- Біоінформатика. (Тема 1)

Содержание
- 2. Лавиноподібне накопичення даних молекулярної та структурної біології, яке відбувається протягом останніх 20 років, кардинальним чином змінило
- 3. «Омік»-технології дозволяють генерувати та оперувати даними в надзвичайно широкому діапазоні, починаючи з досліджень цілих геномів з
- 4. Вузьке місце в біологічних науках зсунулося з отримання первинних результатів до їх зберігання, препроцесінгу, аналізу та
- 5. накопичення великої кількості біологічних даних стимульовало розвиток особливої наукової дисціпліни, що дозволяє інтегрувати і обробляти їх
- 6. множинність визначення біоінформатики вся сукупність методів обчислювальної біології (синоніми – обчислювальна біологія, інформаційна біологія) сукупність програм
- 7. приклад розгорнутого визначення (за Altman, 1998) Біоінформатика досліджує два інформаційних потоки в молекулярній біології: передачу інформації
- 8. власне визначення ;) біоінформатика – спроба інтерпретації біологічних “текстів”, прикладом яких є послідовності макромолекул в живих
- 9. будь-які визначення біоінформатики як правило охоплюють застосування комп’ютерних наближень на рівні не вище клітинного
- 10. основні розділи біоінформатики області інтересу комп’ютерних фахівців в біології біоінформатика послідовностей – класична біоінформатика структурна біоінформатика
- 11. біоінформатика послідовностей – класична біоінформатика
- 13. Статистика надходжень нуклеотидних послідовностей в GenBank На момент свого заснування в 1982 році містив 606 послідовностей,
- 14. Статистика надходжень нуклеотидних послідовностей в GenBank (2007)
- 15. Статистика надходжень нуклеотидних послідовностей в GenBank (2012) детальна статистика доступна за адресою ftp://ftp.ncbi.nih.gov/genbank/gbrel.txt
- 16. Дані щодо послідовностей – розвиток алгоритмів для парного та множинного вирівнювання послідовностей, визначення та дослідження мотивів,
- 17. точки застосування класичної біоінформатики Вирівнювання й визначення подібності двох послідовностей Побудова множинних вирівнювань Розпізнавання генів Передбачення
- 18. структурна біоінформатика – обчислювальна структурна біологія
- 19. структурна біоінформатика з точки зору біоінформатики – підрозділ біоінформатики, що фокусується на представленні, зберіганні, запиті, аналізі
- 20. Статистика надходжень до Міжнародного банка білкових структур (PBD – Protein Data Bank). Синім кольором виділено щорічні
- 21. + структурна біоінформатика біоінформатика структурна біологія біофізика (метод) + цитологія (предмет) молекулярна біологія
- 22. Структурні дані – розвиток обчислювальної геометрії , комп’ютерної графіки, алгоритмів для аналізу кристалографічних даних та даних
- 23. структурна біоінформатика Більш глибоке розуміння, як біологічна функція обумовлена просторовою структурою. Чи можна передбачити просторову структуру,
- 24. задачі структурної біоінформатики класифікація білків за особливостями просторової структури, аналіз та/або передбачення активних сайтів оцінка якості
- 25. точки застосування структурної біоінформатики вибір білків-мішеней трекінг умов кристалізації аналіз кристалографічних даних аналіз даних ЯМР анотування
- 26. труднощі структурно- біоінформатичних обчислень структурні дані є нелінійними, взаємодії між атомами також нелінійні – необхідність використання
- 27. труднощі структурно- біоінформатичних обчислень візуалізація даних – одночасно превага і недолік: вона спрямована на людину і
- 28. комп’ютерна геноміка
- 29. Обчислювальна геноміка фокусується (як цілком зрозуміло з назви) на розмітці та порівняльному аналізі організації геномів різного
- 30. точки застосування комп’ютерної геноміки Передбачення генів у послідовностях. При цьому в деяких випадках вдається навіть знайти
- 31. Задача метапболічної реконструкції є спільною як для обчислювальної геноміки, так і для системної біології – науки,
- 32. структурна біоінформатика – основа структурної геноміки Структурна геноміка – високопропускне визначення просторової структури макромолекул (в першу
- 33. Нова область інтересу – аналіз даних експресії. Необхідність обробки вельми зашумлених даних спричинила розвиток відповідних алгоритмів
- 37. При аналізі первинних структур процедура вирівнювання виявляє сходство між послідовностями (sequence similarity), яке може свідчити про
- 38. Гомологичные последовательности – последовательности, имеющие общее происхождение (общего предка). Признаки гомологичности белков сходная 3D-структура в той
- 39. Что изображено? Название последовательности Номер столбца выравнивания Номер последнего в строке остатка ИЗ ЭТОЙ ПОСЛЕДОВАТЕЛЬНОСТИ Консервативный
- 40. «Идеальное» выравнивание – запись последовательностей одна под другой так, чтобы гомологичные фрагменты оказались друг под другом.
- 42. Паттерн – регулярное выражение UNIX’a: [AC]-x-V-x(4)-{ED} Ala или Cys- х-Val- х- х- х - х- (любой,
- 43. Профиль или весовая матрица (PSSM) F K L L S H C L L V F
- 44. Правильно ли выровнены последовательности?
- 45. В чем биологический смысл выравнивания? Буквы в одной колонке определяют сопоставление аминокислотных остатков двух белков Сопоставленные
- 46. Какое выравнивание “правильнее”? 13 “консервативных” остатков 12 консервативных остатков
- 47. Чтобы понять смысл выравнивания, вернемся к тому, что такое последовательность аминокислотных остатков и что такое белок
- 48. (i)Последовательность – удобный способ закодировать структурную (химическую) формулу молекулы белка (до посттрансляционных модификаций) (iii) Последовательность однозначно
- 49. Пространственное совмещение полипептидных цепей белков mta1_yeast и mat2_yeast На плоской картинке видно плохо ☹
- 50. Схематическое изображение совмещенных структур
- 51. Другой способ отобразить совмещение полипептидных цепей называется структурным выравниванием последовательностей Стрелки как на предыдущем слайде
- 52. Совмещение структур и выравнивание последовательностей
- 53. Еще раз: разметка по совмещенным структурам
- 54. Биологически обоснованное выравнивание гомеодоменов
- 55. Совмещение 5-и гомеодоменов
- 56. Множественное выравнивание гомеодоменов Красным выделены консервативные (одинаковые у всех) остатки; желтым – на 80% консервативные (одинаковые
- 57. Размеченное множественное выравнивание
- 58. Функции аминокислотных остатков Trp48 Arg53 Leu16 Pro442/ Lys442
- 59. В “правильном” выравнивании много консервативных аминокислотных остатков и функционально консервативных позиций
- 60. Выравнивание и эволюция Последовательности белка оболочки из двух штаммов вируса Коксаки
- 61. .. Последовательности белка оболочки из двух штаммов вируса Коксаки и энтеровируса человека
- 62. Аминокислотные остатки в одной колонке биологически обоснованного выравнивания, как правило, “произошли” из одного и того же
- 63. ПРОБЛЕМА: как построить “правильное” выравнивание последовательностей белков если структуры белков неизвестны?
- 64. На сегодня известны: более 10 млн(!!!) последовательностей белков (включая фрагменты и трансляты) пространственные структуры около 65
- 65. Алгоритмические решения проблемы воплощены в программах Программы выравнивания последовательностей тестируются путем сравнения с биологически обоснованными –
- 66. Предположим, известны структуры родственных белков и, значит, биологически обоснованное выравнивание последовательностей При > 60% совпадающих букв
- 67. (*) Справедливы ли положения с предыдущего слайда для выравнивания последовательностей ДНК? последовательностей РНК?
- 68. 2 основних підходи до відтворення просторової структури білка in silico моделювання за гомологією конформаційний пошук
- 69. Утворення тривимірної структури білка in vivo відбувається при біосинтезі або відразу після нього. Чудово, проте, що
- 70. 1999 рік – Rost B. Twilight zone of protein sequence alignment
- 71. схема залежності енергії молекули від її конформації
- 72. Для тубулінів будь-якого походження є характерним явище специфічної взаємодії з низькомолекулярними і не тільки органічними речовинами
- 73. Незважаючи на високу консервативність структури тубулінів різного походження, рослинні тубуліни характеризуються наявністю унікальних властивостей. Насамперед це
- 74. Просторова структура та розподіл електростатичного потенціалу на поверхні представників динітроанілінів (а - трифлюралін, б - орізалін,
- 75. Порівняльне вирівнювання послідовностей тубулінів рослинного (Eleusine indica) та тваринного (Sus scrofa) походження. Вівень тотожності послідовностей складає
- 76. Відсутність досліджень особливостей просторової структури рослинних тубулінів труднощі технологічного характеру при отримані рослинних тубулінів із ступенем
- 77. Стереозображення тривимірної упаковки молекул α-і β-тубулінів Eleusine indica та γ -тубуліну Arabidopsis thaliana
- 78. Фундаментальною особливістю тубулінів є явно виражена метастабільність елементів вторинної структури у часі – явище, яке характеризується
- 79. Діаграма розташування елементів вторинної структури в молекулах α, β та γ-тубуліну рослин на ділянці з 1
- 80. Вид молекулярної поверхні α-тубуліну з боку інтердимерного контакту: а – розподіл електростатичного потенціалу на молекулярній поверхні,
- 81. Особливості рельєфу поверхні та розподілу електростатичного потенціалу в області сайту взаємодії α-тубуліну E. indica з динітроаніліновими
- 82. Мутація Met–>Thr в позиції 268 рослинного α-тубуліну, яка викликає виникнення проміжної стійкості до динітроанілінових гербіцидів, співпадає
- 83. Порівняльний аналіз послідовностей рослинних α-тубулінів Представлено ділянки послідовностей, що безпосередньо прилягають до амінокислотних залишків, для яких
- 84. Карти молекулярної поверхні рослинних тубулінів в області, що відповідає сайту зв’язування на поверхні α-тубуліна. α-тубулін –
- 85. Распознавание генов Поиск открытых рамок считывания Использование статистики (отличия белок-кодирующих и некодирующих областей) Идентификация начал генов
- 86. Ортологи и паралоги Ортологи – гени з різних організмів, що розійшлися при видоутворенні. Мається на увазі,
- 87. Регуляторні послідовності в геномі бактерій
- 88. Регуляторні послідовності в геномі бактерій
- 89. Цель (глобальная) Предсказать свойства организма путем (компьютерного) анализа его генома (возможно, с использованием дополнительной информации: эпигенетика,
- 90. «Неприкладная» биоинформатика Молекулярная эволюция филогения генов таксономия организмов горизонтальные переносы и т.п. положительный и отрицательный отбор
- 91. Задачи С проверяемым ответом предсказание функции, регуляции, структуры и т.п.: ставим эксперимент С непроверяемым ответом эволюционные
- 92. «В принципе не проверяемые ответы» (зависящие от определений) Так ли они непроверяемы? Повторы если иметь все
- 93. Цель (недостижимая?) откуда оно все взялось? первое приближение – реконструкция генома/свойств реально ли заглянуть глубже? реально
- 95. Скачать презентацию

Лавиноподібне накопичення даних молекулярної та структурної біології, яке відбувається протягом останніх
Лавиноподібне накопичення даних молекулярної та структурної біології, яке відбувається протягом останніх

«Омік»-технології дозволяють генерувати та оперувати даними в надзвичайно широкому діапазоні, починаючи
«Омік»-технології дозволяють генерувати та оперувати даними в надзвичайно широкому діапазоні, починаючи

Вузьке місце в біологічних науках зсунулося з отримання первинних результатів до
Вузьке місце в біологічних науках зсунулося з отримання первинних результатів до

накопичення великої кількості біологічних даних стимульовало розвиток особливої наукової дисціпліни, що
накопичення великої кількості біологічних даних стимульовало розвиток особливої наукової дисціпліни, що

множинність визначення
біоінформатики
вся сукупність методів обчислювальної біології (синоніми – обчислювальна біологія, інформаційна
множинність визначення
біоінформатики
вся сукупність методів обчислювальної біології (синоніми – обчислювальна біологія, інформаційна

приклад розгорнутого визначення (за Altman, 1998)
Біоінформатика досліджує два інформаційних потоки в
приклад розгорнутого визначення (за Altman, 1998)
Біоінформатика досліджує два інформаційних потоки в

власне визначення ;)
біоінформатика – спроба інтерпретації біологічних “текстів”, прикладом яких є
власне визначення ;)
біоінформатика – спроба інтерпретації біологічних “текстів”, прикладом яких є

будь-які визначення біоінформатики як правило охоплюють застосування комп’ютерних наближень
на рівні
будь-які визначення біоінформатики як правило охоплюють застосування комп’ютерних наближень
на рівні

основні розділи біоінформатики
області інтересу комп’ютерних фахівців в біології
біоінформатика послідовностей –
основні розділи біоінформатики
області інтересу комп’ютерних фахівців в біології
біоінформатика послідовностей –

біоінформатика послідовностей – класична біоінформатика
біоінформатика послідовностей – класична біоінформатика
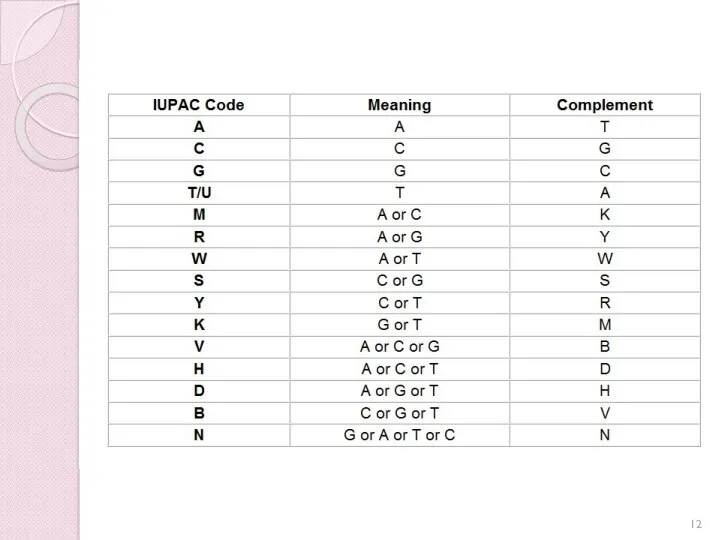

Статистика надходжень нуклеотидних послідовностей в GenBank
На момент свого заснування в 1982
Статистика надходжень нуклеотидних послідовностей в GenBank
На момент свого заснування в 1982
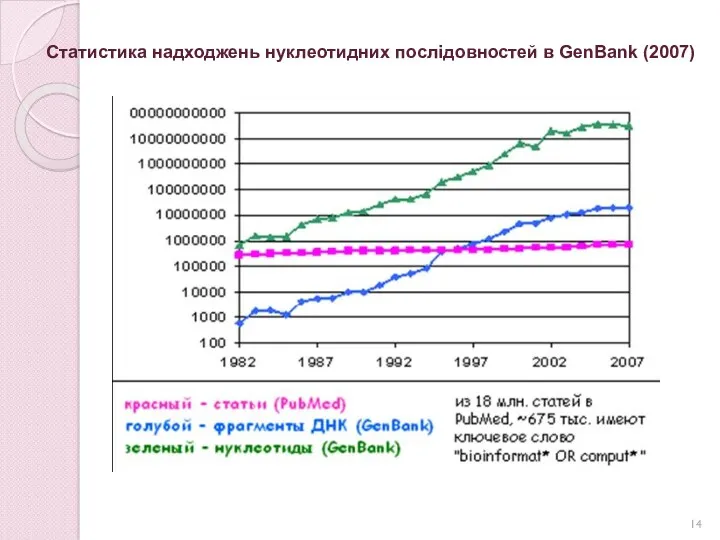
Статистика надходжень нуклеотидних послідовностей в GenBank (2007)
Статистика надходжень нуклеотидних послідовностей в GenBank (2007)
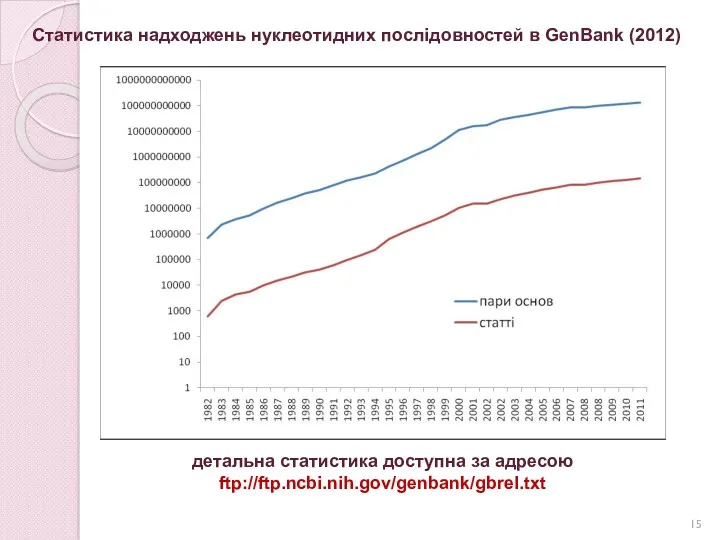
Статистика надходжень нуклеотидних послідовностей в GenBank (2012)
детальна статистика доступна за адресою
Статистика надходжень нуклеотидних послідовностей в GenBank (2012)
детальна статистика доступна за адресою

Дані щодо послідовностей – розвиток алгоритмів для парного та множинного вирівнювання
Дані щодо послідовностей – розвиток алгоритмів для парного та множинного вирівнювання

точки застосування класичної біоінформатики
Вирівнювання й визначення подібності двох послідовностей
Побудова множинних вирівнювань
Розпізнавання
точки застосування класичної біоінформатики
Вирівнювання й визначення подібності двох послідовностей
Побудова множинних вирівнювань
Розпізнавання

структурна біоінформатика – обчислювальна структурна біологія

структурна біоінформатика
з точки зору біоінформатики – підрозділ біоінформатики, що фокусується на
структурна біоінформатика
з точки зору біоінформатики – підрозділ біоінформатики, що фокусується на
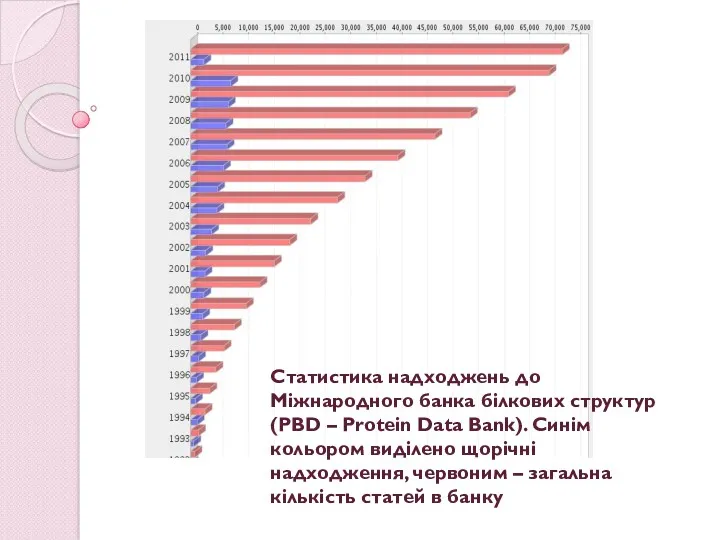
Статистика надходжень до Міжнародного банка білкових структур (PBD – Protein Data
Статистика надходжень до Міжнародного банка білкових структур (PBD – Protein Data
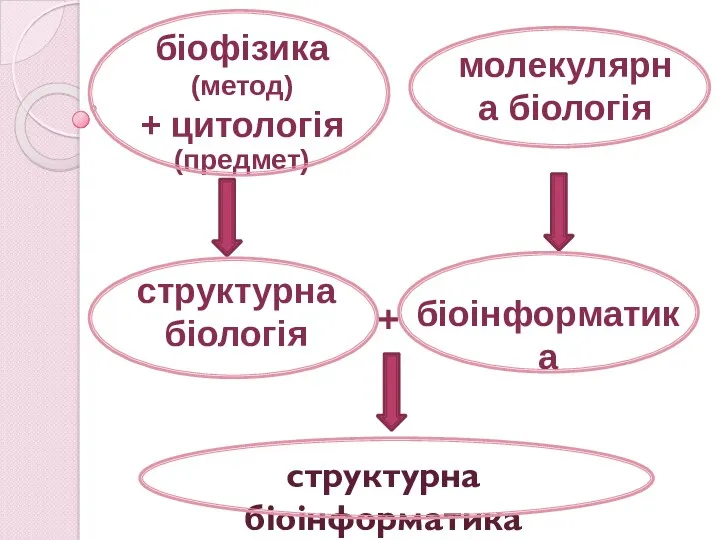
+
структурна біоінформатика
біоінформатика
структурна біологія
біофізика
(метод)
+ цитологія (предмет)
молекулярна біологія
+
структурна біоінформатика
біоінформатика
структурна біологія
біофізика
(метод)
+ цитологія (предмет)
молекулярна біологія
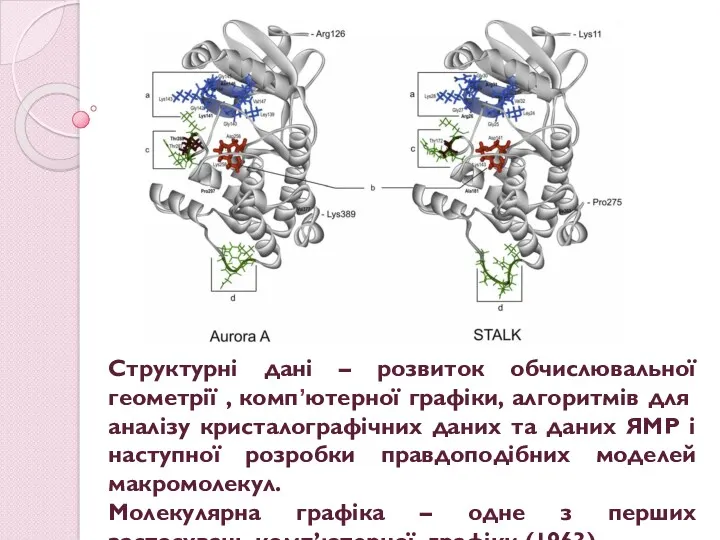
Структурні дані – розвиток обчислювальної геометрії , комп’ютерної графіки, алгоритмів для
Структурні дані – розвиток обчислювальної геометрії , комп’ютерної графіки, алгоритмів для

структурна біоінформатика
Більш глибоке розуміння, як біологічна функція обумовлена просторовою структурою.
структурна біоінформатика
Більш глибоке розуміння, як біологічна функція обумовлена просторовою структурою.

задачі структурної біоінформатики
класифікація білків за особливостями просторової структури, аналіз та/або передбачення
задачі структурної біоінформатики
класифікація білків за особливостями просторової структури, аналіз та/або передбачення

точки застосування структурної біоінформатики
вибір білків-мішеней
трекінг умов кристалізації
аналіз кристалографічних даних
аналіз даних ЯМР
анотування
точки застосування структурної біоінформатики
вибір білків-мішеней
трекінг умов кристалізації
аналіз кристалографічних даних
аналіз даних ЯМР
анотування

труднощі структурно- біоінформатичних обчислень
структурні дані є нелінійними, взаємодії між атомами також
труднощі структурно- біоінформатичних обчислень
структурні дані є нелінійними, взаємодії між атомами також

труднощі структурно- біоінформатичних обчислень
візуалізація даних – одночасно превага і недолік: вона
труднощі структурно- біоінформатичних обчислень
візуалізація даних – одночасно превага і недолік: вона

комп’ютерна геноміка

Обчислювальна геноміка фокусується (як цілком зрозуміло з назви) на розмітці та
Обчислювальна геноміка фокусується (як цілком зрозуміло з назви) на розмітці та

точки застосування комп’ютерної геноміки
Передбачення генів у послідовностях. При цьому в деяких
точки застосування комп’ютерної геноміки
Передбачення генів у послідовностях. При цьому в деяких

Задача метапболічної реконструкції є спільною як для обчислювальної геноміки, так і
Задача метапболічної реконструкції є спільною як для обчислювальної геноміки, так і

структурна біоінформатика – основа структурної геноміки
Структурна геноміка – високопропускне визначення просторової
структурна біоінформатика – основа структурної геноміки
Структурна геноміка – високопропускне визначення просторової

Нова область інтересу – аналіз даних експресії. Необхідність обробки вельми зашумлених
Нова область інтересу – аналіз даних експресії. Необхідність обробки вельми зашумлених

При аналізі первинних структур процедура вирівнювання виявляє сходство між послідовностями (sequence
При аналізі первинних структур процедура вирівнювання виявляє сходство між послідовностями (sequence

Гомологичные последовательности – последовательности, имеющие общее происхождение (общего предка).
Признаки гомологичности
Гомологичные последовательности – последовательности, имеющие общее происхождение (общего предка).
Признаки гомологичности
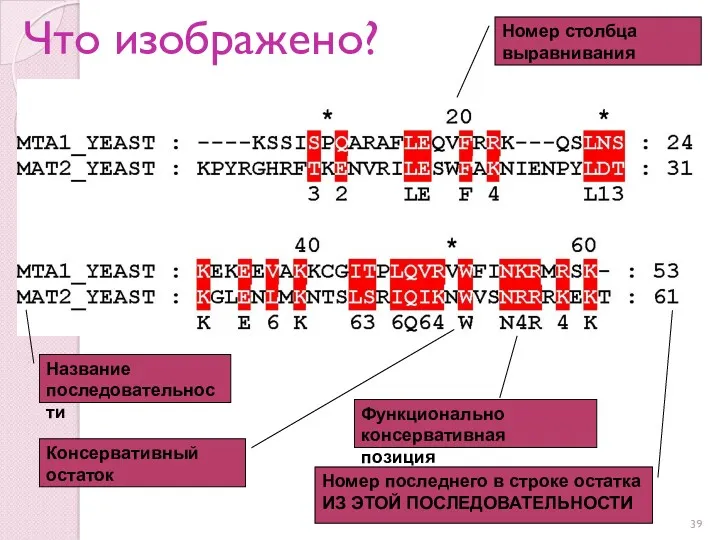
Что изображено?
Название последовательности
Номер столбца выравнивания
Номер последнего в строке остатка ИЗ ЭТОЙ
Что изображено?
Название последовательности
Номер столбца выравнивания
Номер последнего в строке остатка ИЗ ЭТОЙ

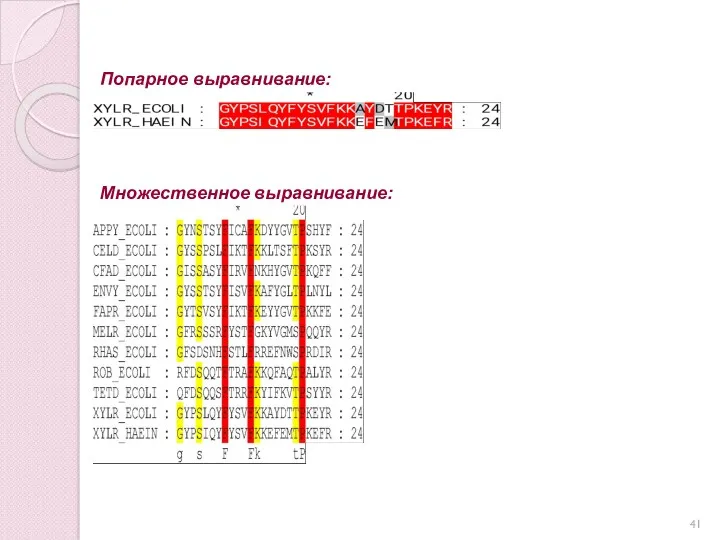
«Идеальное» выравнивание – запись последовательностей одна под другой так, чтобы гомологичные
«Идеальное» выравнивание – запись последовательностей одна под другой так, чтобы гомологичные
![Паттерн – регулярное выражение UNIX’a: [AC]-x-V-x(4)-{ED} Ala или Cys- х-Val-](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/301518/slide-41.jpg)
Паттерн – регулярное выражение UNIX’a:
[AC]-x-V-x(4)-{ED}
Ala или Cys- х-Val- х- х-
Паттерн – регулярное выражение UNIX’a:
[AC]-x-V-x(4)-{ED}
Ala или Cys- х-Val- х- х-
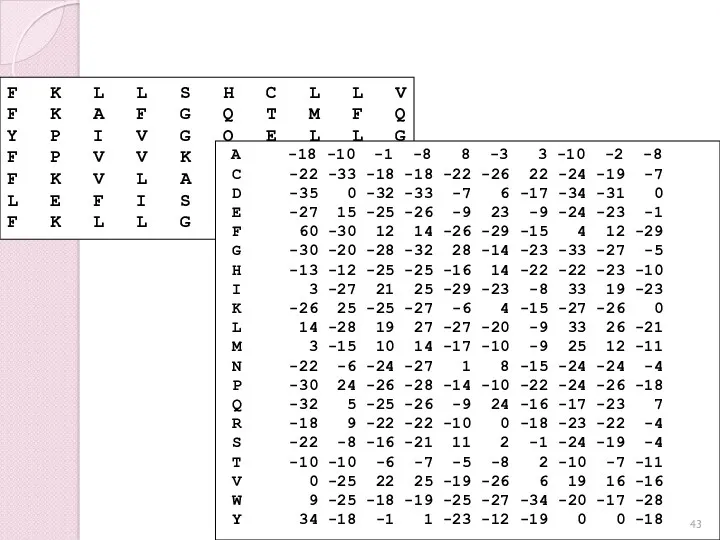
Профиль или весовая матрица (PSSM)
F K L L S H C
Профиль или весовая матрица (PSSM)
F K L L S H C
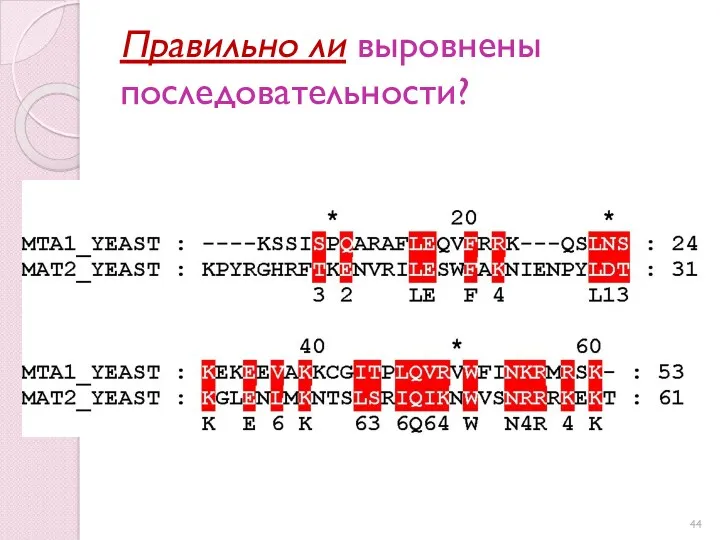
Правильно ли выровнены последовательности?
Правильно ли выровнены последовательности?

В чем биологический смысл выравнивания?
Буквы в одной колонке определяют сопоставление аминокислотных
В чем биологический смысл выравнивания?
Буквы в одной колонке определяют сопоставление аминокислотных
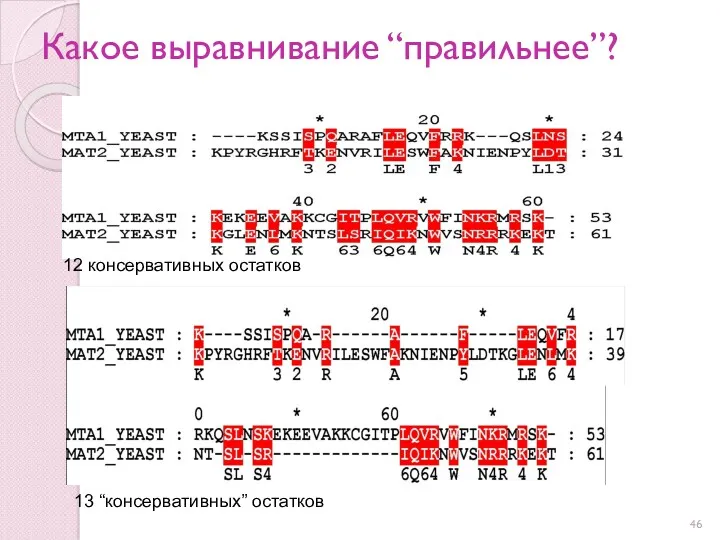
Какое выравнивание “правильнее”?
13 “консервативных” остатков
12 консервативных остатков
Какое выравнивание “правильнее”?
13 “консервативных” остатков
12 консервативных остатков

Чтобы понять смысл выравнивания, вернемся к тому, что такое последовательность аминокислотных
Чтобы понять смысл выравнивания, вернемся к тому, что такое последовательность аминокислотных

(i)Последовательность – удобный способ закодировать структурную (химическую) формулу молекулы белка (до
(i)Последовательность – удобный способ закодировать структурную (химическую) формулу молекулы белка (до
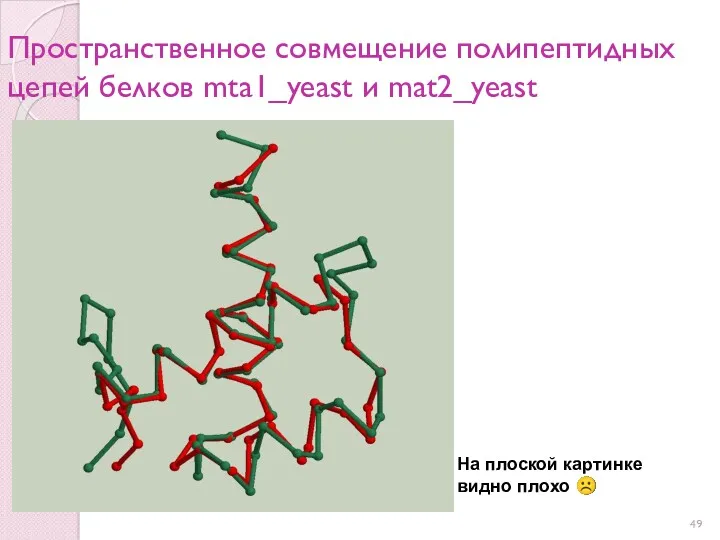
Пространственное совмещение полипептидных цепей белков mta1_yeast и mat2_yeast
На плоской картинке
видно плохо
Пространственное совмещение полипептидных цепей белков mta1_yeast и mat2_yeast
На плоской картинке
видно плохо
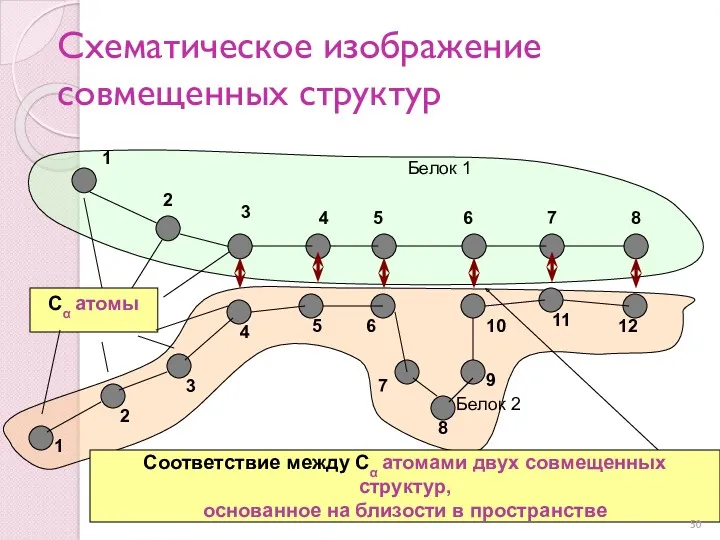
Схематическое изображение совмещенных структур
Схематическое изображение совмещенных структур
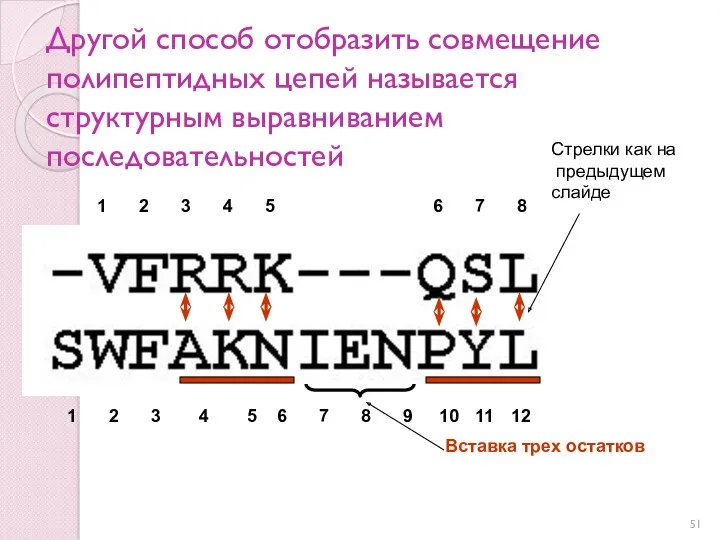
Другой способ отобразить совмещение полипептидных цепей называется структурным выравниванием последовательностей
Стрелки как
Другой способ отобразить совмещение полипептидных цепей называется структурным выравниванием последовательностей
Стрелки как
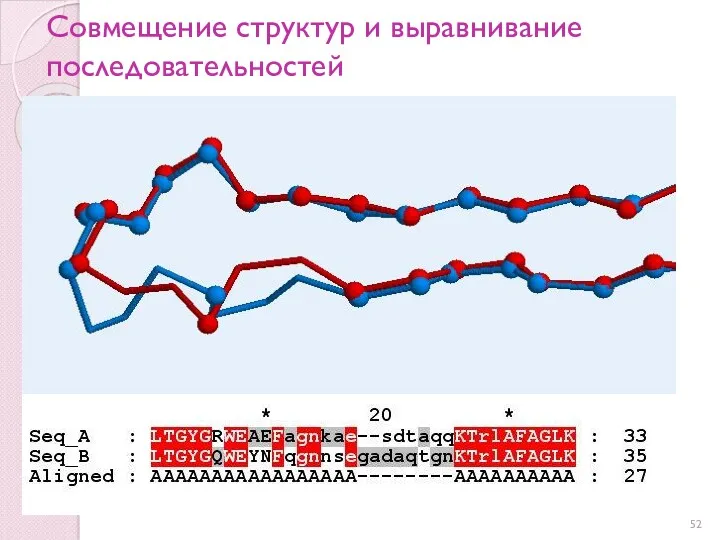
Совмещение структур и выравнивание последовательностей
Совмещение структур и выравнивание последовательностей
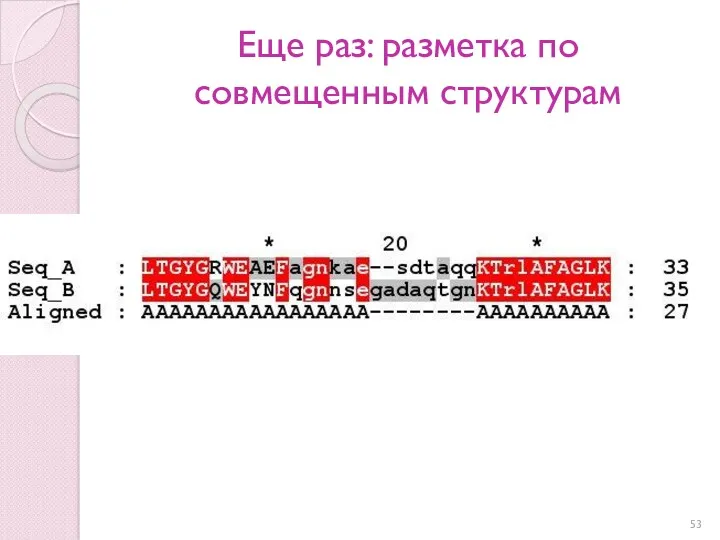
Еще раз: разметка по совмещенным структурам
Еще раз: разметка по совмещенным структурам
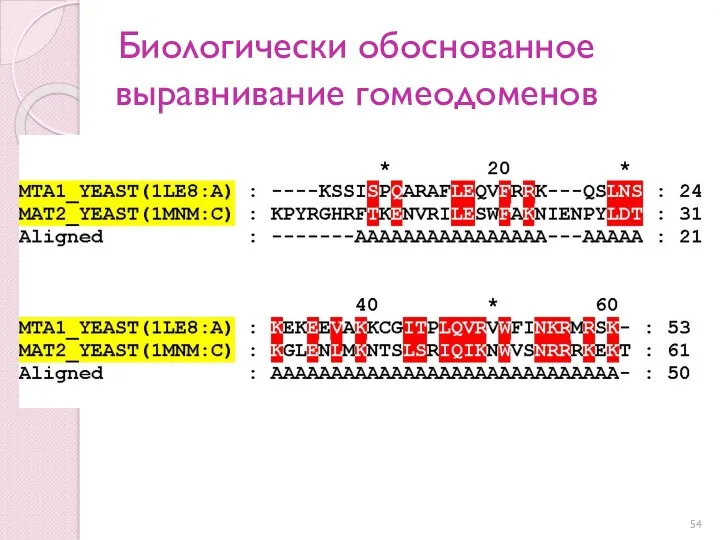
Биологически обоснованное выравнивание гомеодоменов
Биологически обоснованное выравнивание гомеодоменов
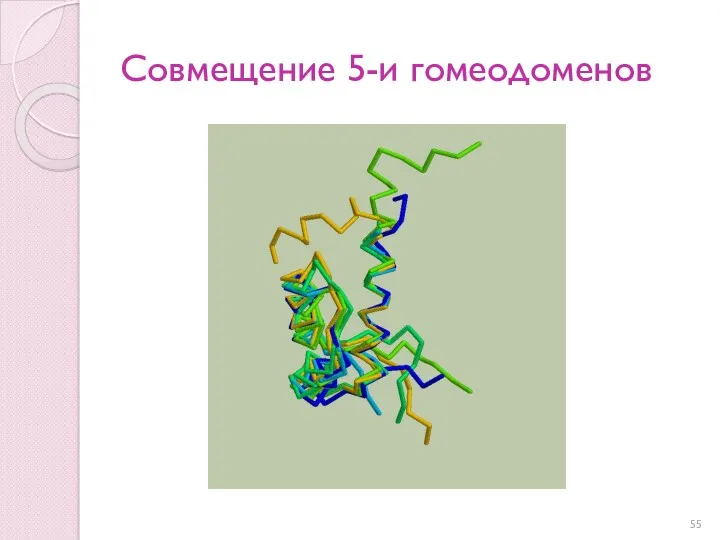
Совмещение 5-и гомеодоменов
Совмещение 5-и гомеодоменов
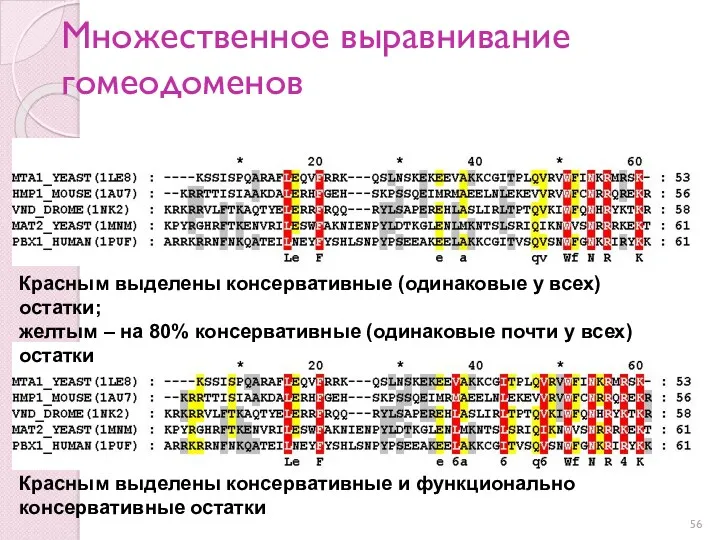
Множественное выравнивание гомеодоменов
Красным выделены консервативные (одинаковые у всех) остатки;
желтым – на
Множественное выравнивание гомеодоменов
Красным выделены консервативные (одинаковые у всех) остатки;
желтым – на

Размеченное множественное выравнивание
Размеченное множественное выравнивание
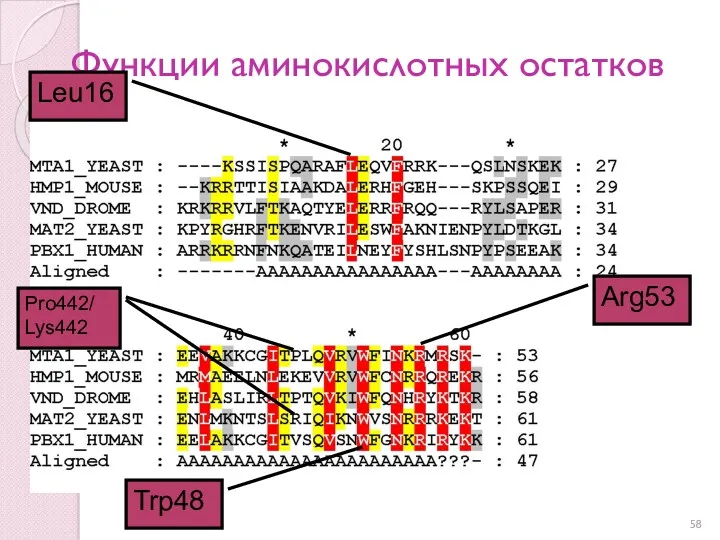
Функции аминокислотных остатков
Trp48
Arg53
Leu16
Pro442/
Lys442
Функции аминокислотных остатков
Trp48
Arg53
Leu16
Pro442/
Lys442

В “правильном” выравнивании много консервативных аминокислотных остатков и функционально консервативных позиций
В “правильном” выравнивании много консервативных аминокислотных остатков и функционально консервативных позиций

Выравнивание и эволюция
Последовательности белка оболочки из двух штаммов
вируса Коксаки
Выравнивание и эволюция
Последовательности белка оболочки из двух штаммов
вируса Коксаки
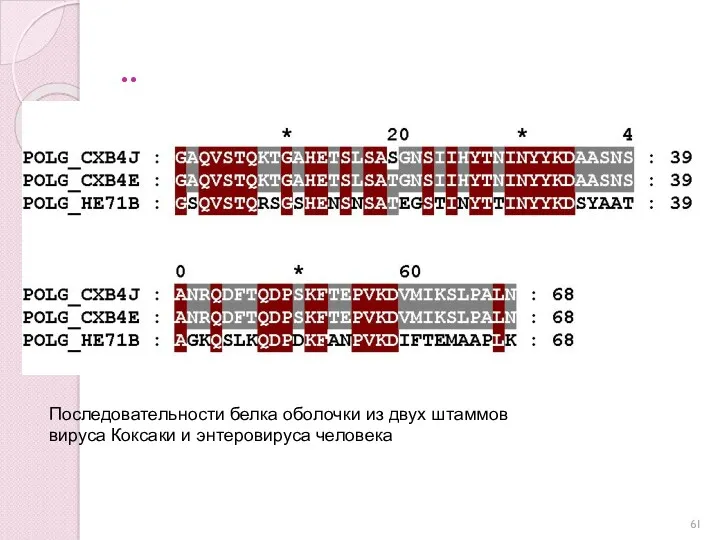
..
Последовательности белка оболочки из двух штаммов
вируса Коксаки и энтеровируса человека
..
Последовательности белка оболочки из двух штаммов
вируса Коксаки и энтеровируса человека

Аминокислотные остатки в одной колонке биологически обоснованного выравнивания, как правило, “произошли”
Аминокислотные остатки в одной колонке биологически обоснованного выравнивания, как правило, “произошли”

ПРОБЛЕМА: как построить “правильное” выравнивание последовательностей белков если структуры белков неизвестны?
ПРОБЛЕМА: как построить “правильное” выравнивание последовательностей белков если структуры белков неизвестны?

На сегодня известны:
более 10 млн(!!!) последовательностей белков (включая фрагменты и
На сегодня известны:
более 10 млн(!!!) последовательностей белков (включая фрагменты и

Алгоритмические решения проблемы воплощены в программах
Программы выравнивания последовательностей тестируются путем
Алгоритмические решения проблемы воплощены в программах
Программы выравнивания последовательностей тестируются путем

Предположим, известны структуры родственных белков и, значит, биологически обоснованное выравнивание последовательностей
При
Предположим, известны структуры родственных белков и, значит, биологически обоснованное выравнивание последовательностей
При

(*) Справедливы ли положения с предыдущего слайда для выравнивания
последовательностей ДНК?
последовательностей РНК?
(*) Справедливы ли положения с предыдущего слайда для выравнивания
последовательностей ДНК?
последовательностей РНК?

2 основних підходи до відтворення просторової структури білка in silico
моделювання за
2 основних підходи до відтворення просторової структури білка in silico
моделювання за

Утворення тривимірної структури білка in vivo відбувається при біосинтезі або
Утворення тривимірної структури білка in vivo відбувається при біосинтезі або
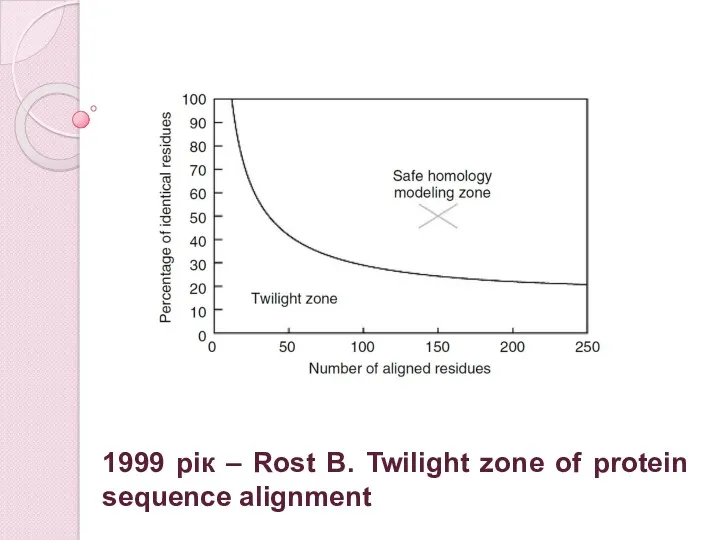
1999 рік – Rost B. Twilight zone of protein sequence alignment
1999 рік – Rost B. Twilight zone of protein sequence alignment
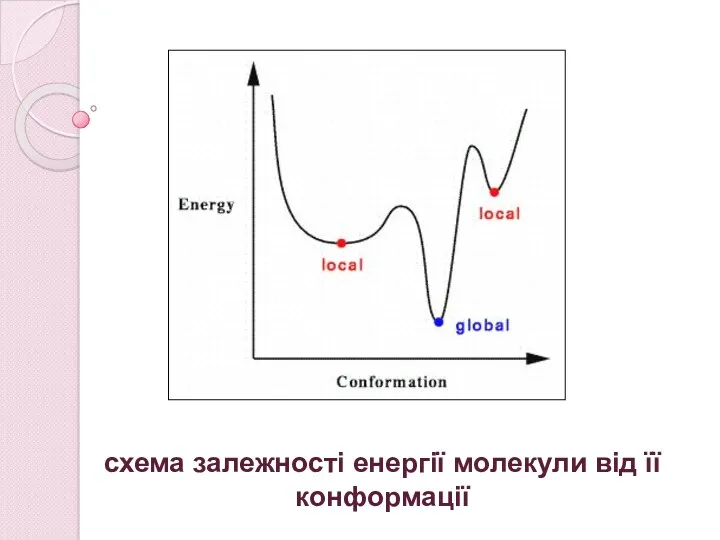
схема залежності енергії молекули від її конформації
схема залежності енергії молекули від її конформації

Для тубулінів будь-якого походження є характерним явище специфічної взаємодії з низькомолекулярними
Для тубулінів будь-якого походження є характерним явище специфічної взаємодії з низькомолекулярними

Незважаючи на високу консервативність структури тубулінів різного походження, рослинні тубуліни характеризуються
Незважаючи на високу консервативність структури тубулінів різного походження, рослинні тубуліни характеризуються
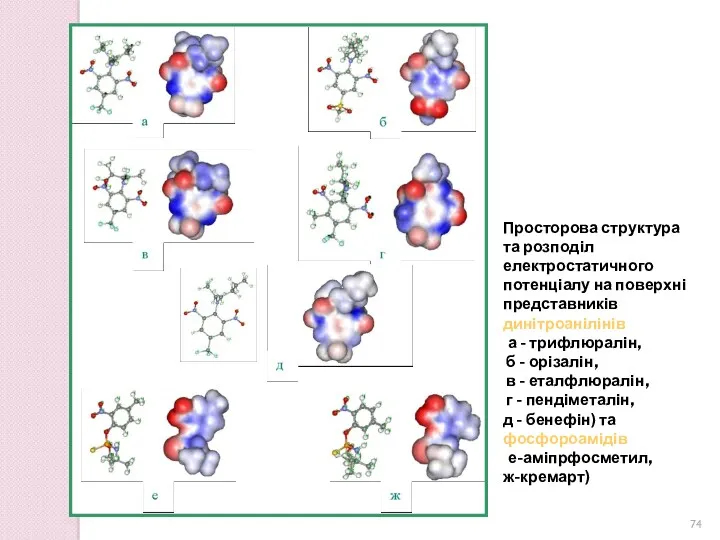
Просторова структура та розподіл електростатичного потенціалу на поверхні представників динітроанілінів
(а
Просторова структура та розподіл електростатичного потенціалу на поверхні представників динітроанілінів
(а
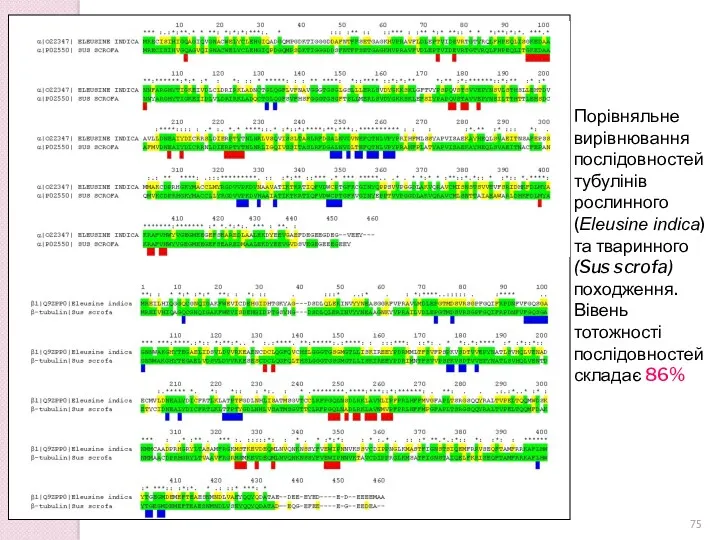
Порівняльне вирівнювання послідовностей тубулінів рослинного (Eleusine indica) та тваринного (Sus scrofa)
Порівняльне вирівнювання послідовностей тубулінів рослинного (Eleusine indica) та тваринного (Sus scrofa)

Відсутність досліджень особливостей просторової структури рослинних тубулінів
труднощі технологічного характеру при
Відсутність досліджень особливостей просторової структури рослинних тубулінів
труднощі технологічного характеру при
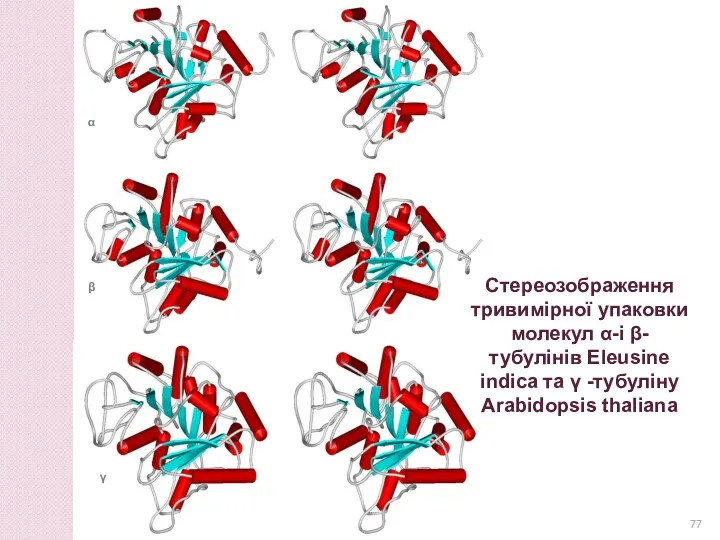
Стереозображення тривимірної упаковки молекул α-і β-тубулінів Eleusine indica та γ -тубуліну
Стереозображення тривимірної упаковки молекул α-і β-тубулінів Eleusine indica та γ -тубуліну

Фундаментальною особливістю тубулінів є явно виражена метастабільність елементів вторинної структури у
Фундаментальною особливістю тубулінів є явно виражена метастабільність елементів вторинної структури у
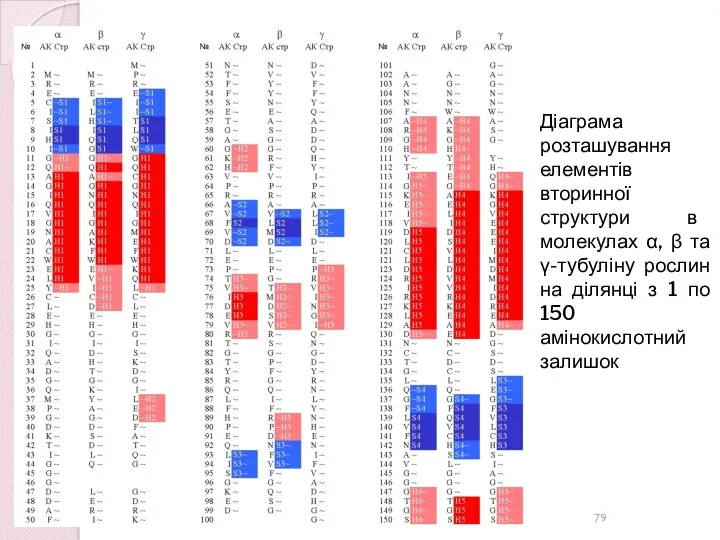
Діаграма розташування елементів вторинної структури в молекулах α, β та γ-тубуліну
Діаграма розташування елементів вторинної структури в молекулах α, β та γ-тубуліну
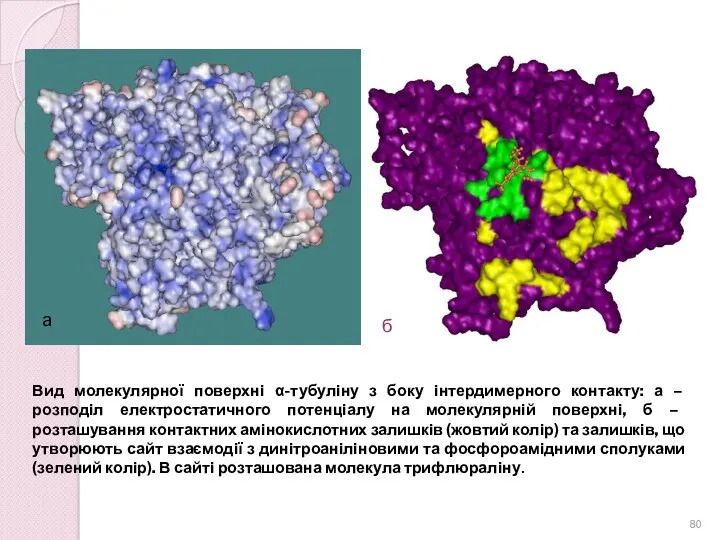
Вид молекулярної поверхні α-тубуліну з боку інтердимерного контакту: а – розподіл
Вид молекулярної поверхні α-тубуліну з боку інтердимерного контакту: а – розподіл

Особливості рельєфу поверхні та розподілу електростатичного потенціалу в області сайту взаємодії
Особливості рельєфу поверхні та розподілу електростатичного потенціалу в області сайту взаємодії

Мутація Met–>Thr в позиції 268 рослинного α-тубуліну, яка викликає виникнення проміжної
Мутація Met–>Thr в позиції 268 рослинного α-тубуліну, яка викликає виникнення проміжної

Порівняльний аналіз послідовностей рослинних α-тубулінів
Представлено ділянки послідовностей, що безпосередньо прилягають до
Порівняльний аналіз послідовностей рослинних α-тубулінів
Представлено ділянки послідовностей, що безпосередньо прилягають до
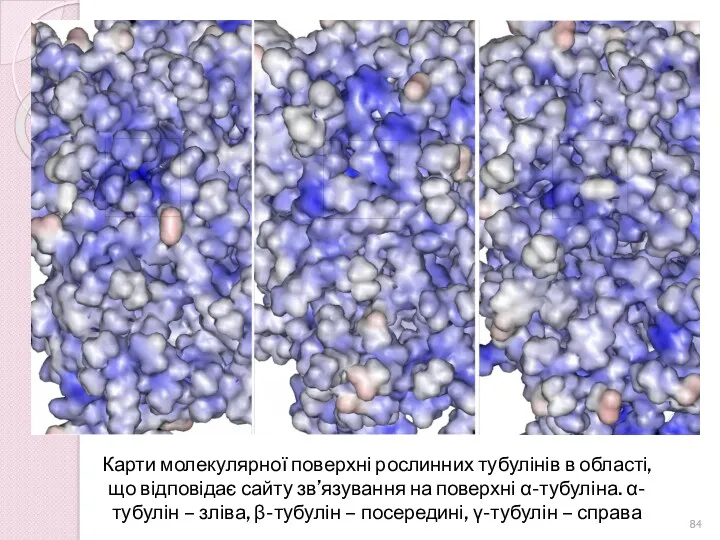
Карти молекулярної поверхні рослинних тубулінів в області, що відповідає сайту зв’язування
Карти молекулярної поверхні рослинних тубулінів в області, що відповідає сайту зв’язування

Распознавание генов
Поиск открытых рамок считывания
Использование статистики (отличия белок-кодирующих и некодирующих областей)
Идентификация
Распознавание генов
Поиск открытых рамок считывания
Использование статистики (отличия белок-кодирующих и некодирующих областей)
Идентификация

Ортологи и паралоги
Ортологи – гени з різних організмів, що розійшлися при
Ортологи и паралоги
Ортологи – гени з різних організмів, що розійшлися при
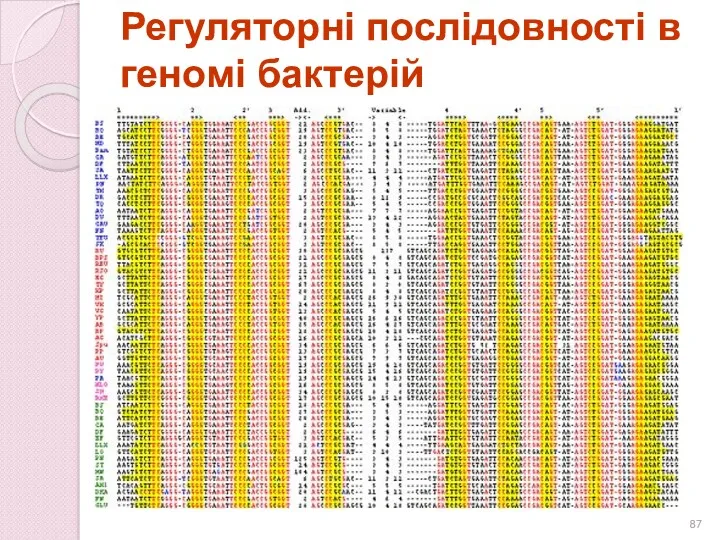
Регуляторні послідовності в геномі бактерій
Регуляторні послідовності в геномі бактерій
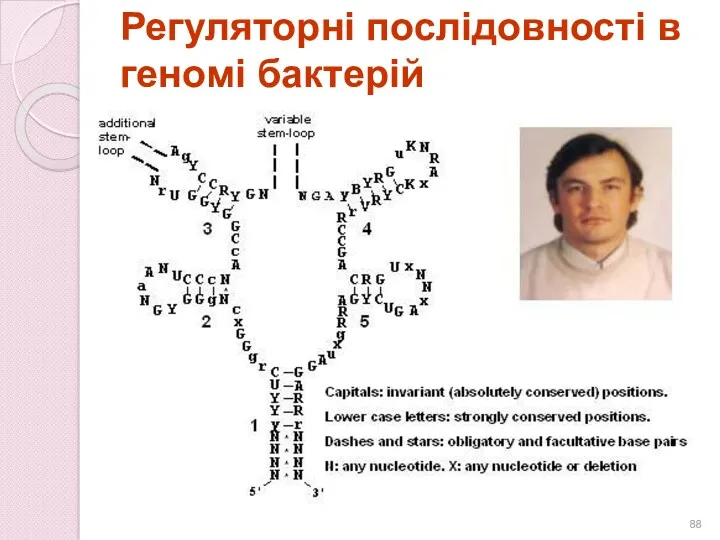
Регуляторні послідовності в геномі бактерій
Регуляторні послідовності в геномі бактерій

Цель (глобальная)
Предсказать свойства организма путем (компьютерного) анализа его генома
(возможно, с использованием
Цель (глобальная)
Предсказать свойства организма путем (компьютерного) анализа его генома
(возможно, с использованием

«Неприкладная» биоинформатика
Молекулярная эволюция
филогения генов
таксономия организмов
горизонтальные переносы и т.п.
положительный и отрицательный отбор
что
«Неприкладная» биоинформатика
Молекулярная эволюция
филогения генов
таксономия организмов
горизонтальные переносы и т.п.
положительный и отрицательный отбор
что

Задачи
С проверяемым ответом
предсказание функции, регуляции, структуры и т.п.:
ставим эксперимент
С непроверяемым ответом
эволюционные
Задачи
С проверяемым ответом
предсказание функции, регуляции, структуры и т.п.:
ставим эксперимент
С непроверяемым ответом
эволюционные

«В принципе не проверяемые ответы» (зависящие от определений)
Так ли они непроверяемы?
Повторы
если
«В принципе не проверяемые ответы» (зависящие от определений)
Так ли они непроверяемы?
Повторы
если

Цель (недостижимая?)
откуда оно все взялось?
первое приближение –
реконструкция генома/свойств
реально ли заглянуть
Цель (недостижимая?)
откуда оно все взялось?
первое приближение –
реконструкция генома/свойств
реально ли заглянуть
 Purinergic signaling
Purinergic signaling Эволюция человека. Предшественники человека
Эволюция человека. Предшественники человека Подцарство Многоклеточные (Metazoa) Тип Круглые, или Первичнополостные черви (Nemathelminthes) класс Нематоды (Nematoda)
Подцарство Многоклеточные (Metazoa) Тип Круглые, или Первичнополостные черви (Nemathelminthes) класс Нематоды (Nematoda) Семейство растений астровые
Семейство растений астровые Презентация к уроку биологии Царство: грибы 6 класс Часть 2 Диск
Презентация к уроку биологии Царство: грибы 6 класс Часть 2 Диск Водная среда обитания
Водная среда обитания Презентация Вид. Критерии вида
Презентация Вид. Критерии вида Разработка урока по биологии Углеводы и липиды 9 класс (проектная деятельность)
Разработка урока по биологии Углеводы и липиды 9 класс (проектная деятельность) Особливості будови клітин
Особливості будови клітин Аптекарский огород
Аптекарский огород Як тварини пристосувалися до життя в різних умовах
Як тварини пристосувалися до життя в різних умовах Опорно-двигательная система человека. Гигиена ОДС
Опорно-двигательная система человека. Гигиена ОДС Заселение Земли человеком. Расы. 7 класс
Заселение Земли человеком. Расы. 7 класс Тварини зоопарку
Тварини зоопарку Выберите признаки классов: 1- паукообразные; 2-ракообразные
Выберите признаки классов: 1- паукообразные; 2-ракообразные Развитие и смена биогеоценозов
Развитие и смена биогеоценозов Витамины. Авитаминоз и гипервитаминоз
Витамины. Авитаминоз и гипервитаминоз Генетика, как наука. Законы Менделя
Генетика, как наука. Законы Менделя Антифризы в живых организмах
Антифризы в живых организмах Артросиндесмология. Виды соединений костей
Артросиндесмология. Виды соединений костей Биотехнология и биоэкономика: состояние и перспективы
Биотехнология и биоэкономика: состояние и перспективы Неделя биологии и химии в школе
Неделя биологии и химии в школе Объект изучения биологии
Объект изучения биологии Современная биология в сюжетах компьютерных игр
Современная биология в сюжетах компьютерных игр Природные сообщества. Биогеоценоз и экосистемы
Природные сообщества. Биогеоценоз и экосистемы Популяції живих організмів та їх основні характеристики
Популяції живих організмів та їх основні характеристики Общие признаки животных
Общие признаки животных Анатомия глазницы
Анатомия глазницы