- Физиология растений. Механизмы поступления минеральных элементов в клетку

Содержание
- 2. Вещества в виде ионов преодолевают мембраны растительной клетки несколькими способами, основные из которых: Диффузия через липидную
- 4. Две движущие силы мембранного транспорта. Пассивный транспорт – перемещение веществ путем диффузии по градиенту электрохимического потенциала
- 5. Расшифрованные геномы высших растений содержат примерно от 100 до 200 генов различных ионных каналов. Практически все
- 6. Геномы водорослей имеют отличные от высших растений семейства гены ионных каналов, более близкие эволюционно и структурно
- 7. Paddle – «весло», «затвор», «воротный механизм» Если мембранный потенциал растет, то «затвор» открывает канал Roderick MacKinnon
- 8. Канал имеет водную пору, которая образуется специальным участком каждой из 4 субъединиц канала. Одна субъединица состоит
- 9. Схема строения катионного канала мембран растений, проницаемого для ионов К+
- 10. Семейства генов катионных каналов Arabidopsis thaliana L. Heynh. и структурная организация их продуктов
- 11. Анионные каналы Поток анионов через плазматическую мембрану растительной клетки реализуется посредством работы анион-селективных ионных каналов (анионных
- 12. Анионные каналы Анализ генома высших растений и электрофизиологические исследования выявили три семейства генов, которые кодируют многообразие
- 13. Транспорт через плазматическую мембрану эпидермальных клеток корня является началом пути ионов до места их назначения в
- 14. Эпидермальные клетки представлены (примерно 1:1) трихобластами (образующими корневые волоски) и атрихобластами (необразующими корневые волоски) 100 μм
- 15. Все эпидермальные клетки корня участвуют в поглощении минеральных элементов из почвы, но наиболее активны зона роста
- 16. Эпидермис зоны роста растяжением корня Arabidopsis thaliana 300 μм Балджи – зачаточные корневые волоски Плотность ионных
- 17. Транспорт веществ из почвенного раствора в цитоплазму через плазматическую мембрану эпидермальных клеток корня: 1) Mg2+, Cа2+
- 18. Гены ионных каналов растений:
- 19. Примеры структуры субъединиц катионных каналов:
- 20. Примеры молекулярно-биологических свойств катионных каналов: - структура и расположение генов, субъединиц, функциональных доменов - влияние факторов
- 21. Изучение ионных каналов: - на уровне популяции каналов целой клетки - на уровне одиночных каналов -
- 22. Пэтч-кламп:
- 23. Регистрация одиночных каналов:
- 24. Вольт-Амперная Характеристика (ВАХ) проводимости или ионного канала
- 25. Кривые временного хода (кинетика) изменения токов через мембрану в ответ на изменение напряжение
- 26. Са2+-люминометрия
- 27. MIFE – microelectrode ion flux estimation внешние сканирующие ион-селективные электроды вибрирующие ион-селективные электроды
- 28. Различают следующие типы активного транспорта минеральных веществ: 1) первичный активный транспорт – трансмембранный векторный перенос иона
- 29. 2) вторичный активный транспорт происходит, когда в качестве энергетического источника используются градиенты других ионов, например, электрохимический
- 30. АТФазы – обширная группа ферментов, имеющих схожую структурную организацию (различают пять классов: F-, V-, A-, E-,
- 31. Наиболее важные АТФазы: F-АТФазы (F1-F0) – АТФ-синтазы – митох., хлоропл. V-АТФазы (V1-V0) – эндомембранные (Н+) Р-АТФазы
- 32. Р-АТФазы (всего 159 у всех организмов). 5 типов Тип I - переносят калий, переходные металлы, тяжелые
- 33. Пример структуры - V-типа АТФаз растений: V1 - цитоплазматический гидрофильный домен, состоит из 8 субъединиц (A-H),
- 34. Front. Plant Sci., 02 December 2011 | https://doi.org/10.3389/fpls.2011.00085 Calcium efflux systems in stress signaling and adaptation
- 35. Более подробно об вакуолярных АТФазах http://jeb.biologists.org/content/209/4/577.full.pdf
- 36. Ион-специфичные активные транспортеры участвуют в транспорте практически всех макро- и микроэлементов минерального питания и представляют собой
- 37. Движущей силой ион-специфичных транспортеров является разность электрохимических потенциалов ионов Н+, Na+ или K+. Всего выделено на
- 38. Азотное питание N – критически-важный компонент аминокислот и белков (18% от массы белка), а также ДНК,
- 39. Азот-фиксирующие бактерии N2 Аммонифи- цирующие бактерии NH3 (аммоний) Органический материал (гумус) атмосфера почва H+ (из почвы)
- 40. Фиксация азота Индустриальная фиксация N2 N2 – самый обильный газ атмосферы (70%), но он не доступен
- 41. клубеньки (нодулы) Корни гороха с клубеньками. корни Корни бобовых разбухают, образуя так-называемые клубеньки или «нодулы», состоящие
- 42. бактероиды в клубеньках сои 5 µм Бактероиды в пределах везикулы Внутри корневых клубеньков Rhizobium живут в
- 43. бактериоиды в клубеньках сои 5 µм Бактероиды в пределах везикулы Бактерии получают из корня сахара и
- 44. Бактерии Rhizobium Инфицирование Химический сигнал привлекает бактерии 1 1 2 2 3 3 4 4 Инфицированный
- 45. Нитрат, попадая в корневые клетки, восстанавливается до аммония, который затем включается в аминокислоты. Аммоний, «синтезированный» из
- 46. Цепочка превращений нитрата к аммиаку начинается на внешней поверхности корня и разворачивается внутри корневых тканей. Другая
- 47. Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937 г. биохимиками А. Е. Браунштейном
- 48. Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937 г. биохимиками А. Е. Браунштейном
- 49. Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937 г. биохимиками А. Е. Браунштейном
- 50. Показано также, что под влиянием соответствующих аминотрансфераз аспарагин и глютамин также могут передавать свои аминные группы
- 51. Цикл фосфора
- 53. Скачать презентацию

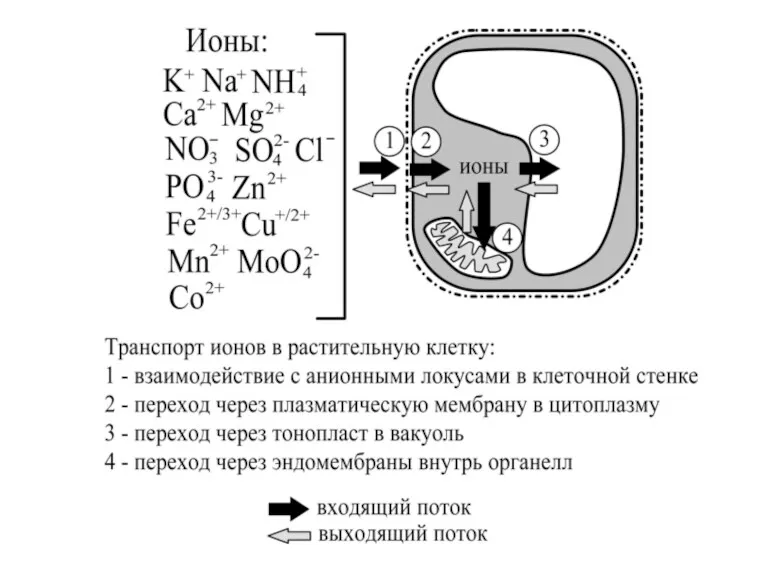
Вещества в виде ионов преодолевают мембраны растительной клетки несколькими способами, основные
Вещества в виде ионов преодолевают мембраны растительной клетки несколькими способами, основные

Две движущие силы мембранного транспорта.
Пассивный транспорт – перемещение веществ путем
Две движущие силы мембранного транспорта.
Пассивный транспорт – перемещение веществ путем

Расшифрованные геномы высших растений содержат примерно от 100 до 200 генов
Расшифрованные геномы высших растений содержат примерно от 100 до 200 генов

Геномы водорослей имеют отличные от высших растений семейства гены ионных каналов,
Геномы водорослей имеют отличные от высших растений семейства гены ионных каналов,
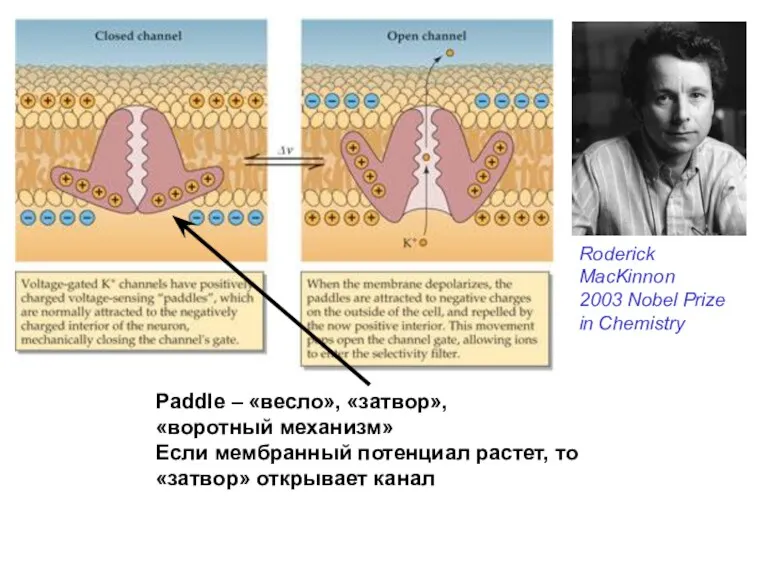
Paddle – «весло», «затвор»,
«воротный механизм»
Если мембранный потенциал растет, то
«затвор» открывает
Paddle – «весло», «затвор»,
«воротный механизм»
Если мембранный потенциал растет, то
«затвор» открывает

Канал имеет водную пору, которая образуется специальным участком каждой из 4
Канал имеет водную пору, которая образуется специальным участком каждой из 4
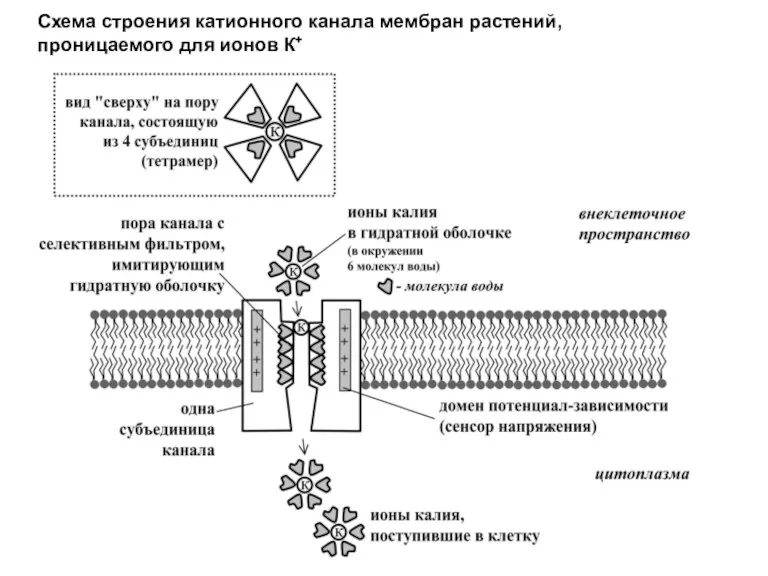
Схема строения катионного канала мембран растений,
проницаемого для ионов К+
Схема строения катионного канала мембран растений,
проницаемого для ионов К+
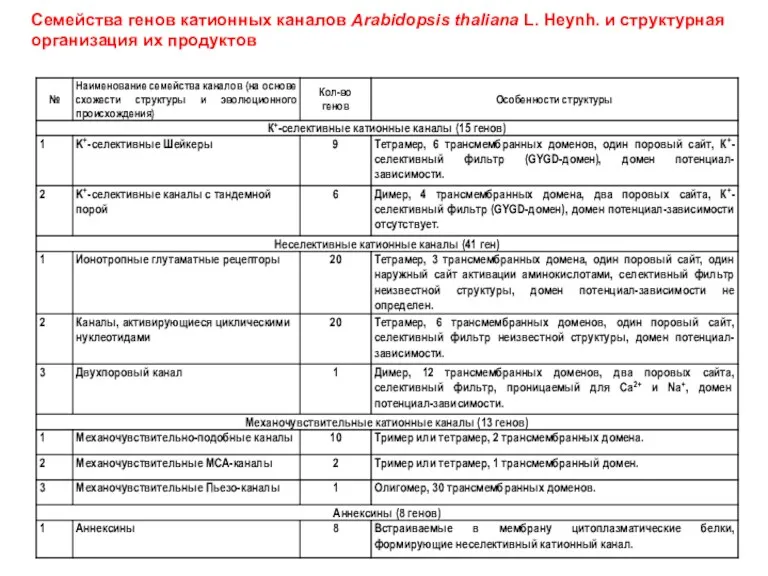
Семейства генов катионных каналов Arabidopsis thaliana L. Heynh. и структурная организация
Семейства генов катионных каналов Arabidopsis thaliana L. Heynh. и структурная организация

Анионные каналы
Поток анионов через плазматическую мембрану растительной клетки реализуется посредством работы
Анионные каналы
Поток анионов через плазматическую мембрану растительной клетки реализуется посредством работы

Анионные каналы
Анализ генома высших растений и электрофизиологические исследования выявили три семейства
Анионные каналы
Анализ генома высших растений и электрофизиологические исследования выявили три семейства

Транспорт через плазматическую мембрану эпидермальных клеток корня является началом пути ионов
Транспорт через плазматическую мембрану эпидермальных клеток корня является началом пути ионов

Эпидермальные клетки представлены (примерно 1:1)
трихобластами (образующими корневые волоски) и атрихобластами (необразующими
Эпидермальные клетки представлены (примерно 1:1)
трихобластами (образующими корневые волоски) и атрихобластами (необразующими

Все эпидермальные клетки корня участвуют в поглощении минеральных элементов из почвы,
Все эпидермальные клетки корня участвуют в поглощении минеральных элементов из почвы,

Эпидермис зоны роста растяжением
корня Arabidopsis thaliana
300 μм
Балджи – зачаточные корневые волоски
Плотность
Эпидермис зоны роста растяжением
корня Arabidopsis thaliana
300 μм
Балджи – зачаточные корневые волоски
Плотность

Транспорт веществ из почвенного раствора в цитоплазму через плазматическую мембрану эпидермальных
Транспорт веществ из почвенного раствора в цитоплазму через плазматическую мембрану эпидермальных
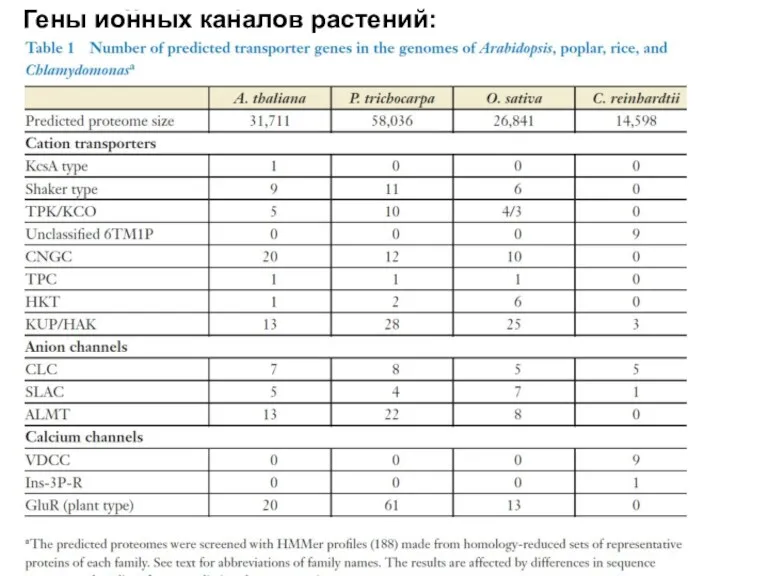
Гены ионных каналов растений:
Гены ионных каналов растений:
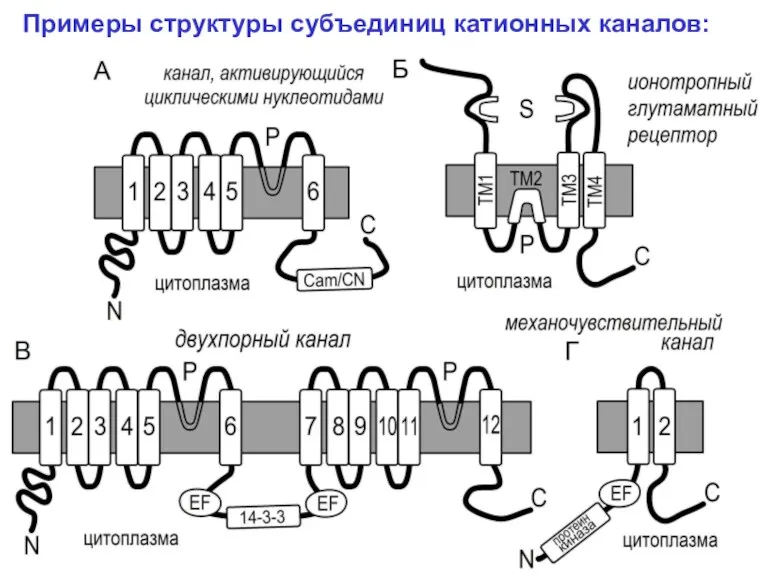
Примеры структуры субъединиц катионных каналов:
Примеры структуры субъединиц катионных каналов:

Примеры молекулярно-биологических свойств катионных каналов:
- структура и расположение генов, субъединиц, функциональных
Примеры молекулярно-биологических свойств катионных каналов:
- структура и расположение генов, субъединиц, функциональных

Изучение ионных каналов:
- на уровне популяции каналов целой клетки
- на
Изучение ионных каналов:
- на уровне популяции каналов целой клетки
- на
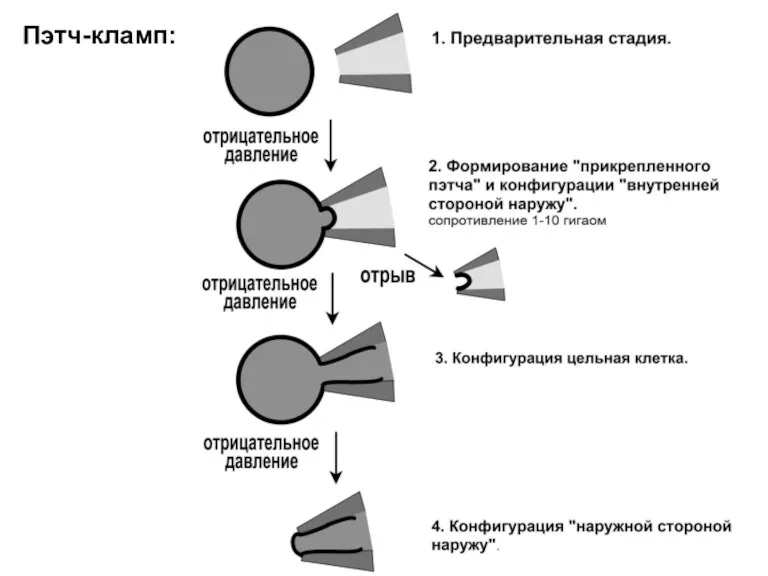
Пэтч-кламп:
Пэтч-кламп:
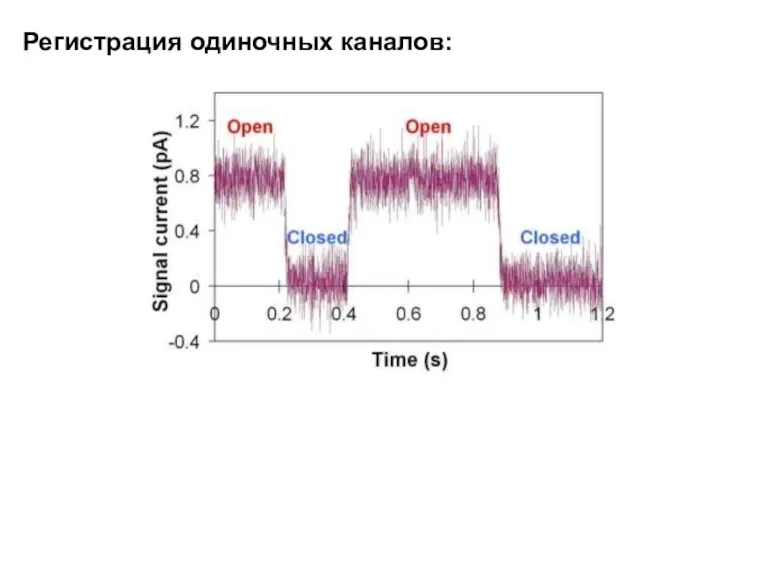
Регистрация одиночных каналов:
Регистрация одиночных каналов:
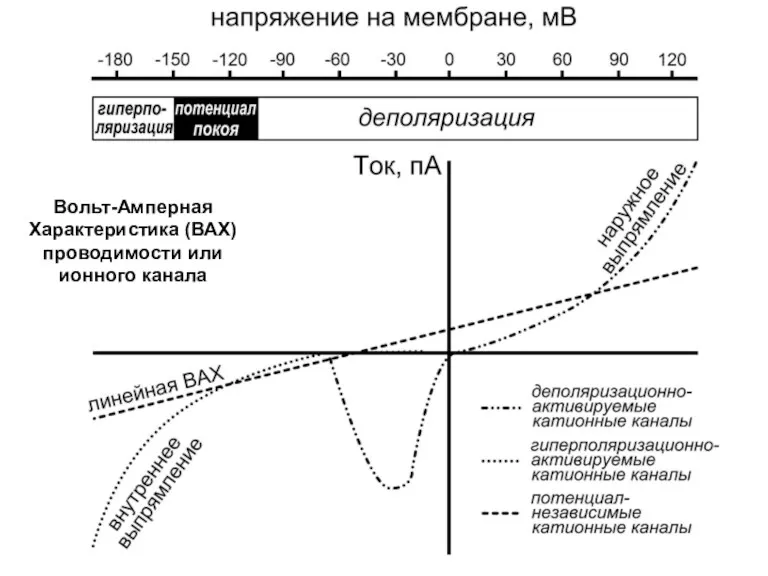
Вольт-Амперная
Характеристика (ВАХ)
проводимости или ионного канала
Вольт-Амперная
Характеристика (ВАХ)
проводимости или ионного канала
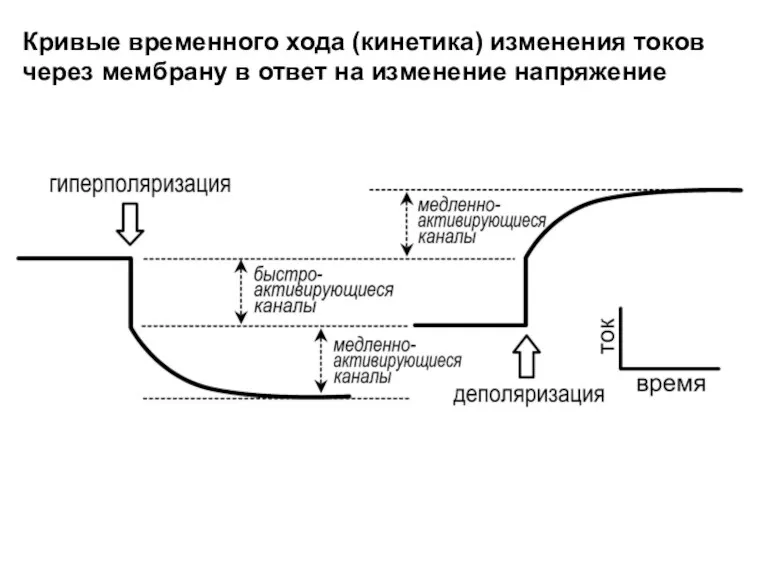
Кривые временного хода (кинетика) изменения токов через мембрану в ответ на
Кривые временного хода (кинетика) изменения токов через мембрану в ответ на
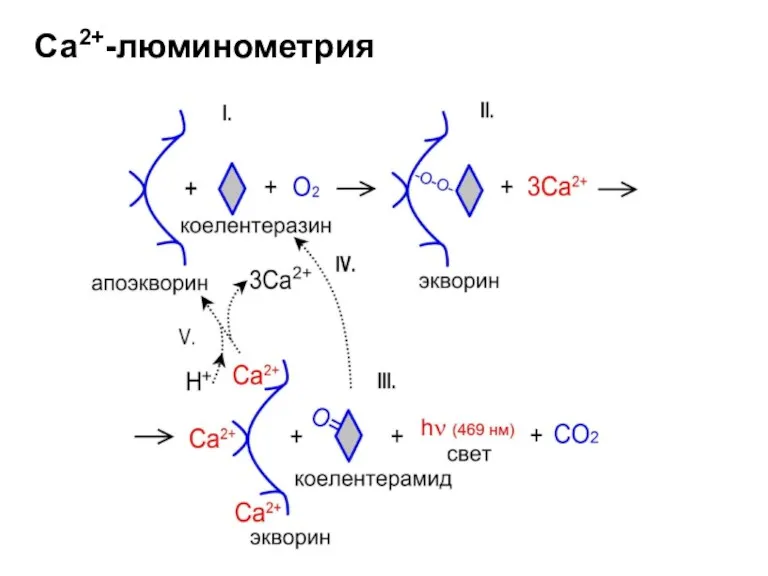
Са2+-люминометрия
Са2+-люминометрия
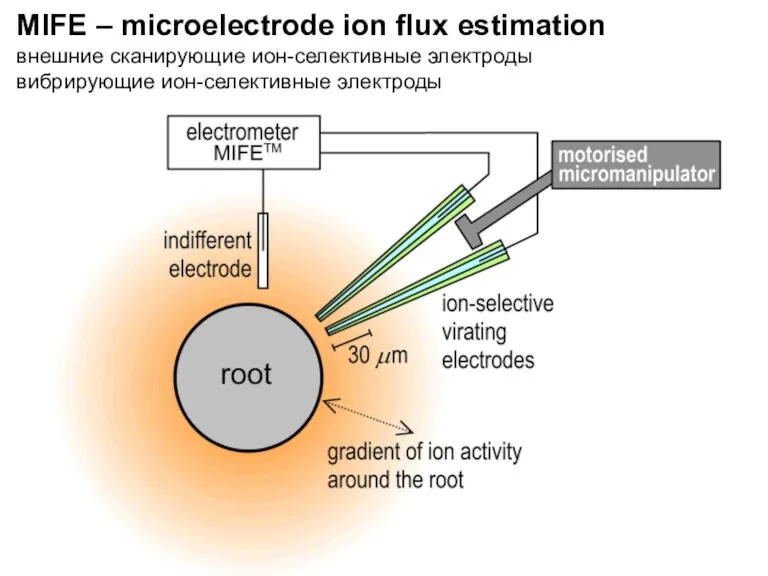
MIFE – microelectrode ion flux estimation
внешние сканирующие ион-селективные электроды
вибрирующие ион-селективные электроды
MIFE – microelectrode ion flux estimation
внешние сканирующие ион-селективные электроды
вибрирующие ион-селективные электроды

Различают следующие типы активного транспорта минеральных веществ:
1) первичный активный транспорт
Различают следующие типы активного транспорта минеральных веществ:
1) первичный активный транспорт

2) вторичный активный транспорт происходит, когда в качестве энергетического источника используются
2) вторичный активный транспорт происходит, когда в качестве энергетического источника используются

АТФазы – обширная группа ферментов, имеющих схожую структурную организацию (различают пять
АТФазы – обширная группа ферментов, имеющих схожую структурную организацию (различают пять

Наиболее важные АТФазы:
F-АТФазы (F1-F0) – АТФ-синтазы – митох., хлоропл.
V-АТФазы (V1-V0)
Наиболее важные АТФазы:
F-АТФазы (F1-F0) – АТФ-синтазы – митох., хлоропл.
V-АТФазы (V1-V0)

Р-АТФазы (всего 159 у всех организмов).
5 типов
Тип I - переносят калий, переходные
Р-АТФазы (всего 159 у всех организмов).
5 типов
Тип I - переносят калий, переходные
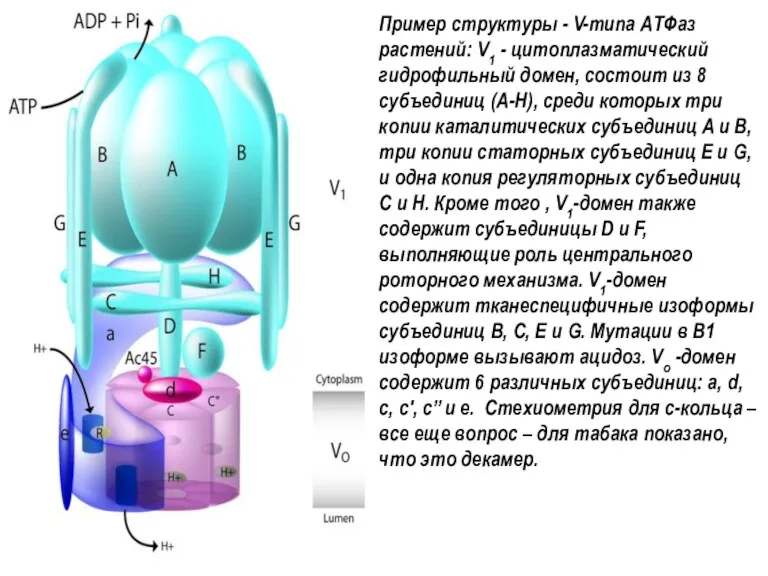
Пример структуры - V-типа АТФаз растений: V1 - цитоплазматический гидрофильный домен, состоит
Пример структуры - V-типа АТФаз растений: V1 - цитоплазматический гидрофильный домен, состоит
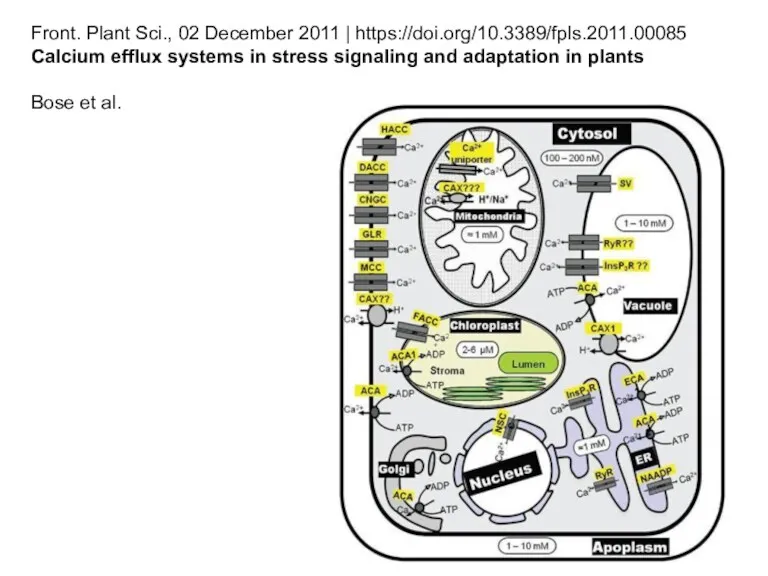
Front. Plant Sci., 02 December 2011 | https://doi.org/10.3389/fpls.2011.00085
Calcium efflux systems in
Front. Plant Sci., 02 December 2011 | https://doi.org/10.3389/fpls.2011.00085
Calcium efflux systems in

Более подробно об вакуолярных АТФазах
http://jeb.biologists.org/content/209/4/577.full.pdf
Более подробно об вакуолярных АТФазах
http://jeb.biologists.org/content/209/4/577.full.pdf
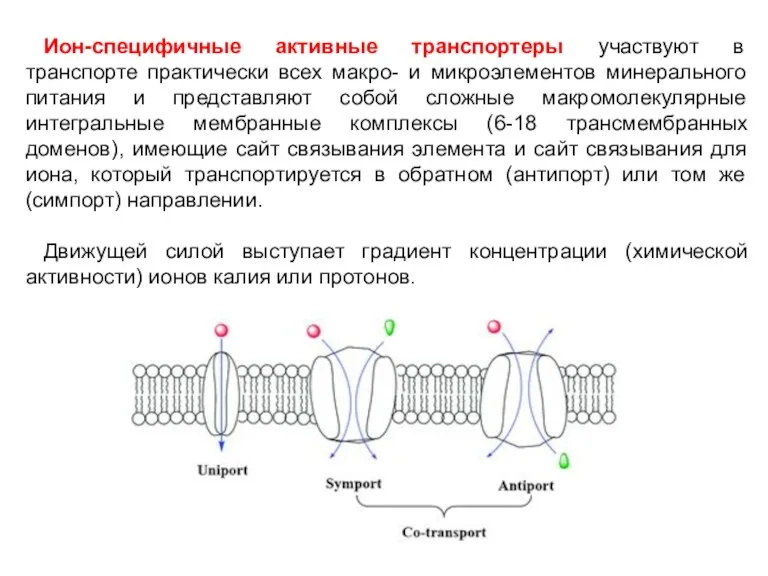
Ион-специфичные активные транспортеры участвуют в транспорте практически всех макро- и микроэлементов
Ион-специфичные активные транспортеры участвуют в транспорте практически всех макро- и микроэлементов

Движущей силой ион-специфичных транспортеров является разность электрохимических потенциалов ионов Н+, Na+
Движущей силой ион-специфичных транспортеров является разность электрохимических потенциалов ионов Н+, Na+

Азотное питание
N – критически-важный компонент аминокислот и белков (18% от массы
Азотное питание
N – критически-важный компонент аминокислот и белков (18% от массы
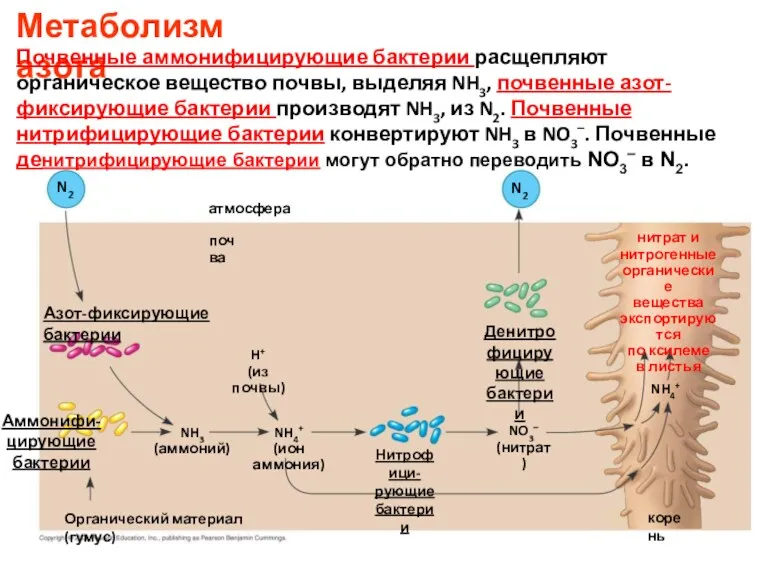
Азот-фиксирующие бактерии
N2
Аммонифи-
цирующие
бактерии
NH3
(аммоний)
Органический материал (гумус)
атмосфера
почва
H+
(из почвы)
NH4+
(ион аммония)
Нитрофици-
рующие бактерии
NO3–
(нитрат)
Денитрофицирующие
бактерии
NH4+
нитрат и
нитрогенные
органические
вещества
экспортируются
по
Азот-фиксирующие бактерии
N2
Аммонифи-
цирующие
бактерии
NH3
(аммоний)
Органический материал (гумус)
атмосфера
почва
H+
(из почвы)
NH4+
(ион аммония)
Нитрофици-
рующие бактерии
NO3–
(нитрат)
Денитрофицирующие
бактерии
NH4+
нитрат и
нитрогенные
органические
вещества
экспортируются
по

Фиксация азота
Индустриальная фиксация N2
N2 – самый обильный газ атмосферы (70%),
Фиксация азота
Индустриальная фиксация N2
N2 – самый обильный газ атмосферы (70%),

клубеньки
(нодулы)
Корни гороха с клубеньками.
корни
Корни бобовых разбухают, образуя так-называемые клубеньки или «нодулы»,
клубеньки
(нодулы)
Корни гороха с клубеньками.
корни
Корни бобовых разбухают, образуя так-называемые клубеньки или «нодулы»,
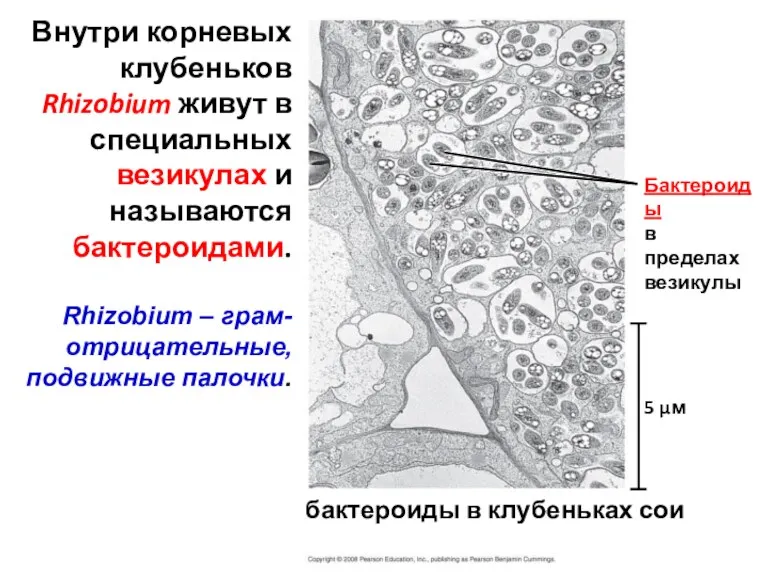
бактероиды в клубеньках сои
5 µм
Бактероиды
в пределах
везикулы
Внутри корневых клубеньков Rhizobium
бактероиды в клубеньках сои
5 µм
Бактероиды
в пределах
везикулы
Внутри корневых клубеньков Rhizobium
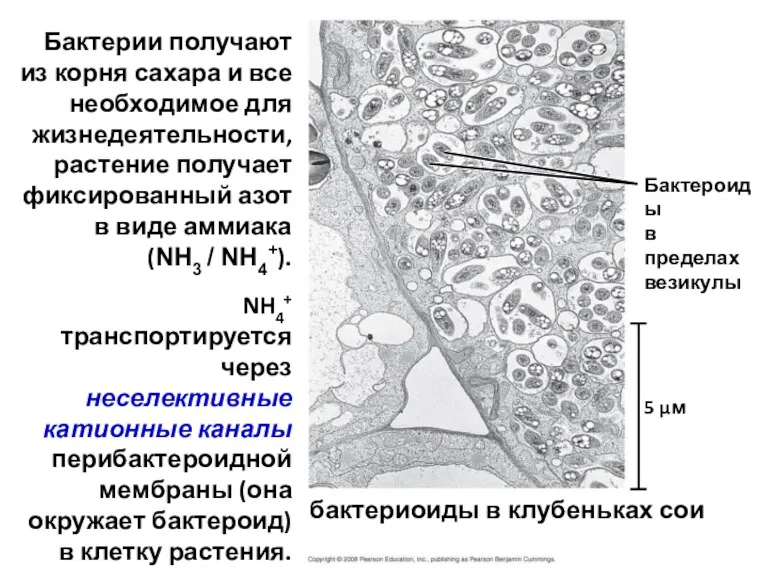
бактериоиды в клубеньках сои
5 µм
Бактероиды
в пределах
везикулы
Бактерии получают из корня
бактериоиды в клубеньках сои
5 µм
Бактероиды
в пределах
везикулы
Бактерии получают из корня
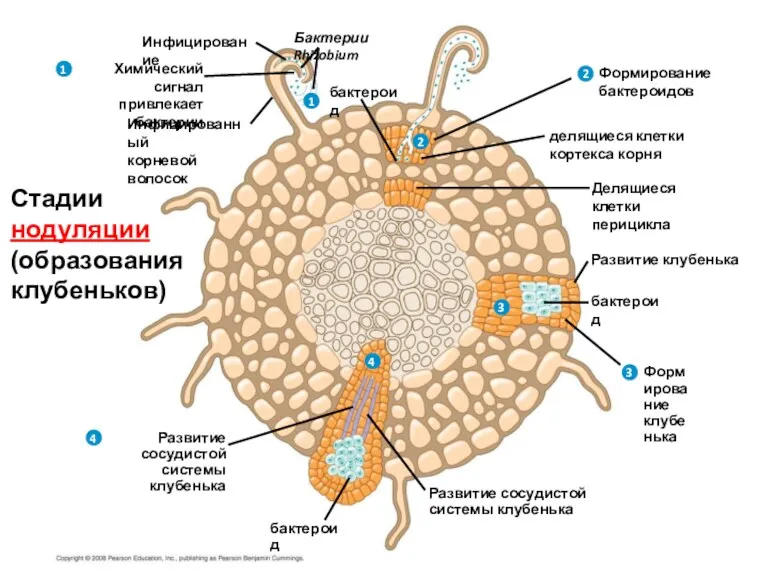
Бактерии Rhizobium
Инфицирование
Химический сигнал
привлекает бактерии
1
1
2
2
3
3
4
4
Инфицированный
корневой
волосок
бактероид
Формирование
бактероидов
делящиеся клетки
кортекса корня
Делящиеся
Бактерии Rhizobium
Инфицирование
Химический сигнал
привлекает бактерии
1
1
2
2
3
3
4
4
Инфицированный
корневой
волосок
бактероид
Формирование
бактероидов
делящиеся клетки
кортекса корня
Делящиеся

Нитрат, попадая в корневые клетки, восстанавливается до аммония, который затем включается
Нитрат, попадая в корневые клетки, восстанавливается до аммония, который затем включается

Цепочка превращений нитрата к аммиаку начинается на внешней поверхности корня и
Цепочка превращений нитрата к аммиаку начинается на внешней поверхности корня и
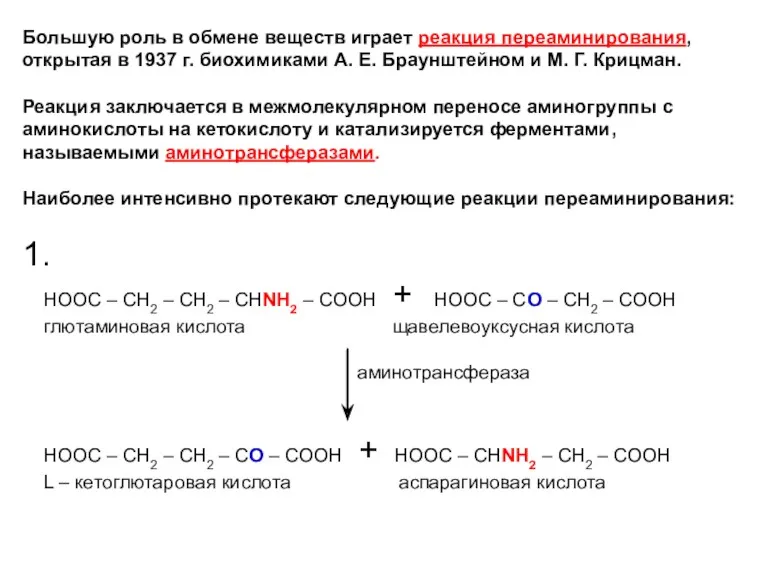
Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937
Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937
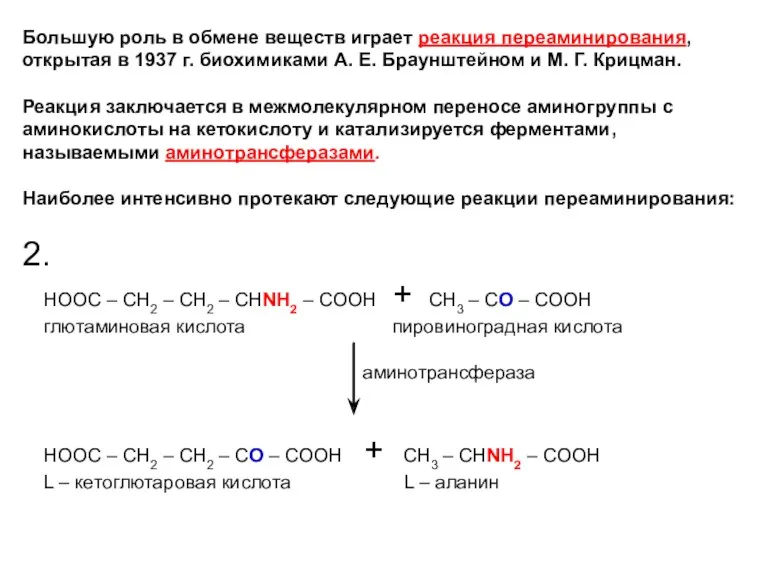
Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937
Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937

Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937
Большую роль в обмене веществ играет реакция переаминирования, открытая в 1937

Показано также, что под влиянием соответствующих аминотрансфераз аспарагин и глютамин также
Показано также, что под влиянием соответствующих аминотрансфераз аспарагин и глютамин также
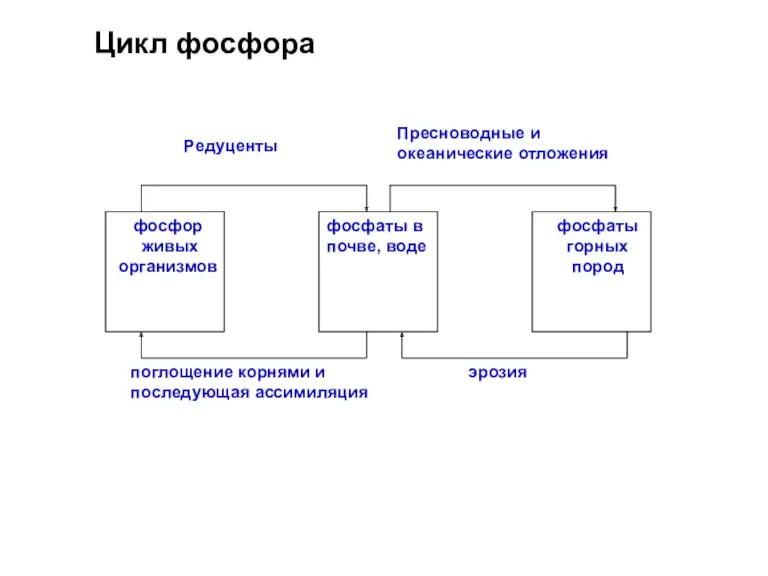
Цикл фосфора
Цикл фосфора
 Биологические модели развития популяций
Биологические модели развития популяций Надклас риби
Надклас риби Аграрний сервіс майбутнього
Аграрний сервіс майбутнього Влияние факторов внешней среды на микроорганизмы
Влияние факторов внешней среды на микроорганизмы Биология как наука. Методы научного познания
Биология как наука. Методы научного познания Анатомия и физиология печени. Лекция № 40
Анатомия и физиология печени. Лекция № 40 Строение и значение нервной системы
Строение и значение нервной системы Популяційна екологія
Популяційна екологія Перелетные птицы средней полосы России
Перелетные птицы средней полосы России Выращивание древесно-кустарниковых культур
Выращивание древесно-кустарниковых культур Фотосинтез: история изучения и условия фотосинтеза
Фотосинтез: история изучения и условия фотосинтеза Система инактивации лекарственных средств в организме человека
Система инактивации лекарственных средств в организме человека Надпочечная железа, glandula suprarenalis
Надпочечная железа, glandula suprarenalis Цели и задачи биотехнологии. История развития биотехнологии. Слагаемые биотехнологического процесса
Цели и задачи биотехнологии. История развития биотехнологии. Слагаемые биотехнологического процесса Повышение продуктивности свиней за счёт скармливания им пророщенного зерна ячменя, в период выращивания
Повышение продуктивности свиней за счёт скармливания им пророщенного зерна ячменя, в период выращивания Процесс дробления клетки
Процесс дробления клетки Способы количественного определения белка. (Лекция 10)
Способы количественного определения белка. (Лекция 10) Введение в изучение анатомии и физиологии человека
Введение в изучение анатомии и физиологии человека Презентация Этапы развития жизни на Земле (1)
Презентация Этапы развития жизни на Земле (1) Организм человека
Организм человека Презентация к уроку биологии 5 класс Как живые организмы переносят неблагоприятные условия
Презентация к уроку биологии 5 класс Как живые организмы переносят неблагоприятные условия Генотипическая изменчивость (наследственная)
Генотипическая изменчивость (наследственная) Вітаміни у нашому житті
Вітаміни у нашому житті Презентация кабинета биологии
Презентация кабинета биологии Типи взаємозв'язків між організмами
Типи взаємозв'язків між організмами Бесполое размножение организмов
Бесполое размножение организмов Витамины, авитаминозы, гиповитаминозы. История открытия витаминов. Многообразие и значение витаминов
Витамины, авитаминозы, гиповитаминозы. История открытия витаминов. Многообразие и значение витаминов Исторический обзор. Место рыб в системе животного мира
Исторический обзор. Место рыб в системе животного мира