- Будова клітинної мембрани. Хімічний склад. Транспортні властивості

Содержание
- 2. План Історичний огляд становлення мембранології Будова мембран Функції мембран Хімія мембран Транспорт через мембрану
- 3. 40-і рр. ХІХ ст. – Дюбуа-Реймон повідомив, що між внутрішньою та зовнішньою поверхнями шкіри жаби існує
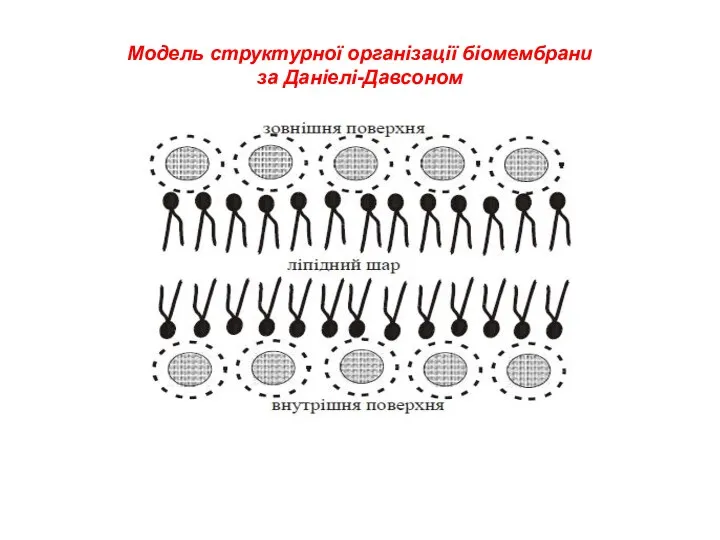
- 4. 1935 р. – Даніелі та Давсон сформулювали гіпотезу про подвійність ліпідного шару, який визначає будову плазматичної
- 5. Модель структурної організації біомембрани за Даніелі-Давсоном
- 6. Модель біомембрани за Сингером –Нікольсоном.
- 7. Загальний план будови біологічної клітинної мембрани
- 8. Функції біомембран 1. Обмеження та відокремлення клітин та органел. Відокремлення клітин від міжклітинного середовища забезпечується плазматичною
- 9. Функції біомембран 2. Контрольований транспорт метаболітів та іонів визначає внутрішнє середовище, що суттєво для гомеостазу, тобто
- 10. Функції біомембран 4. Ферментативний каталіз. У мембранах на межі між ліпідною та водною фазами локалізовані ферменти.
- 11. Функції біомембран 5. Контактна взаємодія з міжклітинним матриксом і взаємодія з іншими клітинами при утворенні тканин.
- 12. Функції біомембран 6. Заякорювання цитоскелету , яке забезпечує підтримку форми клітин та органел, а також клітинну
- 13. Основні властивості мембрани 1) Замкненість. Ліпідні бішари (і мембрани) завжди самостійно замикаються на себе з утворенням
- 14. Основні властивості мембрани 3) Асиметрія. Поверхнева і внутрішня поверхні мембрани переважно різняться за своїм складом: а)
- 15. Ліпіди мембран
- 16. Класи мембранних ліпідів 1. Фосфоліпіди 2. Сфінголіпіди 3. Гліколіпіди 4. Стероїди, а саме холестерин
- 17. Холестерин
- 18. Вплив фосфоліпідів і сфінголіпідів По мірі збільшення вмісту у мембрані ФЛ та СЛ зростають усі показники
- 19. Вплив гліколіпідів і холестерину Ці два види ліпідів здійснюють на лабільність мембрани два протилежні впливи. З
- 20. Білки мембран
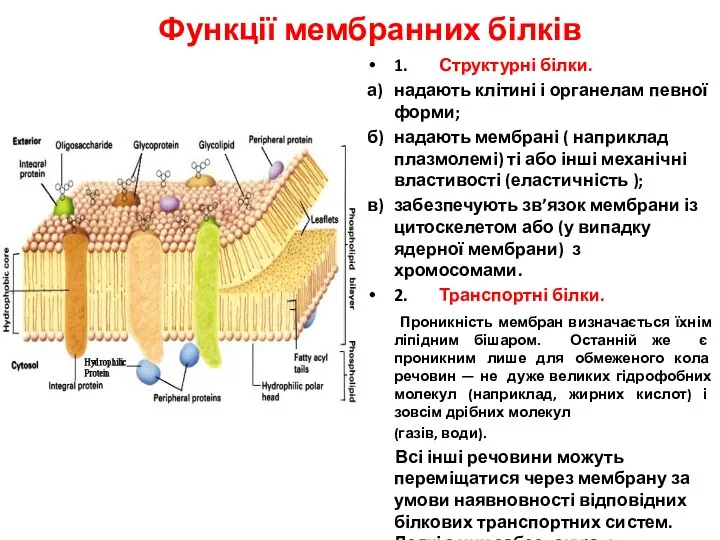
- 21. Функції мембранних білків 1. Структурні білки. а) надають клітині і органелам певної форми; б) надають мембрані
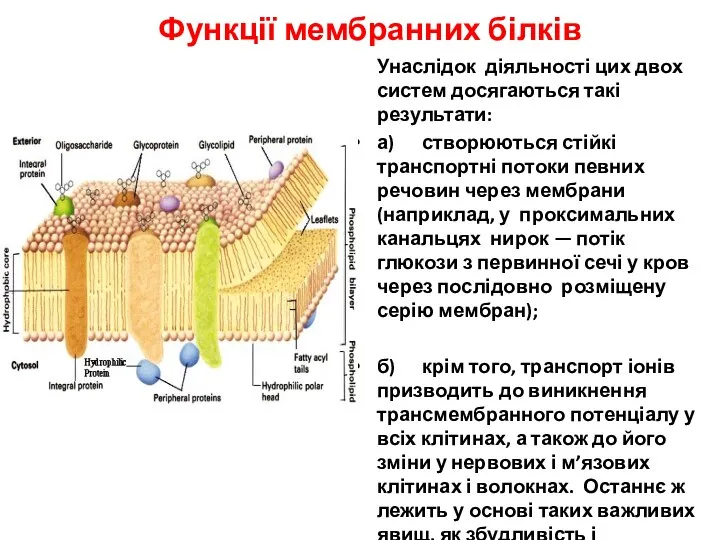
- 22. Функції мембранних білків Унаслідок діяльності цих двох систем досягаються такі результати: а) створюються стійкі транспортні потоки
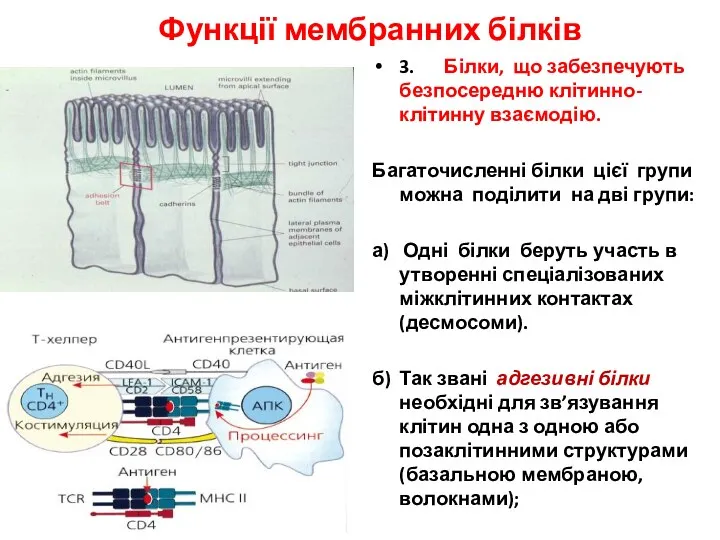
- 23. Функції мембранних білків 3. Білки, що забезпечують безпосередню клітинно-клітинну взаємодію. Багаточисленні білки цієї групи можна поділити
- 24. Triton X-100: (CH3)3C-CH2-C(CH3)2-(C6H6)-(OCH2CH2)10-OH SDS: CH3-(CH2)11-OSO3-Na+ Мембранні білки поділяють на периферичні та інтегральні. Інтегральні зв’язані з мембраною
- 25. Інтегральні білки поділяють на внутрішні трансмембранні і зовнішні, які мають гідрофобний якір. Поліпептидний ланцюг більшості трансмембранних
- 26. У якості якоря може бути фосфатидилінозитольний гліколіпід
- 27. Деякі периферичні білки зв”язуються з зарядженими групами ліпідів безпосередньо за рахунок електростатичних взаємодій (спектрин) Периферичні білки
- 28. Ділянки, що перетинають бішар цілком побудовані з гідрофобних амінокислот і часто мають α-спіральну конфігурацію
- 29. 4. Транспорт речовин через мембрани. Проникність біологічних мембран: пори, канали, помпи.
- 30. 5 різновидів мембранного транспорту: А - Пасивний транспорт Пасивна дифузія Полегшена дифузія В - Активний транспорт
- 31. Транспорт речовин через мембрану а - види переносу; б - пасивний і активний транспорт: 1 -
- 32. Пасивний транспорт
- 33. Полегшена дифузія-один із видів пасивного транспорту Характерний для усіх канальних білків. Відбувається за градієнтом концентрації за
- 34. Полегшена дифузія.Транслокази Транслокази складаються з кількох субодиниць. З урахуванням цього можливі кілька варіантів: Між субодиницями є
- 35. Порівняння пасивного й активного транспорту Пасивний - за електрохімічним градієнтом, відбувається спонтанно або шляхом звичайної дифузії,
- 36. Активний транспорт При активному транспорту речовина проходить через мембрану за допомогою спеціального транспортного білка (транслокази), але
- 37. Симпорт У випадку симпорту обидві речовини переносяться транслоказою в одну сторону. Тобто молекули Y, дифундуючи за
- 38. Антипорт Речовини переносяться транслоказою у взаємно протилежних напрямках. Тобто молекули Y ніби обмінюються на молекули X.
- 39. The activity of membrane transport proteins that regulate the cytosolic pH of mammalian cells changes with
- 40. Nа+-К+-насос Переносник на кожну молекулу АТР, що зазнає гідролізу всередині клітини, активно перекачує 3 іони Nа+
- 41. Nа+,К+-залежна АТФаза Цей білок має дві α- і дві β-субодиниці. Маленький глікозильований β-поліпептид допомагає новосинтезованій α-субодиниці
- 42. EXTRACELLULAR FLUID [Na+] high [K+] low Na+ Na+ Na+ [Na+] low [K+] high CYTOPLASM 1 Цитоплазматичний
- 43. Na+ Na+ Na+ ATP P ADP 2 Зв”язування Na+ стимулює фосфорилювання за допомогою АТР з цитоплазматичного
- 44. Na+ P Na+ Na+ 3 Конформаційні зміни білка, що забезпечує переміщення Nа+ крізь мембрану і вивільнення
- 45. P P K+ K+ 4 Зв”язування К+ на зовнішній поверхні
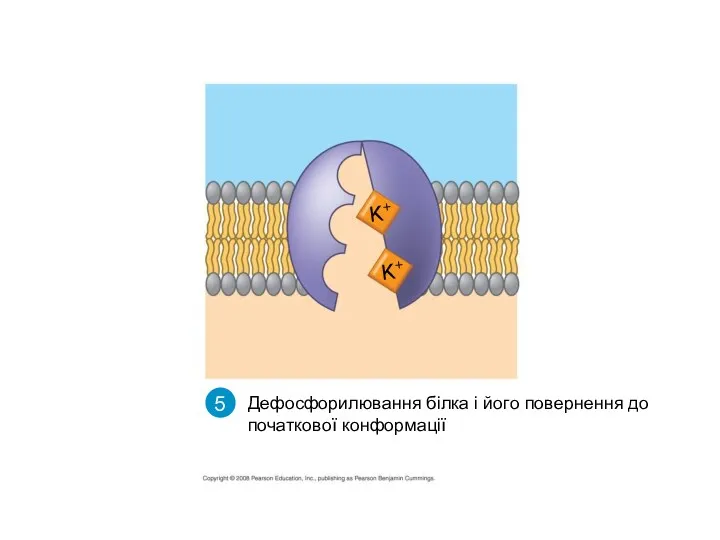
- 46. K+ K+ 5 Дефосфорилювання білка і його повернення до початкової конформації
- 47. Екзоцитоз у еукаріот клітинний процес, при якому внутрішньоклітинні везикули (мембранні пухирці) зливаються з зовнішньої клітинної мембраною.
- 48. Екзоцитоз в синапсі: передача сигналу від нейрона А до нейрона B. 1. Мітохондрія 2. Синаптична везикула
- 49. Екзоцитоз може виконувати три основні завдання: доставка на клітинну мембрану ліпідів, необхідного для росту клітини; вивільнення
- 50. Розрізняють наступні етапи екзоцитозу: Транспортування везикули від місця синтезу і формування ( апарат Гольджі ) до
- 51. Ендоцитоз Ендоцитоз ( англ. endocytosis) - процес захоплення (інтерналізації) зовнішнього матеріалу клітиною, здійснюваний шляхом утворення мембранних
- 52. Типи
- 53. Різні типи ендоцитоза: фагоцитоз , піноцитоз і рецептор-опосередкований ендоцитоз . Фагоцитоз (поїдання клітиною - процес поглинання
- 55. Скачать презентацию
План
Історичний огляд становлення мембранології
Будова мембран
Функції мембран
Хімія мембран
Транспорт через мембрану
План
Історичний огляд становлення мембранології
Будова мембран
Функції мембран
Хімія мембран
Транспорт через мембрану

40-і рр. ХІХ ст. – Дюбуа-Реймон повідомив, що між внутрішньою та зовнішньою поверхнями
40-і рр. ХІХ ст. – Дюбуа-Реймон повідомив, що між внутрішньою та зовнішньою поверхнями

1935 р. – Даніелі та Давсон сформулювали гіпотезу про подвійність ліпідного шару, який
1935 р. – Даніелі та Давсон сформулювали гіпотезу про подвійність ліпідного шару, який

Модель структурної організації біомембрани
за Даніелі-Давсоном
Модель структурної організації біомембрани
за Даніелі-Давсоном
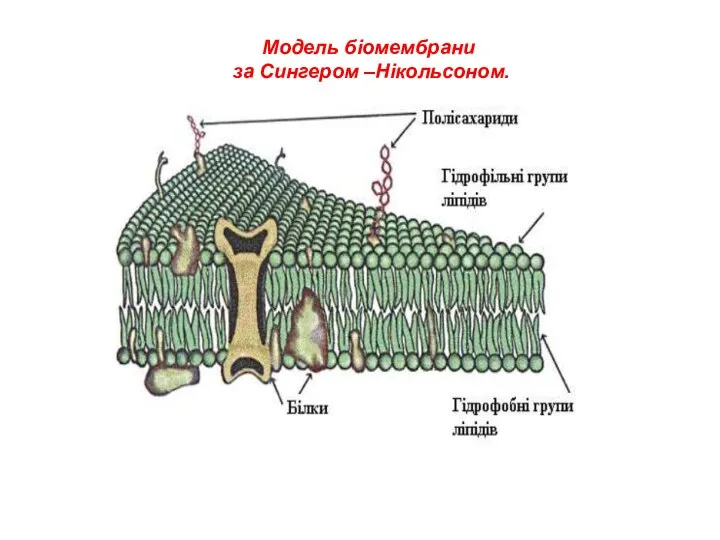
Модель біомембрани
за Сингером –Нікольсоном.
Модель біомембрани
за Сингером –Нікольсоном.
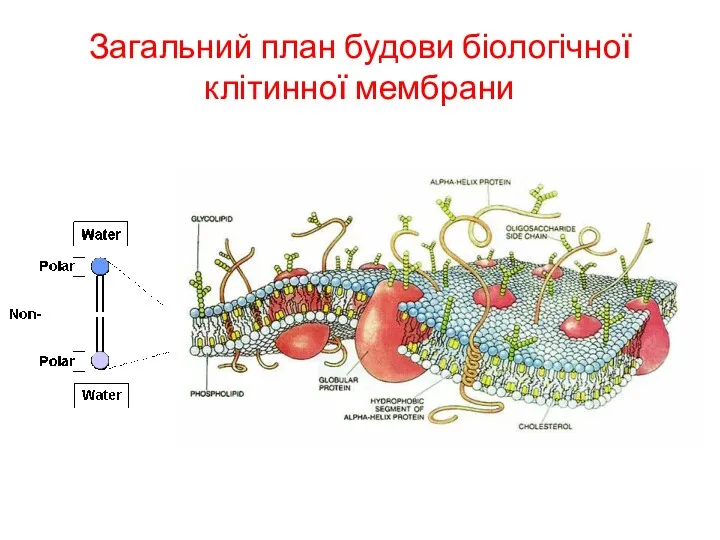
Загальний план будови біологічної клітинної мембрани
Загальний план будови біологічної клітинної мембрани
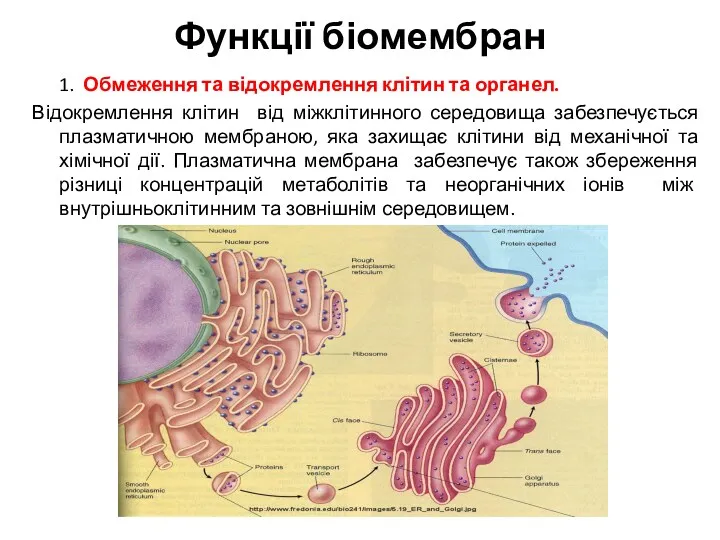
Функції біомембран
1. Обмеження та відокремлення клітин та органел.
Відокремлення клітин від міжклітинного середовища забезпечується
Функції біомембран
1. Обмеження та відокремлення клітин та органел.
Відокремлення клітин від міжклітинного середовища забезпечується
Функції біомембран
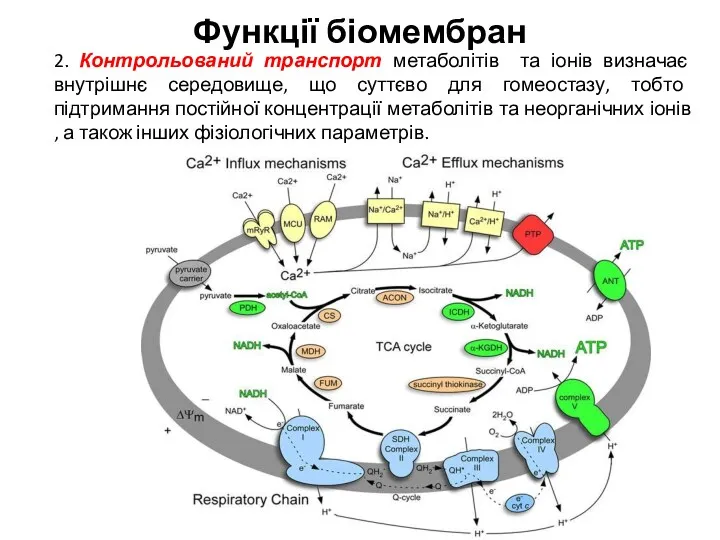
2. Контрольований транспорт метаболітів та іонів визначає внутрішнє середовище, що суттєво для
Функції біомембран
2. Контрольований транспорт метаболітів та іонів визначає внутрішнє середовище, що суттєво для
Функції біомембран
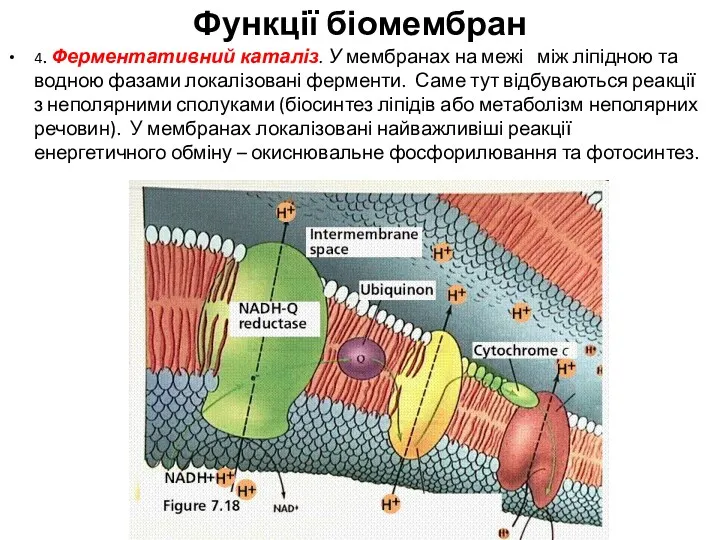
4. Ферментативний каталіз. У мембранах на межі між ліпідною та водною фазами
Функції біомембран
4. Ферментативний каталіз. У мембранах на межі між ліпідною та водною фазами
Функції біомембран
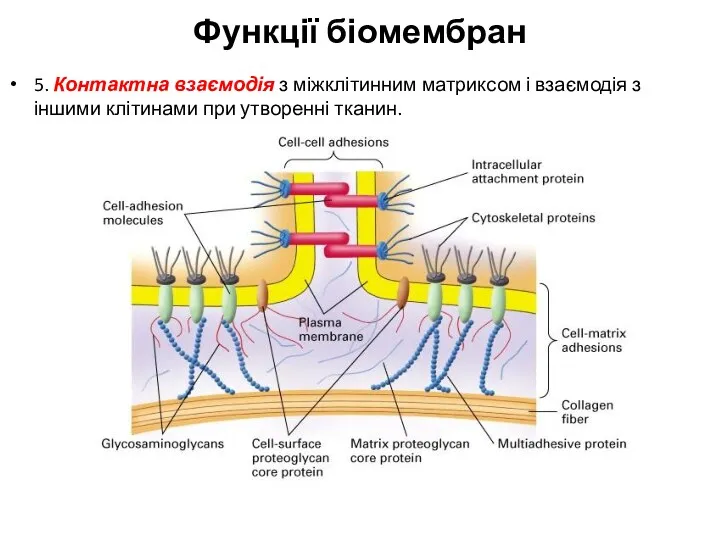
5. Контактна взаємодія з міжклітинним матриксом і взаємодія з іншими клітинами при
Функції біомембран
5. Контактна взаємодія з міжклітинним матриксом і взаємодія з іншими клітинами при
Функції біомембран
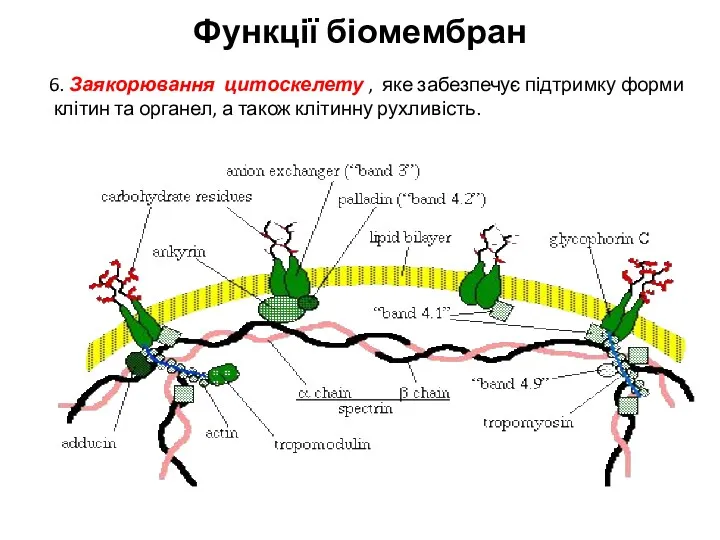
6. Заякорювання цитоскелету , яке забезпечує підтримку форми клітин та органел,
Функції біомембран
6. Заякорювання цитоскелету , яке забезпечує підтримку форми клітин та органел,
Основні властивості мембрани
1) Замкненість. Ліпідні бішари (і мембрани) завжди самостійно замикаються на себе з
Основні властивості мембрани
1) Замкненість. Ліпідні бішари (і мембрани) завжди самостійно замикаються на себе з

Основні властивості мембрани
3) Асиметрія. Поверхнева і внутрішня поверхні мембрани переважно різняться за своїм складом:
а) вуглеводневі
Основні властивості мембрани
3) Асиметрія. Поверхнева і внутрішня поверхні мембрани переважно різняться за своїм складом:
а) вуглеводневі

Ліпіди мембран
Ліпіди мембран
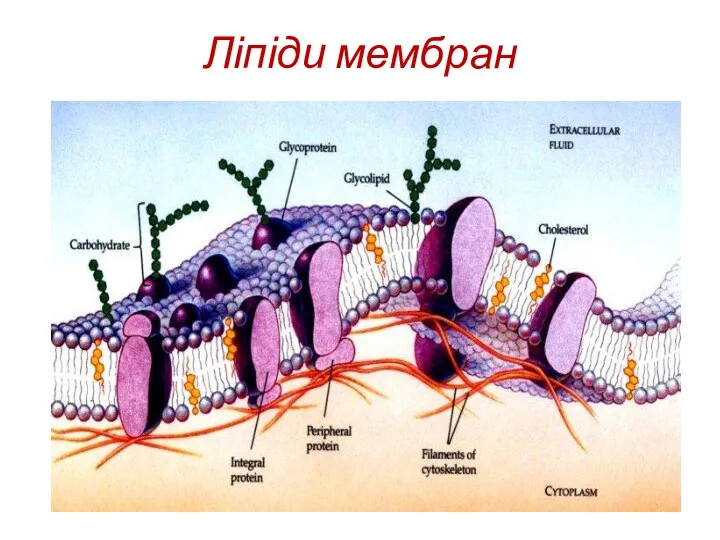
Класи мембранних ліпідів
1. Фосфоліпіди
2. Сфінголіпіди
3. Гліколіпіди
4. Стероїди, а саме холестерин
Класи мембранних ліпідів
1. Фосфоліпіди
2. Сфінголіпіди
3. Гліколіпіди
4. Стероїди, а саме холестерин
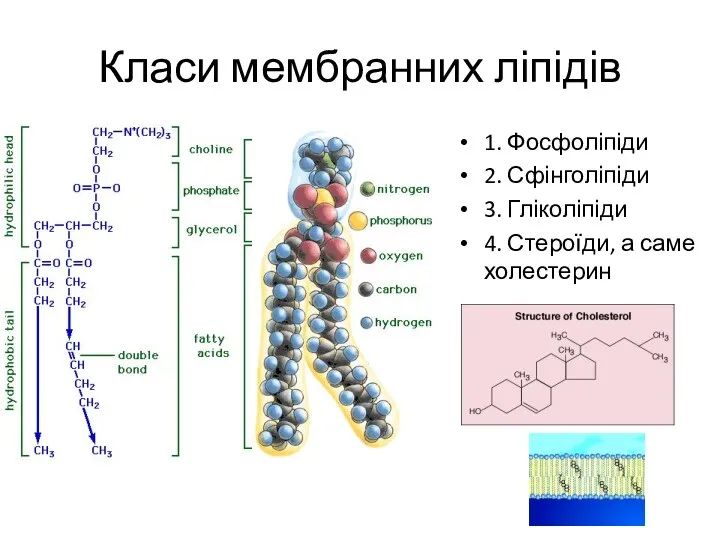
Холестерин
Холестерин
Вплив фосфоліпідів і сфінголіпідів
По мірі збільшення вмісту у мембрані ФЛ та СЛ зростають
Вплив фосфоліпідів і сфінголіпідів
По мірі збільшення вмісту у мембрані ФЛ та СЛ зростають
Вплив гліколіпідів і холестерину
Ці два види ліпідів здійснюють на лабільність мембрани два протилежні
Вплив гліколіпідів і холестерину
Ці два види ліпідів здійснюють на лабільність мембрани два протилежні

Білки мембран
Білки мембран
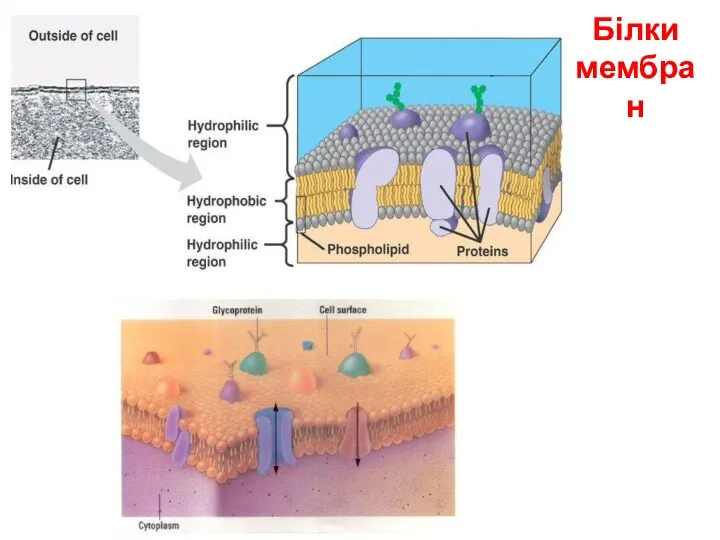
Функції мембранних білків
1. Структурні білки.
а) надають клітині і органелам певної форми;
б) надають мембрані ( наприклад
Функції мембранних білків
1. Структурні білки.
а) надають клітині і органелам певної форми;
б) надають мембрані ( наприклад
Функції мембранних білків
Унаслідок діяльності цих двох систем досягаються такі результати:
а) створюються стійкі транспортні
Функції мембранних білків
Унаслідок діяльності цих двох систем досягаються такі результати:
а) створюються стійкі транспортні
Функції мембранних білків
3. Білки, що забезпечують безпосередню клітинно-клітинну взаємодію.
Багаточисленні білки цієї групи можна поділити
Функції мембранних білків
3. Білки, що забезпечують безпосередню клітинно-клітинну взаємодію.
Багаточисленні білки цієї групи можна поділити
Triton X-100: (CH3)3C-CH2-C(CH3)2-(C6H6)-(OCH2CH2)10-OH
SDS: CH3-(CH2)11-OSO3-Na+
Мембранні білки поділяють на периферичні та інтегральні. Інтегральні зв’язані з
Triton X-100: (CH3)3C-CH2-C(CH3)2-(C6H6)-(OCH2CH2)10-OH
SDS: CH3-(CH2)11-OSO3-Na+
Мембранні білки поділяють на периферичні та інтегральні. Інтегральні зв’язані з
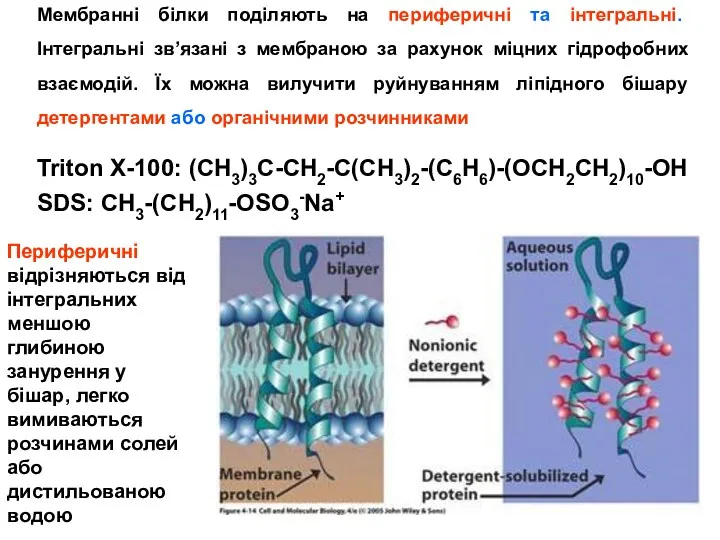
Інтегральні білки поділяють на внутрішні трансмембранні і зовнішні, які мають гідрофобний якір. Поліпептидний
Інтегральні білки поділяють на внутрішні трансмембранні і зовнішні, які мають гідрофобний якір. Поліпептидний
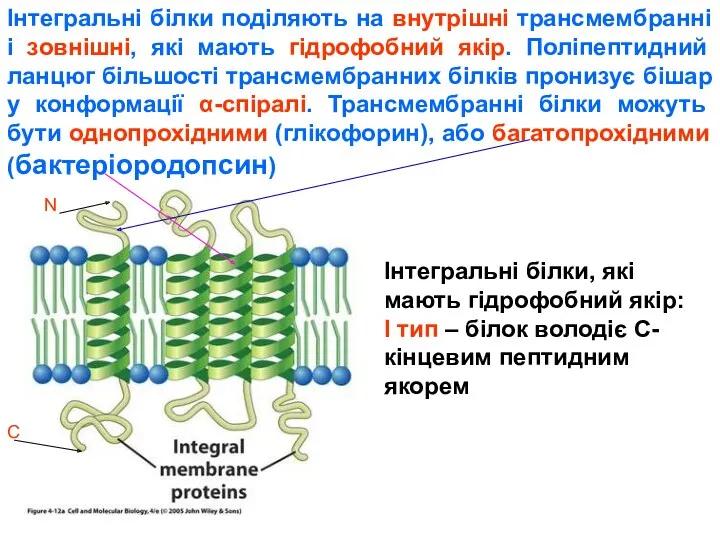
У якості якоря може бути фосфатидилінозитольний гліколіпід
У якості якоря може бути фосфатидилінозитольний гліколіпід
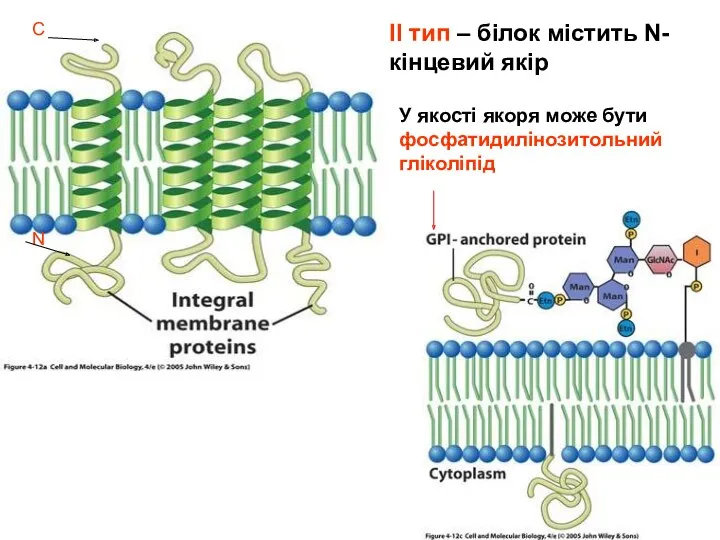
Деякі периферичні білки зв”язуються з зарядженими групами ліпідів безпосередньо за рахунок електростатичних взаємодій
Деякі периферичні білки зв”язуються з зарядженими групами ліпідів безпосередньо за рахунок електростатичних взаємодій
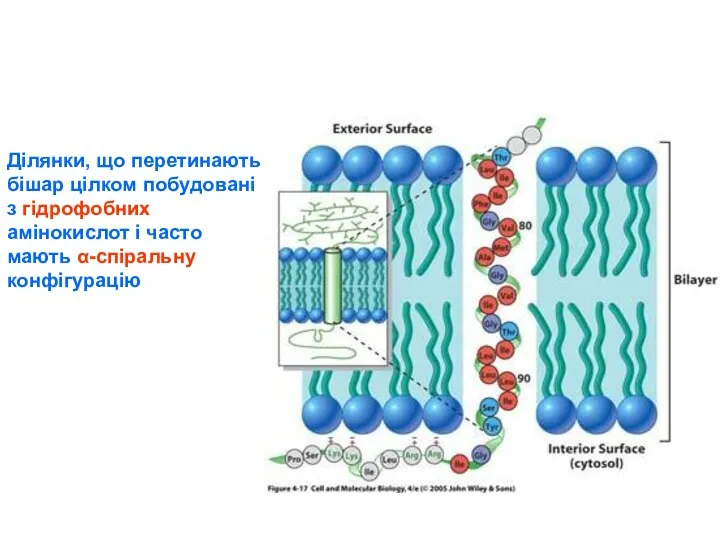
Ділянки, що перетинають бішар цілком побудовані з гідрофобних амінокислот і часто мають α-спіральну
Ділянки, що перетинають бішар цілком побудовані з гідрофобних амінокислот і часто мають α-спіральну
4. Транспорт речовин через мембрани. Проникність біологічних мембран: пори, канали, помпи.
4. Транспорт речовин через мембрани. Проникність біологічних мембран: пори, канали, помпи.

5 різновидів мембранного транспорту:
А - Пасивний транспорт
Пасивна дифузія
Полегшена дифузія
В -
5 різновидів мембранного транспорту:
А - Пасивний транспорт
Пасивна дифузія
Полегшена дифузія
В -

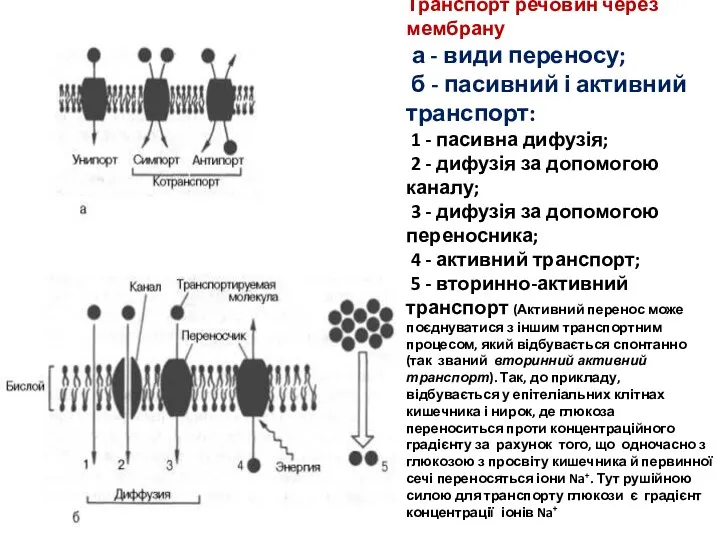
Транспорт речовин через мембрану
а - види переносу;
б - пасивний і активний
Транспорт речовин через мембрану а - види переносу; б - пасивний і активний
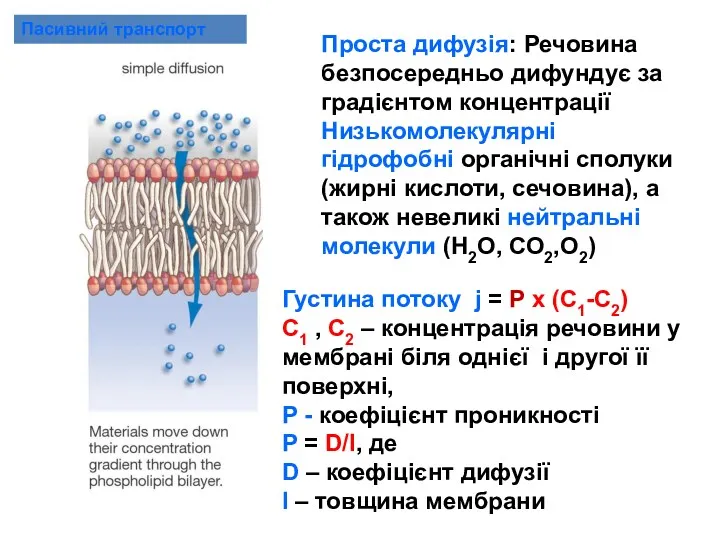
Пасивний транспорт
Пасивний транспорт
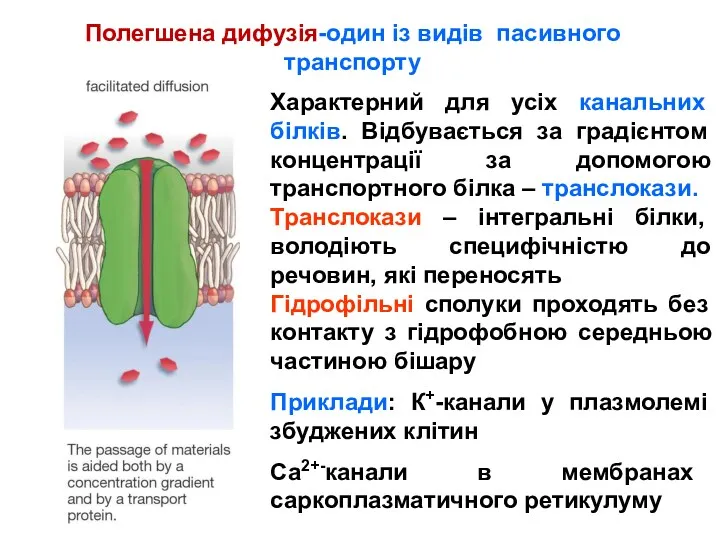
Полегшена дифузія-один із видів пасивного транспорту
Характерний для усіх канальних білків. Відбувається за градієнтом
Полегшена дифузія-один із видів пасивного транспорту
Характерний для усіх канальних білків. Відбувається за градієнтом
Полегшена дифузія.Транслокази
Транслокази складаються з кількох субодиниць. З урахуванням цього можливі кілька варіантів:
Між субодиницями
Полегшена дифузія.Транслокази
Транслокази складаються з кількох субодиниць. З урахуванням цього можливі кілька варіантів:
Між субодиницями

Порівняння пасивного й активного транспорту
Пасивний - за електрохімічним градієнтом, відбувається спонтанно або
Порівняння пасивного й активного транспорту
Пасивний - за електрохімічним градієнтом, відбувається спонтанно або
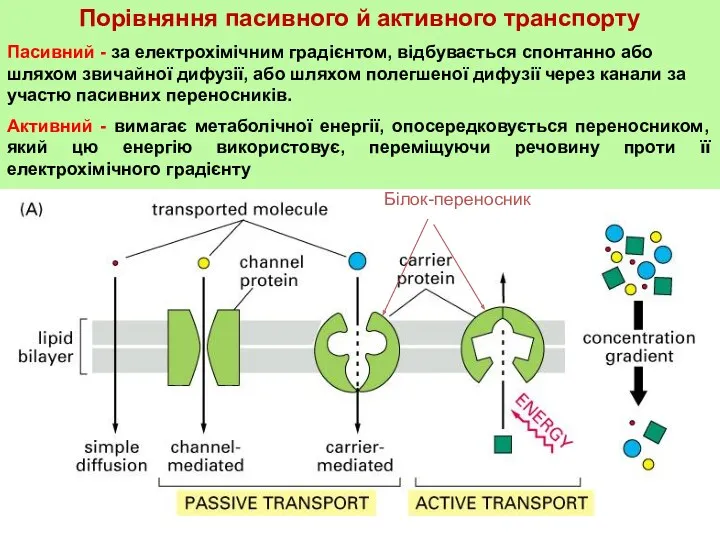
Активний транспорт
При активному транспорту речовина проходить через мембрану за допомогою спеціального транспортного білка
Активний транспорт
При активному транспорту речовина проходить через мембрану за допомогою спеціального транспортного білка

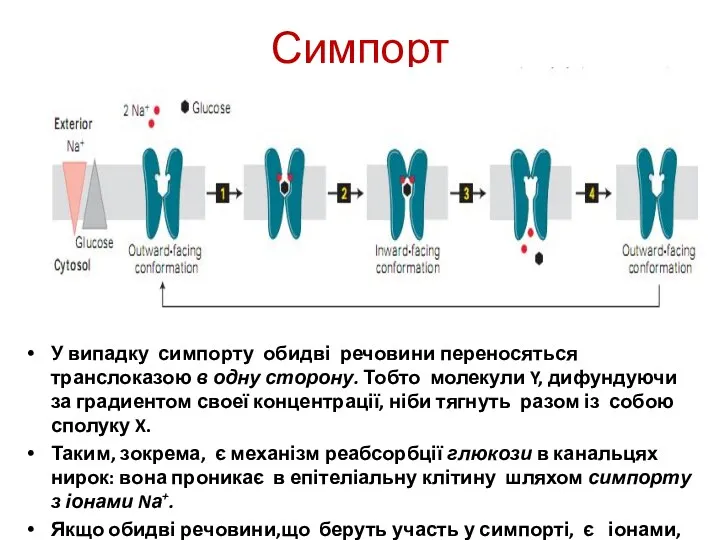
Симпорт
У випадку симпорту обидві речовини переносяться транслоказою в одну сторону. Тобто молекули Y,
Симпорт
У випадку симпорту обидві речовини переносяться транслоказою в одну сторону. Тобто молекули Y,
Антипорт
Речовини переносяться транслоказою у взаємно протилежних напрямках. Тобто молекули Y ніби обмінюються на
Антипорт
Речовини переносяться транслоказою у взаємно протилежних напрямках. Тобто молекули Y ніби обмінюються на

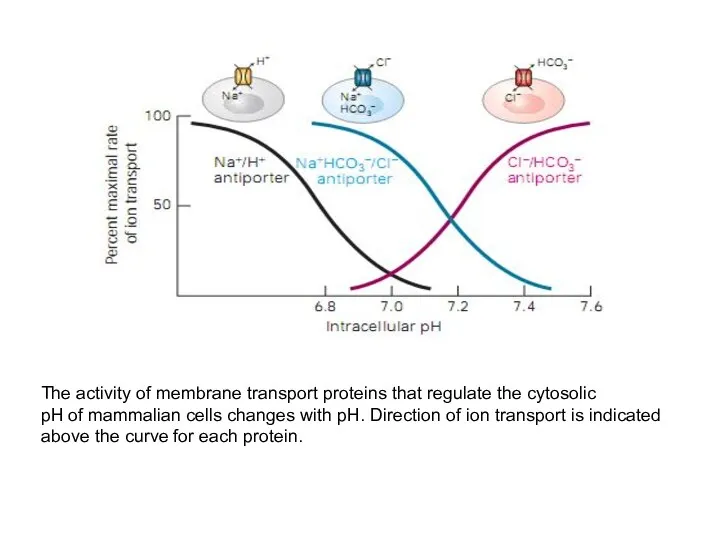
The activity of membrane transport proteins that regulate the cytosolic
pH of mammalian
The activity of membrane transport proteins that regulate the cytosolic
pH of mammalian
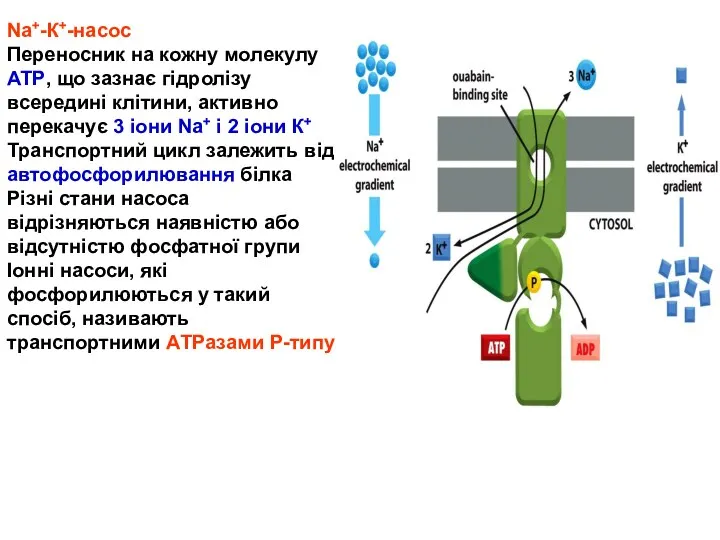
Nа+-К+-насос
Переносник на кожну молекулу АТР, що зазнає гідролізу всередині клітини, активно перекачує 3
Nа+-К+-насос
Переносник на кожну молекулу АТР, що зазнає гідролізу всередині клітини, активно перекачує 3
Nа+,К+-залежна АТФаза
Цей білок має дві α- і дві β-субодиниці. Маленький глікозильований β-поліпептид допомагає
Nа+,К+-залежна АТФаза
Цей білок має дві α- і дві β-субодиниці. Маленький глікозильований β-поліпептид допомагає
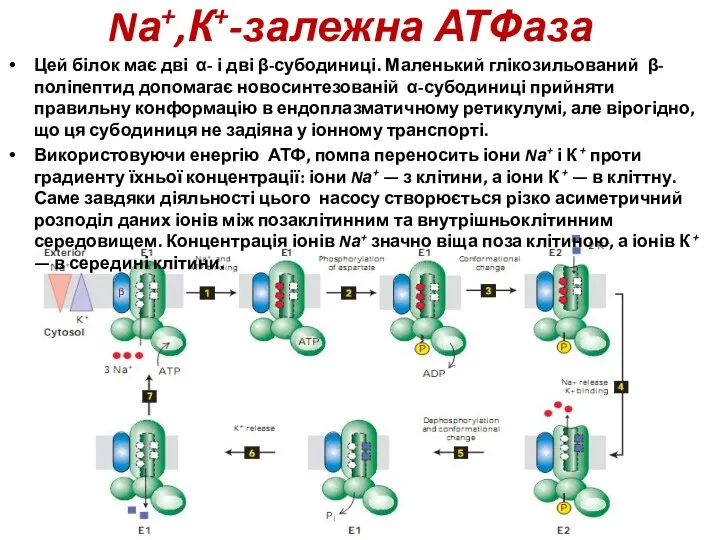
EXTRACELLULAR
FLUID
[Na+] high
[K+] low
Na+
Na+
Na+
[Na+] low
[K+] high
CYTOPLASM
1
EXTRACELLULAR
FLUID
[Na+] high
[K+] low
Na+
Na+
Na+
[Na+] low
[K+] high
CYTOPLASM
1
![EXTRACELLULAR FLUID [Na+] high [K+] low Na+ Na+ Na+ [Na+] low [K+] high](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/314090/slide-41.jpg)
Na+
Na+
Na+
ATP
P
ADP
2
Зв”язування Na+ стимулює фосфорилювання за допомогою АТР
Na+
Na+
Na+
ATP
P
ADP
2
Зв”язування Na+ стимулює фосфорилювання за допомогою АТР
Na+
P
Na+
Na+
3
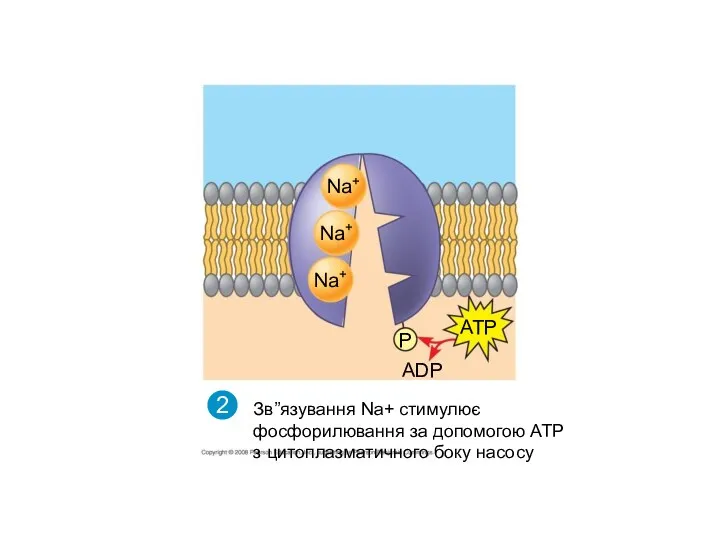
Конформаційні зміни білка, що забезпечує переміщення Nа+ крізь
Na+
P
Na+
Na+
3
Конформаційні зміни білка, що забезпечує переміщення Nа+ крізь
P
P
K+
K+
4
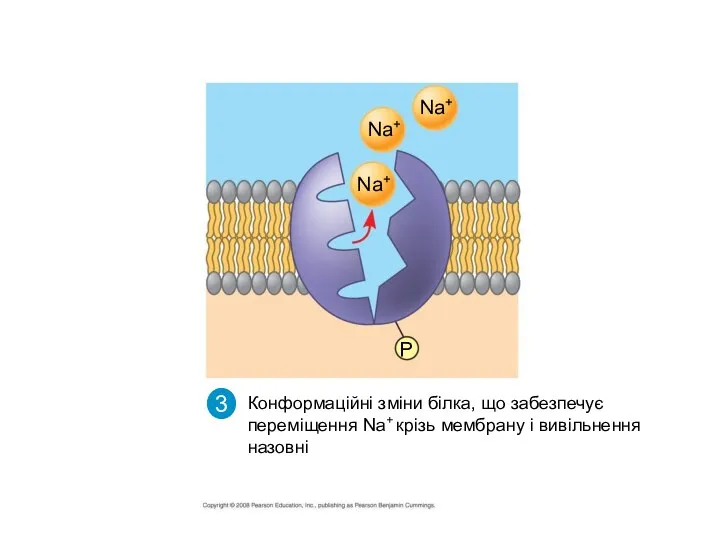
Зв”язування К+ на зовнішній поверхні
P
P
K+
K+
4
Зв”язування К+ на зовнішній поверхні
K+
K+
5
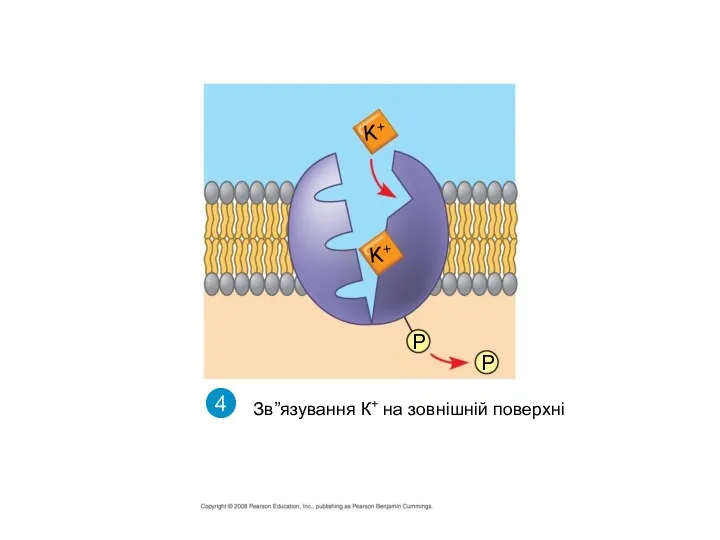
Дефосфорилювання білка і його повернення до початкової конформації
K+
K+
5
Дефосфорилювання білка і його повернення до початкової конформації
Екзоцитоз
у еукаріот клітинний процес, при якому внутрішньоклітинні везикули (мембранні пухирці) зливаються з зовнішньої клітинної мембраною. При екзоцитозі
Екзоцитоз
у еукаріот клітинний процес, при якому внутрішньоклітинні везикули (мембранні пухирці) зливаються з зовнішньої клітинної мембраною. При екзоцитозі

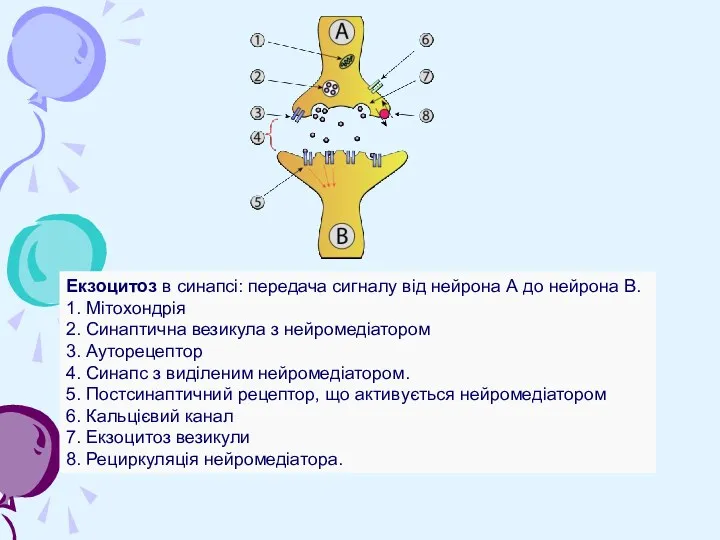
Екзоцитоз в синапсі: передача сигналу від нейрона А до нейрона B.
1. Мітохондрія
2. Синаптична везикула з нейромедіатором
3. Ауторецептор
4. Синапс з виділеним нейромедіатором.
5. Постсинаптичний рецептор, що
Екзоцитоз в синапсі: передача сигналу від нейрона А до нейрона B. 1. Мітохондрія 2. Синаптична везикула з нейромедіатором 3. Ауторецептор 4. Синапс з виділеним нейромедіатором. 5. Постсинаптичний рецептор, що
Екзоцитоз може виконувати три основні завдання:
доставка на клітинну мембрану ліпідів, необхідного для росту
Екзоцитоз може виконувати три основні завдання:
доставка на клітинну мембрану ліпідів, необхідного для росту

Розрізняють наступні етапи екзоцитозу:
Транспортування везикули від місця синтезу і формування ( апарат Гольджі ) до місця
Розрізняють наступні етапи екзоцитозу:
Транспортування везикули від місця синтезу і формування ( апарат Гольджі ) до місця

Ендоцитоз
Ендоцитоз ( англ. endocytosis) - процес захоплення (інтерналізації) зовнішнього матеріалу клітиною, здійснюваний шляхом утворення мембранних везикул. В результаті ендоцитозу
Ендоцитоз
Ендоцитоз ( англ. endocytosis) - процес захоплення (інтерналізації) зовнішнього матеріалу клітиною, здійснюваний шляхом утворення мембранних везикул. В результаті ендоцитозу

Типи
Типи
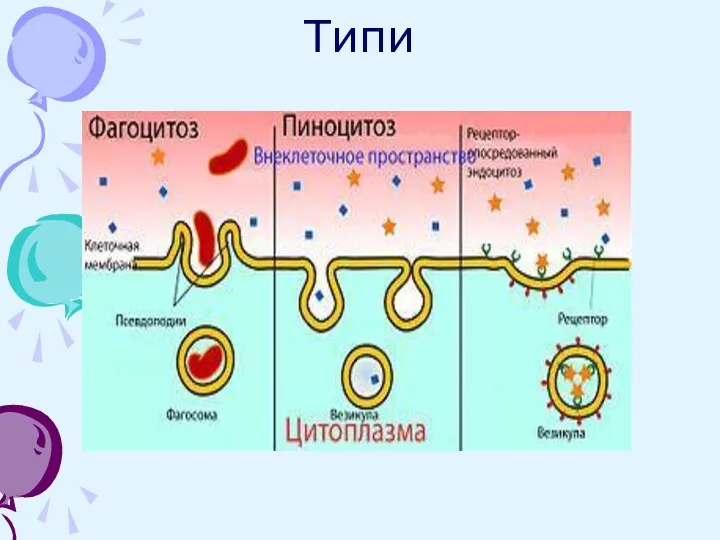
Різні типи ендоцитоза: фагоцитоз , піноцитоз і рецептор-опосередкований ендоцитоз
.
Фагоцитоз (поїдання клітиною - процес поглинання клітиною твердих об'єктів, таких як клітини еукаріот, бактерії , віруси,
Різні типи ендоцитоза: фагоцитоз , піноцитоз і рецептор-опосередкований ендоцитоз
.
Фагоцитоз (поїдання клітиною - процес поглинання клітиною твердих об'єктів, таких як клітини еукаріот, бактерії , віруси,

 Успехи современной биотехнологии
Успехи современной биотехнологии Презентации к теме Химический состав клетки (5 класс)
Презентации к теме Химический состав клетки (5 класс) Возрастные особенности органов пищеварения
Возрастные особенности органов пищеварения Компетентностно-ориентированные задания по биологии 9 класс
Компетентностно-ориентированные задания по биологии 9 класс Человек, как биологический вид. (Лекция 1)
Человек, как биологический вид. (Лекция 1) Обмен веществ и превращение энергии в клетке
Обмен веществ и превращение энергии в клетке Мои наблюдения за деревьями и за изменениями их внешнего вида. 5 класс
Мои наблюдения за деревьями и за изменениями их внешнего вида. 5 класс Дыхательная система
Дыхательная система Птицы. Отряд Совообразные
Птицы. Отряд Совообразные Биология 9 классы 15.04 Кочетова М И
Биология 9 классы 15.04 Кочетова М И Грибы. Общая характеристика грибов. Шляпочные грибы
Грибы. Общая характеристика грибов. Шляпочные грибы Сравнительная характеристика семейств
Сравнительная характеристика семейств Бактериология. Классификация бактерий. Морфология бактерий и методы её изучения. (Лекция 9)
Бактериология. Классификация бактерий. Морфология бактерий и методы её изучения. (Лекция 9) Типы питания живых организмов
Типы питания живых организмов Презентация к уроку биологии в 9 классе
Презентация к уроку биологии в 9 классе Проблемный вопрос: вирус живое существо или вещество? 10 класс
Проблемный вопрос: вирус живое существо или вещество? 10 класс Строение человека. Система кровообращения
Строение человека. Система кровообращения Animals in Russia
Animals in Russia Тип Кишечнополостные
Тип Кишечнополостные Ядовитые растения. Первая помощь, при отравлении ядовитыми растениями
Ядовитые растения. Первая помощь, при отравлении ядовитыми растениями Дыхание растений, животных, человека
Дыхание растений, животных, человека Класс Млекопитающие или Звери. Отряды: Однопроходные, Сумчатые, Насекомоядные, Рукокрылые
Класс Млекопитающие или Звери. Отряды: Однопроходные, Сумчатые, Насекомоядные, Рукокрылые Обмен простых белков. Образование биологически активных аминов
Обмен простых белков. Образование биологически активных аминов Гормональная регуляция обмена веществ
Гормональная регуляция обмена веществ Генная и клеточная инженерия
Генная и клеточная инженерия Эндоплазматическая сеть
Эндоплазматическая сеть Тварини минулого
Тварини минулого Что значит вода для человека
Что значит вода для человека