- Ферменты. Стадии ферментативного катализа

Содержание
- 2. Ферменты По своей функции ферменты являются биологическими катализаторами. Сущность действия ферментов, так же как неорганических катализаторов,
- 3. Ферменты Сходство ферментов и неорганических катализаторов заключается в следующем: Катализируют только энергетически возможные реакции. Не изменяют
- 4. Ферменты Отличия ферментов от неорганических катализаторов позволяют им «работать» внутри живых тел и делают их изучение
- 5. Стадии ферментативного катализа В ферментативной реакции можно выделить следующие этапы: Присоединение субстрата (S) к ферменту (E)
- 6. Механизмы катализа 1. Кислотно-основной катализ – в активном центре фермента находятся группы специфичных аминокислотных остатков, которые
- 7. Типы ферментативных реакций В зависимости от последовательности взаимодействия между ферментом и реактивами выделяют следующие типы ферментативных
- 8. Схема случайных реакций В случайных реакциях последовательность взаимодействия между субстратом и реагентами не имеет значения:
- 9. Схема последовательных реакций В этом случае к ферменту последовательно присоединяются субстраты А и В, образуя "тройной
- 10. Схема «пинг-понг» В этом случае фермент сначала взаимодействует с субстратом А, отбирая у него какие либо
- 11. Строение ферментов Все ферменты являются белками, поэтому, они, как и остальные белки по составу делятся на
- 12. Строение ферментов В составе фермента выделяют две области – активный центр и аллостерический центр. Активный центр
- 13. Схема активного центра
- 14. Строение ферментов Аллостерический центр – центр регуляции активности фермента, который пространственно отделен от активного центра и
- 15. Строение ферментов Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся в разных субъединицах.
- 16. Мультиферментные комплексы В мультиферментном комплексе несколько ферментов прочно связаны между собой в единый комплекс и осуществляют
- 17. Специфичность Ферменты крайне избирательно запускают химические реакции, из всех возможных вариантов направления фермент выбирает один нужный
- 18. Механизм специфичности ферментов В общем случае специфичность обусловлена взаимодействием определенных функциональных групп субстрата и фермента. На
- 19. Механизм специфичности ферментов Согласно другой теории (теория Кошланда, «рука-перчатка») присоединение субстрата вызывает изменения в молекуле фермента,
- 20. Регуляция активности фермента В течении времени активность фермента внутри клетки непостоянна. Ферменты реагируют на изменение внешних
- 21. Кинетические способы регуляции По закону действующих масс скорость пропорциональна произведению концентраций. Уменьшение концентраций приводит к уменьшению
- 22. Аллостерическая регуляция Аллостерические ферменты построены из двух и более субъединиц: одни субъединицы содержат каталитический центр, другие
- 23. Белок-белковое взаимодействие Термин белок-белковое взаимодействие обозначает ситуацию, когда в качестве регулятора выступают не метаболиты биохимических процессов,
- 24. Ковалентная модификация Ковалентная модификация заключается в обратимом присоединении или отщеплении определенной группы, благодаря чему изменяется активность
- 25. Ингибирование В медицине активно разрабатываются и используются соединения, изменяющие активность ферментов с целью регуляции скорости метаболических
- 27. Скачать презентацию

Ферменты
По своей функции ферменты являются биологическими катализаторами. Сущность действия ферментов, так
Ферменты
По своей функции ферменты являются биологическими катализаторами. Сущность действия ферментов, так

Ферменты
Сходство ферментов и неорганических катализаторов заключается в следующем:
Катализируют только энергетически возможные
Ферменты
Сходство ферментов и неорганических катализаторов заключается в следующем:
Катализируют только энергетически возможные

Ферменты
Отличия ферментов от неорганических катализаторов позволяют им «работать» внутри живых тел
Ферменты
Отличия ферментов от неорганических катализаторов позволяют им «работать» внутри живых тел

Стадии ферментативного катализа
В ферментативной реакции можно выделить следующие этапы:
Присоединение субстрата (S) к
Стадии ферментативного катализа
В ферментативной реакции можно выделить следующие этапы:
Присоединение субстрата (S) к

Механизмы катализа
1. Кислотно-основной катализ – в активном центре фермента находятся группы специфичных аминокислотных
Механизмы катализа
1. Кислотно-основной катализ – в активном центре фермента находятся группы специфичных аминокислотных

Типы ферментативных реакций
В зависимости от последовательности взаимодействия между ферментом и реактивами
Типы ферментативных реакций
В зависимости от последовательности взаимодействия между ферментом и реактивами
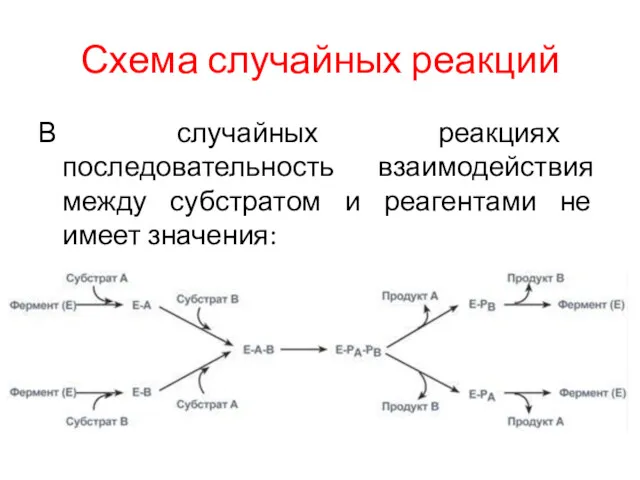
Схема случайных реакций
В случайных реакциях последовательность взаимодействия между субстратом и реагентами
Схема случайных реакций
В случайных реакциях последовательность взаимодействия между субстратом и реагентами
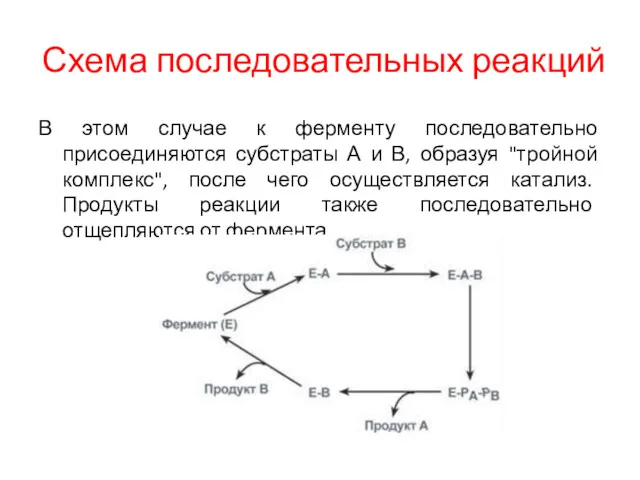
Схема последовательных реакций
В этом случае к ферменту последовательно присоединяются субстраты А
Схема последовательных реакций
В этом случае к ферменту последовательно присоединяются субстраты А
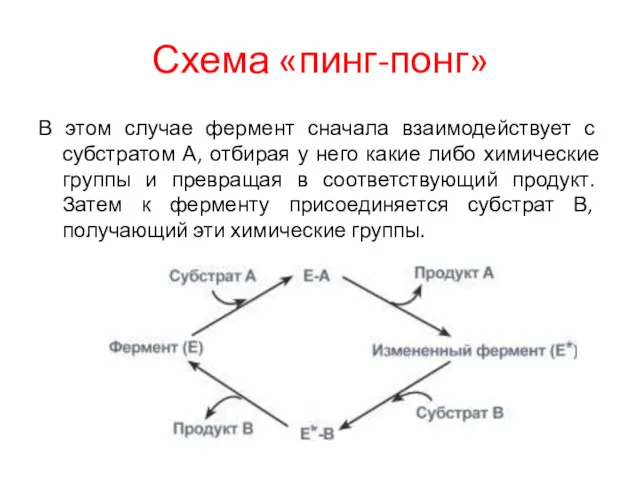
Схема «пинг-понг»
В этом случае фермент сначала взаимодействует с субстратом А, отбирая
Схема «пинг-понг»
В этом случае фермент сначала взаимодействует с субстратом А, отбирая

Строение ферментов
Все ферменты являются белками, поэтому, они, как и остальные белки
Строение ферментов
Все ферменты являются белками, поэтому, они, как и остальные белки

Строение ферментов
В составе фермента выделяют две области – активный центр и
Строение ферментов
В составе фермента выделяют две области – активный центр и
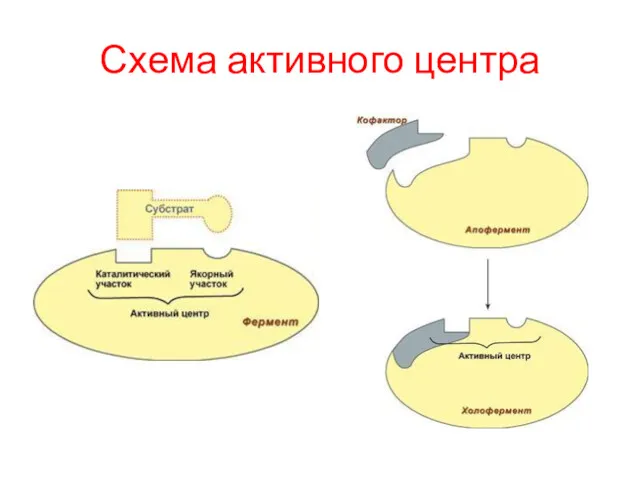
Схема активного центра
Схема активного центра

Строение ферментов
Аллостерический центр – центр регуляции активности фермента, который пространственно отделен от
Строение ферментов
Аллостерический центр – центр регуляции активности фермента, который пространственно отделен от
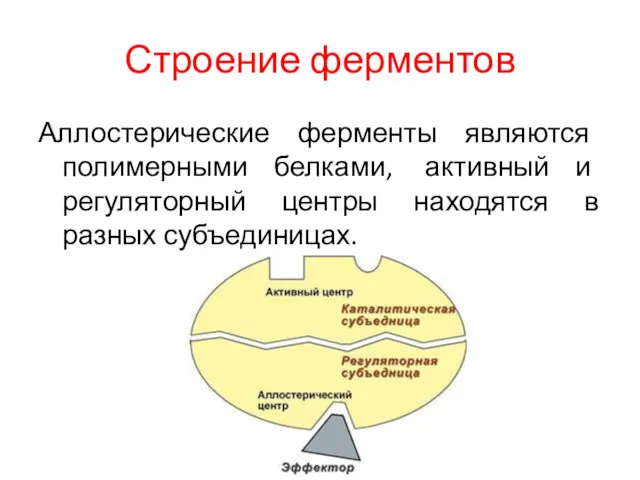
Строение ферментов
Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся
Строение ферментов
Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся
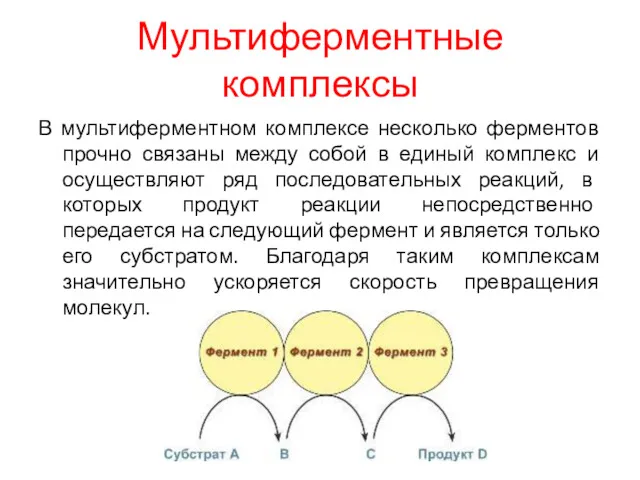
Мультиферментные комплексы
В мультиферментном комплексе несколько ферментов прочно связаны между собой в
Мультиферментные комплексы
В мультиферментном комплексе несколько ферментов прочно связаны между собой в

Специфичность
Ферменты крайне избирательно запускают химические реакции, из всех возможных вариантов направления
Специфичность
Ферменты крайне избирательно запускают химические реакции, из всех возможных вариантов направления

Механизм специфичности ферментов
В общем случае специфичность обусловлена взаимодействием определенных функциональных групп
Механизм специфичности ферментов
В общем случае специфичность обусловлена взаимодействием определенных функциональных групп
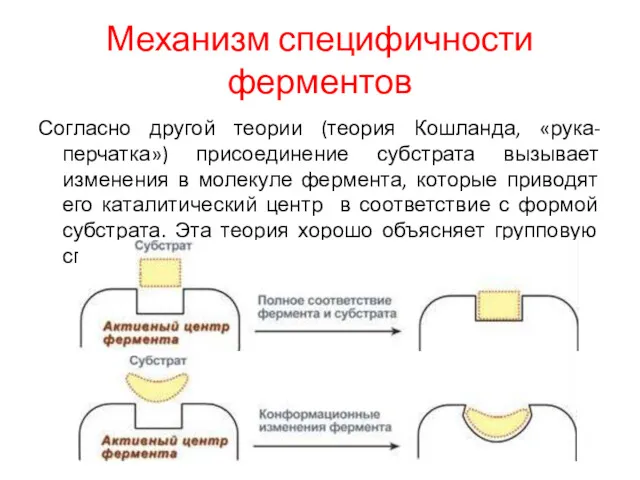
Механизм специфичности ферментов
Согласно другой теории (теория Кошланда, «рука-перчатка») присоединение субстрата вызывает
Механизм специфичности ферментов
Согласно другой теории (теория Кошланда, «рука-перчатка») присоединение субстрата вызывает

Регуляция активности фермента
В течении времени активность фермента внутри клетки непостоянна. Ферменты
Регуляция активности фермента
В течении времени активность фермента внутри клетки непостоянна. Ферменты

Кинетические способы регуляции
По закону действующих масс скорость пропорциональна произведению концентраций. Уменьшение
Кинетические способы регуляции
По закону действующих масс скорость пропорциональна произведению концентраций. Уменьшение

Аллостерическая регуляция
Аллостерические ферменты построены из двух и более субъединиц: одни субъединицы содержат
Аллостерическая регуляция
Аллостерические ферменты построены из двух и более субъединиц: одни субъединицы содержат

Белок-белковое взаимодействие
Термин белок-белковое взаимодействие обозначает ситуацию, когда в качестве регулятора выступают
Белок-белковое взаимодействие
Термин белок-белковое взаимодействие обозначает ситуацию, когда в качестве регулятора выступают

Ковалентная модификация
Ковалентная модификация заключается в обратимом присоединении или отщеплении определенной группы,
Ковалентная модификация
Ковалентная модификация заключается в обратимом присоединении или отщеплении определенной группы,

Ингибирование
В медицине активно разрабатываются и используются соединения, изменяющие активность ферментов с
Ингибирование
В медицине активно разрабатываются и используются соединения, изменяющие активность ферментов с
 Проводящие пути головного и спинного мозга
Проводящие пути головного и спинного мозга Ядовитые змеи мира
Ядовитые змеи мира Проведение опыта по определению pH талой, водопроводной и дистиллированной воды с помощью лакмусовой бумаги
Проведение опыта по определению pH талой, водопроводной и дистиллированной воды с помощью лакмусовой бумаги Хемосинтез и его значение
Хемосинтез и его значение Загальна фізіологія збудливих тканин. Фізіологія м'язів і нервів
Загальна фізіологія збудливих тканин. Фізіологія м'язів і нервів Эпителиальная ткань
Эпителиальная ткань Царство Животные
Царство Животные Sleep for life
Sleep for life Кровеносная система. Кровь
Кровеносная система. Кровь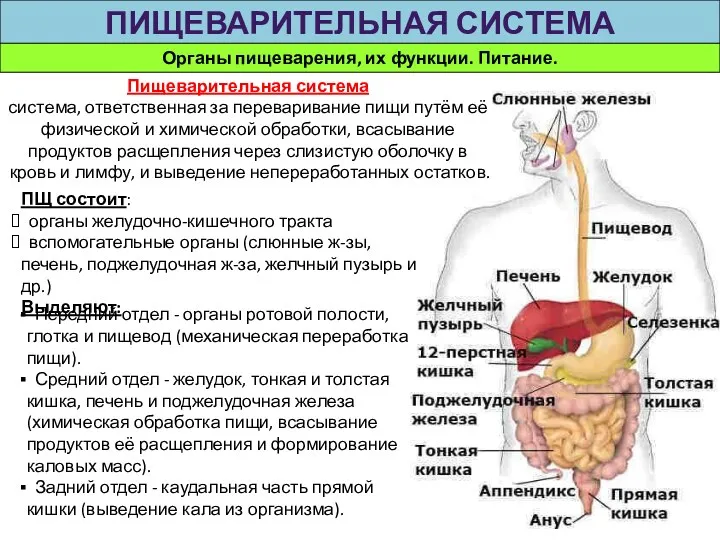 Органы пищеварения, их функции. Питание. Пищеварительная система
Органы пищеварения, их функции. Питание. Пищеварительная система Лабораторная работа. Вегетативное размножение растений
Лабораторная работа. Вегетативное размножение растений Птицы. Отряд Совообразные
Птицы. Отряд Совообразные Ядовитые растения России
Ядовитые растения России Жизнедеятельность клетки
Жизнедеятельность клетки Размножение голосеменных растений
Размножение голосеменных растений анализ учебно-воспитательного процесса
анализ учебно-воспитательного процесса Отряд совы
Отряд совы Геном - совокупность всех генов организма
Геном - совокупность всех генов организма Среда обитания организмов
Среда обитания организмов Общие вопросы миологии. Мышцы головы и шеи
Общие вопросы миологии. Мышцы головы и шеи Дубильні речовини
Дубильні речовини Растениеводство. Окружающий мир 3 класс УМК Школа России
Растениеводство. Окружающий мир 3 класс УМК Школа России Зимующие птицы
Зимующие птицы Генная регуляция поддержания популяции стволовых клеток в организме млекопитающих
Генная регуляция поддержания популяции стволовых клеток в организме млекопитающих Генетикалық код
Генетикалық код Функциональная анатомия артериальной и венозной системы
Функциональная анатомия артериальной и венозной системы Пейте дети молоко - будите здоровыми. Исследовательская работа
Пейте дети молоко - будите здоровыми. Исследовательская работа 16. Внешнее строение листа
16. Внешнее строение листа