Физиология возбудимых тканей. Понятия возбудимость и раздражимость, возбудимые и невозбудимые ткани. Раздражители презентация
- Физиология возбудимых тканей. Понятия возбудимость и раздражимость, возбудимые и невозбудимые ткани. Раздражители

Содержание
- 2. Все живые ткани способностью отвечать на воздействия внешней среды или нарушения их состояния изменением своей структуры
- 3. Строение и свойства плазматической мембраны. Нейроны и мышечные волокна, равно как и другие клетки, отделены от
- 4. Возникновение мембранного потенциала. С внутренней и наружной стороны мембраны находится фактически раствор солей, главными ионами которого
- 5. В итоге на мембране возникает двойной электрический слой – снаружи, главным образом, катионы Na+, изнутри анионы
- 6. 2.2. ПОТЕНЦИАЛ ДЕЙСТВИЯ: ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ, МЕХАНИЗМ ВОЗНИКНОВЕНИЯ. ВОССТАНОВИТЕЛЬНЫЙ ПЕРИОД. ЯВЛЕНИЕ АККОМОДАЦИИ ВОЗБУДИМОЙ ТКАНИ. Потенциал действия.
- 7. При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями
- 8. Кроме пика, в ПД различают два следовых потенциала - следовую де-поляризацию (следовой отрицательный потенциал) и следовую
- 9. Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону называется деполяризацией. Область электроположительности носит
- 10. В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы
- 11. Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости
- 12. 2. 3. ПАРАМЕТРЫ ВОЗБУДИМОСТИ ТКАНИ: ПОРОГ, ПОЛЕЗНОЕ ВРЕМЯ И ХРОНАКСИЯ, КРИТИЧЕСКИЙ НАКЛОН, ЛАБИЛЬНОСТЬ. Зависимость пороговой силы
- 14. Зависимость порога от крутизны нарастания силы раздражителя. Вели-чина порога раздражения нерва или мышцы зависит не только
- 15. В лаборатории показателем скорости аккомодации является та наименьшая крутизна нарастания тока, при которой стимул еще сохраняет
- 16. 2-4. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ЛОКАЛЬНОГО ПОТЕНЦИАЛА И ПОТЕН-ЦИАЛА ДЕЙСТВИЯ. ЗАКОН "ВСЕ ИЛИ НИЧЕГО". ФАЗОВЫЕ ИЗМЕНЕНИЯ ВОЗБУДИМО-СТИ НЕРВА
- 17. Закон "все или ничего". При изучении зависимости эффектов раздражения от силы приложенного стимула был установлен т.н.
- 18. Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД сопровождается многофазными изменениями возбудимости. Для
- 19. Во время локального ответа возбудимость повышена, так как мембрана деполяризована и разность между Е0 и Ек
- 20. 2-5. ДЕЙСТВИЕ ПОСТОЯННОГО ТОКА НА ТКАНЬ: ПОЛЯРНЫЙ ЗАКОН, ЭЛЕКТРОТОН, КАТОДИЧЕСКАЯ ДЕПРЕССИЯ Полярный закон действия тока. При
- 22. Скачать презентацию

Все живые ткани способностью отвечать на воздействия внешней среды или нарушения
Все живые ткани способностью отвечать на воздействия внешней среды или нарушения

Строение и свойства плазматической мембраны. Нейроны и мышечные волокна, равно как
Строение и свойства плазматической мембраны. Нейроны и мышечные волокна, равно как

Возникновение мембранного потенциала.
С внутренней и наружной стороны мембраны находится фактически
Возникновение мембранного потенциала.
С внутренней и наружной стороны мембраны находится фактически

В итоге на мембране возникает двойной электрический слой – снаружи, главным
В итоге на мембране возникает двойной электрический слой – снаружи, главным

2.2. ПОТЕНЦИАЛ ДЕЙСТВИЯ: ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ, МЕХАНИЗМ ВОЗНИКНОВЕНИЯ. ВОССТАНОВИТЕЛЬНЫЙ ПЕРИОД. ЯВЛЕНИЕ
2.2. ПОТЕНЦИАЛ ДЕЙСТВИЯ: ФАЗЫ ПОТЕНЦИАЛА ДЕЙСТВИЯ, МЕХАНИЗМ ВОЗНИКНОВЕНИЯ. ВОССТАНОВИТЕЛЬНЫЙ ПЕРИОД. ЯВЛЕНИЕ

При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень
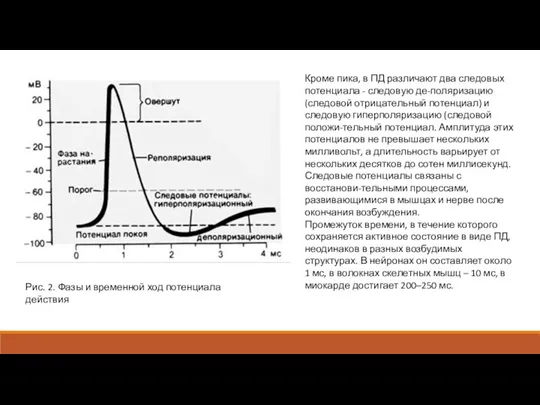
Кроме пика, в ПД различают два следовых потенциала - следовую де-поляризацию
Кроме пика, в ПД различают два следовых потенциала - следовую де-поляризацию

Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону
Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону

В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход

Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо
Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо

2. 3. ПАРАМЕТРЫ ВОЗБУДИМОСТИ ТКАНИ: ПОРОГ, ПОЛЕЗНОЕ ВРЕМЯ И ХРОНАКСИЯ, КРИТИЧЕСКИЙ
2. 3. ПАРАМЕТРЫ ВОЗБУДИМОСТИ ТКАНИ: ПОРОГ, ПОЛЕЗНОЕ ВРЕМЯ И ХРОНАКСИЯ, КРИТИЧЕСКИЙ
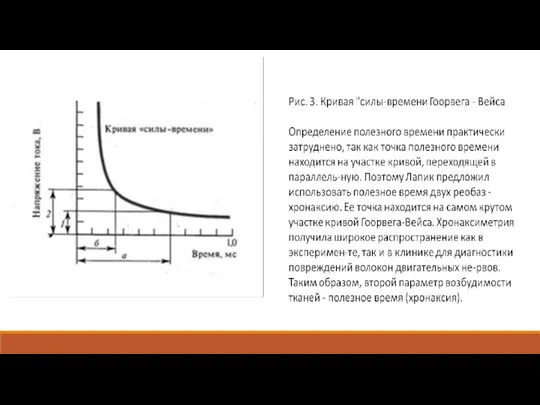

Зависимость порога от крутизны нарастания силы раздражителя.
Вели-чина порога раздражения нерва
Зависимость порога от крутизны нарастания силы раздражителя.
Вели-чина порога раздражения нерва

В лаборатории показателем скорости аккомодации является та наименьшая крутизна нарастания тока,
В лаборатории показателем скорости аккомодации является та наименьшая крутизна нарастания тока,

2-4. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ЛОКАЛЬНОГО ПОТЕНЦИАЛА И ПОТЕН-ЦИАЛА ДЕЙСТВИЯ. ЗАКОН "ВСЕ ИЛИ
2-4. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ЛОКАЛЬНОГО ПОТЕНЦИАЛА И ПОТЕН-ЦИАЛА ДЕЙСТВИЯ. ЗАКОН "ВСЕ ИЛИ

Закон "все или ничего".
При изучении зависимости эффектов раздражения от силы
Закон "все или ничего".
При изучении зависимости эффектов раздражения от силы
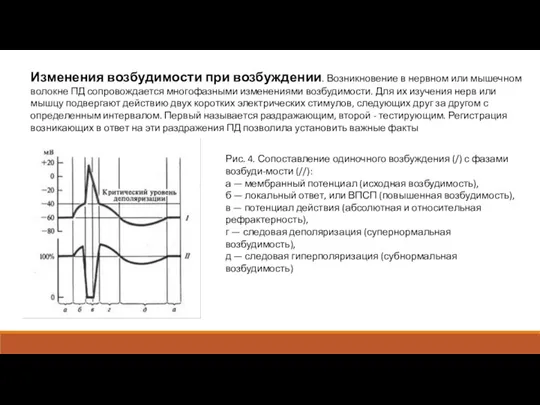
Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД
Изменения возбудимости при возбуждении. Возникновение в нервном или мышечном волокне ПД

Во время локального ответа возбудимость повышена, так как мембрана деполяризована и
Во время локального ответа возбудимость повышена, так как мембрана деполяризована и

2-5. ДЕЙСТВИЕ ПОСТОЯННОГО ТОКА НА ТКАНЬ: ПОЛЯРНЫЙ ЗАКОН, ЭЛЕКТРОТОН, КАТОДИЧЕСКАЯ ДЕПРЕССИЯ
Полярный
2-5. ДЕЙСТВИЕ ПОСТОЯННОГО ТОКА НА ТКАНЬ: ПОЛЯРНЫЙ ЗАКОН, ЭЛЕКТРОТОН, КАТОДИЧЕСКАЯ ДЕПРЕССИЯ
Полярный
 Голосеменные растения
Голосеменные растения Механизм и биологическое значение митоза и мейоза
Механизм и биологическое значение митоза и мейоза Тип Членистоногие
Тип Членистоногие Заяц- русак
Заяц- русак Тварини минулого
Тварини минулого Морфологическая характеристика лекарственных и ядовитых растений
Морфологическая характеристика лекарственных и ядовитых растений Особенности физиологии человека в условиях высокого и низкого давления
Особенности физиологии человека в условиях высокого и низкого давления Немембранные органоиды
Немембранные органоиды Происхождение человека (теории происхождения человека)
Происхождение человека (теории происхождения человека) Биологическая обработка органических отходов , их характеристика и принципы обработки
Биологическая обработка органических отходов , их характеристика и принципы обработки Тип Кишечнополостные
Тип Кишечнополостные Индивидуальное развитие организмов. Онтогенез
Индивидуальное развитие организмов. Онтогенез Общая характеристика пресмыкающихся
Общая характеристика пресмыкающихся Ордовикский период
Ордовикский период Настоящие многоклеточные
Настоящие многоклеточные Систематика бактерий
Систематика бактерий Индивидуальное развитие. Биогенетический закон
Индивидуальное развитие. Биогенетический закон Испарение воды растениями
Испарение воды растениями Ветеринарно - санітарна експертиза молока при інфекційних хворобах тварин і обсіменінні його патогенними мікроорганізмами
Ветеринарно - санітарна експертиза молока при інфекційних хворобах тварин і обсіменінні його патогенними мікроорганізмами Тип моллюски
Тип моллюски Перелетные птицы
Перелетные птицы Деревья леса
Деревья леса Презентация Методика решения задач по генетике
Презентация Методика решения задач по генетике ЕГЭ по биологии
ЕГЭ по биологии Передний мозг
Передний мозг Пізнавальна діяльність людини. Сигнальні системи. Відчуття та сприйняття
Пізнавальна діяльність людини. Сигнальні системи. Відчуття та сприйняття Медицинская арахноэнтомология
Медицинская арахноэнтомология Мускулатура тазовой конечности
Мускулатура тазовой конечности