- Дыхательная цепь или Цепь переноса электронов. Тема 5

Содержание
- 2. Дыхательная цепь В 1961 – 1966 г. г. английский биохимик П. Митчел описал теорию окислительного фосфорилирования,
- 3. Дыхательная цепь Структура и биологическое значение митохондрий Тканевое дыхание происходит в митохондриях. Митохондрии находятся в цитоплазме
- 4. Хемиосмотическая теория Окислительного фосфорилирования Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных кислот, спиртов
- 5. Механизм окислительного фосфорилирования По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных комплексов, включающих множество ферментов.
- 6. В целом работа дыхательной цепи заключается в следующем: 1) Образующиеся в реакциях катаболизма НАДН и ФАДН2
- 7. Принцип работы дыхательной цепи 5) Протоны Н+ стремятся обратно в матрикс и проходят через АТФ-синтазу. 6)При
- 8. Всего цепь переноса электронов включает в себя около 40 разнообразных белков, которые организованы в 4 больших
- 9. АТФ-синтаза
- 10. Комплекс I. НАД-коэнзим Q редуктаза, содержит НАД дегидрогеназу, с простетической группой ФМН, и 5 FeS-(железосерных) белков
- 11. Комплекс II. Сукцинат-коэнзим Q редуктаза. Содержит ФАД-зависимую сукцинатдегидрогеназу, ассоциированную с железосерными белками и катализирует перенос электронов
- 12. Комплекс III. КоQН2-цитохром-С-оксидоредуктаза. Состоит из цитохромов b и c и одного железосерного белка. Комплекс осуществляет перенос
- 13. Комплекс IV. Цитохром c оксидаза. Состоит из двух цитохромов (а и а3), (которые обозначаются как цитохромоксидаза),
- 14. Общие принцип окислительного фосфорилирования
- 15. Блок-схема дыхательной цепи
- 16. Первый комплекс. 1 комплекс - НАДН-КоQ-оксидоредуктаза Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН, 22
- 17. Второй комплекс 2 комплекс - ФАД-зависимые дегидрогеназы Данный комплекс как таковой не существует, его выделение условно.
- 18. Третий комплекс 3 комплекс - КоQ-цитохром с-оксидоредуктаза Данный комплекс включает цитохромы b и c1. Кроме цитохромов
- 19. Четвёртый комплекс 4 комплекс. - Цитохром с-кислород-оксидоредуктаза В этом комплексе находятся цитохромы а и а3, он
- 20. Пятый комплекс 5 комплекс – это фермент АТФ-синтаза, состоящий из множества белковых цепей, подразделенных на две
- 21. Строение дыхательной цепи и механизм окислительного фосфорилирования
- 22. Механизм окислительного фосфорилирования На основании строения и функций компонентов дыхательной цепи предложен механизм окислительного фосфорилирования: 1)
- 23. Механизм окислительного фосфорилирования (продолжение) 3) На внутренней мембране митохондрий атомы водорода (от НАДН и ФАДН2) передают
- 24. Використання енергії електронів Спрощено сказане можна представити у вигляді рівняння:
- 25. Участки сопряжения 5) Перенос ионов водорода через мембрану (выкачивание) происходит не случайно, а в строго определенных
- 26. Участки сопряжения Участки сопряжения представлены I, III, IV комплексами дыхательных ферментов. В результате работы этих комплексов
- 27. Наработка АТФ 6. Как завершение всех предыдущих событий и необходимый их результат происходит наработка АТФ: ионы
- 28. На клеточное дыхание можно влиять В клетке часто может создаваться ситуация, когда реакции окислительного фосфорилирования идут
- 29. Гипоэнергетические состояния Причины: гиповитаминозы экзогенные и/или эндогенные – снижается скорость и эффективность окислительных реакций. Возникает обычно
- 30. Разобщители окисления и фосфорилирования К разобщителям в первую очередь относят "протонофоры" – вещества переносящие ионы водорода.
- 31. Ингибиторы ферментов дыхательной цепи Ряд веществ может ингибировать ферменты дыхательной цепи и блокировать движение электронов от
- 33. Скачать презентацию

Дыхательная цепь
В 1961 – 1966 г. г. английский биохимик П. Митчел
Дыхательная цепь
В 1961 – 1966 г. г. английский биохимик П. Митчел

Дыхательная цепь
Структура и биологическое значение митохондрий
Тканевое дыхание происходит в митохондриях. Митохондрии
Дыхательная цепь
Структура и биологическое значение митохондрий
Тканевое дыхание происходит в митохондриях. Митохондрии

Хемиосмотическая теория
Окислительного фосфорилирования
Молекулы НАДН и ФАДН2, образуемые в реакциях окисления
Хемиосмотическая теория
Окислительного фосфорилирования
Молекулы НАДН и ФАДН2, образуемые в реакциях окисления

Механизм окислительного фосфорилирования
По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных
Механизм окислительного фосфорилирования
По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных

В целом работа дыхательной цепи заключается в следующем:
1) Образующиеся в реакциях
В целом работа дыхательной цепи заключается в следующем:
1) Образующиеся в реакциях

Принцип работы дыхательной цепи
5) Протоны Н+ стремятся обратно в матрикс и
Принцип работы дыхательной цепи
5) Протоны Н+ стремятся обратно в матрикс и

Всего цепь переноса электронов включает в себя около 40 разнообразных белков,
Всего цепь переноса электронов включает в себя около 40 разнообразных белков,
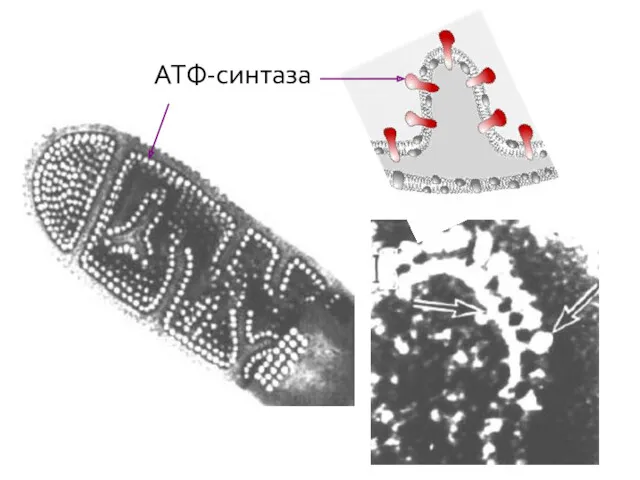
АТФ-синтаза
АТФ-синтаза
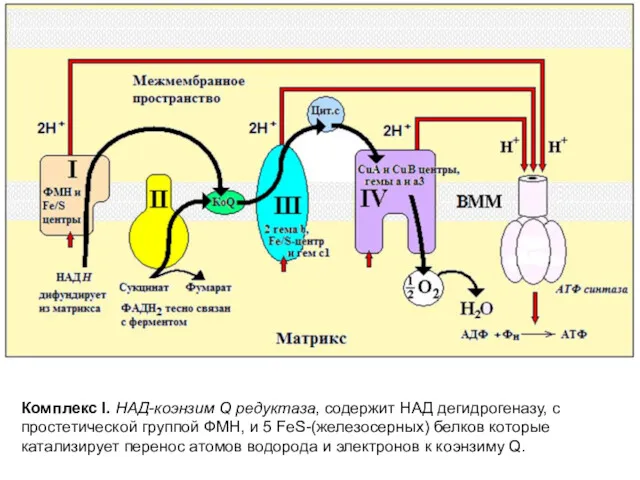
Комплекс I. НАД-коэнзим Q редуктаза, содержит НАД дегидрогеназу, с простетической группой
Комплекс I. НАД-коэнзим Q редуктаза, содержит НАД дегидрогеназу, с простетической группой
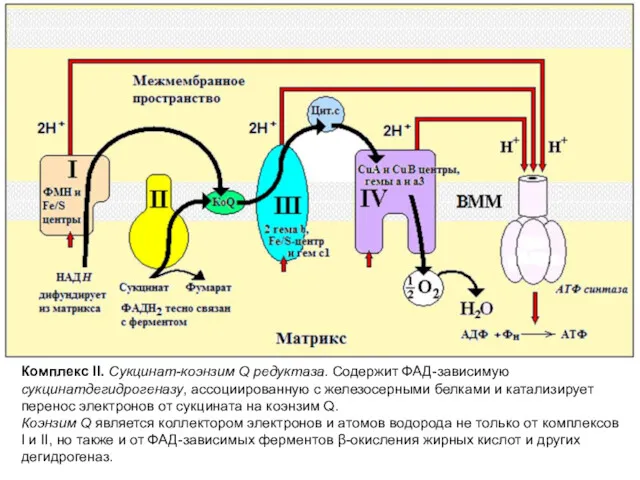
Комплекс II. Сукцинат-коэнзим Q редуктаза. Содержит ФАД-зависимую сукцинатдегидрогеназу, ассоциированную с железосерными
Комплекс II. Сукцинат-коэнзим Q редуктаза. Содержит ФАД-зависимую сукцинатдегидрогеназу, ассоциированную с железосерными
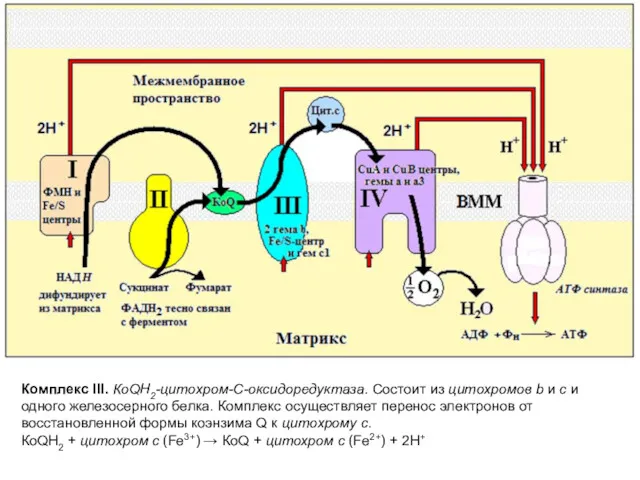
Комплекс III. КоQН2-цитохром-С-оксидоредуктаза. Состоит из цитохромов b и c и одного
Комплекс III. КоQН2-цитохром-С-оксидоредуктаза. Состоит из цитохромов b и c и одного
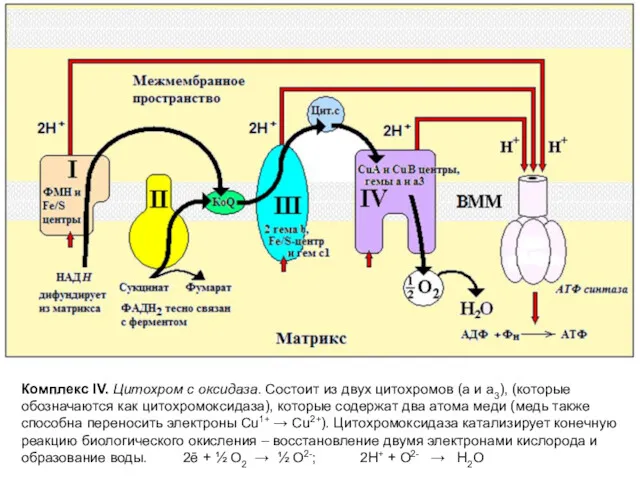
Комплекс IV. Цитохром c оксидаза. Состоит из двух цитохромов (а и
Комплекс IV. Цитохром c оксидаза. Состоит из двух цитохромов (а и
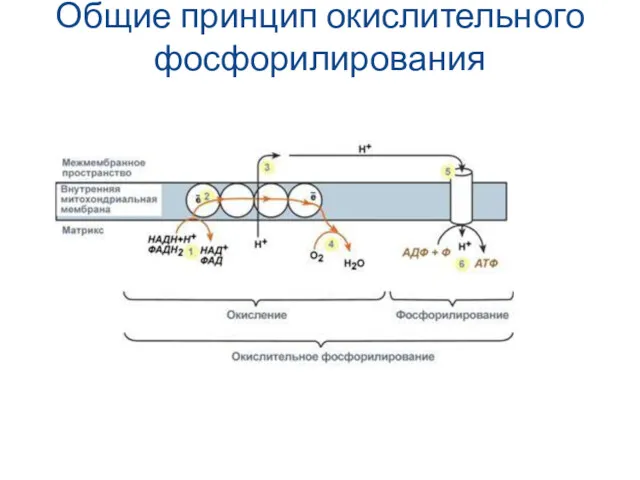
Общие принцип окислительного фосфорилирования
Общие принцип окислительного фосфорилирования
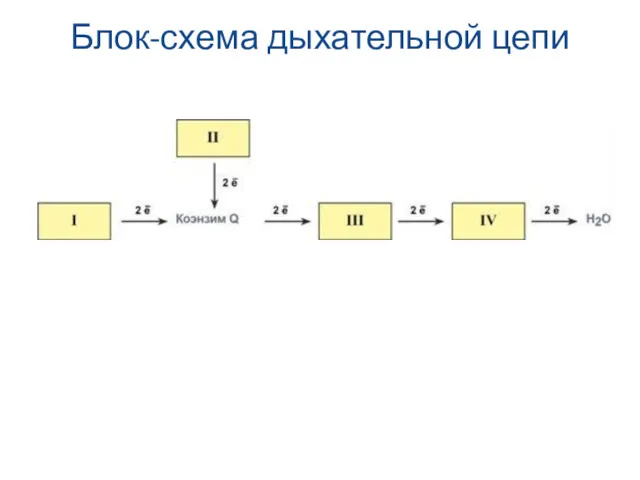
Блок-схема дыхательной цепи
Блок-схема дыхательной цепи

Первый комплекс.
1 комплекс - НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН,
Первый комплекс.
1 комплекс - НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН,

Второй комплекс
2 комплекс - ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует,
Второй комплекс
2 комплекс - ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует,

Третий комплекс
3 комплекс - КоQ-цитохром с-оксидоредуктаза
Данный комплекс включает цитохромы b и
Третий комплекс
3 комплекс - КоQ-цитохром с-оксидоредуктаза
Данный комплекс включает цитохромы b и

Четвёртый комплекс
4 комплекс. - Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а
Четвёртый комплекс
4 комплекс. - Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а

Пятый комплекс
5 комплекс – это фермент АТФ-синтаза, состоящий из множества белковых
Пятый комплекс
5 комплекс – это фермент АТФ-синтаза, состоящий из множества белковых
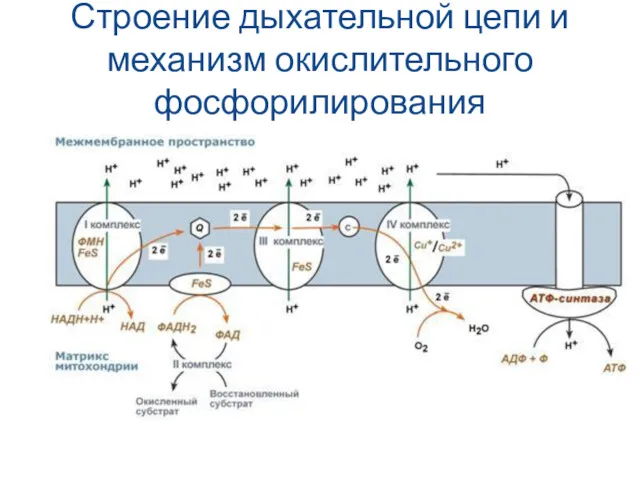
Строение дыхательной цепи и механизм окислительного фосфорилирования
Строение дыхательной цепи и механизм окислительного фосфорилирования

Механизм окислительного фосфорилирования
На основании строения и функций компонентов дыхательной цепи предложен
Механизм окислительного фосфорилирования
На основании строения и функций компонентов дыхательной цепи предложен

Механизм окислительного фосфорилирования (продолжение)
3) На внутренней мембране митохондрий атомы водорода (от
Механизм окислительного фосфорилирования (продолжение)
3) На внутренней мембране митохондрий атомы водорода (от

Використання енергії електронів
Спрощено сказане можна представити у вигляді рівняння:
Використання енергії електронів
Спрощено сказане можна представити у вигляді рівняння:

Участки сопряжения
5) Перенос ионов водорода через мембрану (выкачивание) происходит не случайно,
Участки сопряжения
5) Перенос ионов водорода через мембрану (выкачивание) происходит не случайно,

Участки сопряжения
Участки сопряжения представлены I, III, IV комплексами дыхательных ферментов. В
Участки сопряжения
Участки сопряжения представлены I, III, IV комплексами дыхательных ферментов. В

Наработка АТФ
6. Как завершение всех предыдущих событий и необходимый их результат
Наработка АТФ
6. Как завершение всех предыдущих событий и необходимый их результат

На клеточное дыхание можно влиять
В клетке часто может создаваться ситуация,
На клеточное дыхание можно влиять
В клетке часто может создаваться ситуация,

Гипоэнергетические состояния
Причины:
гиповитаминозы экзогенные и/или эндогенные – снижается скорость и эффективность окислительных реакций.
Гипоэнергетические состояния
Причины:
гиповитаминозы экзогенные и/или эндогенные – снижается скорость и эффективность окислительных реакций.

Разобщители окисления и фосфорилирования
К разобщителям в первую очередь относят "протонофоры" –
Разобщители окисления и фосфорилирования
К разобщителям в первую очередь относят "протонофоры" –

Ингибиторы ферментов дыхательной цепи
Ряд веществ может ингибировать ферменты дыхательной цепи и
Ингибиторы ферментов дыхательной цепи
Ряд веществ может ингибировать ферменты дыхательной цепи и
 Газообмен рыб
Газообмен рыб Хвощи. Внешний вид хвоща
Хвощи. Внешний вид хвоща Принципы и механизмы регуляции онтогенеза
Принципы и механизмы регуляции онтогенеза Урок-презентция по биологи 8 класс Строение клетки
Урок-презентция по биологи 8 класс Строение клетки Турбота про потомство
Турбота про потомство Нервная система
Нервная система Растительные ткани
Растительные ткани Стебель, его значение и строение. Многообразие стеблей
Стебель, его значение и строение. Многообразие стеблей Дыхание. Органы дыхательной системы
Дыхание. Органы дыхательной системы Мир комнатных растений. Окружающий мир 2 класс
Мир комнатных растений. Окружающий мир 2 класс Годовой цикл жизни земноводных
Годовой цикл жизни земноводных Всё о Кушнаренковском винограде
Всё о Кушнаренковском винограде Микроорганизмдердің морфологиясы
Микроорганизмдердің морфологиясы Онтогенез. Эмбриогенез. Постэмбриогенез
Онтогенез. Эмбриогенез. Постэмбриогенез Дикие животные Г. Бирска
Дикие животные Г. Бирска Презентация к Дню космонавтики Что едят космонавты?
Презентация к Дню космонавтики Что едят космонавты? Эволюция дыхательной системы
Эволюция дыхательной системы Терінің қосалқы мүшелері: тері және май бездері, шаш, тырнақ
Терінің қосалқы мүшелері: тері және май бездері, шаш, тырнақ Что нужно знать улитководу
Что нужно знать улитководу Нуклеиновые кислоты. ДНК, РНК
Нуклеиновые кислоты. ДНК, РНК Химический состав клетки
Химический состав клетки Загадочный Иван-чай или чудесное превращение Кипрея
Загадочный Иван-чай или чудесное превращение Кипрея Систематика растений
Систематика растений Обмен веществ и энергии в клетке
Обмен веществ и энергии в клетке Сравнение характеристик процессов фотосинтеза и дыхания
Сравнение характеристик процессов фотосинтеза и дыхания Анатомия, физиология и гигиена сенсорных систем
Анатомия, физиология и гигиена сенсорных систем Ядовитые растения Крыма
Ядовитые растения Крыма История развития экологических связей человечества
История развития экологических связей человечества