДФ-глюкозы, идет на гликозидную затравку (праймер) гликогена.
- 8. Инициация синтеза гликогена гликогенином
- 18. Основное понятие Глюконеогенез – процесс синтеза глюкозы de novo из неуглеводных предшественников. Главная функция этого процесса
- 19. Образование оксалоацетата, его транспорт в цитозоль и превращение его в фосфоенолпируват Образование фосфоенолпирувата из пирувата происходит
- 20. Образование оксалоацетата из пирувата Митохондриальный фермент пируваткарбоксилаза, катализирующая данное превращение пирувата, в качестве кофермента содержит биотин.
- 21. Превращение оксалоацетата в малат Для оксалоацетата внутренняя мембрана митохондрий непроницаема, и транспорт оксалоацетата в цитоплазму клетки
- 22. В последующей реакции, катализируемой ферментом фосфоеноилпируваткарбоксикиназой из оксалоацетата образуется фосфоенолпируват. Реакция Mg-зависимая, и донором фосфата служит
- 23. Дефосфорилирование фруктозо-1,6-дифосфата После образования фосфоенолпирувата процесс глюконеогенеза идет по обратимым реакциям гликолиза вплоть до синтеза фруктозо-1,6-дифосфата.
- 24. Дефосфорилирование глюкозо-6-фосфата Образовавшийся фрукфозо-6-фосфат фосфоглюкоизомеразой переводится в глюкозо-6-фосфат, которая под действием глюкозо-6-фосфатазы (в процессе гликолиза этот
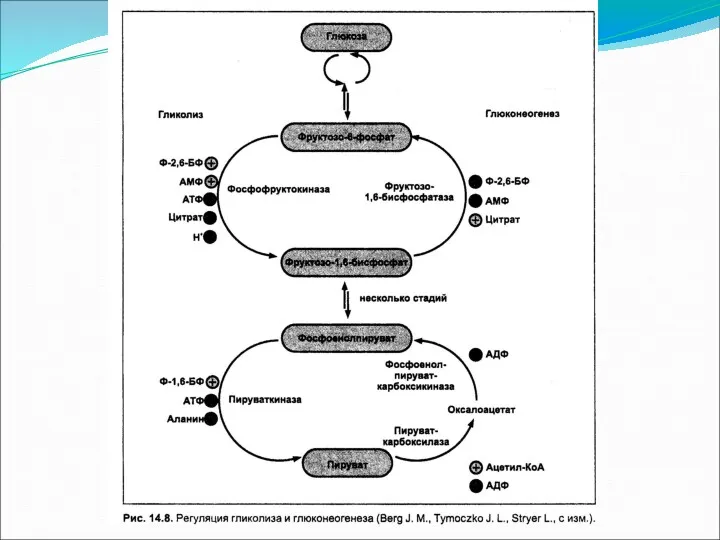
- 25. Механизмы регуляции глюконеогенеза Регуляторным ферментом в глюконеогенезе является пируваткарбоксилаза, которая активируется ацетил-КоА тогда, когда в митохондриях
- 28. Цикл Кори (глюкозолактатный цикл) Схема цикла Синтез глюкозы из лактата и превращение его в пируват есть
- 29. Метаболизм гексоз
- 34. Скачать презентацию
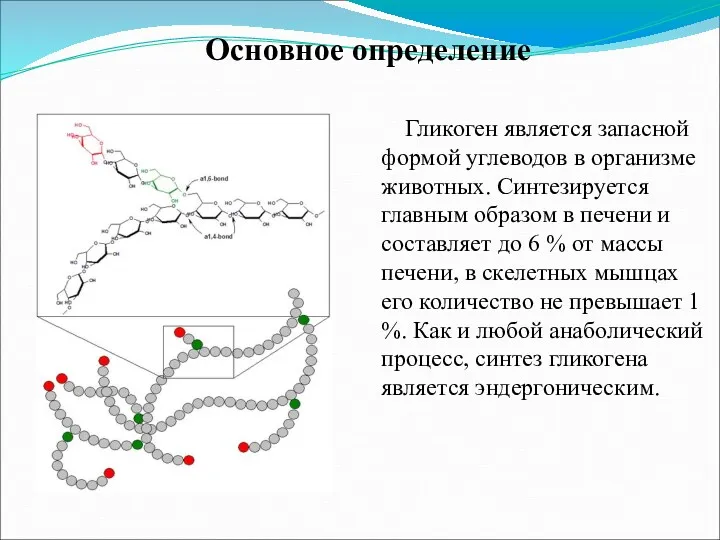
Гликоген является запасной формой углеводов в организме животных. Синтезируется главным
Гликоген является запасной формой углеводов в организме животных. Синтезируется главным
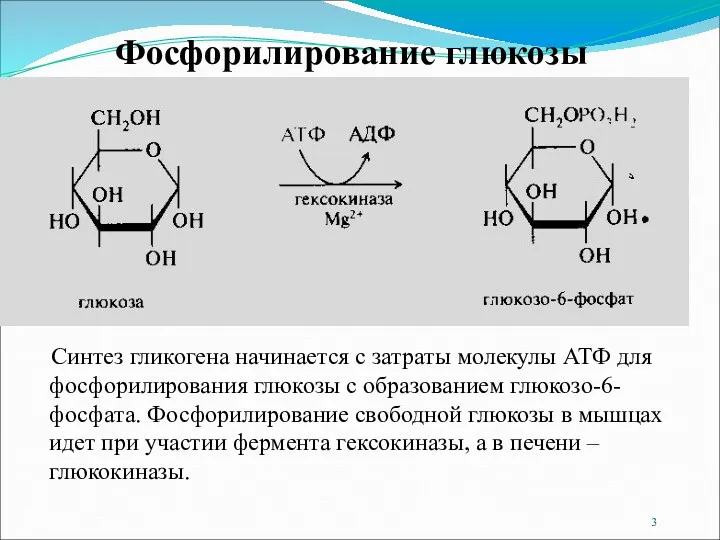
Синтез гликогена начинается с затраты молекулы АТФ для фосфорилирования глюкозы
Синтез гликогена начинается с затраты молекулы АТФ для фосфорилирования глюкозы
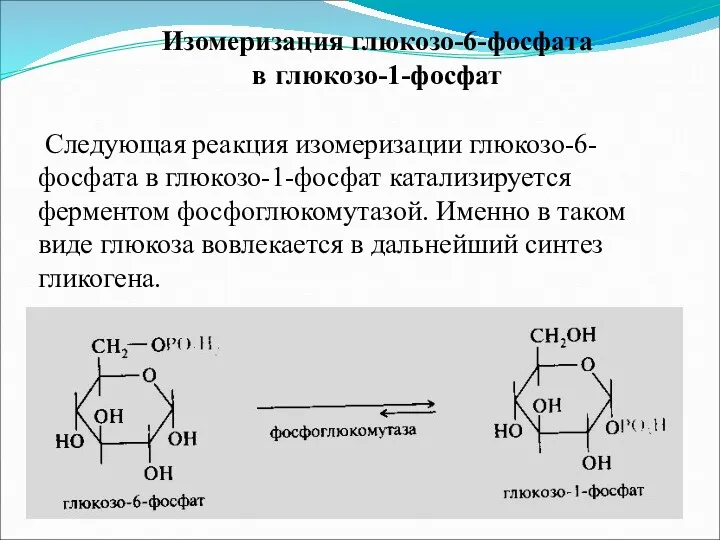
Изомеризация глюкозо-6-фосфата
в глюкозо-1-фосфат
Следующая реакция изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат катализируется
Изомеризация глюкозо-6-фосфата
в глюкозо-1-фосфат
Следующая реакция изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат катализируется
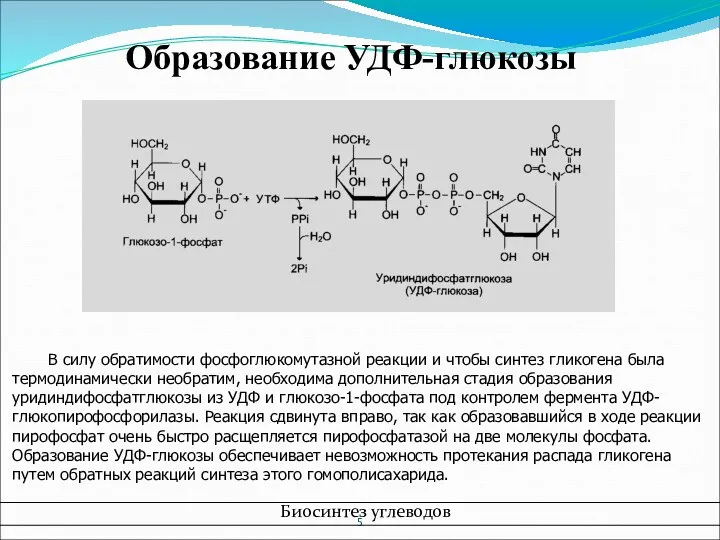
Образование УДФ-глюкозы
В силу обратимости фосфоглюкомутазной реакции и чтобы синтез гликогена была
Образование УДФ-глюкозы
В силу обратимости фосфоглюкомутазной реакции и чтобы синтез гликогена была

Схема синтеза гликогена
Перенос глюкозидного остатка, входящего
в состав УДФ-глюкозы, идет на гликозидную
Схема синтеза гликогена
Перенос глюкозидного остатка, входящего в состав УДФ-глюкозы, идет на гликозидную
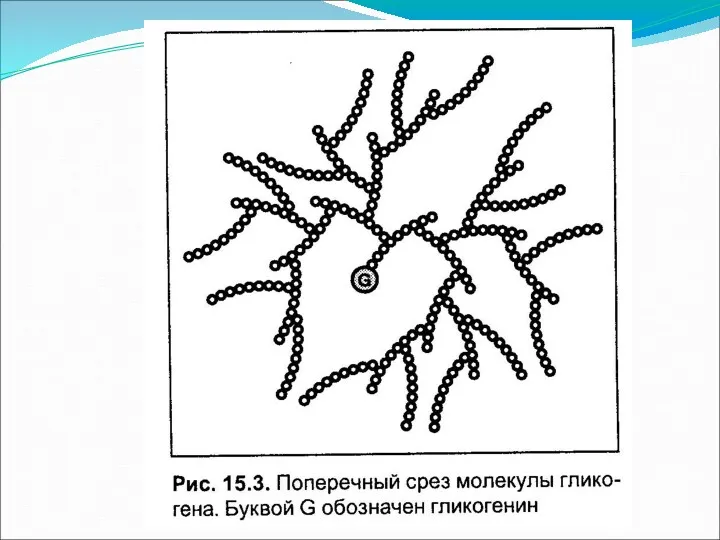
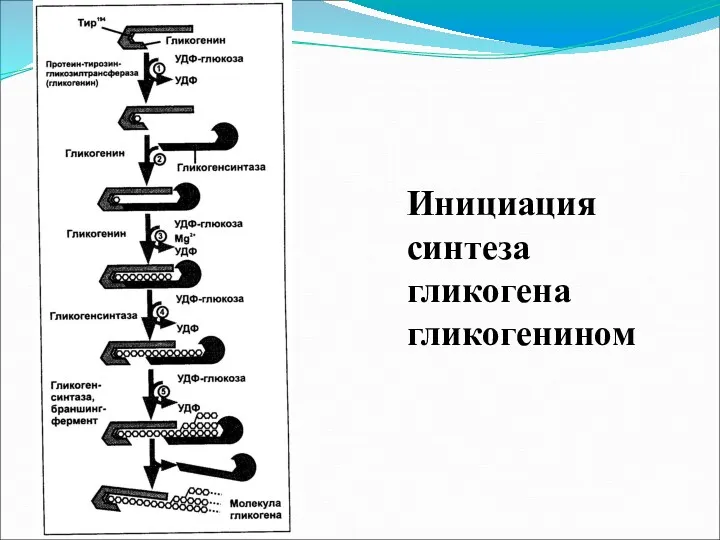
Инициация синтеза гликогена гликогенином
Инициация синтеза гликогена гликогенином
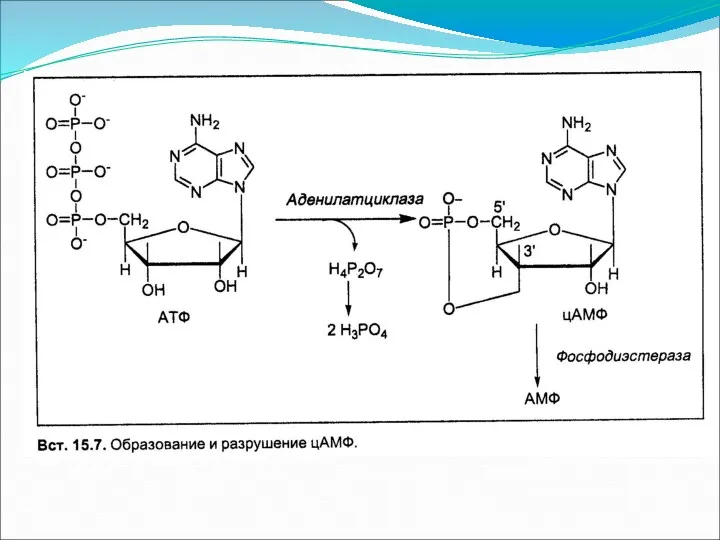
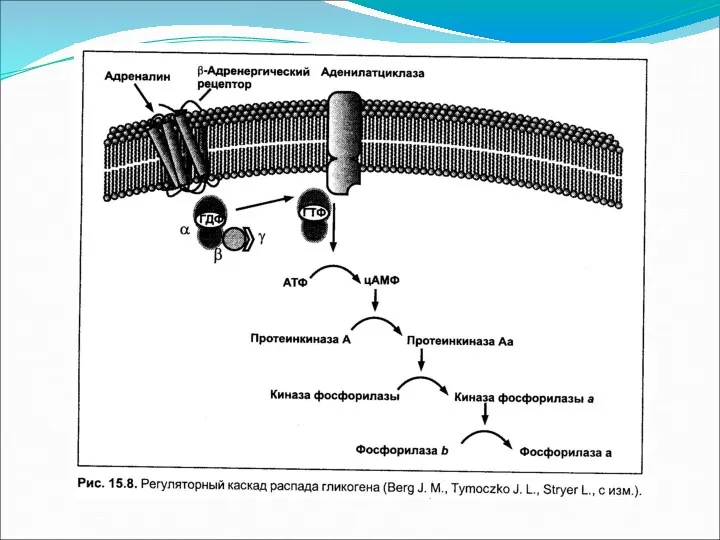
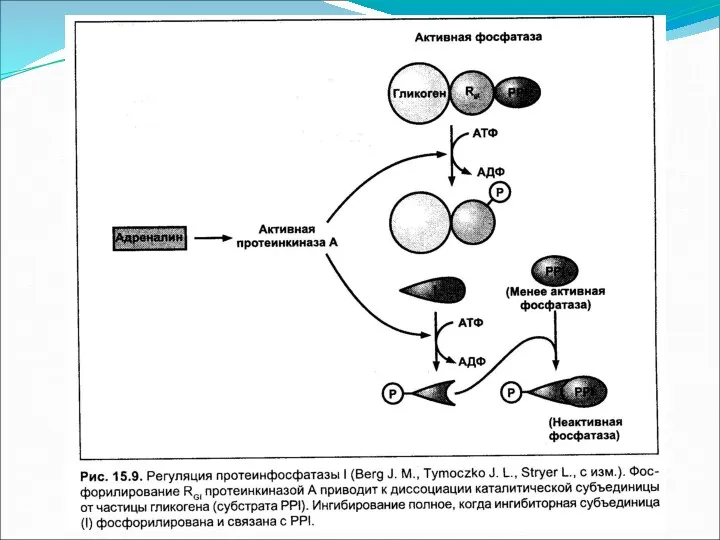
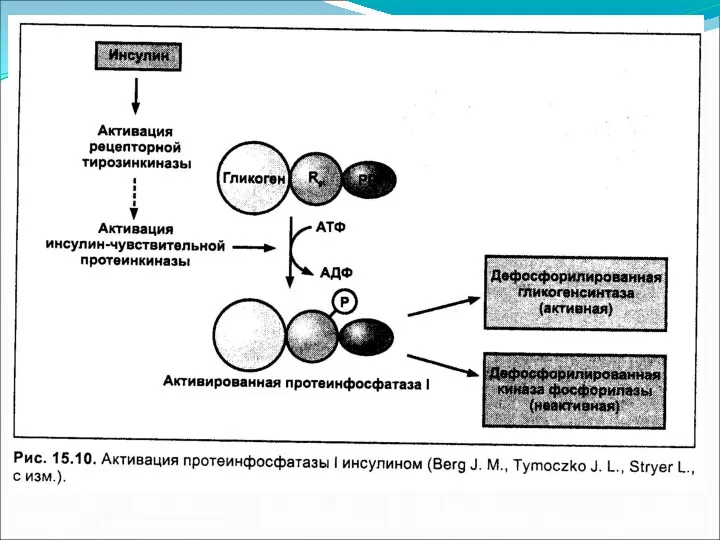
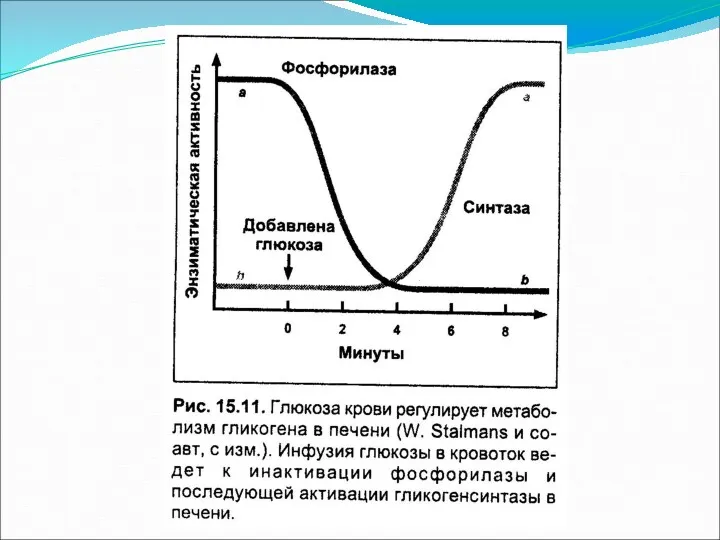
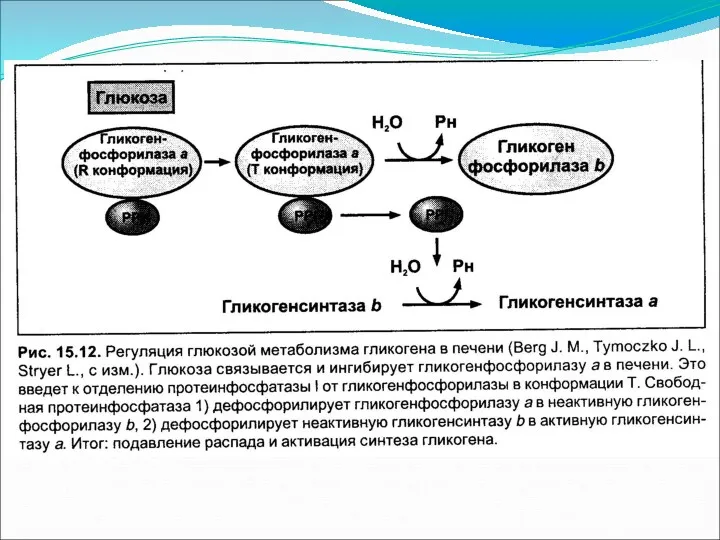


Основное понятие
Глюконеогенез – процесс синтеза глюкозы de novo из неуглеводных предшественников.
Основное понятие
Глюконеогенез – процесс синтеза глюкозы de novo из неуглеводных предшественников.
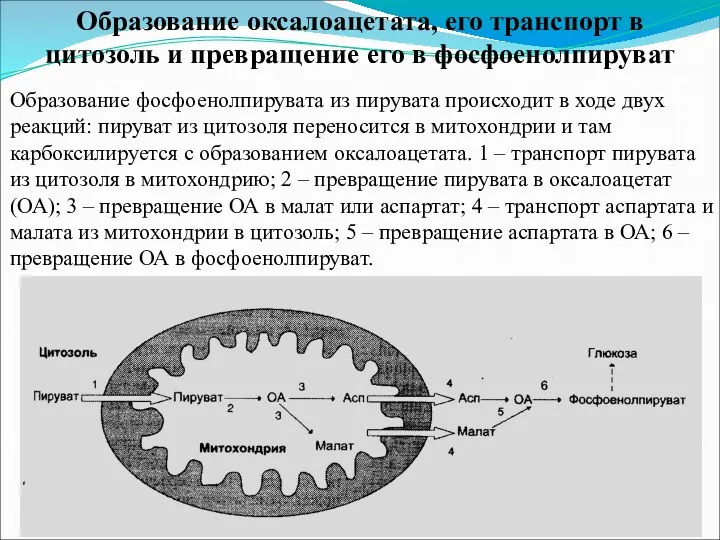
Образование оксалоацетата, его транспорт в цитозоль и превращение его в фосфоенолпируват
Образование
Образование оксалоацетата, его транспорт в цитозоль и превращение его в фосфоенолпируват
Образование
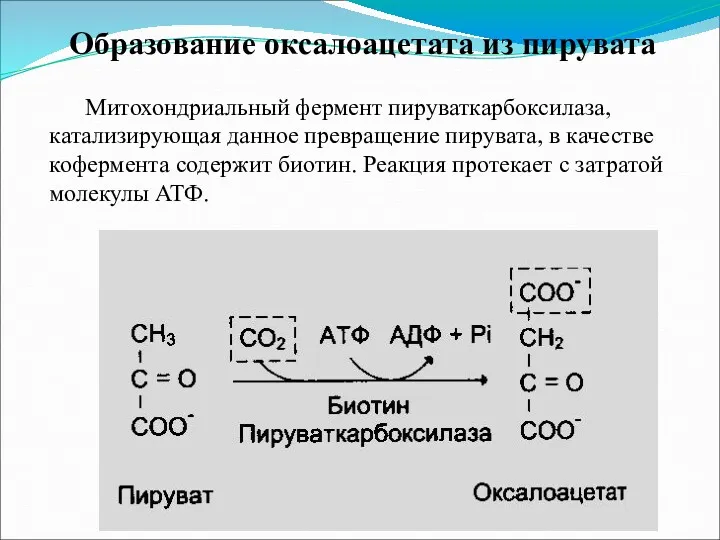
Образование оксалоацетата из пирувата
Митохондриальный фермент пируваткарбоксилаза, катализирующая данное превращение пирувата, в
Образование оксалоацетата из пирувата
Митохондриальный фермент пируваткарбоксилаза, катализирующая данное превращение пирувата, в
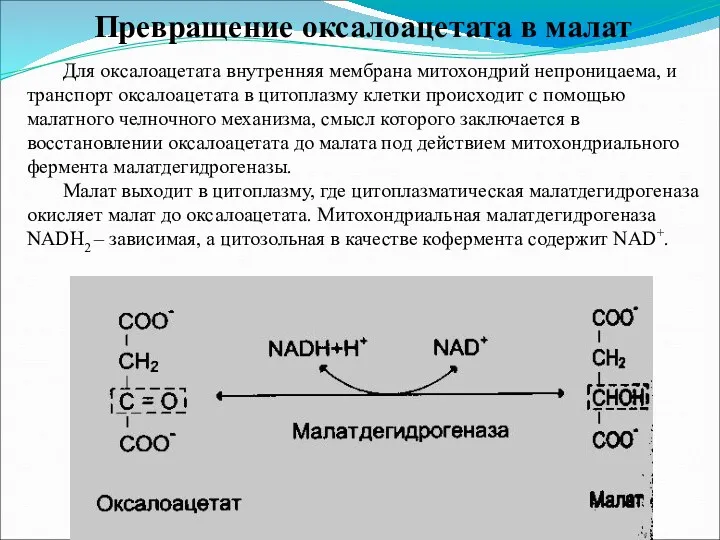
Превращение оксалоацетата в малат
Для оксалоацетата внутренняя мембрана митохондрий непроницаема, и транспорт
Превращение оксалоацетата в малат
Для оксалоацетата внутренняя мембрана митохондрий непроницаема, и транспорт
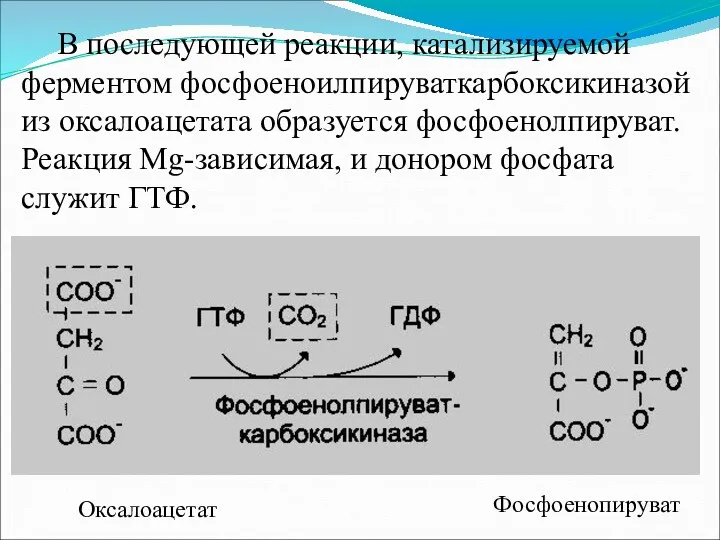
В последующей реакции, катализируемой ферментом фосфоеноилпируваткарбоксикиназой из оксалоацетата образуется фосфоенолпируват. Реакция
В последующей реакции, катализируемой ферментом фосфоеноилпируваткарбоксикиназой из оксалоацетата образуется фосфоенолпируват. Реакция

Дефосфорилирование фруктозо-1,6-дифосфата
После образования фосфоенолпирувата процесс глюконеогенеза идет по обратимым реакциям
Дефосфорилирование фруктозо-1,6-дифосфата
После образования фосфоенолпирувата процесс глюконеогенеза идет по обратимым реакциям
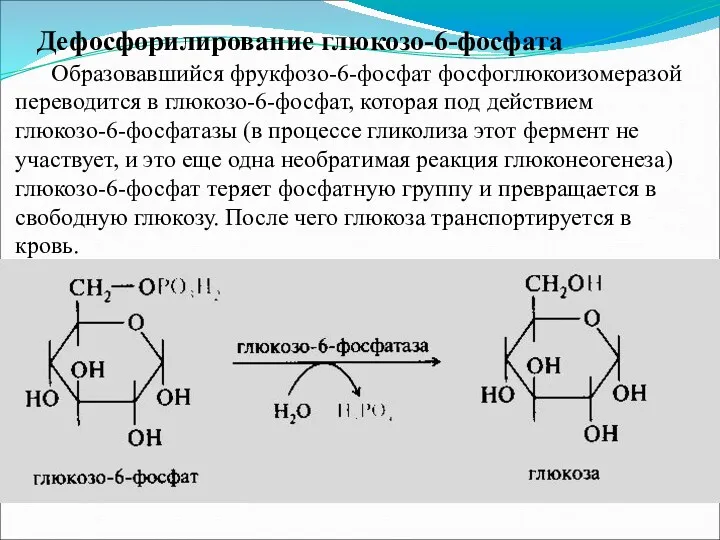
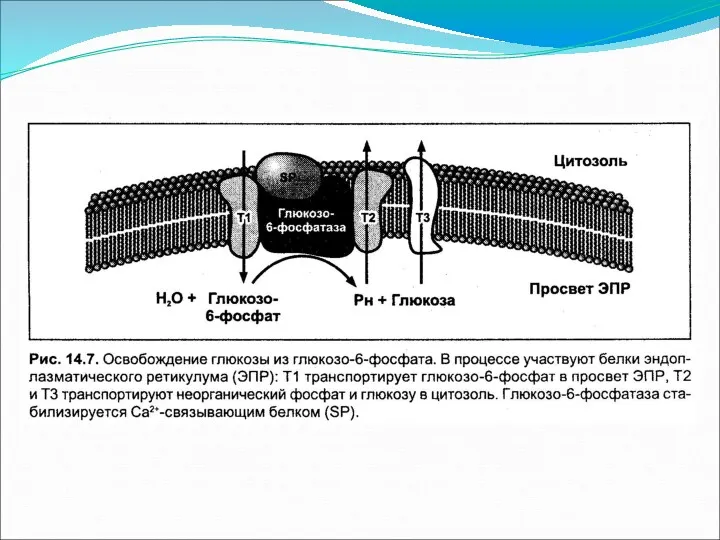
Дефосфорилирование глюкозо-6-фосфата
Образовавшийся фрукфозо-6-фосфат фосфоглюкоизомеразой переводится в глюкозо-6-фосфат, которая под действием глюкозо-6-фосфатазы
Дефосфорилирование глюкозо-6-фосфата
Образовавшийся фрукфозо-6-фосфат фосфоглюкоизомеразой переводится в глюкозо-6-фосфат, которая под действием глюкозо-6-фосфатазы

Механизмы регуляции глюконеогенеза
Регуляторным ферментом в глюконеогенезе является пируваткарбоксилаза, которая активируется ацетил-КоА
Механизмы регуляции глюконеогенеза
Регуляторным ферментом в глюконеогенезе является пируваткарбоксилаза, которая активируется ацетил-КоА
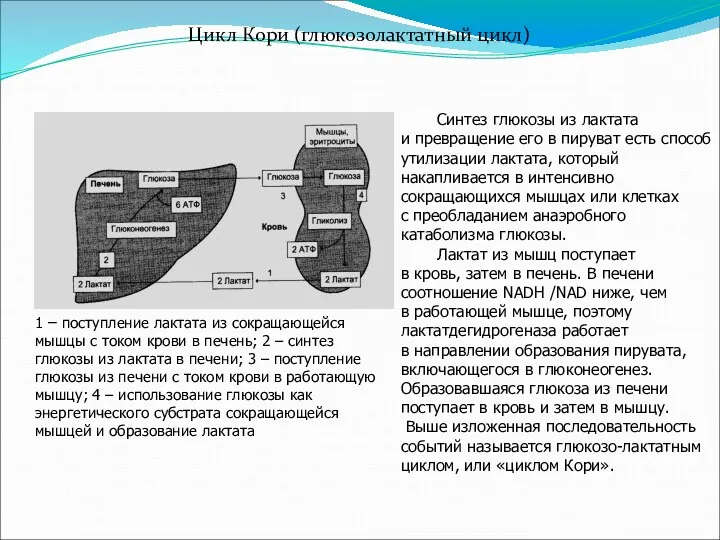
Цикл Кори (глюкозолактатный цикл)
Схема цикла
Синтез глюкозы из лактата
и превращение
Цикл Кори (глюкозолактатный цикл)
Схема цикла
Синтез глюкозы из лактата и превращение

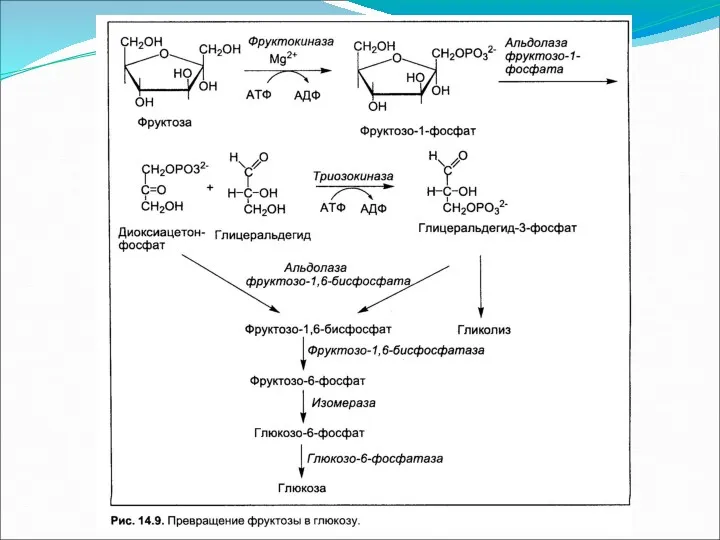
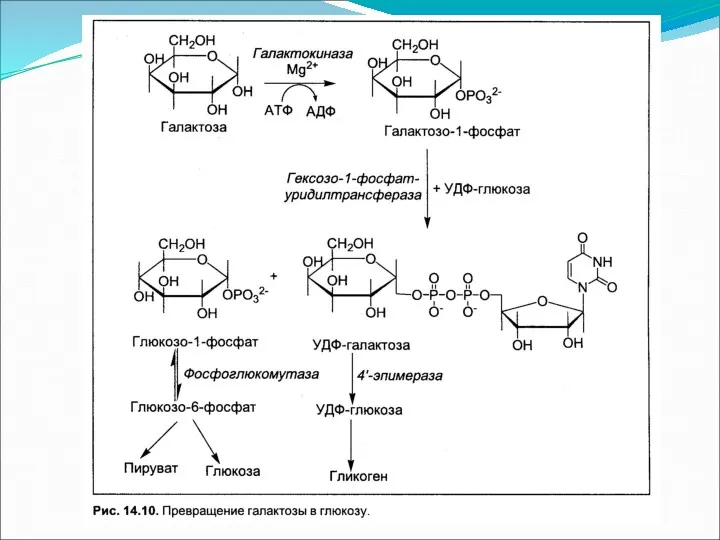
Метаболизм гексоз
Метаболизм гексоз
 Дыхание растений
Дыхание растений презентация к теме Эволюционное учение
презентация к теме Эволюционное учение Рецепция
Рецепция Классификацая органического мира
Классификацая органического мира Шеміршекті балықтар: акулалар және скаттар. Бекіре балықтар маңызы және оларды қалпына келтіру шаралары. Қостынысты балықтар
Шеміршекті балықтар: акулалар және скаттар. Бекіре балықтар маңызы және оларды қалпына келтіру шаралары. Қостынысты балықтар Мышцы шеи
Мышцы шеи Ноосфера. Вернадский В. И
Ноосфера. Вернадский В. И Школьная клумба
Школьная клумба Функциональная анатомия промежуточного мозга
Функциональная анатомия промежуточного мозга Гербарий КГПУ им. В.П. Астафьева
Гербарий КГПУ им. В.П. Астафьева Гаструляция. Дробление и гаструляция в кишечнодышащих
Гаструляция. Дробление и гаструляция в кишечнодышащих Тесты по биологии
Тесты по биологии Нутриенты для попугаев, при содержании в искусственных условиях
Нутриенты для попугаев, при содержании в искусственных условиях Өсімдіктердің стресс механизмдері
Өсімдіктердің стресс механизмдері ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Многобразие Простейших 5-7 класс.
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Многобразие Простейших 5-7 класс. Тірі ағзалардың көптүрлілігі
Тірі ағзалардың көптүрлілігі Растение в интерьере жилого дома
Растение в интерьере жилого дома Химический состав бактериальной клетки
Химический состав бактериальной клетки Хемосинтез (автотрофное питание)
Хемосинтез (автотрофное питание) Строение и функции стебля
Строение и функции стебля Оживление грибов дрожжей
Оживление грибов дрожжей Аграрний сервіс майбутнього
Аграрний сервіс майбутнього Квантовые методы в медицине. Ядерный магнитный резонанс. Электронный парамагнитный резонанс. (Лекция 14)
Квантовые методы в медицине. Ядерный магнитный резонанс. Электронный парамагнитный резонанс. (Лекция 14) Презентация по биологии для учащихся 8 класса на тему: Гигиена пищеарения
Презентация по биологии для учащихся 8 класса на тему: Гигиена пищеарения Органы пищеварения
Органы пищеварения Своя игра Общие сведения о мире животных
Своя игра Общие сведения о мире животных Класс Пресмыкающиеся, или рептилии
Класс Пресмыкающиеся, или рептилии Липиды.
Липиды.