- Эндокринная система рыб. Лекция 3

Содержание
- 2. Эндокринная система часть системы нейрогуморальной регуляции, прошедшей длинный эволюционный путь. системе гуморальная регуляция (в исто-рическом плане)
- 3. Нервная и гуморальная системы функционируют как единое целое. Взаимодействие между ними осуществляется на трех уровнях: клеточном,
- 4. Примером взаимодействия на клеточном уровне может служить работа нейрохимического синапса
- 5. Межорганное взаимодействие иллюстрирует работа гипоталамо-гипофизарно-адреналовой системы Афферентация рецепторного аппарата приводит в возбуждение определенные структуры таламуса. В
- 7. Примером взаимодействия нервной и гуморальной систем на организменном уровне является реакция адаптации
- 8. Гормоны Химическая природа гормонов различна. Это и стероиды, и пептиды, и производные аминокислот, и производные жирных
- 9. Стероидные гормоны кортизол, тестостерон, эстрадиол, прогестерон образуются из холестерина. К этой же группе специалисты причисляют арахидоновую
- 10. Рецепторы этих гормонов обнаруживают в цитоплазме клеток-мишеней. Белки-рецепторы одновременно выступают и транспортным средством в пределах клетки,
- 11. В ядре стероиды взаимодействуют с ДНК и вызывают синтез матричной РНК, затем рибосомальной РНК и полисом
- 13. Белковые гормоны (СТГ, ТТГ, ФСГ, ЛГ, пролактин, инсулин и др.) имеют слишком крупные размеры молекулы и
- 14. Этому предшествует этап синтеза ряда химических веществ называемых месенджерами, под влиянием комплекса из гормона и белка-рецептора
- 16. цАМФ
- 17. цАМФ-зависимыми гормонами являются АКЛТ, ТТГ, ФСГ, ЛГ, адреналин (через р-рецепторы). Кальцийзависимыми считаются окситоцин, вазопрессин, гастрин, катехоламины
- 18. эндокринной системе рыб больше подходит название "паракринная система", так как истинной железой можно считать только гипофиз.
- 19. Гипофиз В отличие от гипофиза высших животных гипофиз рыб имеет упрощенное строение: он не имеет четко
- 20. Передняя доля гипофиза (аденогипофиз) у рыб может занимать большую часть железы. В этом случае его подразделяют
- 21. Гипофиз костистых рыб: 1, 2- аденогипофиз; 3- промежуточная доля. 4- нейрогипофиз
- 22. Филогенетические корни аденогипофиза и нейрогипофиза различны. Аденогипофиз берет свое начало от органов пищеварения. В эмбриогенезе он
- 23. Из гипофиза у рыб выделен тот же набор гормонов, что и у высших животных. Однако их
- 24. Пролактин имеет клетки-мишени в ряде органов: почках, жабрах, желудочно-кишечном тракте, плавательном пузыре, коже. Играет большую роль
- 25. (ФСГ, ЛГ) гипофиза регулируют сперматогенез у самцов и овогенез у самок рыб. Влияют на половое поведение.
- 26. Меланофорный гормон (МФГ) метааденогипофиза у рыб, как и у других животных, регулирует пигментный обмен. Секреторная активность
- 27. Щитовидная железа у рыб не оправдывает своего названия, заимствованного у высших позвоночных. У рыб под "щитовидной
- 28. Функция тиреоидных гормонов у рыб до конца неясна. Тироксин не дает у рыб такого однозначного эффекта
- 29. Интерреналовые тельца (функциональный аналог корковой части надпочечников) невооруженным глазом увидеть невозможно так как они представляют собой
- 30. Кортизол обеспечивает адаптацию организма рыб в стрессовых ситуациях. Он стимулирует глюконеогенез в печени, поддерживает на постоянном
- 31. Хромаффиновые железы - аналоги мозгового слоя надпочечников - секретируют в кровь катехоламины. Хромаффиновые клетки можно обнаружить
- 32. Ренин-ангиотензиновая система у рыб представлена окологломелулярными клетками, т. е. является еще одной эндокринной системой в составе
- 33. Улътимобранхиальная железа - структура, обнаруженная на мембране между сердцем и печенью, имеет вид белесой полоски. Клетки,
- 34. Урофиз - нейросекреторная зона концевого отдела спинного мозга - был обнаружен при изучении гистоструктуры спинного мозга
- 35. Поджелудочная железа - железа смешанного типа. Она представляет собой компактный орган только у хрящевых рыб. У
- 36. Гонады как железы смешанной секреции имеют хорошо развитый эндосекреторный аппарат, который вырабатывает половые стероиды. В ястыках
- 38. Скачать презентацию

Эндокринная система
часть системы нейрогуморальной регуляции, прошедшей длинный эволюционный путь.
системе гуморальная
Эндокринная система
часть системы нейрогуморальной регуляции, прошедшей длинный эволюционный путь.
системе гуморальная

Нервная и гуморальная системы функционируют как единое целое. Взаимодействие между ними
Нервная и гуморальная системы функционируют как единое целое. Взаимодействие между ними

Примером взаимодействия на клеточном уровне может служить работа нейрохимического синапса
Примером взаимодействия на клеточном уровне может служить работа нейрохимического синапса

Межорганное взаимодействие иллюстрирует работа гипоталамо-гипофизарно-адреналовой системы
Афферентация рецепторного аппарата приводит в возбуждение
Межорганное взаимодействие иллюстрирует работа гипоталамо-гипофизарно-адреналовой системы
Афферентация рецепторного аппарата приводит в возбуждение


Примером взаимодействия нервной и гуморальной систем на организменном уровне является реакция
Примером взаимодействия нервной и гуморальной систем на организменном уровне является реакция

Гормоны
Химическая природа гормонов различна. Это и стероиды, и пептиды, и производные
Гормоны
Химическая природа гормонов различна. Это и стероиды, и пептиды, и производные

Стероидные гормоны
кортизол, тестостерон, эстрадиол, прогестерон
образуются из холестерина.
К этой же
Стероидные гормоны
кортизол, тестостерон, эстрадиол, прогестерон
образуются из холестерина.
К этой же

Рецепторы этих гормонов обнаруживают в цитоплазме клеток-мишеней.
Белки-рецепторы одновременно выступают и
Рецепторы этих гормонов обнаруживают в цитоплазме клеток-мишеней.
Белки-рецепторы одновременно выступают и

В ядре стероиды
взаимодействуют с ДНК и вызывают синтез матричной РНК, затем
В ядре стероиды
взаимодействуют с ДНК и вызывают синтез матричной РНК, затем


Белковые гормоны
(СТГ, ТТГ, ФСГ, ЛГ, пролактин, инсулин и др.) имеют слишком
Белковые гормоны
(СТГ, ТТГ, ФСГ, ЛГ, пролактин, инсулин и др.) имеют слишком

Этому предшествует этап синтеза ряда химических веществ называемых месенджерами, под влиянием
Этому предшествует этап синтеза ряда химических веществ называемых месенджерами, под влиянием

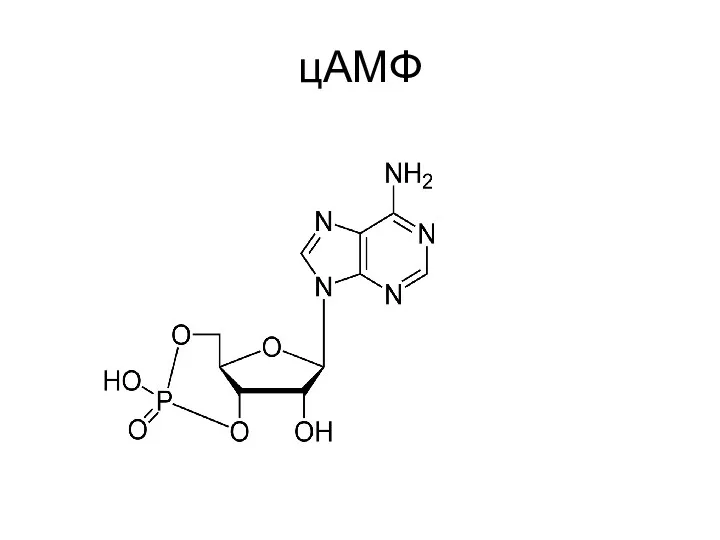
цАМФ
цАМФ

цАМФ-зависимыми гормонами являются АКЛТ, ТТГ, ФСГ, ЛГ, адреналин (через р-рецепторы).
Кальцийзависимыми считаются
цАМФ-зависимыми гормонами являются АКЛТ, ТТГ, ФСГ, ЛГ, адреналин (через р-рецепторы).
Кальцийзависимыми считаются

эндокринной системе рыб
больше подходит название "паракринная система", так как истинной железой
эндокринной системе рыб
больше подходит название "паракринная система", так как истинной железой

Гипофиз
В отличие от гипофиза высших животных гипофиз рыб имеет упрощенное строение:
Гипофиз
В отличие от гипофиза высших животных гипофиз рыб имеет упрощенное строение:

Передняя доля гипофиза (аденогипофиз) у рыб может занимать большую часть железы.
Передняя доля гипофиза (аденогипофиз) у рыб может занимать большую часть железы.
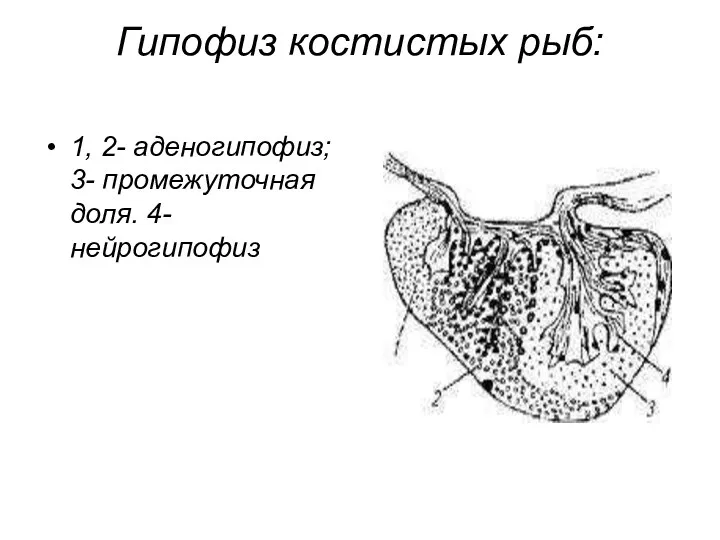
Гипофиз костистых рыб:
1, 2- аденогипофиз; 3- промежуточная доля. 4- нейрогипофиз
Гипофиз костистых рыб:
1, 2- аденогипофиз; 3- промежуточная доля. 4- нейрогипофиз

Филогенетические корни аденогипофиза и нейрогипофиза различны. Аденогипофиз берет свое начало от
Филогенетические корни аденогипофиза и нейрогипофиза различны. Аденогипофиз берет свое начало от

Из гипофиза у рыб выделен тот же набор гормонов, что и
Из гипофиза у рыб выделен тот же набор гормонов, что и

Пролактин имеет клетки-мишени в ряде органов: почках, жабрах, желудочно-кишечном тракте, плавательном
Пролактин имеет клетки-мишени в ряде органов: почках, жабрах, желудочно-кишечном тракте, плавательном

(ФСГ, ЛГ) гипофиза регулируют сперматогенез у самцов и овогенез у самок
(ФСГ, ЛГ) гипофиза регулируют сперматогенез у самцов и овогенез у самок

Меланофорный гормон (МФГ)
метааденогипофиза у рыб, как и у других животных, регулирует
Меланофорный гормон (МФГ)
метааденогипофиза у рыб, как и у других животных, регулирует

Щитовидная железа
у рыб не оправдывает своего названия, заимствованного у высших позвоночных.
Щитовидная железа
у рыб не оправдывает своего названия, заимствованного у высших позвоночных.

Функция тиреоидных гормонов у рыб до конца неясна. Тироксин не дает
Функция тиреоидных гормонов у рыб до конца неясна. Тироксин не дает

Интерреналовые тельца (функциональный аналог корковой части надпочечников) невооруженным глазом увидеть невозможно
Интерреналовые тельца (функциональный аналог корковой части надпочечников) невооруженным глазом увидеть невозможно

Кортизол обеспечивает адаптацию организма рыб в стрессовых ситуациях. Он стимулирует глюконеогенез
Кортизол обеспечивает адаптацию организма рыб в стрессовых ситуациях. Он стимулирует глюконеогенез

Хромаффиновые железы - аналоги мозгового слоя надпочечников - секретируют в кровь
Хромаффиновые железы - аналоги мозгового слоя надпочечников - секретируют в кровь

Ренин-ангиотензиновая система
у рыб представлена окологломелулярными клетками, т. е. является еще одной
Ренин-ангиотензиновая система
у рыб представлена окологломелулярными клетками, т. е. является еще одной

Улътимобранхиальная железа
- структура, обнаруженная на мембране между сердцем и печенью, имеет
Улътимобранхиальная железа
- структура, обнаруженная на мембране между сердцем и печенью, имеет

Урофиз
- нейросекреторная зона концевого отдела спинного мозга - был обнаружен при
Урофиз
- нейросекреторная зона концевого отдела спинного мозга - был обнаружен при

Поджелудочная железа - железа смешанного типа. Она представляет собой компактный орган
Поджелудочная железа - железа смешанного типа. Она представляет собой компактный орган

Гонады как железы смешанной секреции имеют хорошо развитый эндосекреторный аппарат, который
Гонады как железы смешанной секреции имеют хорошо развитый эндосекреторный аппарат, который
 Презентация Пресмыкающиеся
Презентация Пресмыкающиеся Адаптация к физическим упражнениям
Адаптация к физическим упражнениям Кто живет в Африке
Кто живет в Африке Значение знаний об особенностях строения и жизнедеятельности организма
Значение знаний об особенностях строения и жизнедеятельности организма Черепаха в моём доме. Исследовательский проект
Черепаха в моём доме. Исследовательский проект Обеспечение человечества продовольствием
Обеспечение человечества продовольствием Значение водорослей в природе и жизни человека
Значение водорослей в природе и жизни человека Эволюция кровеносной системы
Эволюция кровеносной системы Дикие и домашние животные
Дикие и домашние животные Мейоз. Типы и фазы мейоза
Мейоз. Типы и фазы мейоза Биологи во время Великой Отечественной войны
Биологи во время Великой Отечественной войны Биотехнологии. Генная инженерия
Биотехнологии. Генная инженерия Гормональна діяльність ендокринних залоз
Гормональна діяльність ендокринних залоз Фитопатогенные нематоды. (Лекция 5)
Фитопатогенные нематоды. (Лекция 5) Половое размножение. Оплодотворение, его значение (10 класс)
Половое размножение. Оплодотворение, его значение (10 класс) Пищевые добавки в продуктах питания и их влияние на здоровье человека
Пищевые добавки в продуктах питания и их влияние на здоровье человека презентация на тему Движущие силы эволюции
презентация на тему Движущие силы эволюции Царства живой природы
Царства живой природы 22_01_8_klass (2)
22_01_8_klass (2) Органический мир докембрия
Органический мир докембрия Как зимой помочь птицам (урок окружающего мира, 1 класс)
Как зимой помочь птицам (урок окружающего мира, 1 класс) биотические связи в природе
биотические связи в природе Плеснивые грибы и грибы паразиты
Плеснивые грибы и грибы паразиты Бобр. Жизнь бобров
Бобр. Жизнь бобров Белковые катализаторы ферменты
Белковые катализаторы ферменты Витамины. Определение
Витамины. Определение Взаємозв'язки організмів в екосистемах. Стабільність екосистем
Взаємозв'язки організмів в екосистемах. Стабільність екосистем Селекция
Селекция