- Физиология высшей нервной деятельности

Содержание
- 2. Цель лекции: Дать механизмы развития сна и сновидений, сознательного и бессознательного Мотивация лекции: Данный раздел физиологии
- 3. План лекции: - физиология сна, сновидений и гипноза - физиология сознательного и бессознательного - физиология боли
- 4. Сон - физиологическое состояние неподвижности с ослабленным тонусом мышц и резко ограниченным сенсорным контактом с внешней
- 5. Сон – это состояние, характеризующееся: отсутствием моторной активности, пониженным уровнем сознания (у человека), внешним покоем, характерной
- 6. Функции сна: покой и отдых, восстановление сил организма, переработка дневной информации и перевод её в долговременную
- 7. Виды сна Ежесуточный естественный Монофазный и полифазный сон Медленный или ортодоксальный сон Быстрый или парадоксальный сон
- 8. Фазы сна Стадия дремоты - постепенное замещение альфа-ритма низкоамплитудными тета-волнами Стадия сонных веретен - между двух-трехфазными
- 9. Соотношение сна и бодрствования, а также БДГ– и медленноволнового сна в различные периоды жизни человека. Наиболее
- 10. Основные формы ЭЭГ основной α–ритм (α–волны с частотой 8–13 Гц, в среднем 10 Гц), β–ритм частотой
- 11. Классификация стадий сна у человека по особенностям ЭЭГ по Лумису Стадия W — бодрствование в расслабленном
- 16. Центры сна Синхронизирующие (сомногенные) структуры - структуры вокруг сильвиевого водопровода и задней стенки IV желудочка, -
- 18. Нервные теории сна Гесс: в ядрах гипоталамуса существует центр сна. Поражение гипоталамуса – увеличение сонливости. Павлов:
- 20. Представление о нейрофизиологии и нейрохимии сна А — бодрствование; Б — медленноволновая фаза сна; В —
- 21. Структуры мозга, принимающие участие в регуляции уровня бодрствования и глубины сна (по А. Н. Шеповальникову) 1
- 22. Физиология эндокринной системы
- 23. Классические эндокринные железы (голубовато-зеленые) и некоторые органы диффузной эндокринной системы (фиолетовые точки). С – клетки ЩЖ
- 24. Эндокринная система 1. Эндокринные железы ГИПОФИЗ (аденогипофиз и нейрогипофиз) НАДПОЧЕЧНИКИ (кора и мозговое в-во) ЩИТОВИДНАЯ ЖЕЛЕЗА
- 25. Железы внутренней секреции и их гормоны
- 28. Химическая природа и пути действия гормонов Простые и сложные белки, - через мембранные пептиды - гормоны
- 30. Виды действия гормонов Метаболическое Морфогенетическое Кинетическое Корригирующее
- 31. Нейроэндокринные системы регуляции гормональной системы по принципу механизма обратной отрицательной связи, тормозящих секрецию гипоталамических и гипофизарных
- 32. Динамики секреции в гормональных системах происходит в соответствии с характерными ритмами (от минут до периодов жизни),
- 34. Нейроны гипоталамуса Обладают рецепторной функцией и способны улавливать изменения химического состава крови и цереброспинальной жидкости, что
- 35. Двусторонние связи гипоталамуса с: Таламусом Лимбической системой КБП Центральным серым веществом среднего мозга Соматическими ядрами ствола
- 36. Гипоталамус Вырабатывает собственные гормоны Эффекторные: окситоцин и вазопрессин (супраоптическое и паравентрикулярное) Релизинг-гормоны: либерины и статины
- 39. Гормоны гипофизотропной части гипоталамуса выделяются в области срединного возвышения и через воротную систему гипофиза транспортируются в
- 40. Топография циркумвентрикулярных органов (ЦВО), которые находятся вне гематоэнцефалического барьера и гормоны могут действовать здесь как гуморальные
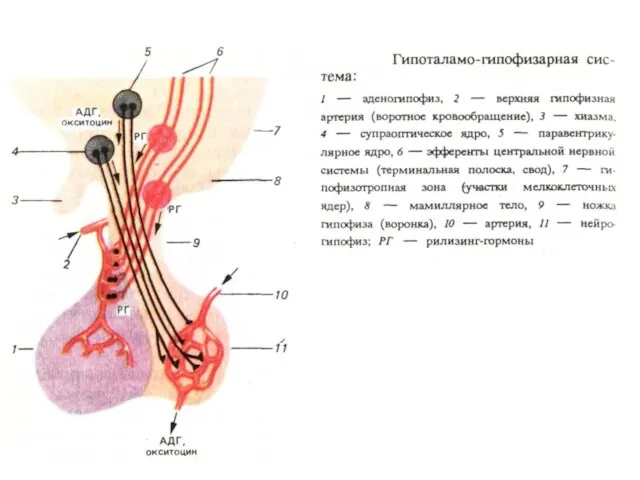
- 41. Гуморальные связи ЦНС с периферией организма. Слева: варианты выделения гормонов 1- гипофизтропные гормоны определяют высвобождение гормонов
- 43. Система гормональной регуляции организма позвоночных — гипоталамо—гипофизарная система; — эндокринные железы—мишени гормонов аденогипофиза. Висперотропные нейрогормоны: АДГ
- 44. Задняя доля гипофиза состоит из терминалей аксонов крупных клеток супраоптического и паравентрикулярного ядер (СЯ и ПЯ
- 46. Основные принципы регуляции секреции аденогипофизарных гормонов (нижние прямоугольники) со стороны гипоталамуса (верхний прямоугольник).
- 47. Прямые и обратные связи в системе нейроэндокринной регуляции 1 — медленно развивающееся и продолжительное ингибирование секреции
- 48. Рилизинг–гормоны группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза. Их подразделяют на либерины и
- 49. А: либерины и статины, выделяемые окончаниями нейросекреторных клеток в медиальном возвышении, поступают по портальным сосудам в
- 50. Соматостатин 1 – в передней доле гипофиза подавляет синтез и секрецию СТГ, АКТГ и ТТГ; 2
- 51. Соматолиберин синтезируется нейросекреторными нейронами дугового ядра (n. arcuatus) гипоталамуса и некоторыми опухолями островковых клеток поджелудочной железы
- 52. Гонадолиберин Мишени гонадолиберина — гонадотрофы передней доли гипофиза. Функции: Гонадолиберин — ключевой нейрорегулятор репродуктивной функции, стимулирует
- 53. Пролактиностатин Мишени пролактиностатина — лактотрофы передней доли гипофиза. Функции: Пролактиностатин подавляет секрецию пролактина из лактотрофных клеток
- 54. Тиреолиберин синтезируется многими нейронами ЦНС (в т.ч. нейросекреторными нейронами паравентрикулярного ядра). Мишени — тиреотрофы и лактотрофы
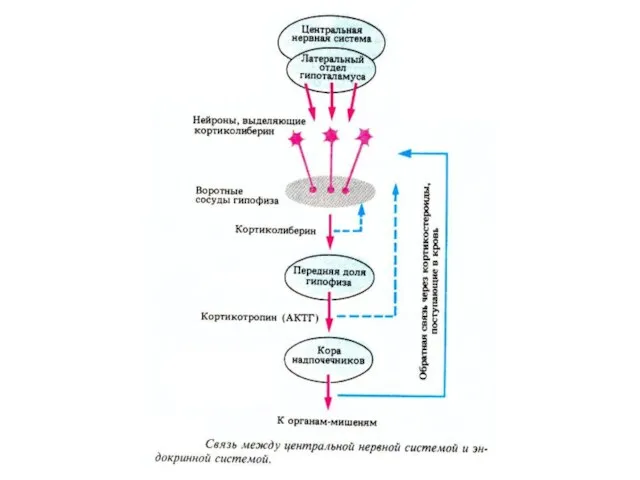
- 55. Кортиколиберин синтезируется в нейросекреторных нейронах паравентрикулярного ядра гипоталамуса, плаценте, Т-лимфоцитах. Глюкокортикоиды подавляют синтез гипоталамического кортиколиберина и
- 56. Кортиколиберин Функции: 1. Передняя доля гипофиза - стимуляция синтеза и секреции АКТГ 2. Во время стресса
- 57. Меланостатин подавляет образование меланотропинов.
- 58. Дофамин Промежуточный продукт метаболизма тирозина и предшественник норадреналина и адреналина — поступающий к клеткам передней доли
- 60. Гормоны задней доли гипофиза Вазопрессин и окситоцин синтезируются в нейросекреторных нейронах паравентрикулярного и супраоптического ядер гипоталамуса,
- 61. Вазопрессин (АДГ) Оказывает антидиуретический и сосудосуживающий эффекты (повышает системное АД). Главная функция АДГ — регуляция обмена
- 62. Регуляция секреции АДГ Стимулируют секрецию: 1. гиповолемия, 2. гиперосмоляльность, 3. переход в вертикальное положение, 4. стресс,
- 63. Окситоцин: - стимулирует сокращение ГМК миометрия. - стимулирует сокращение миоэпителиальных клеток альвеол лактирующей молочной железы (рефлекс
- 64. Гормоны передней доли гипофиза По химической структуре гормоны аденогипофиза являются либо пептидами, либо гликопротеинами. Пептидные гормоны:
- 65. СТГ Рецепторы СТГ относятся к семейству мембранных цитокиновых рецепторов. Регуляция синтеза СТГ происходит за счет обратной
- 66. Метаболические эффекты СТГ 1. Начальная фаза (инсулиноподобный эффект): А) СТГ увеличивает поглощение глюкозы мышцами и жировой
- 67. Секрецию СТГ стимулируют: Уменьшение глюкозы в крови, Уменьшение свободных ЖК в крови, Голодание, белковая недостаточность, Травма,
- 68. Регуляция секреции АКТГ Синтез и секрецию АКТГ стимулирует кортиколиберин. Глюкокортикоиды в высоких дозах - ингибируют секрецию
- 69. АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (преимущественно кортизола). Гиперсекреция АКТГ ведёт к гиперплазии коры
- 70. Секрецию пролактина стимулируют: тиреолиберин, стимуляция соска и околососкового поля. стресс. подавляют: пролактиностатин дофамин.
- 71. Функции пролактина Главная функция пролактина — стимуляция функции молочной железы. Рецептор пролактина — мембранный цитокиновый рецептор.
- 72. Тиреотропный гормон (ТТГ) Соматостатин подавляет секрецию ТТГ. Тиреолиберин стимулирует синтез и секрецию ТТГ. Гормоны щитовидной железы
- 73. Гонадотропные гормоны: фолликулостимулирующий гормон (ФСГ) лютеинизирующий гормон (ЛГ), хорионический гонадотропин (ХГТ) плаценты. ФСГ: - у женщин
- 74. Гонадолиберин стимулирует синтез и секрецию ФСГ и ЛГ. a- и b-ингибины (фолликулы яичника и клетки Сертоли
- 75. Пролактин, его функции и регуляция. Прямой контроль секреции пролактина с помощью механизма отрицательной обратной связи не
- 76. Гормон роста СТГ, его функции и регуляция. СТГ стимулирует в печени секрецию ИФР-1 (соматомедин С) и
- 77. Множественные функции соматостатина. Соматостатин присутствует во многих тканях. Он действует: Через кровь как гормон Паракринным путем
- 78. Множественное действие кортикотропин-РГ (КРГ) КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует также в управлении центрами
- 79. Функции антидиуретического гормона (вазопрессина). АДГ действует на ЦНС как нейротрансмиттер и нейромодулятор, на ПОМК-клетки аденогипофиза –
- 80. Взаимодействие гипоталамуса и гипофиза
- 81. Гормоны гипофиза и их функции
- 82. Щитовидная железа
- 83. Фолликулы щитовидной железы при неактивном (А) и активном (Б) состояниях железы А — эпителий плоский, фолликулы
- 85. В клетках щитовидной железы происходит синтез двух классов гормонов: йодсодержащих и кальцитониновых. Йодсодержащие гормоны, Кальцитонин, Катакальцин,
- 86. Молекула тиреоглобулина и строение основных гормонов щитовидной железы. Представленные три- или тетрайодтиронины, а также моно- и
- 87. Тироксин и трийодтиронин Йодсодержащие гормоны - тироксин (T4) и трийодтиронин (Т3) Образование Т3 из тироксина частично
- 88. Функции йодсодержащих гормонов Необходимы для нормального развития ЦНС, Увеличивают обменные процессы, ускоряют катаболизм белков, жиров и
- 89. Кальцитонин и катакальцин Их функции антагонистичны эффектам ПТГ: кальцитонин уменьшает [Са2+] в крови, стимулирует минерализацию кости,
- 90. относящиеся к кальцитониновому гену, участвуют в ноцицепции, пищевом поведении, а также в регуляции тонуса сосудов. Рецепторы
- 91. Уровень гормонов щитовидной железы и связывающих белков в плазме и функциональные тесты. А) приведены характерные для
- 92. Связи щитовидной железы у млекопитающих
- 94. Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иоде покрывается за счет всасывания из в
- 97. Транспорт тироидного гормона в сыворотке крови (тироксин связывающий глобулин, транстиретин, альбумин)
- 98. Гормоны щитовидной (1) и паращитовидных (2) желез и их функции
- 99. Cуточная потребность в йоде покрывается за счет всасывания в кишечнике в виде ионов. Последние поглощаются клетками
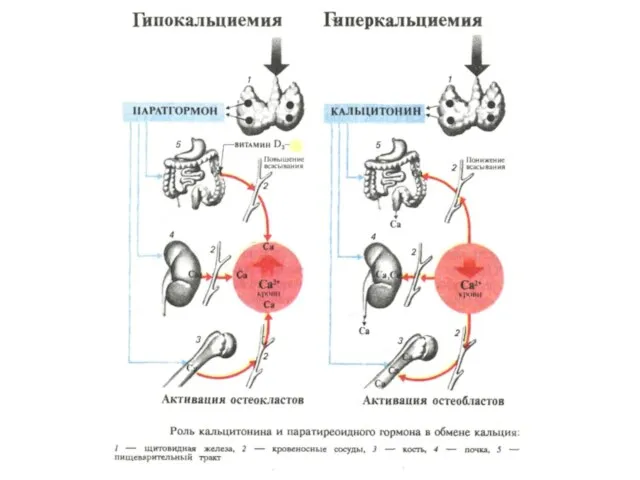
- 100. Регуляция содержания кальция в крови кальцитонином и паратгормоном
- 102. Эпифиз
- 103. Эпифиз Гормоны эпифиза представлены в основном индоламинами (серотонин и мелатонин), пептидами и белками. Выделяясь в ликвор
- 104. Гормон эпифиза — мелатонин Кроме того: снижение метаболизма в светочувствительных нейронах СХЯ, иннервирующих и активирующих все
- 105. НАДПОЧЕЧНИКИ
- 106. Схема зон надпочечника и вырабатываемые ими гормоны 1 — пучковая зона, 2 — сетчатая зона, 3
- 107. Строение и функции надпочечника Три слоя надпочечников имеют характерный набор ферментов обеспечивающий синтез специфических конечных продуктов:
- 109. ГОРМОНЫ НАДПОЧЕЧНИКА Кора надпочечника Мозговое вещество КОРТИЗОЛ КОРТИКОСТЕРОН АЛЬДОСТЕРОН ДЕЗОКСИКОРТИКОСТЕРОН ДЕГИДРОЭПИАНДРОСТЕРОН АНДРОСТЕНДИОН АДРЕНАЛИН НОРАДРЕНАЛИН
- 110. Глюкокортикоиды Основной глюкокортикоид – кортизол Регуляция секреции Активирующие (нисходящие) влияния. Непосредственный активатор синтеза и секреции кортизола
- 111. Углеводный обмен. Стимуляция глюконеогенеза и уменьшение потребления глюкозы внутренними органами (кроме головного мозга). Основной эффект —
- 112. Липидный обмен. Липолиз усиливается в конечностях. Липогенез усиливается в других частях тела (туловище и лицо) (синдром
- 113. - В высоких дозах выступают как иммунодепрессанты (применяют для предупреждения отторжения трансплантатов). - имеют выраженный противовоспалительный
- 114. Основной минералокортикоид - альдостерон Регуляция секреции альдостерона : Ангиотензин II — главный регулятор, стимулирует выброс альдостерона.
- 115. Функции альдостерона: - увеличивает реабсорбцию Na+. Задержка Na+ приводит к увеличению содержания воды в организме и
- 116. Гормоны надпочечников
- 118. Система гипоталамус гипофиз–кора надпочечников.
- 119. Паракринное действие кортизола на синтез катехоламинов в мозговом веществе надпочечников. Важный этап синтеза от тирозина к
- 120. Регуляция оси «гипоталамус – гипофиз – кора надпочечников», обратная связь с участием кортизола, а также взаимодействия
- 121. Множественное действие КРГ. КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует в управлении центрами ВНС. Действуя
- 122. МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ СЛИЗИСТОЙ ОБОЛОЧКИ
- 123. СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ Реакция тревоги СТАДИЯ РЕЗИСТЕНТНОСТИ СТАДИЯ ИСТОЩЕНИЯ ИСХОДНЫЙ СРЕДНИЙ УРОВЕНЬ УСТОЙЧИВОСТИ
- 124. ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
- 125. В островках Лангерганса идентифицировано несколько типов эндокринных клеток, синтезирующих и секретирующих пептидные гормоны: инсулин (b‑клетки, 70%
- 126. Действие инсулина на углеводный обмен активация утилизации глюкозы клетками, усиление фосфорилирования; подавление распада гликогена; стимуляция синтеза
- 127. Действие инсулина на белковый обмен повышение проницаемости мембран для аминоокислот; усиление синтеза иРНК; активация в печени
- 128. Действие инсулина на липидный обмен стимуляция синтеза свободных жирных кислот из глюкозы; стимуляция синтеза липопротеиновой липазы
- 129. Скорость проявления эффектов инсулина Физиологические эффекты инсулина по скорости их наступления подразделяют на: Быстрый (секунды) Увеличение
- 130. Регуляция Глюкоза - ведущий регулятор секреции инсулина: при участии Са2+ стимулирует секрецию инсулина и тормозит секрецию
- 131. Гомеостаз глюкозы поддерживают следующие механизмы: Печень демпфирует колебания концентрации глюкозы за счет обратимой реакции глюкоза -
- 132. Функциональная организация островков Лангерганса сплошные стрелки — стимуляция, пунктирные — подавление гормональных секретов. Ведущий регулятор —
- 133. Сахарный диабет типа I развивается в результате аутоиммунной деструкции b-клеток островков Лангерганса и значительно реже вследствие
- 134. Сахарный диабет типа II развивается в результате : либо нарушения секреции инсулина (избыточное содержание сахара в
- 135. Тимус Тимус (вилочковая железа) является центральным органом иммунитета, обеспечивающим продукцию специфических Т-лимфоцитов. Тимоциты секретируют в кровь
- 136. Яички В яичках синтезируются стероидные андрогены и a‑ингибин. Тестостерон — основной циркулирующий андроген. В эмбриогенезе андрогены
- 137. Яичники В яичниках синтезируются стероидные женские половые гормоны, гликопротеиновые гормоны ингибины и пептидной природы релаксины. Эстрогены
- 138. Гормоны, вырабатываемые в плаценте Плацента тесно анатомически и функционально связана с организмами матери и плода. Синтез
- 139. Гормоны, вырабатываемые в почке Ренин не является гормоном, этот фермент — начальное звено в ренин-ангиотензиновой системе,
- 140. Гормоны, вырабатываемые в сердце Натрийуретический фактор синтезируют кардиомиоциты правого предсердия и некоторые нейроны ЦНС. Мишени —
- 141. Гормоны, вырабатываемые в ЖКТ В стенке трубчатых органов ЖКТ присутствует огромное количество секретирующих гормоны разнообразных эндокринных
- 142. АПУД-система APUD — аббревиатура, образованная из первых букв англ. слов amines - амины, precursor - предшественник,
- 143. АПУД-система Предполагают, что единым эмбриональным предшественником клеток АПУД-системы является так называемый нейроэндокринно-программированный эпибласт. Кроме этого апудоциты
- 144. Гастроэнтеропанкреатическая эндокринная система
- 145. Гормоны поджелудочной железы и их функции: α- альфа-клетки, продуцирующие глюкогон, β – бетта-клетки, продуцирующие инсулин, D
- 146. Регуляция активности островковых клеток поджелудочной железы
- 148. Скачать презентацию

Цель лекции:
Дать механизмы развития сна и сновидений, сознательного и бессознательного
Мотивация лекции:
Данный
Цель лекции:
Дать механизмы развития сна и сновидений, сознательного и бессознательного
Мотивация лекции:
Данный

План лекции:
- физиология сна, сновидений и гипноза
- физиология сознательного и бессознательного
План лекции:
- физиология сна, сновидений и гипноза
- физиология сознательного и бессознательного

Сон - физиологическое состояние неподвижности с ослабленным тонусом мышц и резко
Сон - физиологическое состояние неподвижности с ослабленным тонусом мышц и резко

Сон – это состояние, характеризующееся:
отсутствием моторной активности,
пониженным уровнем сознания (у человека),
внешним
Сон – это состояние, характеризующееся:
отсутствием моторной активности,
пониженным уровнем сознания (у человека),
внешним

Функции сна:
покой и отдых,
восстановление сил организма,
переработка дневной информации и перевод её
Функции сна:
покой и отдых,
восстановление сил организма,
переработка дневной информации и перевод её

Виды сна
Ежесуточный естественный
Монофазный и полифазный сон
Медленный или ортодоксальный сон
Быстрый или парадоксальный
Виды сна
Ежесуточный естественный
Монофазный и полифазный сон
Медленный или ортодоксальный сон
Быстрый или парадоксальный

Фазы сна
Стадия дремоты - постепенное замещение альфа-ритма низкоамплитудными тета-волнами
Стадия сонных
Фазы сна
Стадия дремоты - постепенное замещение альфа-ритма низкоамплитудными тета-волнами
Стадия сонных
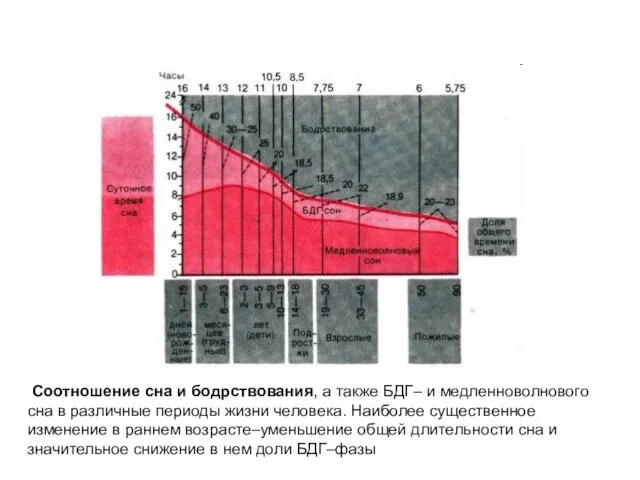
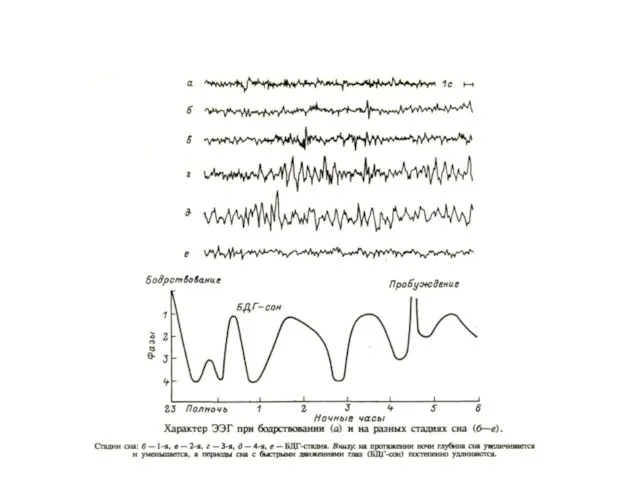
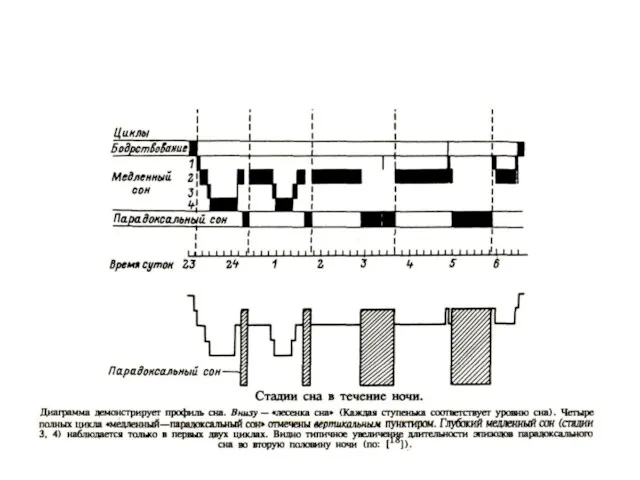
Соотношение сна и бодрствования, а также БДГ– и медленноволнового сна
Соотношение сна и бодрствования, а также БДГ– и медленноволнового сна
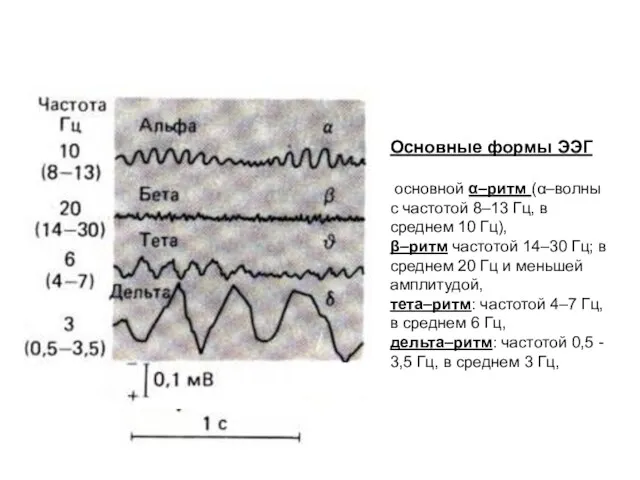
Основные формы ЭЭГ
основной α–ритм (α–волны с частотой 8–13 Гц,
Основные формы ЭЭГ
основной α–ритм (α–волны с частотой 8–13 Гц,
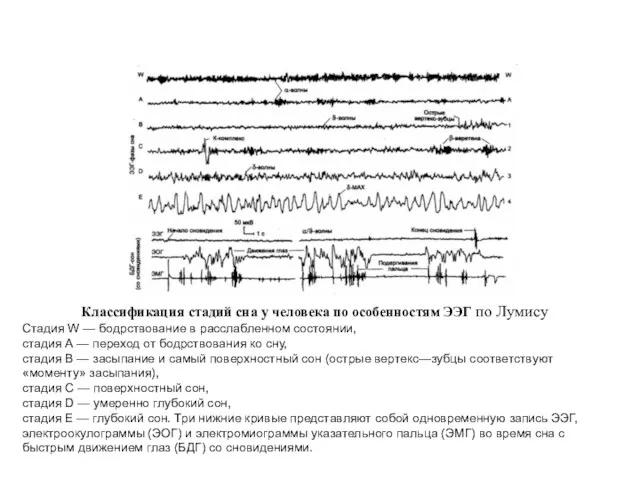
Классификация стадий сна у человека по особенностям ЭЭГ по Лумису
Стадия
Классификация стадий сна у человека по особенностям ЭЭГ по Лумису
Стадия


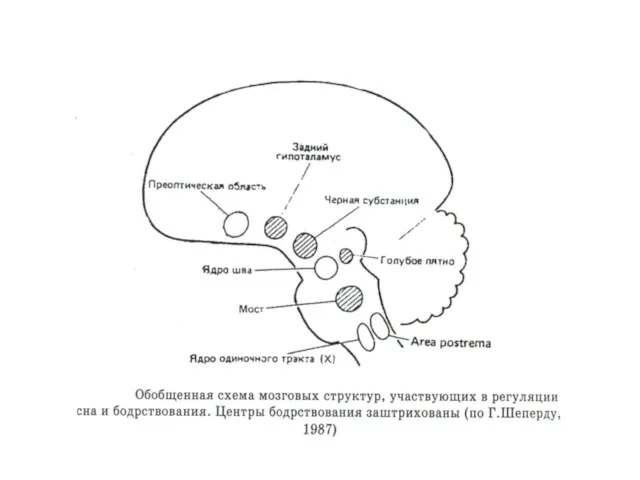
Центры сна
Синхронизирующие (сомногенные) структуры
- структуры вокруг сильвиевого водопровода и
Центры сна
Синхронизирующие (сомногенные) структуры
- структуры вокруг сильвиевого водопровода и

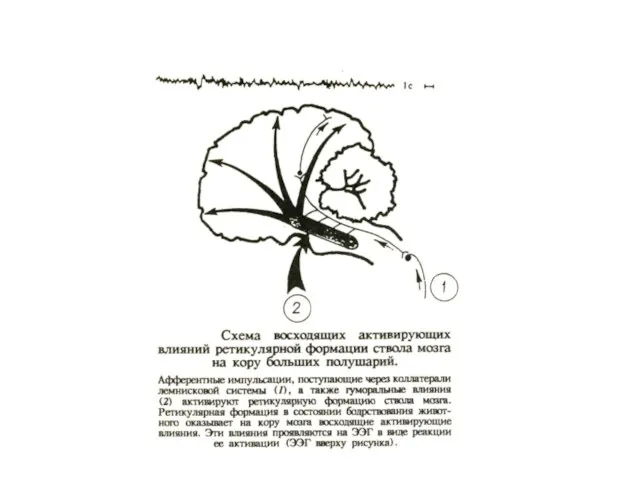
Нервные теории сна
Гесс: в ядрах гипоталамуса существует центр сна. Поражение
Нервные теории сна
Гесс: в ядрах гипоталамуса существует центр сна. Поражение
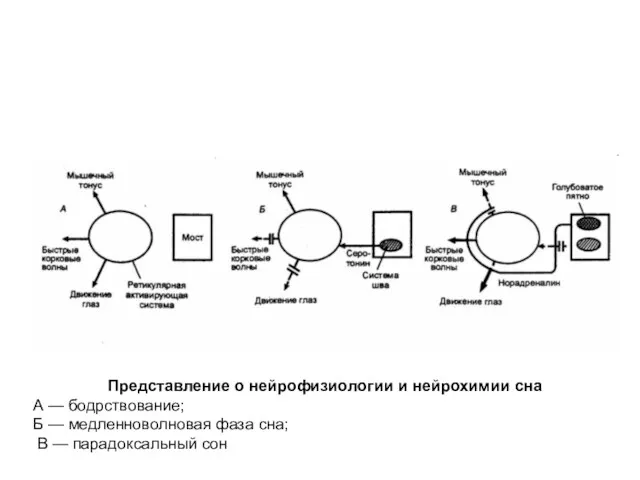
Представление о нейрофизиологии и нейрохимии сна
А — бодрствование;
Б —
Представление о нейрофизиологии и нейрохимии сна
А — бодрствование;
Б —
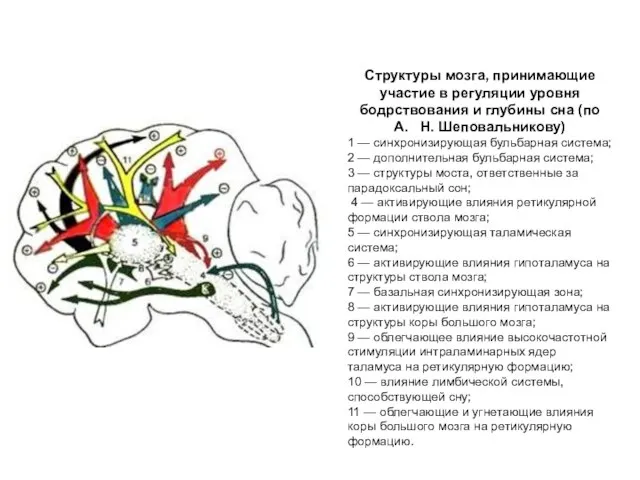
Структуры мозга, принимающие участие в регуляции уровня бодрствования и глубины сна
Структуры мозга, принимающие участие в регуляции уровня бодрствования и глубины сна
Физиология
эндокринной
системы
Физиология
эндокринной
системы
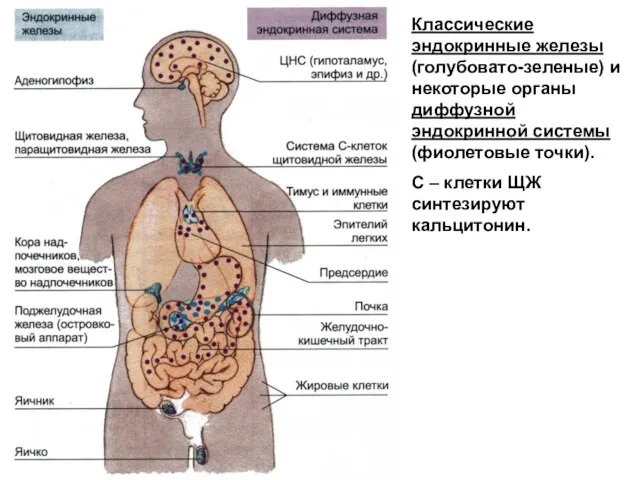
Классические эндокринные железы (голубовато-зеленые) и некоторые органы диффузной эндокринной системы (фиолетовые
Классические эндокринные железы (голубовато-зеленые) и некоторые органы диффузной эндокринной системы (фиолетовые

Эндокринная система
1. Эндокринные железы
ГИПОФИЗ (аденогипофиз и нейрогипофиз)
НАДПОЧЕЧНИКИ (кора и мозговое в-во)
ЩИТОВИДНАЯ
Эндокринная система
1. Эндокринные железы
ГИПОФИЗ (аденогипофиз и нейрогипофиз)
НАДПОЧЕЧНИКИ (кора и мозговое в-во)
ЩИТОВИДНАЯ
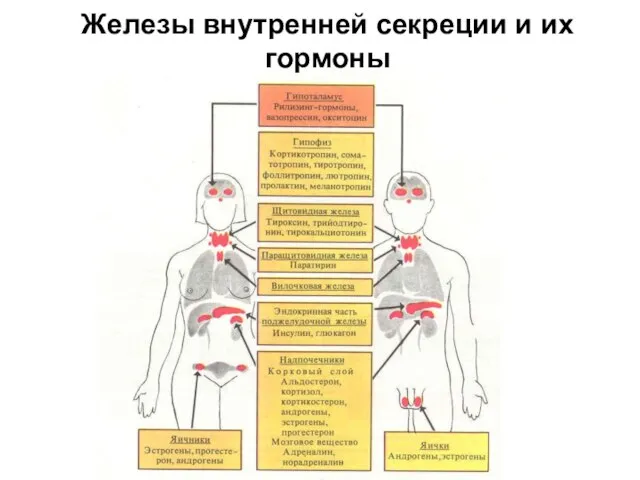
Железы внутренней секреции и их гормоны
Железы внутренней секреции и их гормоны

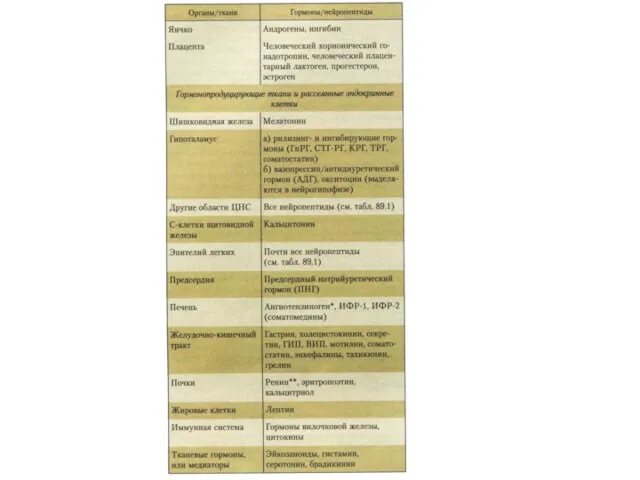

Химическая природа и пути действия гормонов
Простые и сложные белки, - через
Химическая природа и пути действия гормонов
Простые и сложные белки, - через

Виды действия гормонов
Метаболическое
Морфогенетическое
Кинетическое
Корригирующее
Виды действия гормонов
Метаболическое
Морфогенетическое
Кинетическое
Корригирующее
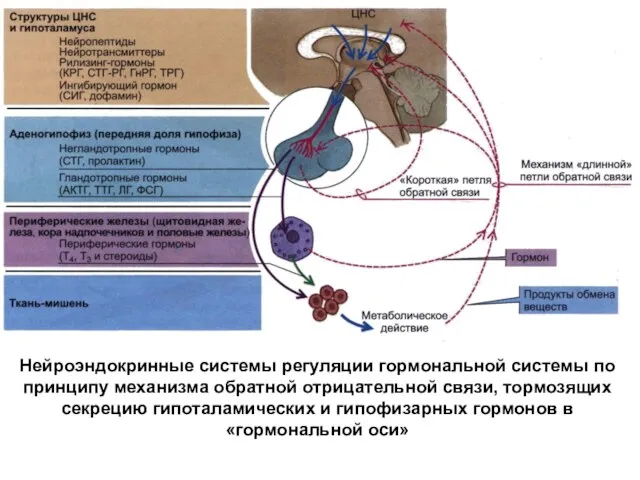
Нейроэндокринные системы регуляции гормональной системы по принципу механизма обратной отрицательной связи,
Нейроэндокринные системы регуляции гормональной системы по принципу механизма обратной отрицательной связи,
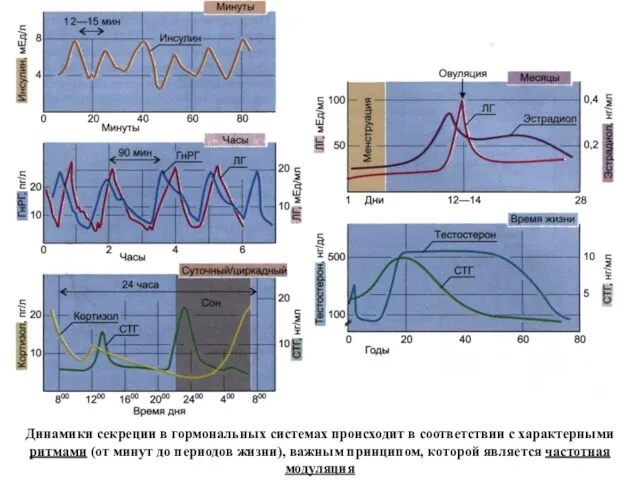
Динамики секреции в гормональных системах происходит в соответствии с характерными ритмами
Динамики секреции в гормональных системах происходит в соответствии с характерными ритмами

Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливать изменения химического состава крови
Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливать изменения химического состава крови

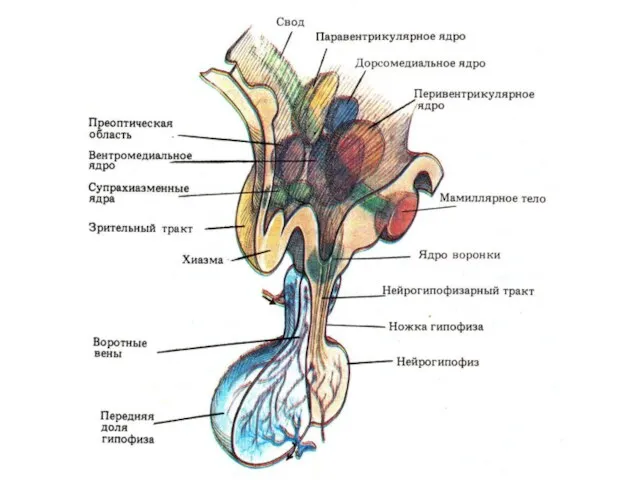
Двусторонние связи гипоталамуса с:
Таламусом
Лимбической системой
КБП
Двусторонние связи гипоталамуса с:
Таламусом
Лимбической системой
КБП

Гипоталамус
Вырабатывает собственные гормоны
Эффекторные: окситоцин и вазопрессин (супраоптическое и паравентрикулярное)
Гипоталамус
Вырабатывает собственные гормоны
Эффекторные: окситоцин и вазопрессин (супраоптическое и паравентрикулярное)
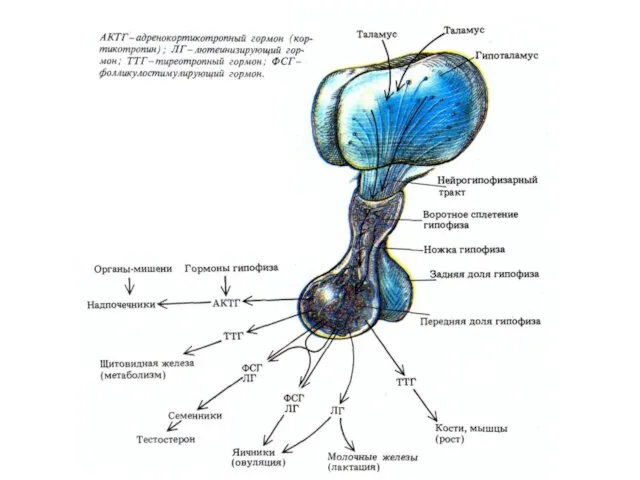
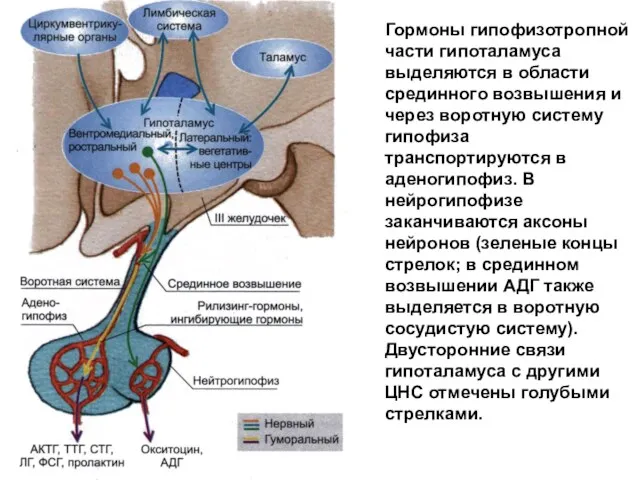
Гормоны гипофизотропной части гипоталамуса выделяются в области срединного возвышения и через
Гормоны гипофизотропной части гипоталамуса выделяются в области срединного возвышения и через
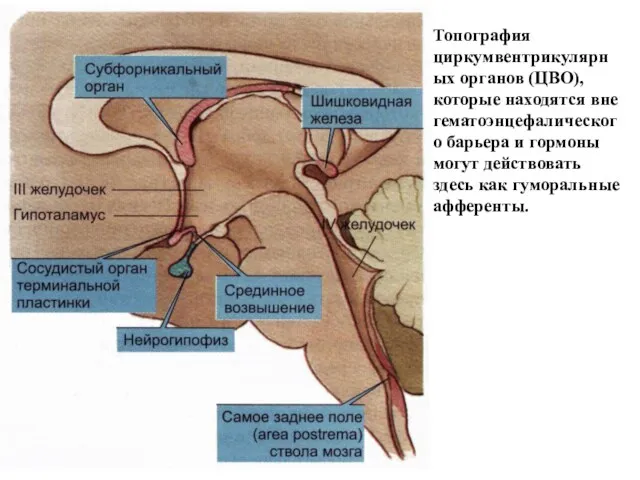
Топография циркумвентрикулярных органов (ЦВО), которые находятся вне гематоэнцефалического барьера и гормоны
Топография циркумвентрикулярных органов (ЦВО), которые находятся вне гематоэнцефалического барьера и гормоны
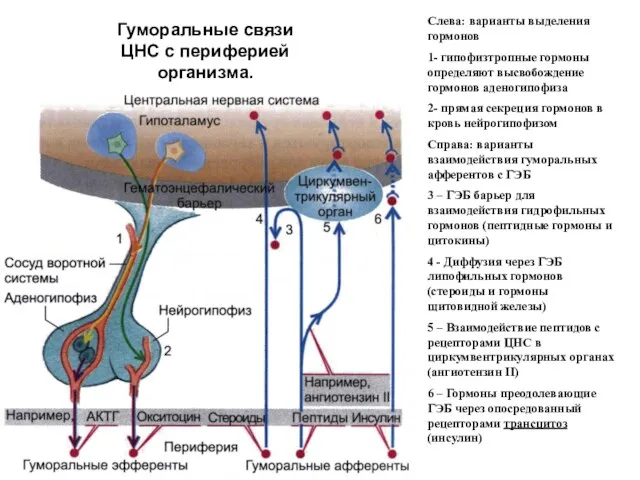
Гуморальные связи ЦНС с периферией организма.
Слева: варианты выделения гормонов
1- гипофизтропные гормоны
Гуморальные связи ЦНС с периферией организма.
Слева: варианты выделения гормонов
1- гипофизтропные гормоны
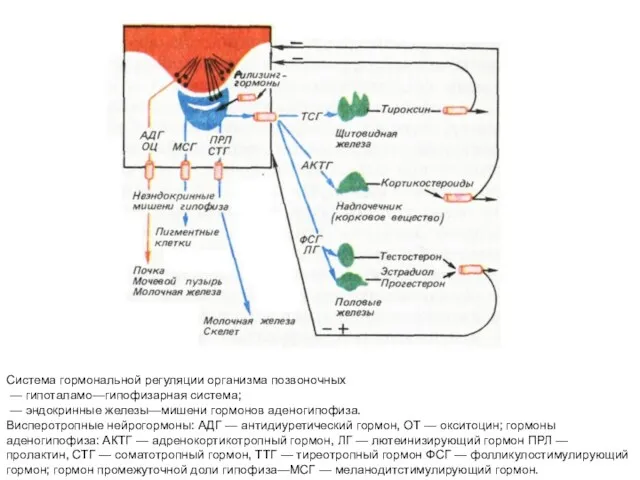
Система гормональной регуляции организма позвоночных
— гипоталамо—гипофизарная система;
—
Система гормональной регуляции организма позвоночных
— гипоталамо—гипофизарная система;
—
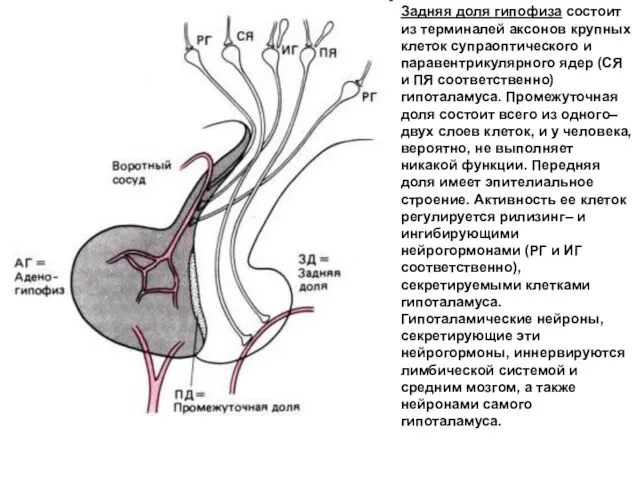
Задняя доля гипофиза состоит из терминалей аксонов крупных клеток супраоптического и
Задняя доля гипофиза состоит из терминалей аксонов крупных клеток супраоптического и
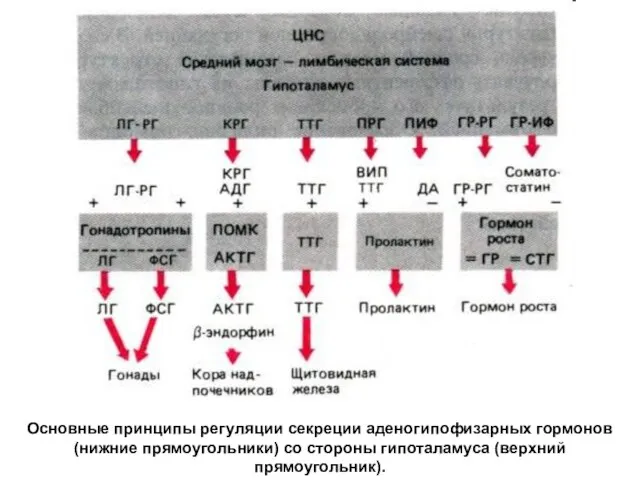
Основные принципы регуляции секреции аденогипофизарных гормонов (нижние прямоугольники) со стороны гипоталамуса
Основные принципы регуляции секреции аденогипофизарных гормонов (нижние прямоугольники) со стороны гипоталамуса
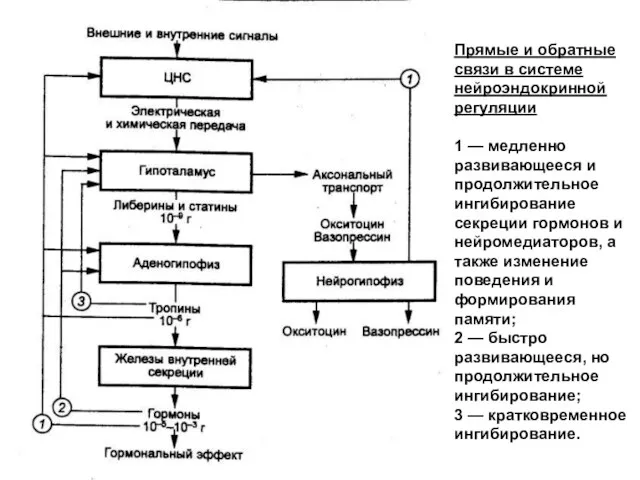
Прямые и обратные связи в системе нейроэндокринной регуляции
1 — медленно
Прямые и обратные связи в системе нейроэндокринной регуляции
1 — медленно

Рилизинг–гормоны
группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза.
Их
Рилизинг–гормоны
группа нейрогормонов, мишенями которых являются эндокринные клетки передней доли гипофиза.
Их
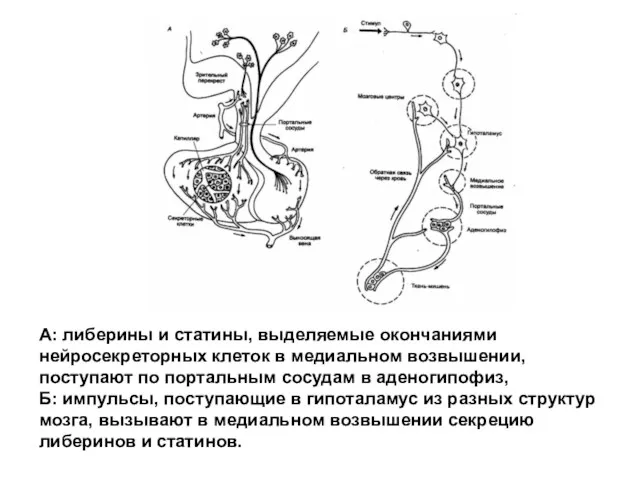
А: либерины и статины, выделяемые окончаниями нейросекреторных клеток в медиальном возвышении,
А: либерины и статины, выделяемые окончаниями нейросекреторных клеток в медиальном возвышении,
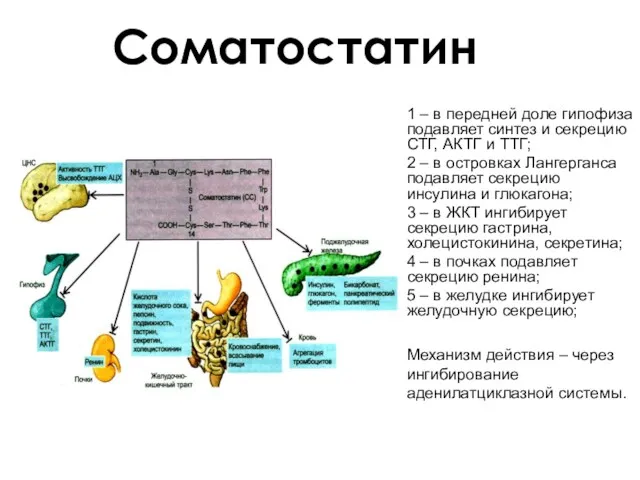
Соматостатин
1 – в передней доле гипофиза подавляет синтез и секрецию СТГ,
Соматостатин
1 – в передней доле гипофиза подавляет синтез и секрецию СТГ,

Соматолиберин
синтезируется нейросекреторными нейронами дугового ядра (n. arcuatus) гипоталамуса и некоторыми опухолями островковых
Соматолиберин
синтезируется нейросекреторными нейронами дугового ядра (n. arcuatus) гипоталамуса и некоторыми опухолями островковых

Гонадолиберин
Мишени гонадолиберина — гонадотрофы передней доли гипофиза.
Функции:
Гонадолиберин — ключевой нейрорегулятор репродуктивной функции,
Гонадолиберин
Мишени гонадолиберина — гонадотрофы передней доли гипофиза.
Функции:
Гонадолиберин — ключевой нейрорегулятор репродуктивной функции,

Пролактиностатин
Мишени пролактиностатина — лактотрофы передней доли гипофиза.
Функции:
Пролактиностатин подавляет секрецию пролактина из
Пролактиностатин
Мишени пролактиностатина — лактотрофы передней доли гипофиза.
Функции:
Пролактиностатин подавляет секрецию пролактина из

Тиреолиберин
синтезируется многими нейронами ЦНС (в т.ч. нейросекреторными нейронами паравентрикулярного ядра).
Мишени —
Тиреолиберин
синтезируется многими нейронами ЦНС (в т.ч. нейросекреторными нейронами паравентрикулярного ядра).
Мишени —
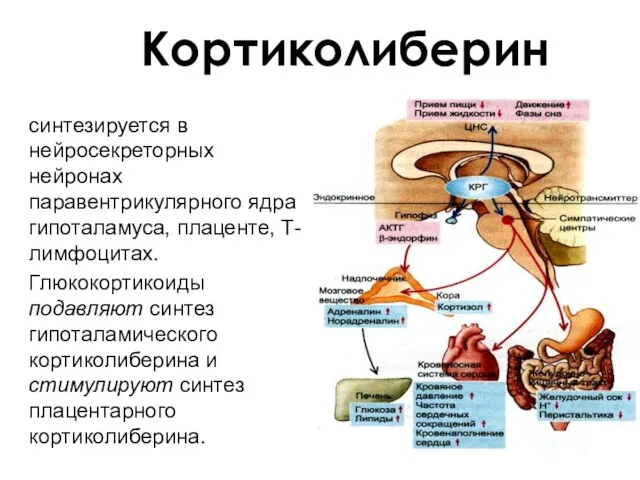
Кортиколиберин
синтезируется в нейросекреторных нейронах паравентрикулярного ядра гипоталамуса, плаценте, Т-лимфоцитах.
Глюкокортикоиды подавляют синтез
Кортиколиберин
синтезируется в нейросекреторных нейронах паравентрикулярного ядра гипоталамуса, плаценте, Т-лимфоцитах.
Глюкокортикоиды подавляют синтез

Кортиколиберин
Функции:
1. Передняя доля гипофиза - стимуляция синтеза и секреции АКТГ
2. Во
Кортиколиберин
Функции:
1. Передняя доля гипофиза - стимуляция синтеза и секреции АКТГ
2. Во

Меланостатин
подавляет образование меланотропинов.
Меланостатин
подавляет образование меланотропинов.

Дофамин
Промежуточный продукт метаболизма тирозина и предшественник норадреналина и адреналина — поступающий
Дофамин
Промежуточный продукт метаболизма тирозина и предшественник норадреналина и адреналина — поступающий
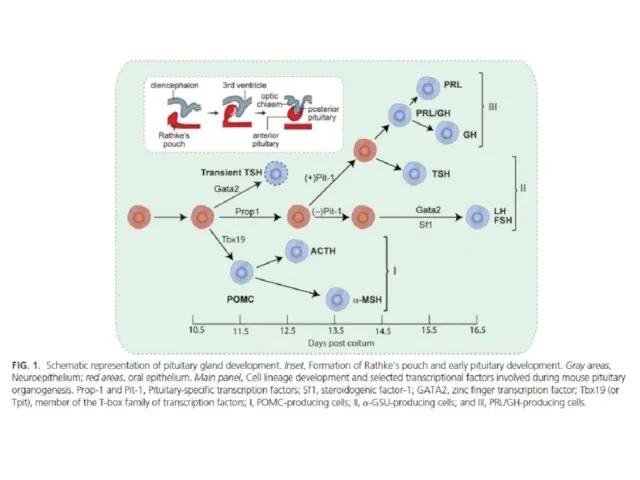

Гормоны задней доли гипофиза
Вазопрессин и окситоцин синтезируются в нейросекреторных нейронах паравентрикулярного
Гормоны задней доли гипофиза
Вазопрессин и окситоцин синтезируются в нейросекреторных нейронах паравентрикулярного
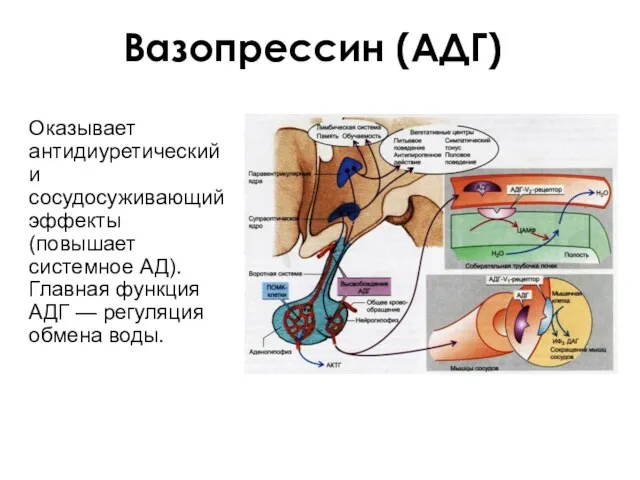
Вазопрессин (АДГ)
Оказывает антидиуретический и сосудосуживающий эффекты (повышает системное АД). Главная функция
Вазопрессин (АДГ)
Оказывает антидиуретический и сосудосуживающий эффекты (повышает системное АД). Главная функция

Регуляция секреции АДГ
Стимулируют секрецию:
1. гиповолемия,
2. гиперосмоляльность,
3. переход в
Регуляция секреции АДГ
Стимулируют секрецию:
1. гиповолемия,
2. гиперосмоляльность,
3. переход в
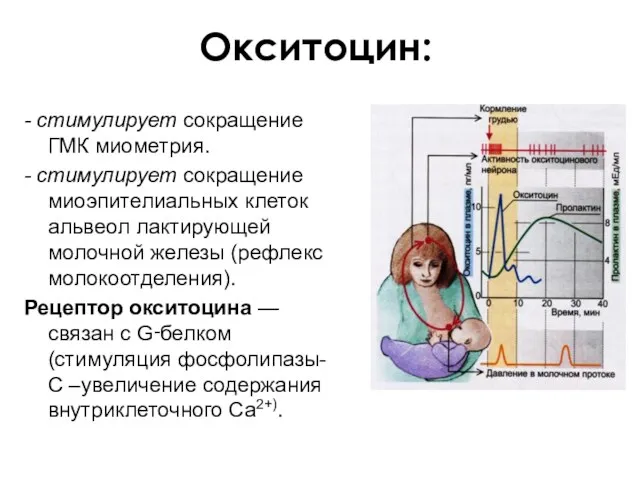
Окситоцин:
- стимулирует сокращение ГМК миометрия.
- стимулирует сокращение миоэпителиальных клеток альвеол лактирующей
Окситоцин:
- стимулирует сокращение ГМК миометрия.
- стимулирует сокращение миоэпителиальных клеток альвеол лактирующей

Гормоны передней доли гипофиза
По химической структуре гормоны аденогипофиза являются либо пептидами,
Гормоны передней доли гипофиза
По химической структуре гормоны аденогипофиза являются либо пептидами,
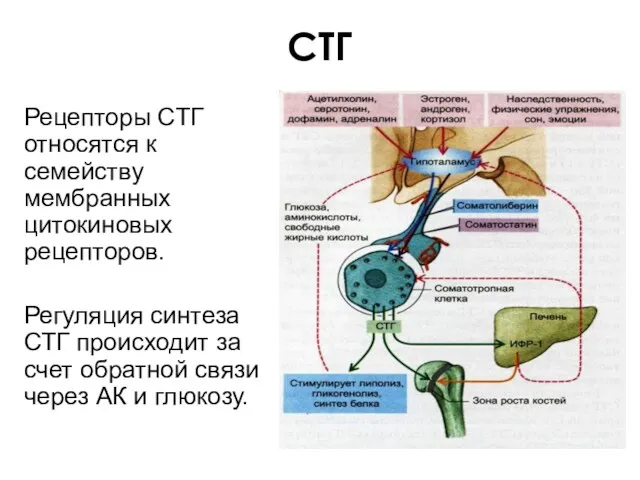
СТГ
Рецепторы СТГ относятся к семейству мембранных цитокиновых рецепторов.
Регуляция синтеза СТГ происходит
СТГ
Рецепторы СТГ относятся к семейству мембранных цитокиновых рецепторов.
Регуляция синтеза СТГ происходит

Метаболические эффекты СТГ
1. Начальная фаза (инсулиноподобный эффект):
А) СТГ увеличивает поглощение
Метаболические эффекты СТГ
1. Начальная фаза (инсулиноподобный эффект):
А) СТГ увеличивает поглощение

Секрецию СТГ
стимулируют:
Уменьшение глюкозы в крови,
Уменьшение свободных ЖК в крови,
Голодание, белковая недостаточность,
Травма,
Секрецию СТГ
стимулируют:
Уменьшение глюкозы в крови,
Уменьшение свободных ЖК в крови,
Голодание, белковая недостаточность,
Травма,

Регуляция секреции АКТГ
Синтез и секрецию АКТГ стимулирует кортиколиберин.
Глюкокортикоиды в высоких дозах -
Регуляция секреции АКТГ
Синтез и секрецию АКТГ стимулирует кортиколиберин.
Глюкокортикоиды в высоких дозах -

АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (преимущественно кортизола).
Гиперсекреция АКТГ
АКТГ стимулирует синтез и секрецию гормонов коры надпочечников (преимущественно кортизола).
Гиперсекреция АКТГ
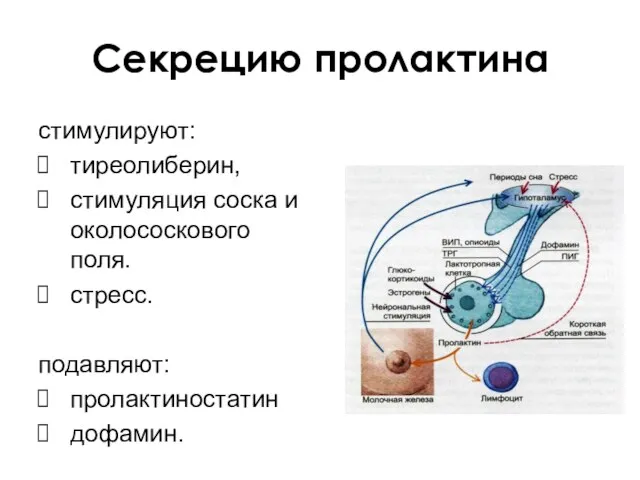
Секрецию пролактина
стимулируют:
тиреолиберин,
стимуляция соска и околососкового поля.
стресс.
подавляют:
пролактиностатин
дофамин.
Секрецию пролактина
стимулируют:
тиреолиберин,
стимуляция соска и околососкового поля.
стресс.
подавляют:
пролактиностатин
дофамин.

Функции пролактина
Главная функция пролактина — стимуляция функции молочной железы.
Рецептор пролактина — мембранный цитокиновый
Функции пролактина
Главная функция пролактина — стимуляция функции молочной железы.
Рецептор пролактина — мембранный цитокиновый

Тиреотропный гормон (ТТГ)
Соматостатин подавляет секрецию ТТГ.
Тиреолиберин стимулирует синтез и секрецию ТТГ.
Гормоны
Тиреотропный гормон (ТТГ)
Соматостатин подавляет секрецию ТТГ.
Тиреолиберин стимулирует синтез и секрецию ТТГ.
Гормоны

Гонадотропные гормоны:
фолликулостимулирующий гормон (ФСГ)
лютеинизирующий гормон (ЛГ),
хорионический гонадотропин (ХГТ) плаценты.
ФСГ:
- у женщин
Гонадотропные гормоны:
фолликулостимулирующий гормон (ФСГ)
лютеинизирующий гормон (ЛГ),
хорионический гонадотропин (ХГТ) плаценты.
ФСГ:
- у женщин

Гонадолиберин стимулирует синтез и секрецию ФСГ и ЛГ.
a- и b-ингибины (фолликулы яичника
Гонадолиберин стимулирует синтез и секрецию ФСГ и ЛГ.
a- и b-ингибины (фолликулы яичника
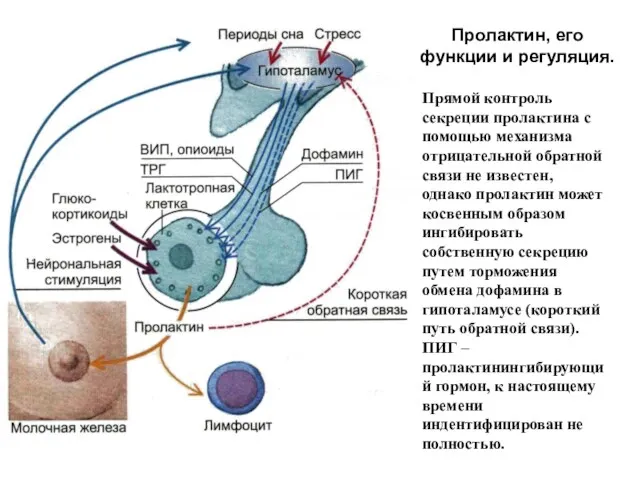
Пролактин, его функции и регуляция.
Прямой контроль секреции пролактина с помощью механизма
Пролактин, его функции и регуляция.
Прямой контроль секреции пролактина с помощью механизма
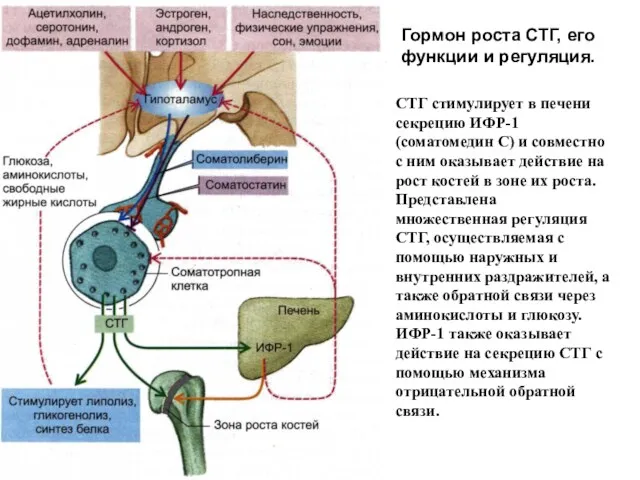
Гормон роста СТГ, его функции и регуляция.
СТГ стимулирует в печени
Гормон роста СТГ, его функции и регуляция.
СТГ стимулирует в печени
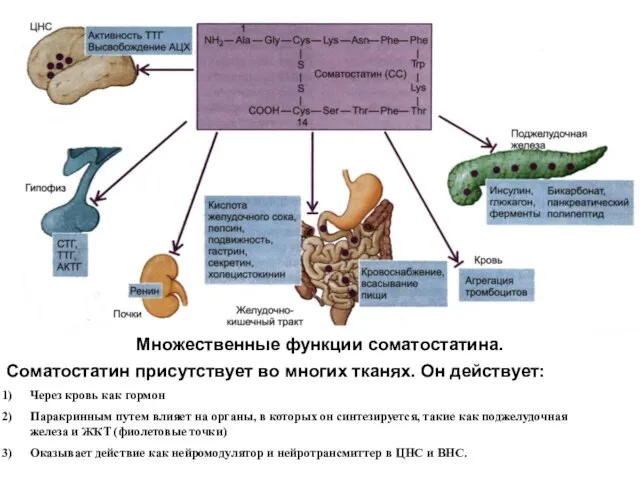
Множественные функции соматостатина.
Соматостатин присутствует во многих тканях. Он действует:
Через кровь
Множественные функции соматостатина.
Соматостатин присутствует во многих тканях. Он действует:
Через кровь
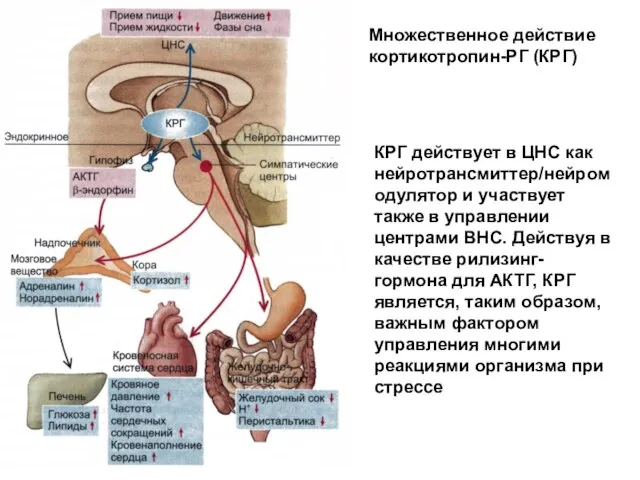
Множественное действие кортикотропин-РГ (КРГ)
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует
Множественное действие кортикотропин-РГ (КРГ)
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует
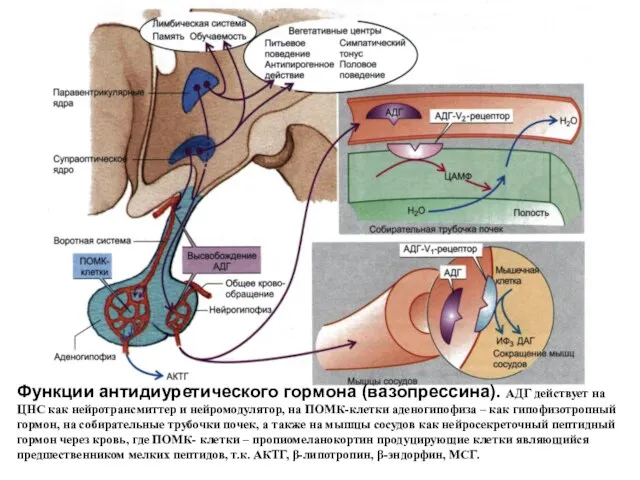
Функции антидиуретического гормона (вазопрессина). АДГ действует на ЦНС как нейротрансмиттер и
Функции антидиуретического гормона (вазопрессина). АДГ действует на ЦНС как нейротрансмиттер и
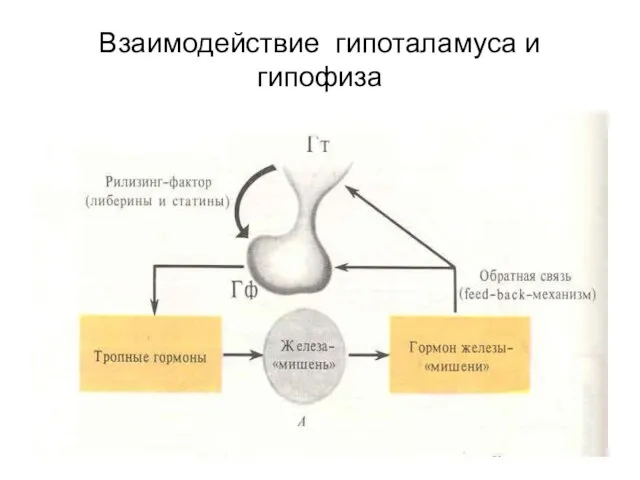
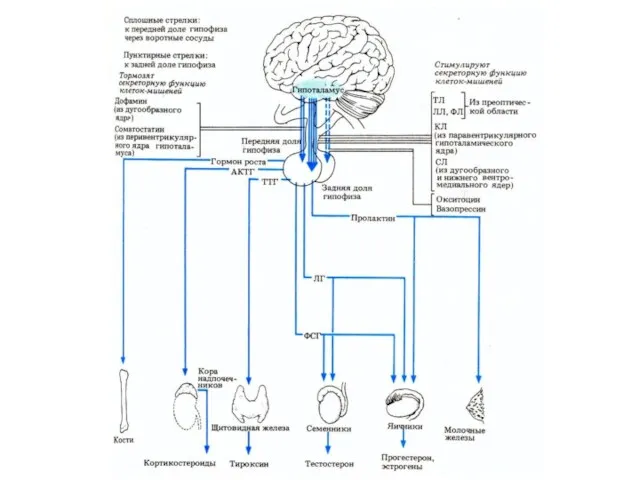
Взаимодействие гипоталамуса и гипофиза
Взаимодействие гипоталамуса и гипофиза
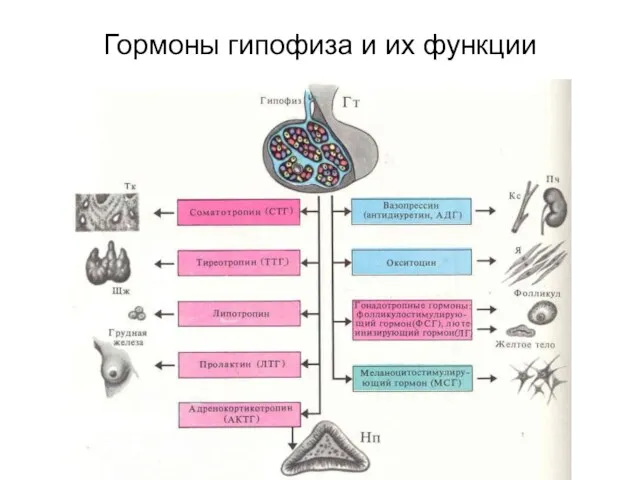
Гормоны гипофиза и их функции
Гормоны гипофиза и их функции
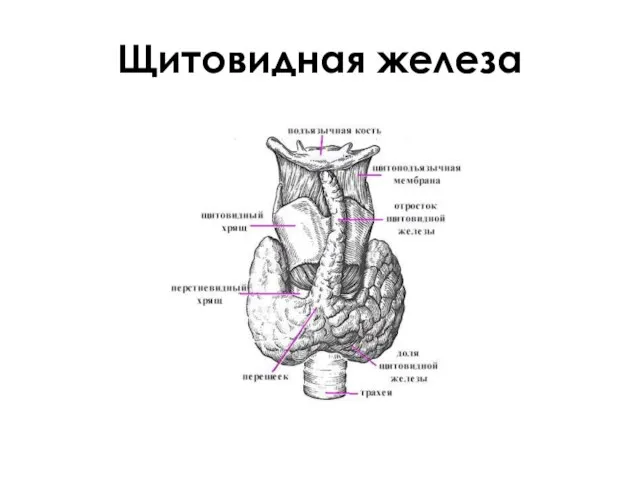
Щитовидная железа
Щитовидная железа
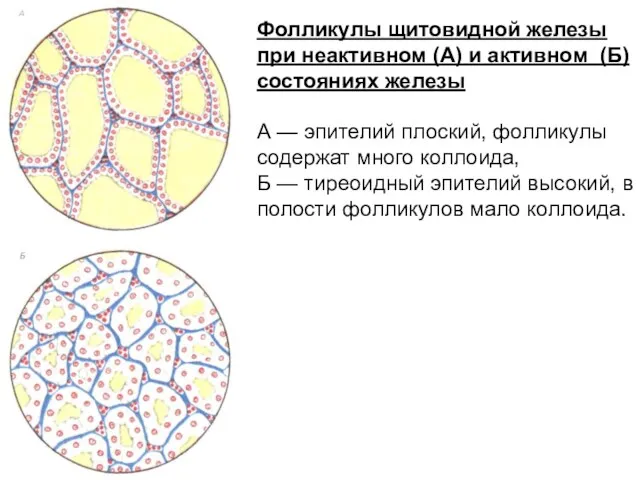
Фолликулы щитовидной железы при неактивном (А) и активном (Б) состояниях железы
А
Фолликулы щитовидной железы при неактивном (А) и активном (Б) состояниях железы
А
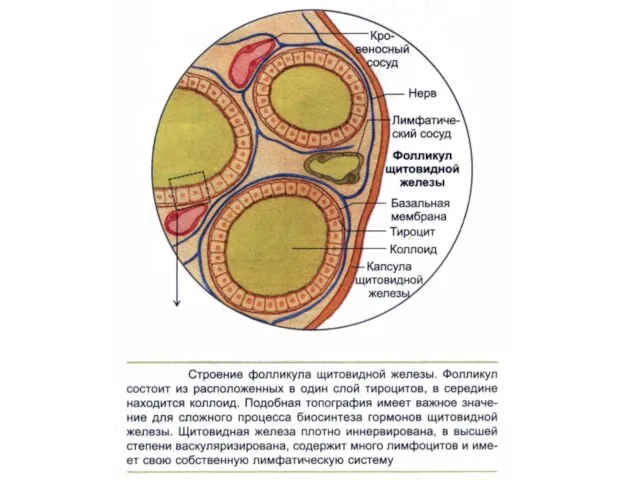

В клетках щитовидной железы происходит синтез двух классов гормонов:
йодсодержащих и кальцитониновых.
Йодсодержащие
В клетках щитовидной железы происходит синтез двух классов гормонов:
йодсодержащих и кальцитониновых.
Йодсодержащие
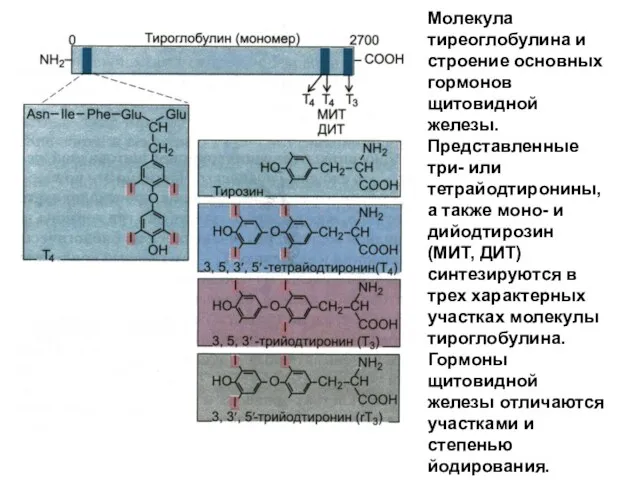
Молекула тиреоглобулина и строение основных гормонов щитовидной железы. Представленные три- или
Молекула тиреоглобулина и строение основных гормонов щитовидной железы. Представленные три- или

Тироксин и трийодтиронин
Йодсодержащие гормоны - тироксин (T4) и трийодтиронин (Т3)
Образование Т3
Тироксин и трийодтиронин
Йодсодержащие гормоны - тироксин (T4) и трийодтиронин (Т3)
Образование Т3

Функции йодсодержащих гормонов
Необходимы для нормального развития ЦНС,
Увеличивают обменные процессы, ускоряют
Функции йодсодержащих гормонов
Необходимы для нормального развития ЦНС,
Увеличивают обменные процессы, ускоряют

Кальцитонин и катакальцин
Их функции антагонистичны эффектам ПТГ:
кальцитонин уменьшает [Са2+]
Кальцитонин и катакальцин
Их функции антагонистичны эффектам ПТГ:
кальцитонин уменьшает [Са2+]

относящиеся к кальцитониновому гену, участвуют в ноцицепции, пищевом поведении, а также
относящиеся к кальцитониновому гену, участвуют в ноцицепции, пищевом поведении, а также

Уровень гормонов щитовидной железы и связывающих белков в плазме и функциональные
Уровень гормонов щитовидной железы и связывающих белков в плазме и функциональные
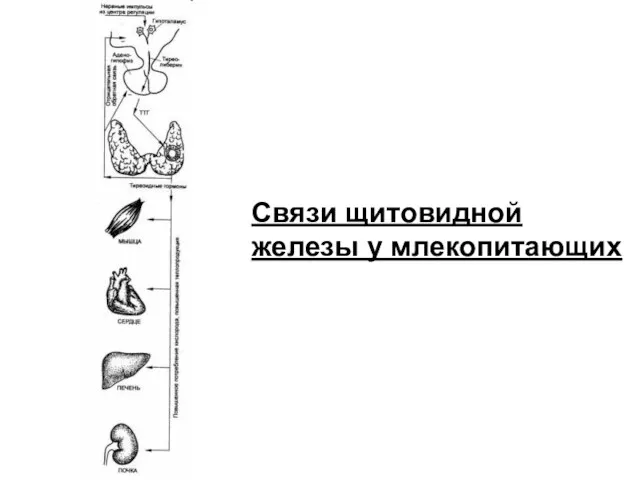
Связи щитовидной железы у млекопитающих
Связи щитовидной железы у млекопитающих
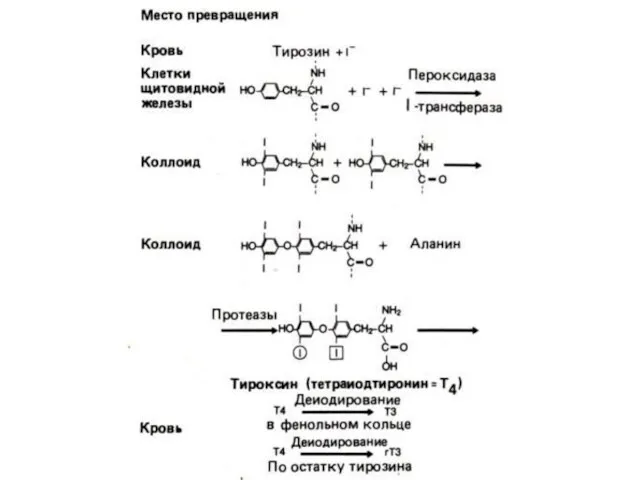
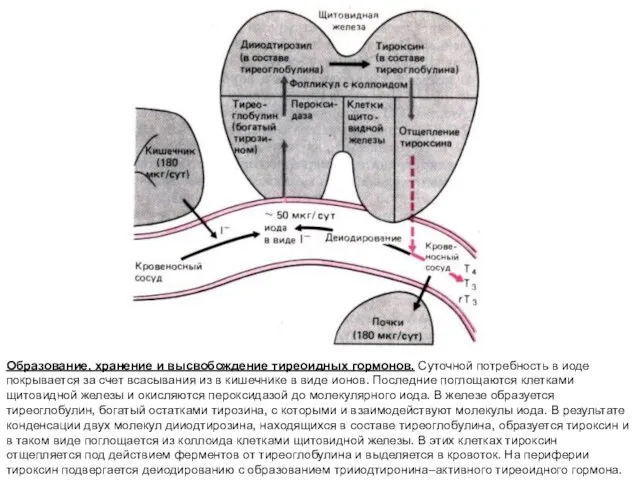
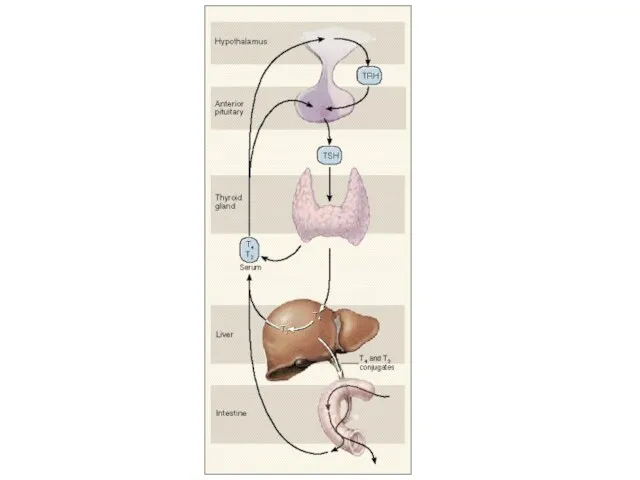
Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иоде покрывается
Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иоде покрывается
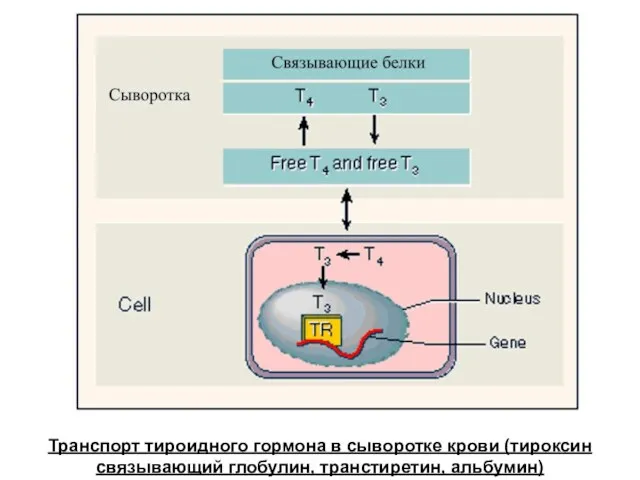
Транспорт тироидного гормона в сыворотке крови (тироксин связывающий глобулин, транстиретин, альбумин)
Транспорт тироидного гормона в сыворотке крови (тироксин связывающий глобулин, транстиретин, альбумин)
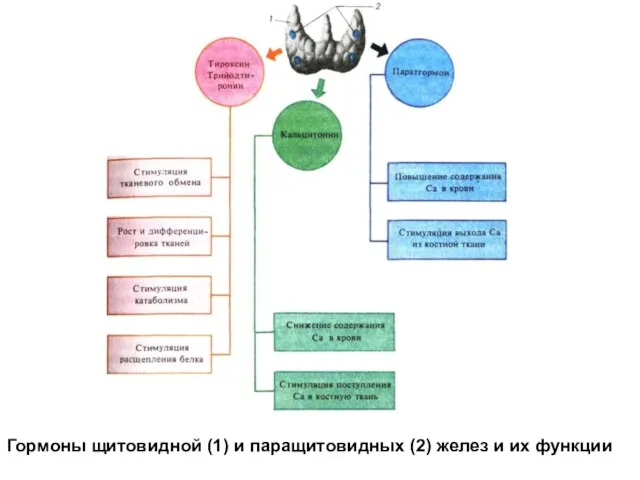
Гормоны щитовидной (1) и паращитовидных (2) желез и их функции
Гормоны щитовидной (1) и паращитовидных (2) желез и их функции

Cуточная потребность в йоде покрывается за счет всасывания в кишечнике в
Cуточная потребность в йоде покрывается за счет всасывания в кишечнике в
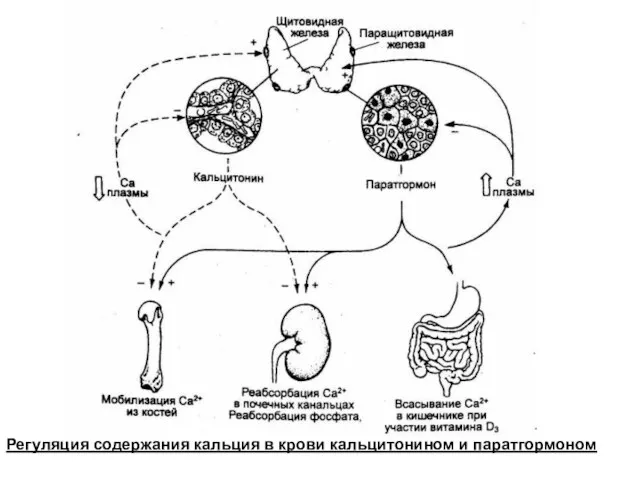
Регуляция содержания кальция в крови кальцитонином и паратгормоном
Регуляция содержания кальция в крови кальцитонином и паратгормоном
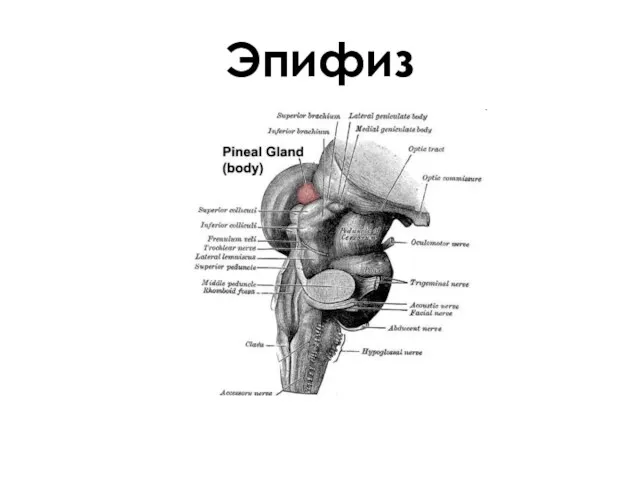
Эпифиз
Эпифиз

Эпифиз
Гормоны эпифиза представлены в основном индоламинами (серотонин и мелатонин), пептидами и
Эпифиз
Гормоны эпифиза представлены в основном индоламинами (серотонин и мелатонин), пептидами и
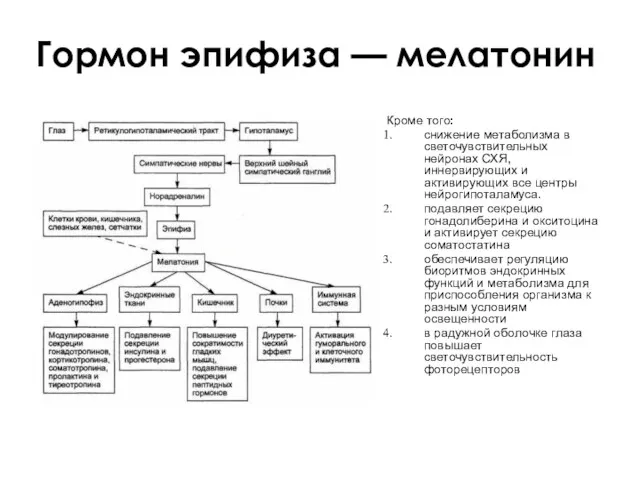
Гормон эпифиза — мелатонин
Кроме того:
снижение метаболизма в светочувствительных нейронах СХЯ, иннервирующих
Гормон эпифиза — мелатонин
Кроме того:
снижение метаболизма в светочувствительных нейронах СХЯ, иннервирующих
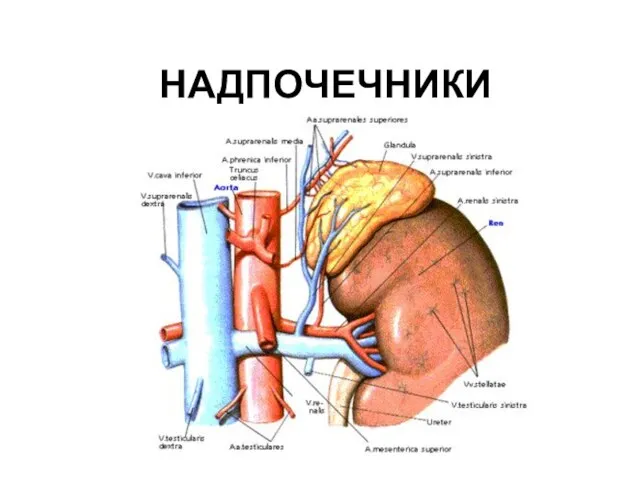
НАДПОЧЕЧНИКИ
НАДПОЧЕЧНИКИ
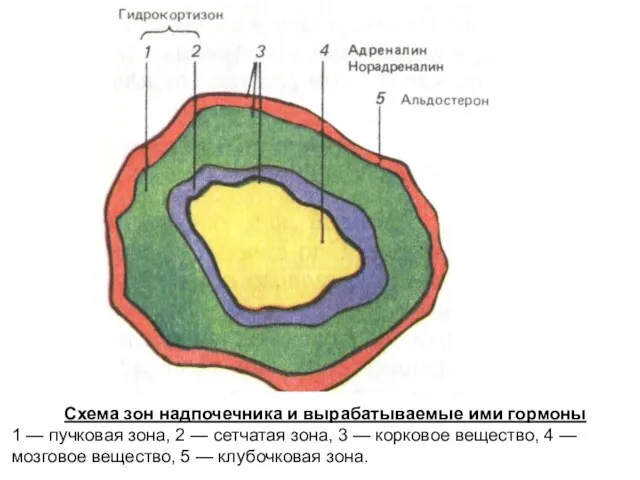
Схема зон надпочечника и вырабатываемые ими гормоны
1 — пучковая зона, 2
Схема зон надпочечника и вырабатываемые ими гормоны
1 — пучковая зона, 2

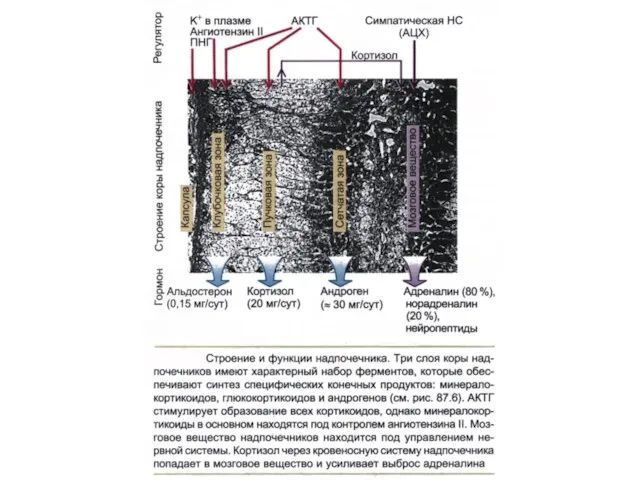
Строение и функции надпочечника
Три слоя надпочечников имеют характерный набор ферментов обеспечивающий
Строение и функции надпочечника
Три слоя надпочечников имеют характерный набор ферментов обеспечивающий
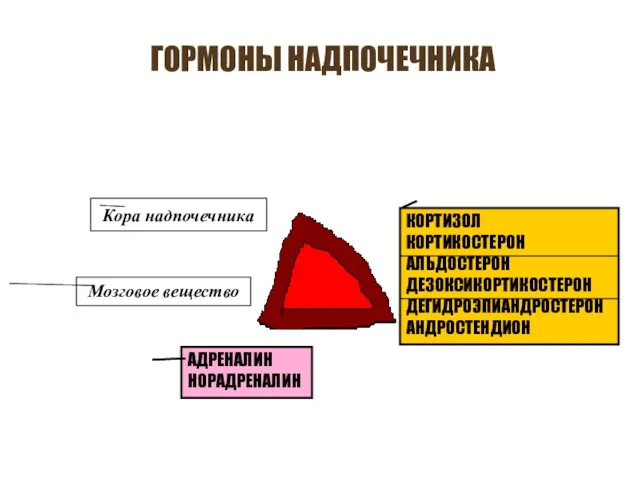
ГОРМОНЫ НАДПОЧЕЧНИКА
Кора надпочечника
Мозговое вещество
КОРТИЗОЛ
КОРТИКОСТЕРОН
АЛЬДОСТЕРОН
ДЕЗОКСИКОРТИКОСТЕРОН
ДЕГИДРОЭПИАНДРОСТЕРОН
АНДРОСТЕНДИОН
АДРЕНАЛИН
НОРАДРЕНАЛИН
ГОРМОНЫ НАДПОЧЕЧНИКА
Кора надпочечника
Мозговое вещество
КОРТИЗОЛ
КОРТИКОСТЕРОН
АЛЬДОСТЕРОН
ДЕЗОКСИКОРТИКОСТЕРОН
ДЕГИДРОЭПИАНДРОСТЕРОН
АНДРОСТЕНДИОН
АДРЕНАЛИН
НОРАДРЕНАЛИН
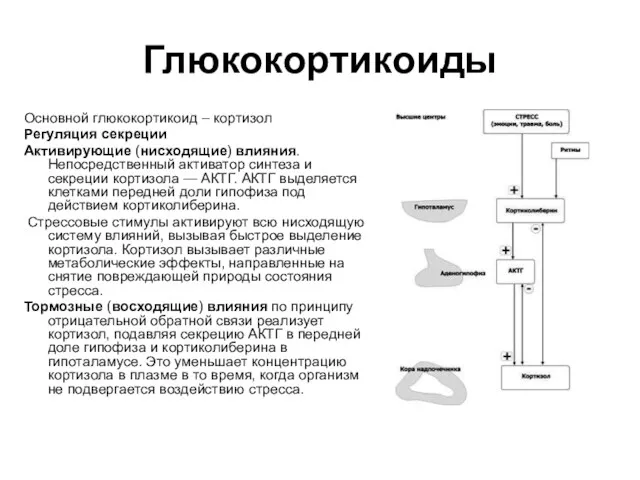
Глюкокортикоиды
Основной глюкокортикоид – кортизол
Регуляция секреции
Активирующие (нисходящие) влияния. Непосредственный активатор синтеза и
Глюкокортикоиды
Основной глюкокортикоид – кортизол
Регуляция секреции
Активирующие (нисходящие) влияния. Непосредственный активатор синтеза и

Углеводный обмен.
Стимуляция глюконеогенеза и уменьшение потребления глюкозы внутренними органами (кроме
Углеводный обмен.
Стимуляция глюконеогенеза и уменьшение потребления глюкозы внутренними органами (кроме

Липидный обмен.
Липолиз усиливается в конечностях.
Липогенез усиливается в других частях тела
Липидный обмен.
Липолиз усиливается в конечностях.
Липогенез усиливается в других частях тела

- В высоких дозах выступают как иммунодепрессанты (применяют для предупреждения отторжения
- В высоких дозах выступают как иммунодепрессанты (применяют для предупреждения отторжения

Основной минералокортикоид - альдостерон
Регуляция секреции альдостерона :
Ангиотензин II — главный регулятор, стимулирует выброс
Основной минералокортикоид - альдостерон
Регуляция секреции альдостерона :
Ангиотензин II — главный регулятор, стимулирует выброс

Функции альдостерона:
- увеличивает реабсорбцию Na+. Задержка Na+ приводит к увеличению содержания
Функции альдостерона:
- увеличивает реабсорбцию Na+. Задержка Na+ приводит к увеличению содержания
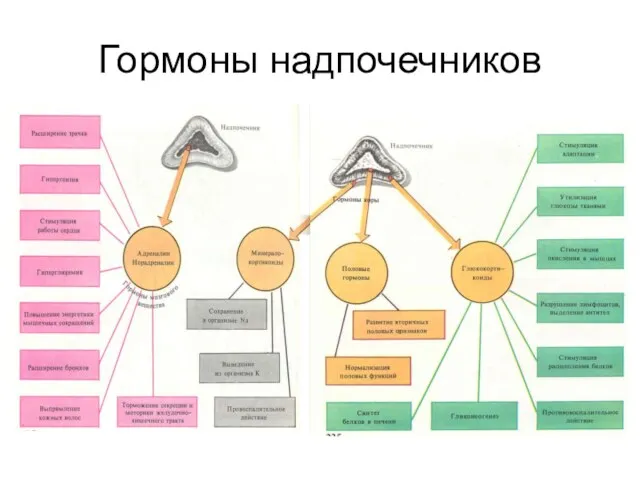
Гормоны надпочечников
Гормоны надпочечников
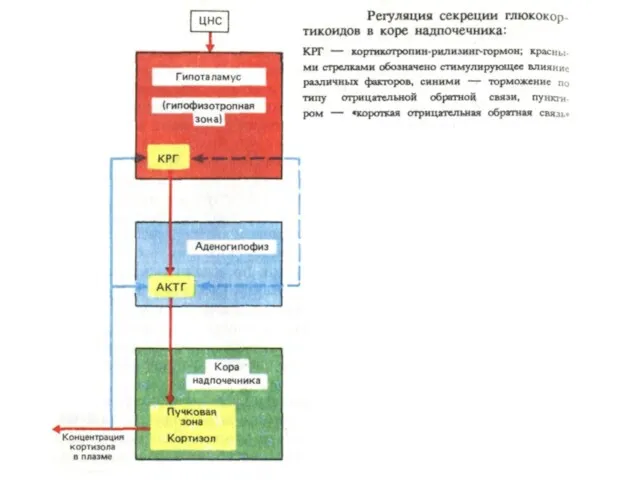
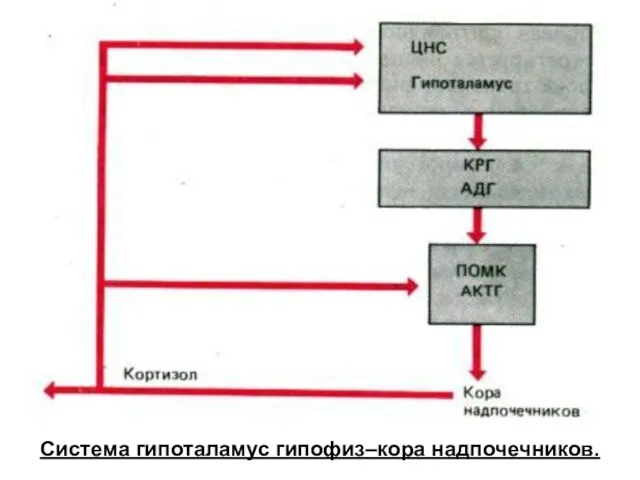
Система гипоталамус гипофиз–кора надпочечников.
Система гипоталамус гипофиз–кора надпочечников.
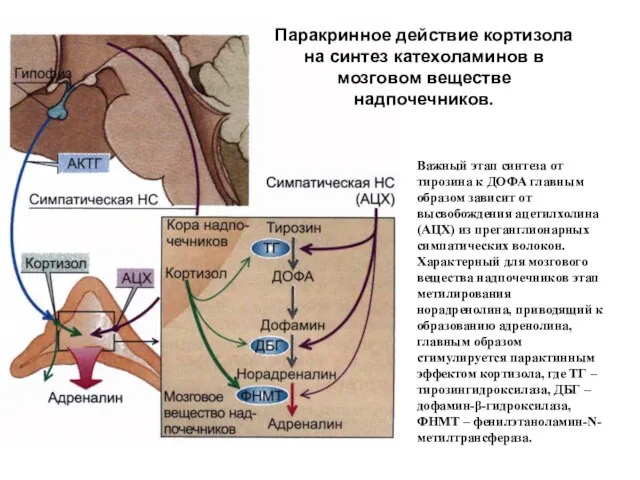
Паракринное действие кортизола на синтез катехоламинов в мозговом веществе надпочечников.
Важный этап
Паракринное действие кортизола на синтез катехоламинов в мозговом веществе надпочечников.
Важный этап
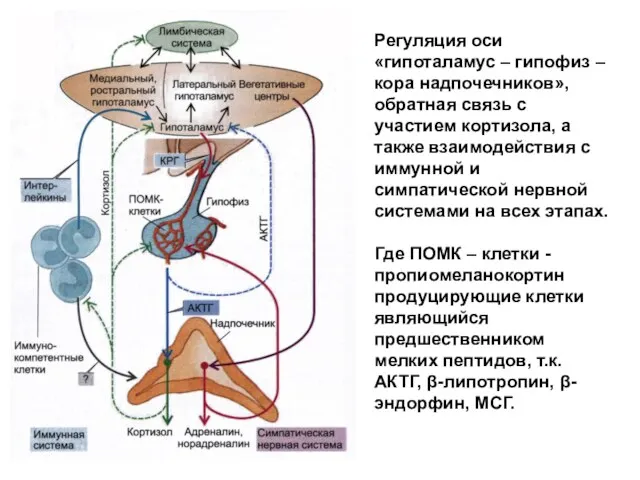
Регуляция оси «гипоталамус – гипофиз – кора надпочечников», обратная связь с
Регуляция оси «гипоталамус – гипофиз – кора надпочечников», обратная связь с
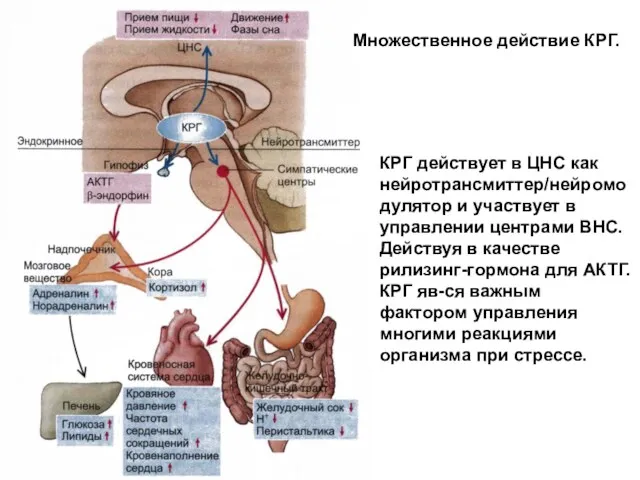
Множественное действие КРГ.
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует
Множественное действие КРГ.
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует

МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА
ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ
ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ
КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ
МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА
ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ
ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ
КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ
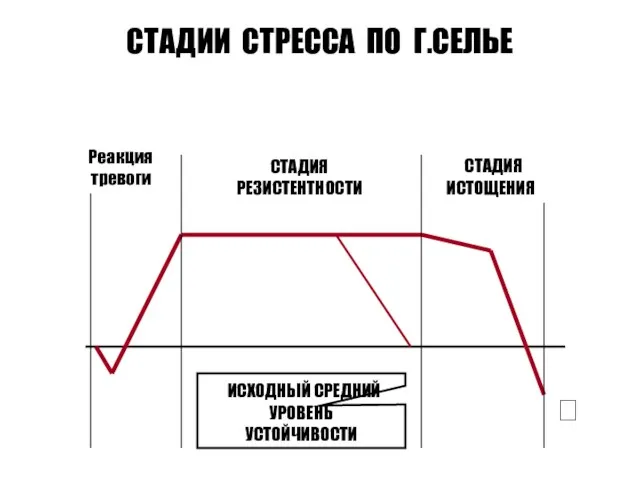
СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ
Реакция
тревоги
СТАДИЯ
РЕЗИСТЕНТНОСТИ
СТАДИЯ
ИСТОЩЕНИЯ
ИСХОДНЫЙ СРЕДНИЙ
УРОВЕНЬ
УСТОЙЧИВОСТИ
СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ
Реакция
тревоги
СТАДИЯ
РЕЗИСТЕНТНОСТИ
СТАДИЯ
ИСТОЩЕНИЯ
ИСХОДНЫЙ СРЕДНИЙ
УРОВЕНЬ
УСТОЙЧИВОСТИ
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА

В островках Лангерганса идентифицировано несколько типов эндокринных клеток, синтезирующих и секретирующих
В островках Лангерганса идентифицировано несколько типов эндокринных клеток, синтезирующих и секретирующих

Действие инсулина на углеводный обмен
активация утилизации глюкозы клетками,
усиление фосфорилирования;
подавление распада гликогена;
стимуляция
Действие инсулина на углеводный обмен
активация утилизации глюкозы клетками,
усиление фосфорилирования;
подавление распада гликогена;
стимуляция

Действие инсулина на белковый обмен
повышение проницаемости мембран для аминоокислот;
усиление синтеза
Действие инсулина на белковый обмен
повышение проницаемости мембран для аминоокислот;
усиление синтеза

Действие инсулина на липидный обмен
стимуляция синтеза свободных жирных кислот из глюкозы;
Действие инсулина на липидный обмен
стимуляция синтеза свободных жирных кислот из глюкозы;

Скорость проявления эффектов инсулина
Физиологические эффекты инсулина по скорости их наступления подразделяют
Скорость проявления эффектов инсулина
Физиологические эффекты инсулина по скорости их наступления подразделяют

Регуляция
Глюкоза - ведущий регулятор секреции инсулина: при участии Са2+ стимулирует секрецию
Регуляция
Глюкоза - ведущий регулятор секреции инсулина: при участии Са2+ стимулирует секрецию

Гомеостаз глюкозы
поддерживают следующие механизмы:
Печень демпфирует колебания концентрации глюкозы за счет обратимой
Гомеостаз глюкозы
поддерживают следующие механизмы:
Печень демпфирует колебания концентрации глюкозы за счет обратимой
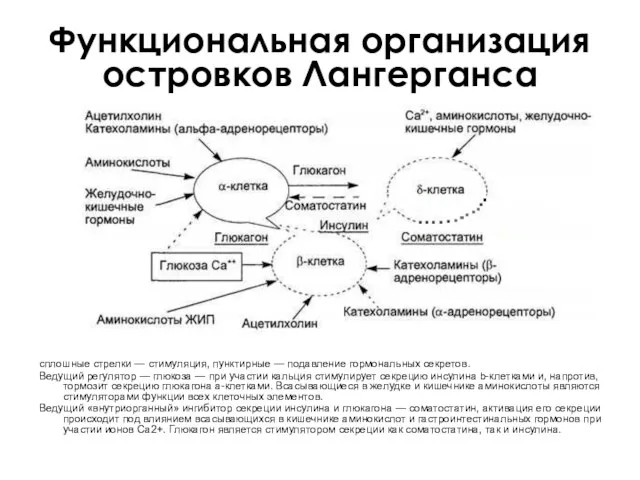
Функциональная организация островков Лангерганса
сплошные стрелки — стимуляция, пунктирные — подавление гормональных
Функциональная организация островков Лангерганса
сплошные стрелки — стимуляция, пунктирные — подавление гормональных

Сахарный диабет типа I
развивается в результате аутоиммунной деструкции b-клеток островков Лангерганса
Сахарный диабет типа I
развивается в результате аутоиммунной деструкции b-клеток островков Лангерганса

Сахарный диабет типа II
развивается в результате :
либо нарушения секреции инсулина (избыточное
Сахарный диабет типа II
развивается в результате :
либо нарушения секреции инсулина (избыточное
Тимус
Тимус (вилочковая железа) является центральным органом иммунитета, обеспечивающим продукцию специфических Т-лимфоцитов.
Тимус
Тимус (вилочковая железа) является центральным органом иммунитета, обеспечивающим продукцию специфических Т-лимфоцитов.

Яички
В яичках синтезируются стероидные андрогены и a‑ингибин.
Тестостерон — основной циркулирующий андроген.
Яички
В яичках синтезируются стероидные андрогены и a‑ингибин.
Тестостерон — основной циркулирующий андроген.

Яичники
В яичниках синтезируются стероидные женские половые гормоны, гликопротеиновые гормоны ингибины и
Яичники
В яичниках синтезируются стероидные женские половые гормоны, гликопротеиновые гормоны ингибины и

Гормоны, вырабатываемые в плаценте
Плацента тесно анатомически и функционально связана с организмами
Гормоны, вырабатываемые в плаценте
Плацента тесно анатомически и функционально связана с организмами

Гормоны, вырабатываемые в почке
Ренин не является гормоном, этот фермент — начальное
Гормоны, вырабатываемые в почке
Ренин не является гормоном, этот фермент — начальное

Гормоны, вырабатываемые в сердце
Натрийуретический фактор синтезируют кардиомиоциты правого предсердия и некоторые
Гормоны, вырабатываемые в сердце
Натрийуретический фактор синтезируют кардиомиоциты правого предсердия и некоторые

Гормоны, вырабатываемые в ЖКТ
В стенке трубчатых органов ЖКТ присутствует огромное количество
Гормоны, вырабатываемые в ЖКТ
В стенке трубчатых органов ЖКТ присутствует огромное количество

АПУД-система
APUD — аббревиатура, образованная из первых букв англ. слов amines - амины,
АПУД-система
APUD — аббревиатура, образованная из первых букв англ. слов amines - амины,

АПУД-система
Предполагают, что единым эмбриональным предшественником клеток АПУД-системы является так называемый нейроэндокринно-программированный
АПУД-система
Предполагают, что единым эмбриональным предшественником клеток АПУД-системы является так называемый нейроэндокринно-программированный
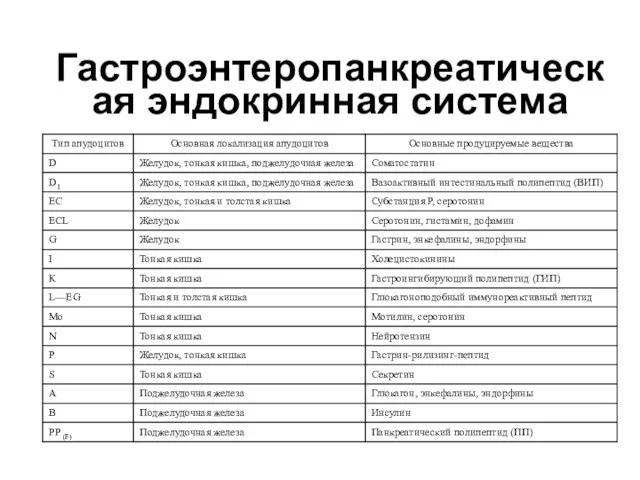
Гастроэнтеропанкреатическая эндокринная система
Гастроэнтеропанкреатическая эндокринная система
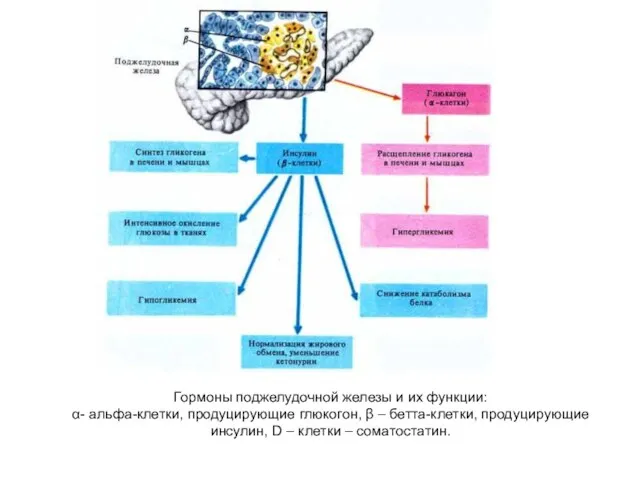
Гормоны поджелудочной железы и их функции:
α- альфа-клетки, продуцирующие глюкогон, β –
Гормоны поджелудочной железы и их функции: α- альфа-клетки, продуцирующие глюкогон, β –
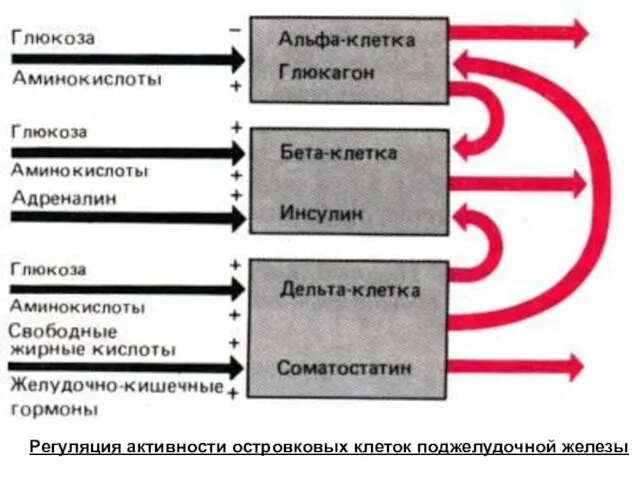
Регуляция активности островковых клеток поджелудочной железы
Регуляция активности островковых клеток поджелудочной железы
 Многообразие земноводных
Многообразие земноводных Способы классификации. Лекция 01
Способы классификации. Лекция 01 Будова і функції шкіри
Будова і функції шкіри Эмбриональное развитие человеческого организма
Эмбриональное развитие человеческого организма § 18 Водоросли
§ 18 Водоросли Неживая природа осенью (2 класс)
Неживая природа осенью (2 класс) Классификация микроорганизмов
Классификация микроорганизмов Тигрова Акула
Тигрова Акула презентация Значение медоносной пчелы в жизни человека
презентация Значение медоносной пчелы в жизни человека Ендокринна регуляція функцій організму людини
Ендокринна регуляція функцій організму людини Отряды насекомых с неполным превращением. Отряд Стрекозы
Отряды насекомых с неполным превращением. Отряд Стрекозы Основы разведения животных
Основы разведения животных Презентация 6 класс
Презентация 6 класс Гены, обусловливающие темно-синий окрас глаз
Гены, обусловливающие темно-синий окрас глаз Биогеоценоз и экосистема
Биогеоценоз и экосистема Четвероногие друзья
Четвероногие друзья Получение ранней продукции кабачков при выращивании посевом семян в открытый грунт и рассадным способом
Получение ранней продукции кабачков при выращивании посевом семян в открытый грунт и рассадным способом Загрязнение атмосферы автономным транспортом
Загрязнение атмосферы автономным транспортом Попугаи. Виды
Попугаи. Виды Нервная ткань. Органы нервной системы. Лекция №6
Нервная ткань. Органы нервной системы. Лекция №6 Обобщающий урок в форме конференции по итогам изучения раздела Нервная система и органы чувств
Обобщающий урок в форме конференции по итогам изучения раздела Нервная система и органы чувств Мышцы конечностей
Мышцы конечностей Механизм мышечного сокращения
Механизм мышечного сокращения Опорно-двигательная система (животные)
Опорно-двигательная система (животные) Строение и значение нервной системы
Строение и значение нервной системы Сообщества. Экосистемы
Сообщества. Экосистемы Бактериялардың өсу және көбеюі
Бактериялардың өсу және көбеюі Гименолепидоз. Систематическое положение
Гименолепидоз. Систематическое положение