- Физиология мышечной системы

Содержание
- 2. Потенциал действия, его фазы. Соотношение фаз возбудимости с фазами ПД.
- 3. Потенциал действия Потенциал действия — быстрое изменение мембранного потенциала во время возбуждения. ПД возникает при действии
- 4. ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ Екр Ео 1 2 3 4 5 1- местное возбуждение (начальная деполяризация) 2
- 5. 1) Местное возбуждение (локальный ответ, начальная деполяризация) Под влиянием раздражителя пороговой или сверхпороговой величины проницаемость мембраны
- 6. Критический уровень деполяризации (КУД, Екр) тот уровень, при котором деполяризация мембраны принимает регенеративный (самоусиливающийся) характер, свидетельствующий
- 7. 2) Деполяризация (восходящая фаза ПД) проницаемость мембраны для ионов натрия увеличивается в 500 раз и превышает
- 8. Деполяризация (2) В результате проникновения ионов натрия в цитоплазму и их взаимодействия с анионами разность потенциалов
- 9. 3)Реполяризация Открыты калиевые каналы, преобладает выходящий из клетки калиевый ток Происходит восстановление поляризации мембраны (из клетки
- 10. 4)Следовая деполяризация Отрицательный следовой потенциал Связана с резким снижением калиевого тока и остаточным током натрия
- 11. 5)Следовая гиперполяризация Положительный следовой потенциал Связан с преобладанием выходящего калиевого тока
- 12. СООТНОШЕНИЕ ФАЗ ПД И ВОЗБУДИМОСТИ I - фаза первичной супервозбудимости (экзальтации) II - фаза абсолютной рефрактерности
- 13. Локальный ответ возбудимость ткани повышена( фаза первичной экзальтации). Во время развития локального ответа мембранный потенциал покоя
- 14. фаза деполяризации ПД В период развития фазы деполяризации мембрана утрачивает способность отвечать возбуждением на раздражители даже
- 15. фаза реполяризации ПД Соответствует фазе относительной рефрактерности Новый ПД в этой фазе можно получить при действии
- 16. фаза следовой деполяризации соответствует фазе вторичной экзальтации Так как мембранный потенциал в эту фазу ближе к
- 17. Следовая гиперполяризация Соответствует фазе субнормальной (пониженной) возбудимости В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны),
- 18. Физиология мышц
- 19. Виды мышц • поперечно-полосатые скелетные мышцы; • поперечно-полосатая сердечная мышца; • гладкие мышцы внутренних органов, кожи,
- 20. Мышечная клетка (мышечное волокно) – вытянутой формы, многоядерная (результат слияния многих недифференцированных клеток), содержит сократительные белки
- 22. Сократительные белки мышечных клеток – филаменты тонкие актин: сокращение небулин: длина актина – Са++-кальмодулин; тропонин, тропомиозин:
- 23. саркомеры – сократительная единица м. клетки поперечная исчерченность м. Z линии – места связок саркомеров (крепление
- 26. Тонкие филаменты Закреплены в области Z линии внешне напоминает две нитки бус, закрученные в двойную спираль,
- 27. в покое места связывания миозина на актине закрыты тропомиозином, три формы тропонина внедрены через регулярные интервалы
- 29. Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку и хвост .
- 32. Пузырьки на концах продольных трубочек - терминальные цистерны - подходят очень близко к поперечным трубочкам, образуя
- 34. Саркоплазматический ретикулум форма ЭР - депо Ca2+ Ca2+ -АТФаза (закачивает кальций в СР), Кальсеквестрин (внутри СР)
- 35. рецептор рианодина в мембране цистерны связан с Ca2+-каналом. Работу канала контролируют рецепторы дигидропиридина плазмолеммы, в ответ
- 36. НЕЙРО-МЫШЕЧНЫЙ СИНАПС. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ
- 37. Структура нервно-мышечного соединения концевая пластинка двигательного нерва на мышце (пресинаптическая мембрана) концевая мышечная пластинка (постсинаптическая мембрана)
- 38. Внутри терминального участка аксона находятся везикул- пузырьки, содержащие медиатор- вещество, при помощи которого передается возбуждение. Также
- 39. Постсинаптическая мембрана часть мембраны мышечного волокна, на которой находятся специальных рецепторы, чувствительные к определенному медиатору и
- 42. Процесс передачи импульса в нервно-мышечном синапсе 1. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической
- 43. 2. Ионы Са++ поступают в цитоплазму синаптического окончания и активируют опорожнение синаптических пузырьков в синаптическую щель.
- 44. 3. Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных
- 45. 4. Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы
- 47. 6. ПКП(потенциал концевой пластинки) создает ток, который приводит к генерации потенциала действия на постсинаптической мембране.
- 48. Медиатором в нервно-мышечном синапсе является ацетилхолин. После взаимодействия АХ с рецептором специальный фермент холинэстераза расщепляет его
- 49. Два механизма высвобождения медиатора в синапсе: 1.с полным слиянием везикулы с плазмалеммой и 2.модель «поцеловал и
- 50. Полное слияние везикулы с плазмолеммой
- 51. Синапсины – семейство фосфопротеинов на поверхности синаптических пузырьков, регулируют процесс выброса нейромедиатора в дефосфорилированной форме способствуют
- 52. Три пула пузырьков в пресинаптическом пространстве: готовый к выбросу, рециклируемый, и резервный
- 53. Процессы, происходящие после генерации ПД на сарколемме ПД распространяется по сарколемме в Т-трубочки Деполяризация Т-трубочек вызывает
- 55. Теория скользящих нитей Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых
- 56. В покое в межфибриллярном пространстве низкая [Ca2+ ] 10-7 М - связывание актина и миозина блокировано
- 57. После выделения Ca2+ из СР (вследствие активации рианодиновых рецепторов) кальций связывается с тропонином С, устраняется блокирующий
- 58. Теория скольжения Хаксли Рабочий цикл поперечных мостиков: Образование поперечных мостиков между актином и миозином (энергия АТФ)
- 61. В цикле поперечных мостиков АТФ выполняет две разные роли: 1) гидролиз АТФ поставляет энергию для движения
- 63. Режимы мышечного сокращения В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по
- 64. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех
- 65. Одиночные мышечные сокращения Развивается после одного потенциала действия возникают при низкой частоте электрических импульсов при стимуляции.
- 66. Характеристика одиночного мышечного сокращения Изменение длины мышцы показано синим цветом, потенциал действия в мышце - красным,
- 67. Тетанус (лат. tetanus, от греч. tétanos — напряжение, оцепенение, судорога) При слишком частой стимуляции мышцы частотная
- 68. Во время тетанического сокращения в мышце многократно возникают ПД, обеспечивающие продолжительный выход ионов Са2+ из саркоплазматического
- 70. Зубчатый тетанус очередной импульс совпадает с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет
- 71. Гладкий тетанус следующий импульс действует на мышцу во время фазы укорочения (сокращения), в результате чего возникнет
- 74. Оптимум и пессимум частоты Амплитуда тетанического сокращения, основанного на суммации, зависит от частоты импульсов, раздражающих мышцу.
- 76. Скачать презентацию

Потенциал действия, его фазы. Соотношение фаз возбудимости с фазами ПД.
Потенциал действия, его фазы. Соотношение фаз возбудимости с фазами ПД.

Потенциал действия
Потенциал действия — быстрое изменение мембранного потенциала во время возбуждения.
ПД
Потенциал действия
Потенциал действия — быстрое изменение мембранного потенциала во время возбуждения.
ПД
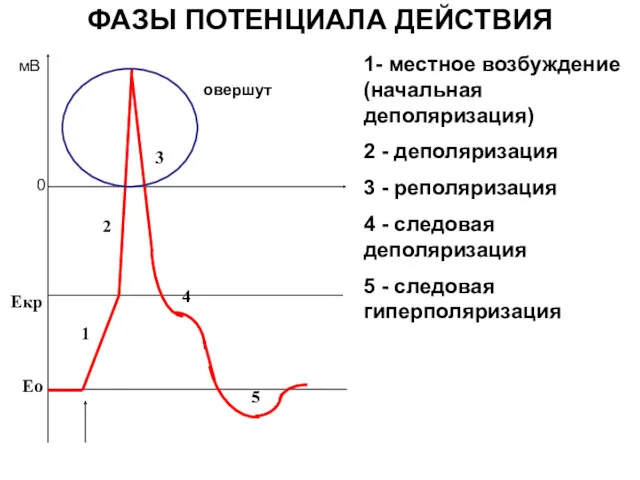
ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Екр
Ео
1
2
3
4
5
1- местное возбуждение (начальная деполяризация)
2 - деполяризация
3 - реполяризация
4
ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Екр
Ео
1
2
3
4
5
1- местное возбуждение (начальная деполяризация)
2 - деполяризация
3 - реполяризация
4

1) Местное возбуждение (локальный ответ, начальная деполяризация)
Под влиянием раздражителя пороговой или
1) Местное возбуждение (локальный ответ, начальная деполяризация)
Под влиянием раздражителя пороговой или

Критический уровень деполяризации (КУД, Екр)
тот уровень, при котором деполяризация мембраны принимает
Критический уровень деполяризации (КУД, Екр)
тот уровень, при котором деполяризация мембраны принимает
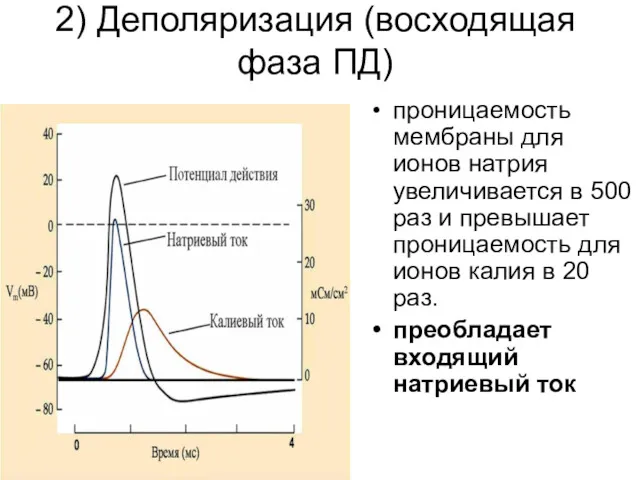
2) Деполяризация (восходящая фаза ПД)
проницаемость мембраны для ионов натрия увеличивается в
2) Деполяризация (восходящая фаза ПД)
проницаемость мембраны для ионов натрия увеличивается в

Деполяризация (2)
В результате проникновения ионов натрия в цитоплазму и их взаимодействия
Деполяризация (2)
В результате проникновения ионов натрия в цитоплазму и их взаимодействия
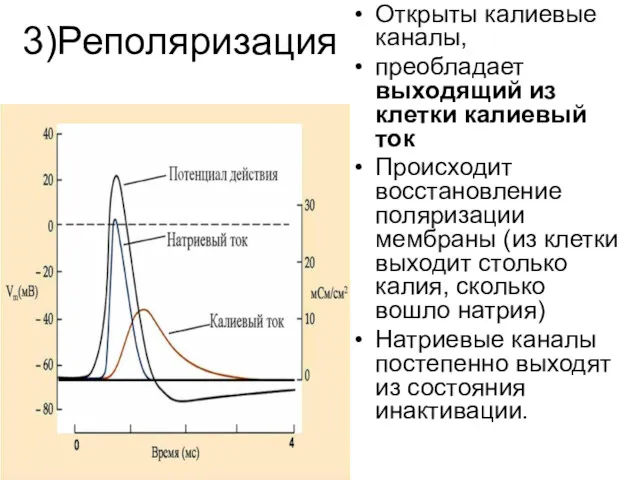
3)Реполяризация
Открыты калиевые каналы,
преобладает выходящий из клетки калиевый ток
Происходит восстановление поляризации
3)Реполяризация
Открыты калиевые каналы,
преобладает выходящий из клетки калиевый ток
Происходит восстановление поляризации

4)Следовая деполяризация
Отрицательный следовой потенциал
Связана с резким снижением калиевого тока и остаточным
4)Следовая деполяризация
Отрицательный следовой потенциал
Связана с резким снижением калиевого тока и остаточным

5)Следовая гиперполяризация
Положительный следовой потенциал
Связан с преобладанием выходящего калиевого тока
5)Следовая гиперполяризация
Положительный следовой потенциал
Связан с преобладанием выходящего калиевого тока
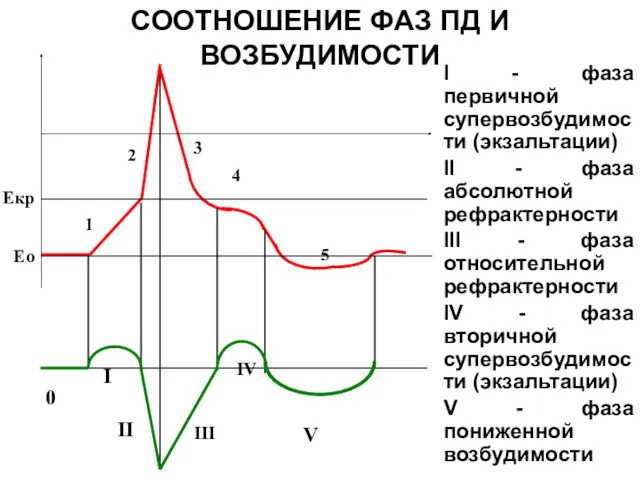
СООТНОШЕНИЕ ФАЗ ПД И ВОЗБУДИМОСТИ
I - фаза первичной супервозбудимости (экзальтации)
II -
СООТНОШЕНИЕ ФАЗ ПД И ВОЗБУДИМОСТИ
I - фаза первичной супервозбудимости (экзальтации)
II -
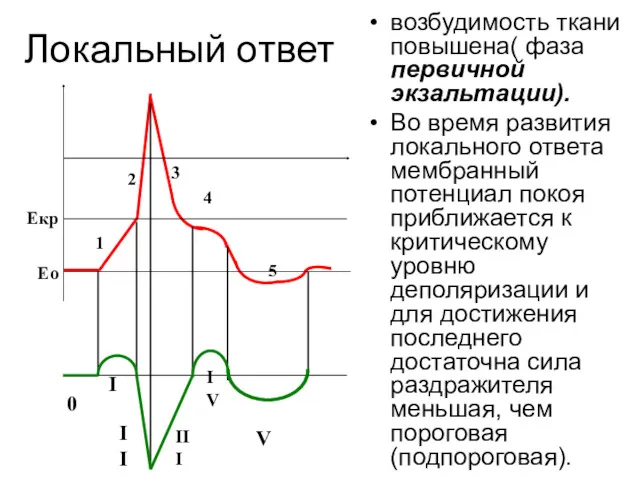
Локальный ответ
возбудимость ткани повышена( фаза первичной экзальтации).
Во время развития локального
Локальный ответ
возбудимость ткани повышена( фаза первичной экзальтации).
Во время развития локального
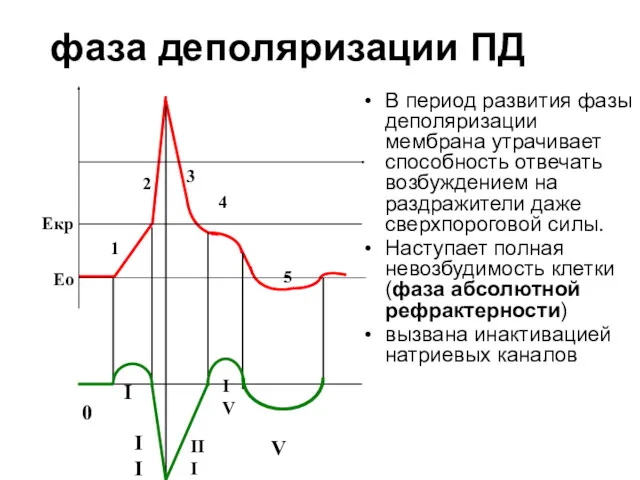
фаза деполяризации ПД
В период развития фазы деполяризации мембрана утрачивает способность отвечать
фаза деполяризации ПД
В период развития фазы деполяризации мембрана утрачивает способность отвечать
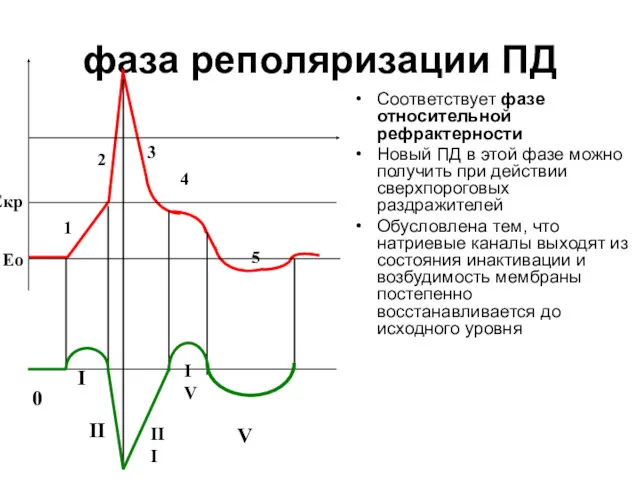
фаза реполяризации ПД
Соответствует фазе относительной рефрактерности
Новый ПД в этой фазе можно
фаза реполяризации ПД
Соответствует фазе относительной рефрактерности
Новый ПД в этой фазе можно
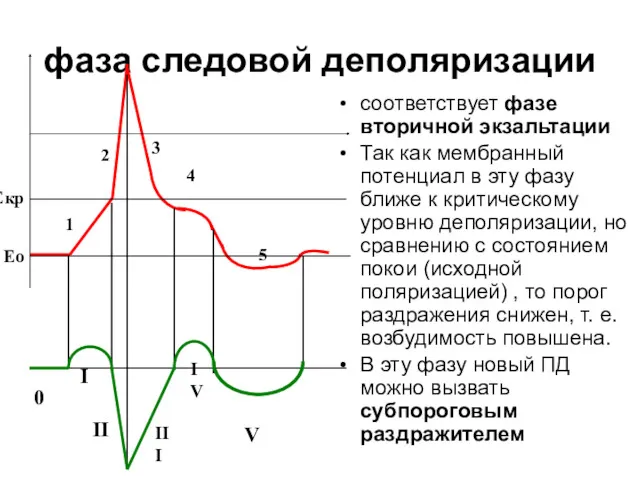
фаза следовой деполяризации
соответствует фазе вторичной экзальтации
Так как мембранный потенциал в
фаза следовой деполяризации
соответствует фазе вторичной экзальтации
Так как мембранный потенциал в
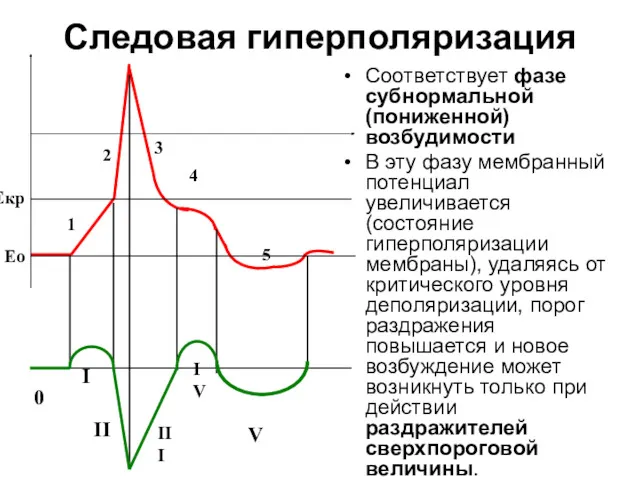
Следовая гиперполяризация
Соответствует фазе субнормальной (пониженной) возбудимости
В эту фазу мембранный потенциал
Следовая гиперполяризация
Соответствует фазе субнормальной (пониженной) возбудимости
В эту фазу мембранный потенциал

Физиология мышц
Физиология мышц
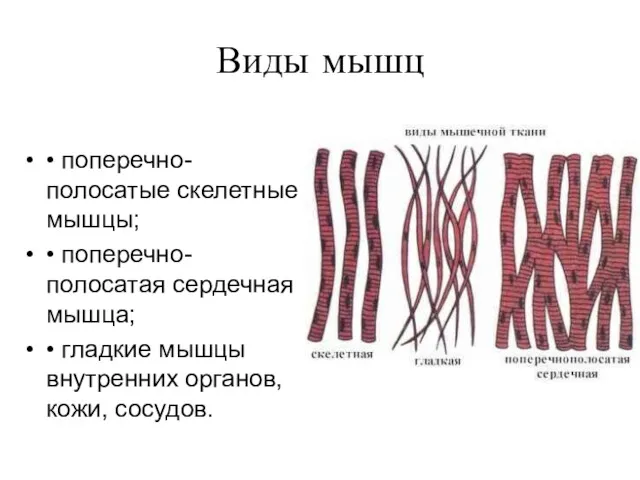
Виды мышц
• поперечно-полосатые скелетные мышцы;
• поперечно-полосатая сердечная мышца;
• гладкие мышцы внутренних
Виды мышц
• поперечно-полосатые скелетные мышцы;
• поперечно-полосатая сердечная мышца;
• гладкие мышцы внутренних
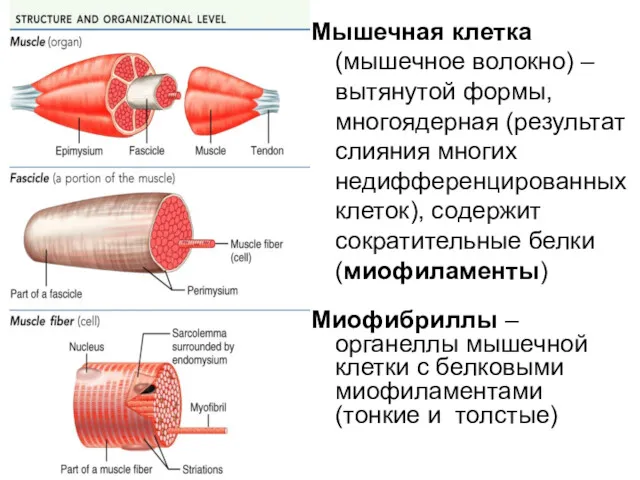
Мышечная клетка (мышечное волокно) – вытянутой формы, многоядерная (результат слияния многих
Мышечная клетка (мышечное волокно) – вытянутой формы, многоядерная (результат слияния многих

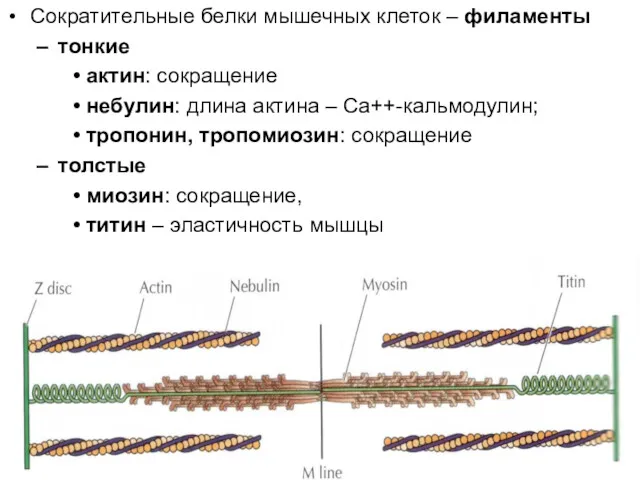
Сократительные белки мышечных клеток – филаменты
тонкие
актин: сокращение
небулин: длина актина –
Сократительные белки мышечных клеток – филаменты
тонкие
актин: сокращение
небулин: длина актина –
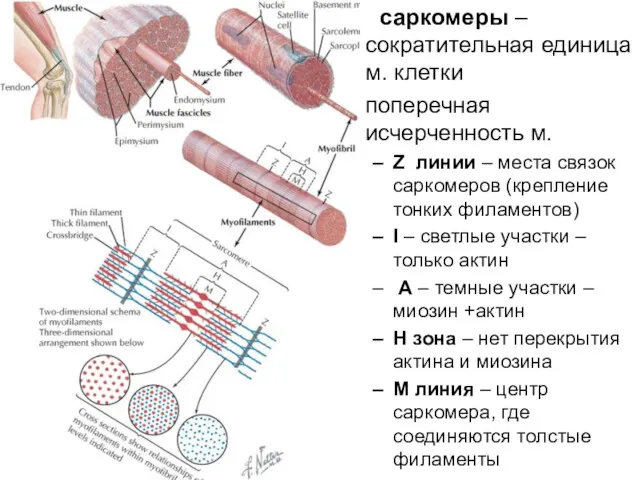
саркомеры – сократительная единица м. клетки
поперечная исчерченность м.
Z линии –
саркомеры – сократительная единица м. клетки
поперечная исчерченность м.
Z линии –


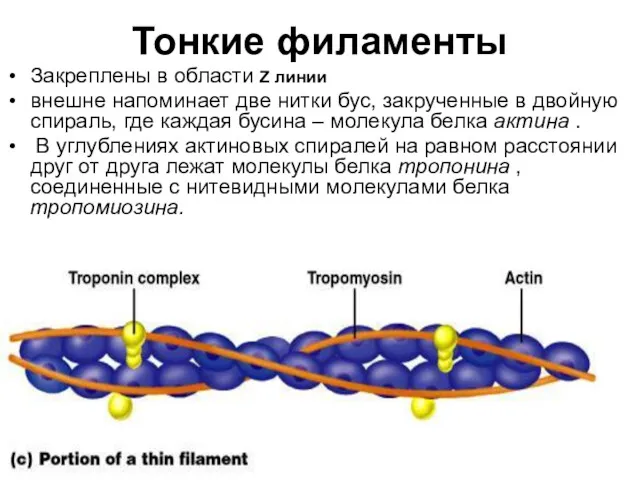
Тонкие филаменты
Закреплены в области Z линии
внешне напоминает две нитки бус,
Тонкие филаменты
Закреплены в области Z линии
внешне напоминает две нитки бус,
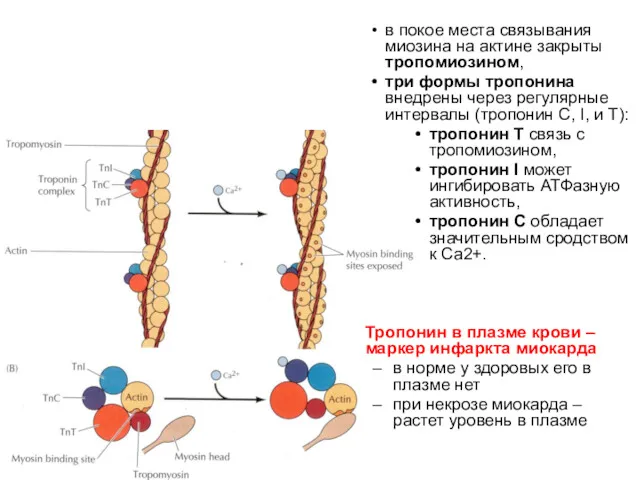
в покое места связывания миозина на актине закрыты тропомиозином,
три формы
три формы
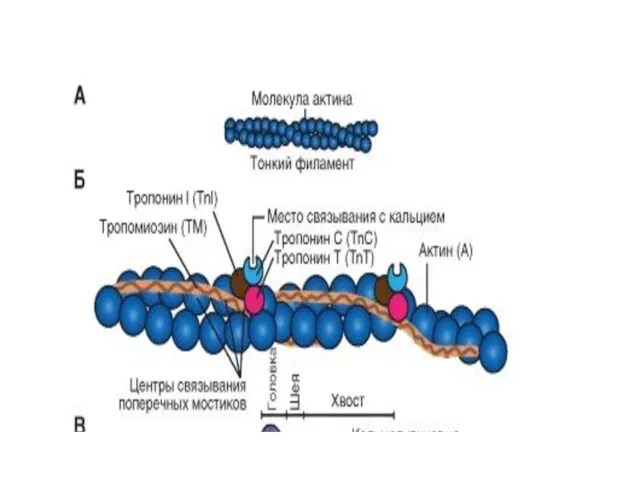
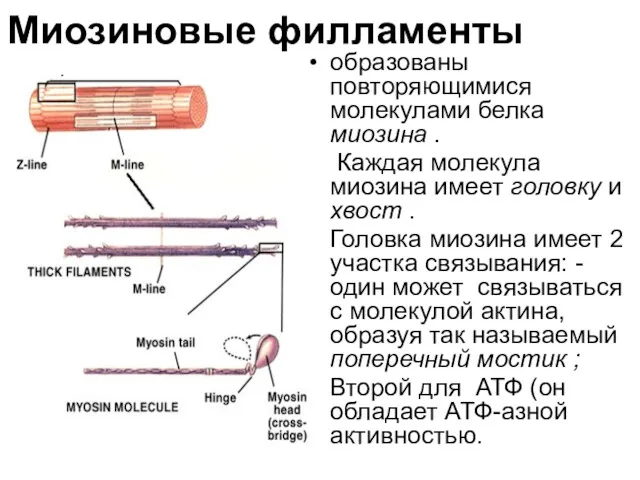
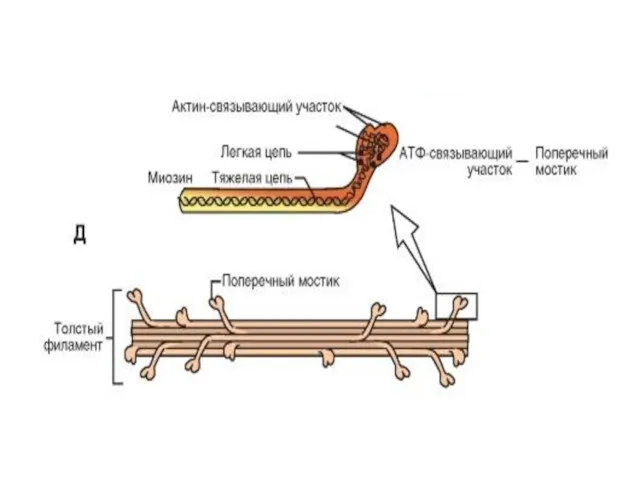
Миозиновые филламенты
образованы повторяющимися молекулами белка миозина .
Каждая молекула миозина
Миозиновые филламенты
образованы повторяющимися молекулами белка миозина .
Каждая молекула миозина

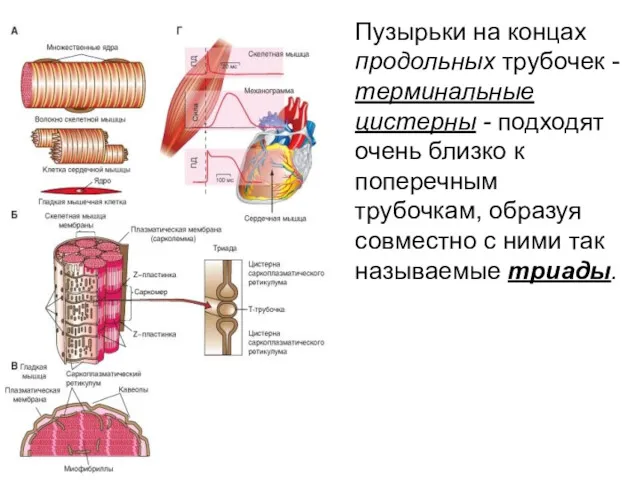
Пузырьки на концах продольных трубочек - терминальные цистерны - подходят очень
Пузырьки на концах продольных трубочек - терминальные цистерны - подходят очень

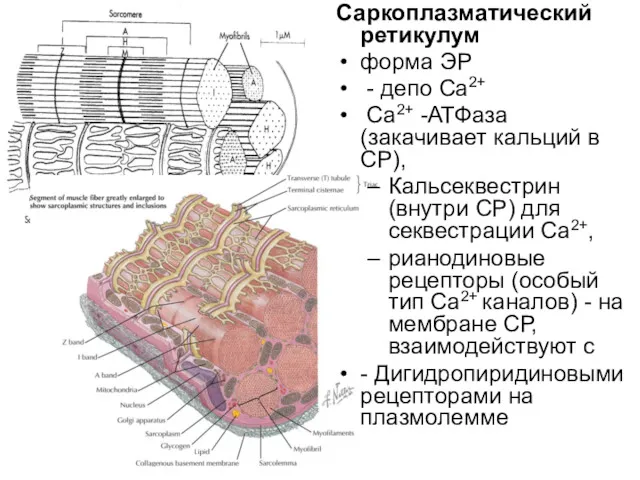
Саркоплазматический ретикулум
форма ЭР
- депо Ca2+
Ca2+ -АТФаза
Саркоплазматический ретикулум
форма ЭР
- депо Ca2+
Ca2+ -АТФаза
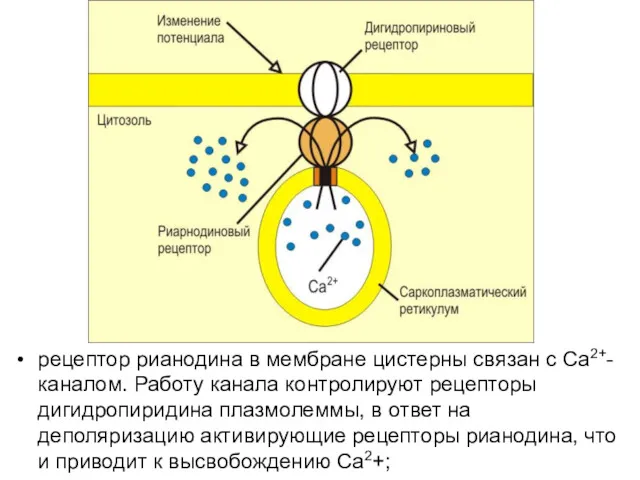
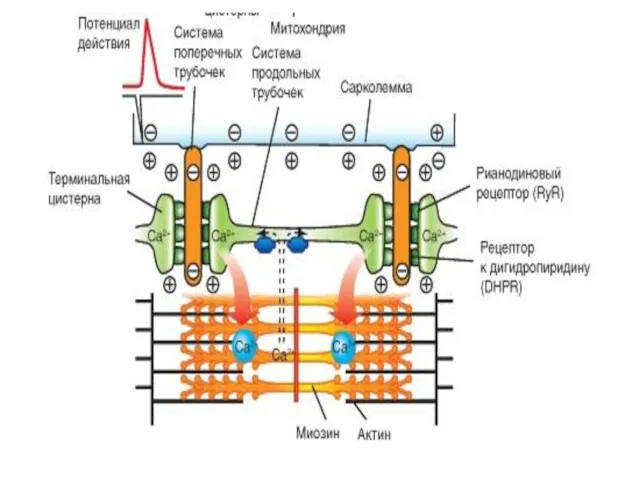
рецептор рианодина в мембране цистерны связан с Ca2+-каналом. Работу канала контролируют
рецептор рианодина в мембране цистерны связан с Ca2+-каналом. Работу канала контролируют
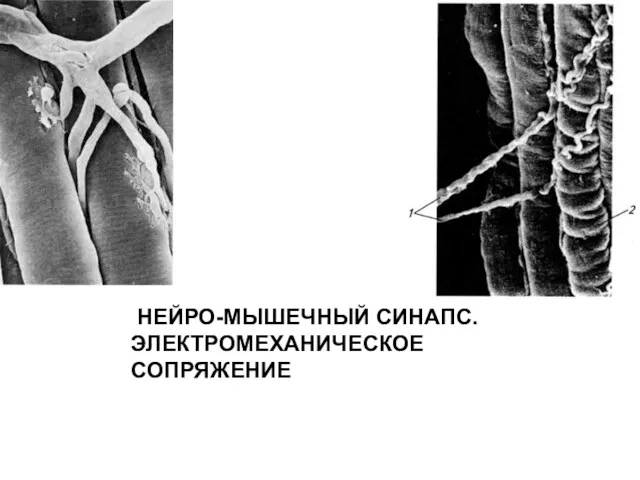
НЕЙРО-МЫШЕЧНЫЙ СИНАПС. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ
НЕЙРО-МЫШЕЧНЫЙ СИНАПС. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ
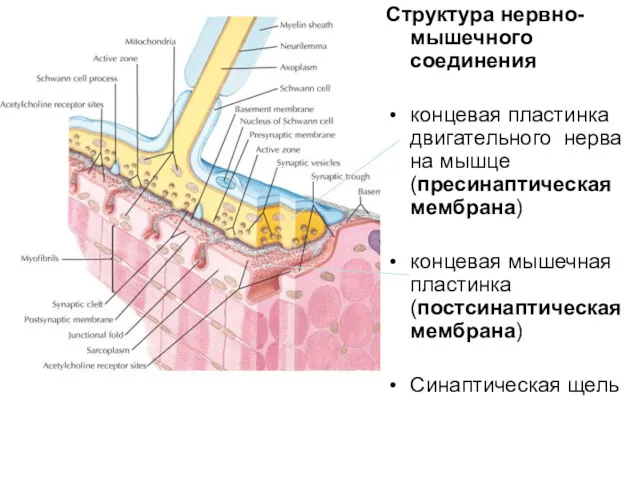
Структура нервно-мышечного соединения
концевая пластинка двигательного нерва на мышце (пресинаптическая мембрана)
концевая мышечная
Структура нервно-мышечного соединения
концевая пластинка двигательного нерва на мышце (пресинаптическая мембрана)
концевая мышечная

Внутри терминального участка аксона находятся везикул- пузырьки, содержащие медиатор- вещество, при
Внутри терминального участка аксона находятся везикул- пузырьки, содержащие медиатор- вещество, при
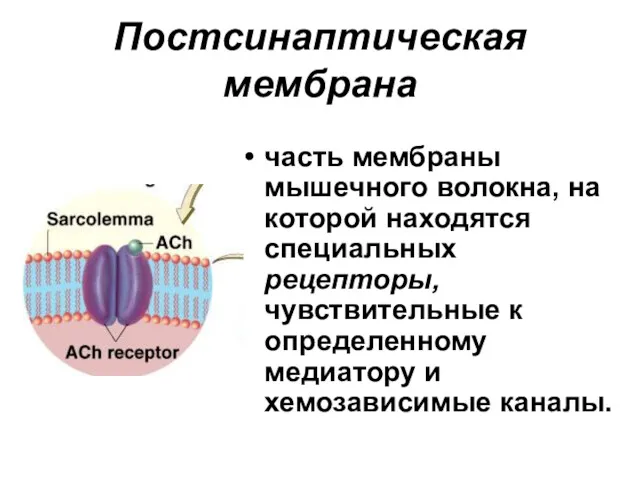
Постсинаптическая мембрана
часть мембраны мышечного волокна, на которой находятся специальных рецепторы,
Постсинаптическая мембрана
часть мембраны мышечного волокна, на которой находятся специальных рецепторы,


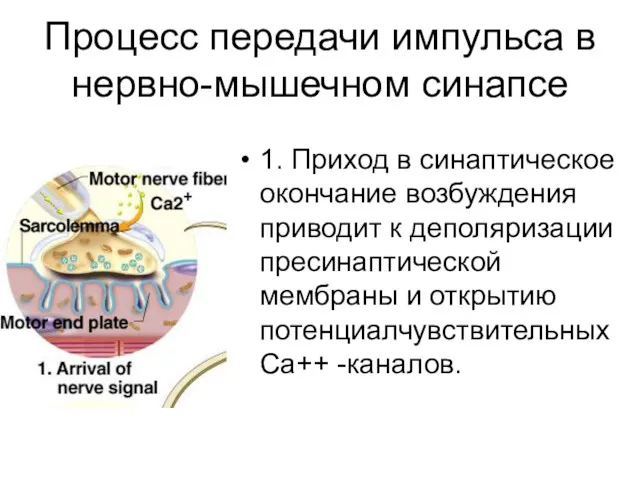
Процесс передачи импульса в нервно-мышечном синапсе
1. Приход в синаптическое окончание возбуждения
Процесс передачи импульса в нервно-мышечном синапсе
1. Приход в синаптическое окончание возбуждения
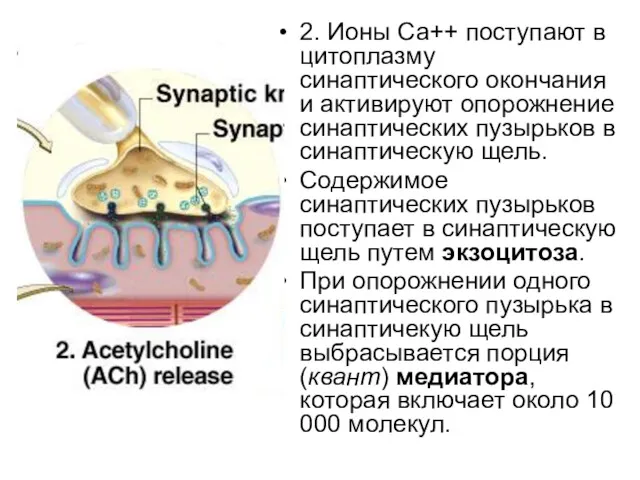
2. Ионы Са++ поступают в цитоплазму синаптического окончания и активируют опорожнение
2. Ионы Са++ поступают в цитоплазму синаптического окончания и активируют опорожнение
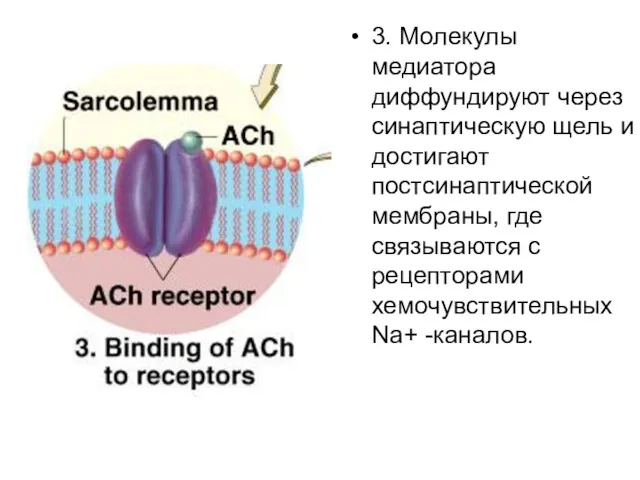
3. Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны,
3. Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны,
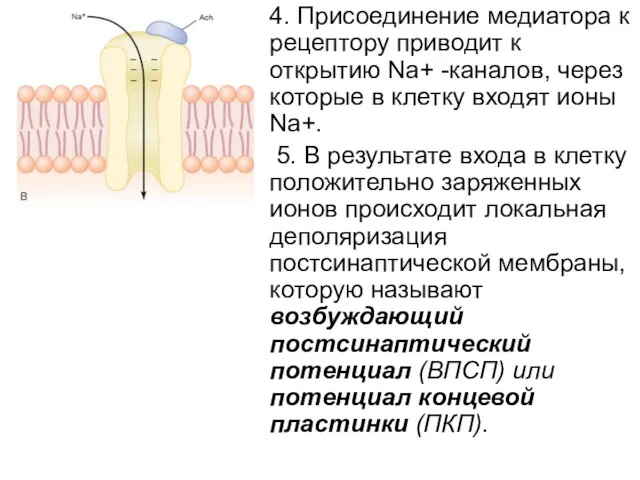
4. Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через
4. Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через
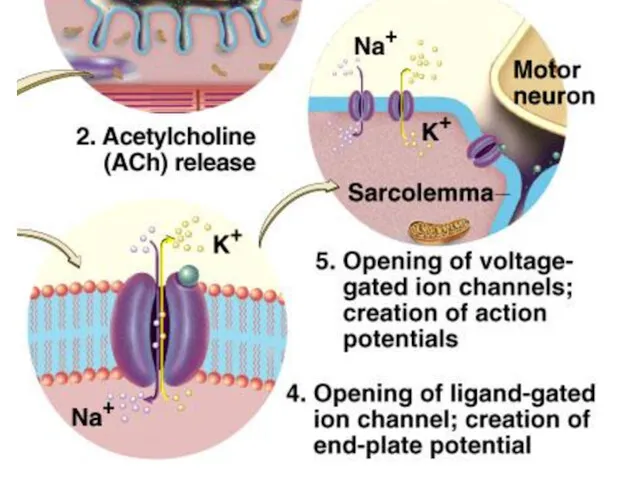
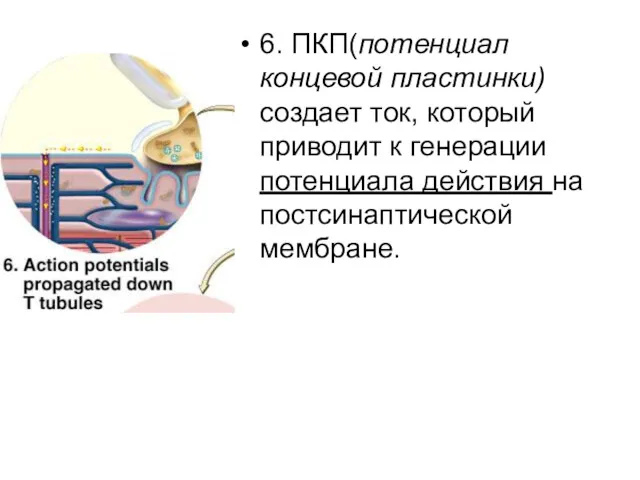
6. ПКП(потенциал концевой пластинки) создает ток, который приводит к генерации потенциала
6. ПКП(потенциал концевой пластинки) создает ток, который приводит к генерации потенциала

Медиатором в нервно-мышечном синапсе является ацетилхолин.
После взаимодействия АХ с рецептором специальный
Медиатором в нервно-мышечном синапсе является ацетилхолин.
После взаимодействия АХ с рецептором специальный

Два механизма высвобождения медиатора в синапсе:
1.с полным слиянием везикулы с плазмалеммой
Два механизма высвобождения медиатора в синапсе:
1.с полным слиянием везикулы с плазмалеммой
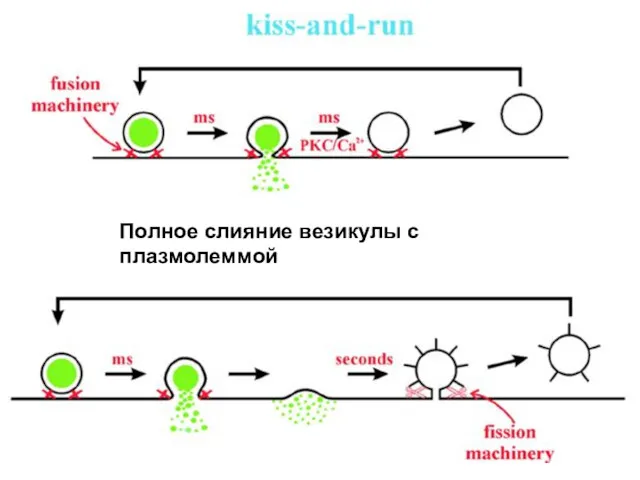
Полное слияние везикулы с плазмолеммой
Полное слияние везикулы с плазмолеммой
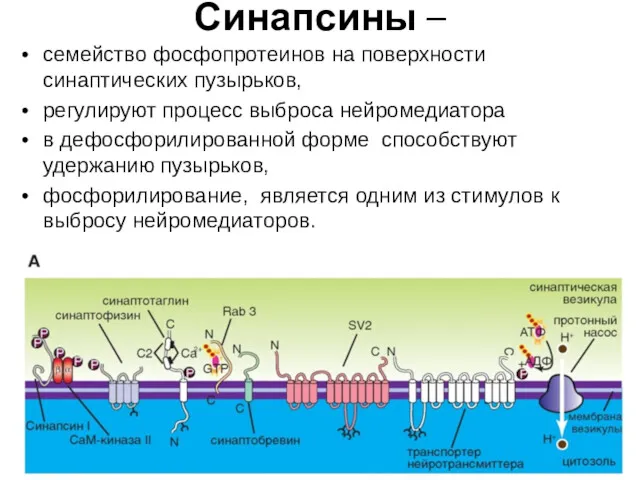
Синапсины –
семейство фосфопротеинов на поверхности синаптических пузырьков,
регулируют процесс выброса
Синапсины –
семейство фосфопротеинов на поверхности синаптических пузырьков,
регулируют процесс выброса
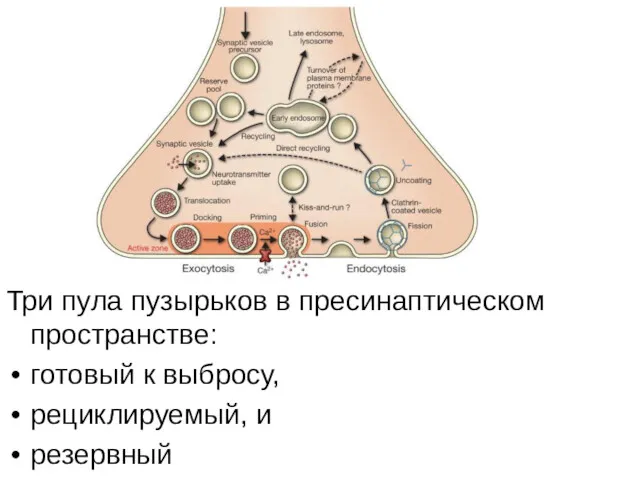
Три пула пузырьков в пресинаптическом пространстве:
готовый к выбросу,
рециклируемый, и
Три пула пузырьков в пресинаптическом пространстве:
готовый к выбросу,
рециклируемый, и
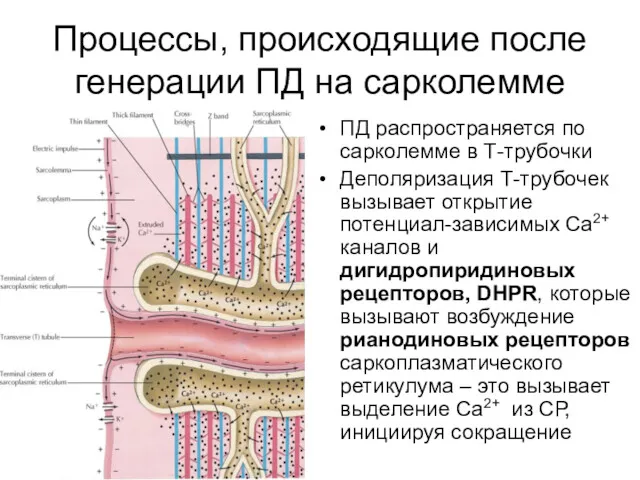
Процессы, происходящие после генерации ПД на сарколемме
ПД распространяется по сарколемме в
Процессы, происходящие после генерации ПД на сарколемме
ПД распространяется по сарколемме в

Теория скользящих нитей
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему
Теория скользящих нитей
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему
![В покое в межфибриллярном пространстве низкая [Ca2+ ] 10-7 М](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/138865/slide-55.jpg)
В покое в межфибриллярном пространстве низкая [Ca2+ ] 10-7 М - связывание
В покое в межфибриллярном пространстве низкая [Ca2+ ] 10-7 М - связывание

После выделения Ca2+ из СР (вследствие активации рианодиновых рецепторов) кальций связывается
После выделения Ca2+ из СР (вследствие активации рианодиновых рецепторов) кальций связывается
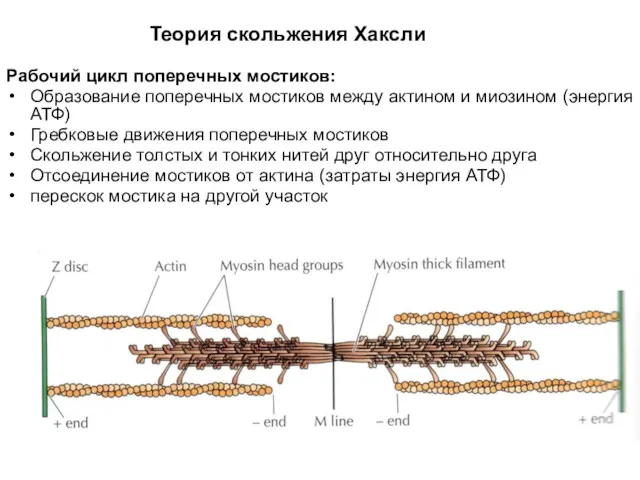
Теория скольжения Хаксли
Рабочий цикл поперечных мостиков:
Образование поперечных мостиков между актином и
Теория скольжения Хаксли
Рабочий цикл поперечных мостиков:
Образование поперечных мостиков между актином и



В цикле поперечных мостиков АТФ выполняет две разные роли:
1) гидролиз АТФ поставляет энергию
В цикле поперечных мостиков АТФ выполняет две разные роли:
1) гидролиз АТФ поставляет энергию
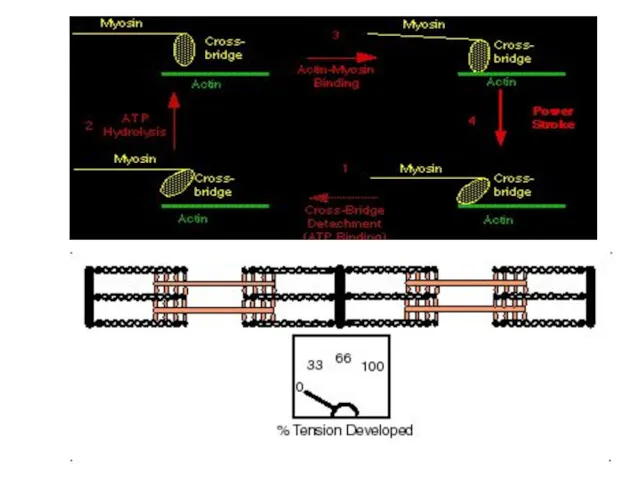

Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения

В зависимости от частоты приходящих к мышце нервных импульсов мышца может
В зависимости от частоты приходящих к мышце нервных импульсов мышца может
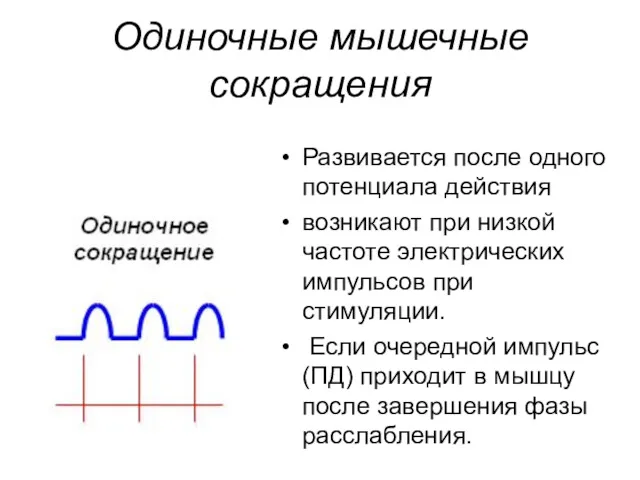
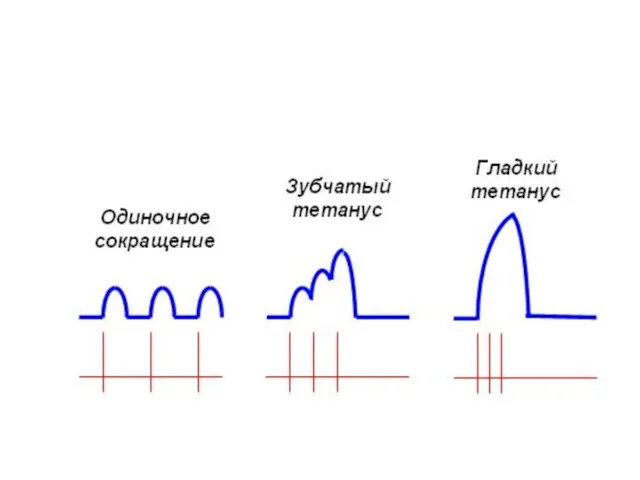
Одиночные мышечные сокращения
Развивается после одного потенциала действия
возникают при низкой частоте электрических
Одиночные мышечные сокращения
Развивается после одного потенциала действия
возникают при низкой частоте электрических
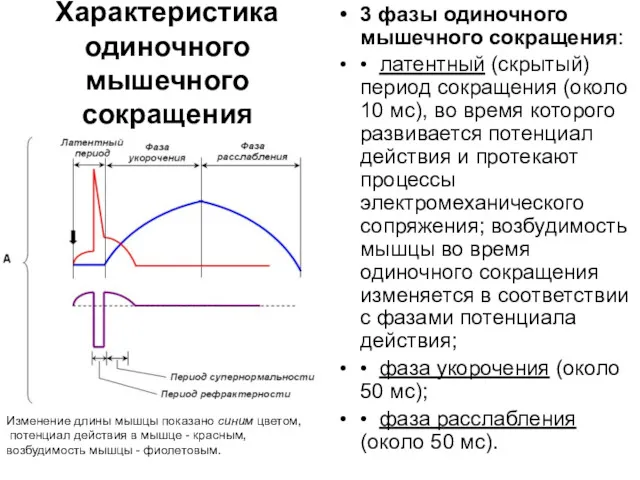
Характеристика одиночного мышечного сокращения
Изменение длины мышцы показано синим цветом,
потенциал действия
Характеристика одиночного мышечного сокращения
Изменение длины мышцы показано синим цветом,
потенциал действия

Тетанус (лат. tetanus, от греч. tétanos — напряжение, оцепенение, судорога)
При слишком частой стимуляции мышцы частотная суммация обусловливает появление
тетануса
Тетанус (лат. tetanus, от греч. tétanos — напряжение, оцепенение, судорога)
При слишком частой стимуляции мышцы частотная суммация обусловливает появление
тетануса

Во время тетанического сокращения в мышце многократно возникают ПД, обеспечивающие продолжительный
Во время тетанического сокращения в мышце многократно возникают ПД, обеспечивающие продолжительный
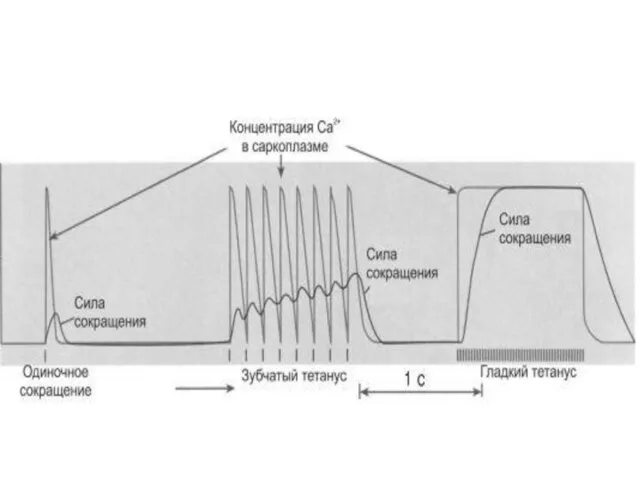
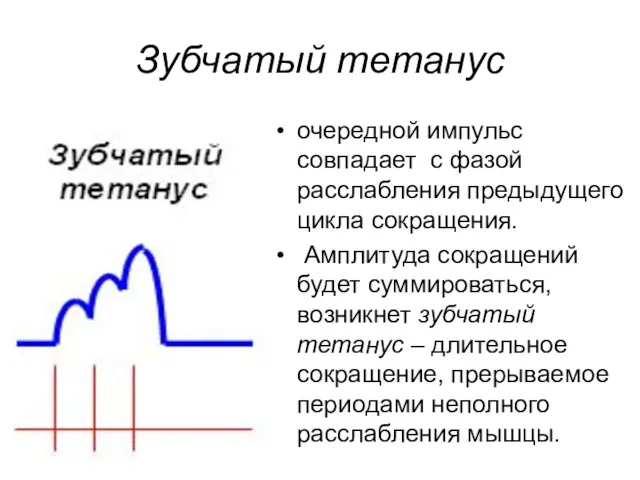
Зубчатый тетанус
очередной импульс совпадает с фазой расслабления предыдущего цикла сокращения.
Зубчатый тетанус
очередной импульс совпадает с фазой расслабления предыдущего цикла сокращения.
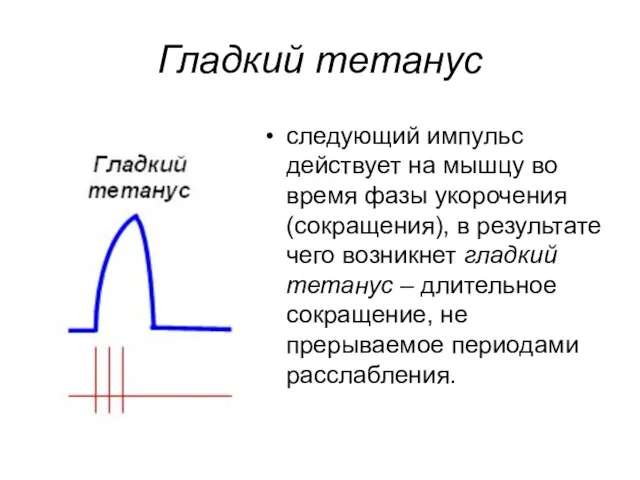
Гладкий тетанус
следующий импульс действует на мышцу во время фазы укорочения
Гладкий тетанус
следующий импульс действует на мышцу во время фазы укорочения


Оптимум и пессимум частоты
Амплитуда тетанического сокращения, основанного на суммации, зависит
Оптимум и пессимум частоты
Амплитуда тетанического сокращения, основанного на суммации, зависит
 Основные процессы жизнедеятельности растений
Основные процессы жизнедеятельности растений Земноводные, или амфибии
Земноводные, или амфибии Разнообразие пасленовых (акцент на цветки)
Разнообразие пасленовых (акцент на цветки) Методы дрессировки немецких овчарок по общерозыскному профилю и по поиску взрывчатых веществ
Методы дрессировки немецких овчарок по общерозыскному профилю и по поиску взрывчатых веществ Выделительная система
Выделительная система История семейства розоцветных
История семейства розоцветных Эволюция пищеварительной системы
Эволюция пищеварительной системы Травы и цветы полей и лугов
Травы и цветы полей и лугов Состав крови
Состав крови Многообразие птиц в природе
Многообразие птиц в природе многообразие мхов
многообразие мхов Жизнь в аквариуме
Жизнь в аквариуме Тест. Вариант № 3. Основы общей и медицинской генетики
Тест. Вариант № 3. Основы общей и медицинской генетики Кровеносная система человека
Кровеносная система человека :Нервная система рыб
:Нервная система рыб Extinction of animals
Extinction of animals Самые - самые растения
Самые - самые растения Качкодзьоб. Спеціальні пристосування качкодзьоба
Качкодзьоб. Спеціальні пристосування качкодзьоба Теория И.П. Павлова о типах высшей нервной деятельности
Теория И.П. Павлова о типах высшей нервной деятельности Что такое адаптации? Какие бывают адаптации?
Что такое адаптации? Какие бывают адаптации? Микроскоп и его составляющие
Микроскоп и его составляющие Семейство пасленовых. Презентация по биологии для 6 класса
Семейство пасленовых. Презентация по биологии для 6 класса Ткани человека. Виды и строение
Ткани человека. Виды и строение Волк. Охота волков
Волк. Охота волков Навіщо Землі атмосфера. Урок №35. 5 клас
Навіщо Землі атмосфера. Урок №35. 5 клас Наследственность и изменчивость (модификационная, комбинативная). Модуль 1
Наследственность и изменчивость (модификационная, комбинативная). Модуль 1 Паук каракурт
Паук каракурт Интерактивная игра Кто? Как? Почему? Играя, вспоминаем для 8 класса (по материалу I полугодия)
Интерактивная игра Кто? Как? Почему? Играя, вспоминаем для 8 класса (по материалу I полугодия)