- Функциональная морфология синтетического аппарата клетки

Содержание
- 2. Синтетический аппарат клетки Представлен органеллами, синтезирующими различные вещества, которые используются самой клеткой или идут на нужды
- 3. Рибосомы Немембранные, электронноплотные, округлые (диаметром 15 -30 нм) органеллы, обеспечивающие создание первичной структуры белка путем образования
- 4. Рибосомы Каждая рибосома состоит из двух (большей и меньшей) субъединиц, которые диссоциированно располагаются в цитоплазме и
- 5. 1. Синтез мРНК рибосомных белков РНК полимеразой II. 2. Экспорт мРНК из ядра. 3. Узнавание мРНК
- 6. Виды рибосом Морфологически (по размерам, индексу седиментации и молекулярной массе) различают рибосомы прокариот, цитоплазматические рибосомы эукариот
- 7. Строение рибосомы
- 8. Строение рибосомы Схема РНК-связывающих участков рибосомы. Буквами обозначены участки связывания тРНК. А — аминоацил-тРНК-связывающий участок, Р
- 9. Виды РНК матричные РНК являются матрицей для синтеза белка в рибосомах; транспортные РНК доставляют аминокислоты к
- 10. Нуклеотиды — фосфатные эфиры нуклеозидов. Нуклеозиды — N–гликозильные производные (N–гликозиды) разных азотистых оснований (пурины, пиримидины), содержащих
- 11. Биосинтез белка (схематично) Биосинтез белка осуществляется поэтапно в ядре и цитоплазме. В ядре происходит: 1. Образование
- 12. Биосинтез белка (схематично) В цитоплазме идет трансляция (синтез) на свободных полисомах или грЭПС белковых молекул по
- 13. Биосинтез белка (схематично) Во время инициации последовательно происходит образование комплекса полной рибосомы – иРНК и присоединение
- 14. Общая схема трансляции. Инициация. 1. Узнавание стартового кодона (AUG), сопровождается присоединением тРНК аминоацилированной метионином (М) и
- 15. Эндоплазматическая сеть (ЭПС) Это замкнутая, единая система цистерн, трубочек, уплощенных или расширенных канальцев, окруженных биологической мембраной.
- 16. Эндоплазматическая сеть (ЭПС) Выделяют: гранулярную или шероховатую ЭПС (грЭПС, шЭПС), агранулярную или гладкую (аЭПС, глЭПС)
- 17. Гранулярная ЭПС Гранулярная ЭПС обеспечивает синтез всех мембранных белков, в том числе циторецепторов (за исключением некоторых
- 18. Сигнальная гипотеза поступления секреторных, мембранных и лизосомных белков в гранулярную эндоплазматическую сеть.
- 19. Функции грЭПС Обеспечивает синтез белков, липидов и углеводов, а также начальное посттрансляционное преобразование белков (их дополнительное
- 20. Агранулярная ЭПС Это участки эндоплазматической сети, на которых отсутствуют рибосомы и осуществляет: синтез липидов (в т.ч.
- 21. Пластинчатый комплекс Гольджи (ПКГ) в световом микроскопе имеет вид сети или отдельных диктиосом, расположенных возле ядра
- 22. Пластинчатый комплекс Гольджи (ПКГ) Под электронным микроскопом представляет собой мембранную органеллу, состоящую из: а) стопки уплощенных
- 23. Пластинчатый комплекс Гольджи (ПКГ) Пространственно ПКГ представляет поляризованную структуру, имеющую две морфологические и функционально различные поверхности:
- 24. Пластинчатый комплекс Гольджи (ПКГ) Цистерны имеют вид изогнутых дисков, диаметром 0,5 – 5 мкм, объединяясь по
- 26. Функции ПКГ Доработка (процессинг) продуктов, поступивших из ЭПС (включение углеводных или липидных компонентов в белковые молекулы,
- 28. Скачать презентацию

Синтетический аппарат клетки
Представлен органеллами, синтезирующими различные вещества, которые используются самой
Синтетический аппарат клетки
Представлен органеллами, синтезирующими различные вещества, которые используются самой

Рибосомы
Немембранные, электронноплотные, округлые (диаметром 15 -30 нм) органеллы, обеспечивающие создание
Рибосомы
Немембранные, электронноплотные, округлые (диаметром 15 -30 нм) органеллы, обеспечивающие создание

Рибосомы
Каждая рибосома состоит из двух (большей и меньшей) субъединиц, которые диссоциированно
Рибосомы
Каждая рибосома состоит из двух (большей и меньшей) субъединиц, которые диссоциированно
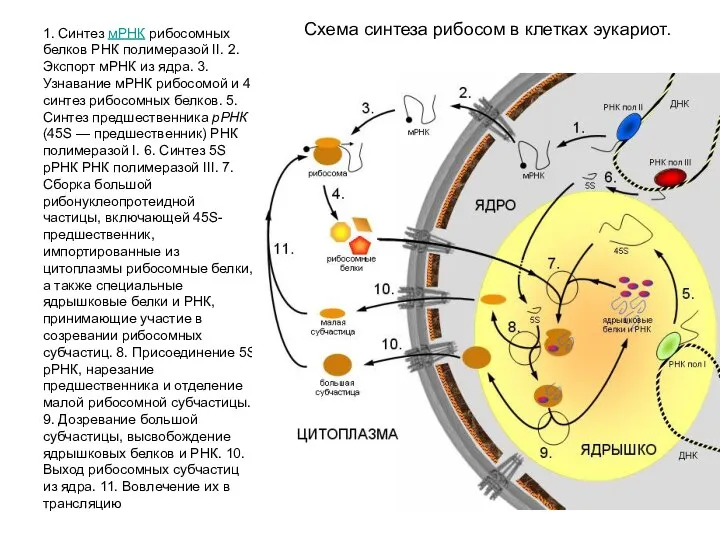
1. Синтез мРНК рибосомных белков РНК полимеразой II. 2. Экспорт мРНК
1. Синтез мРНК рибосомных белков РНК полимеразой II. 2. Экспорт мРНК
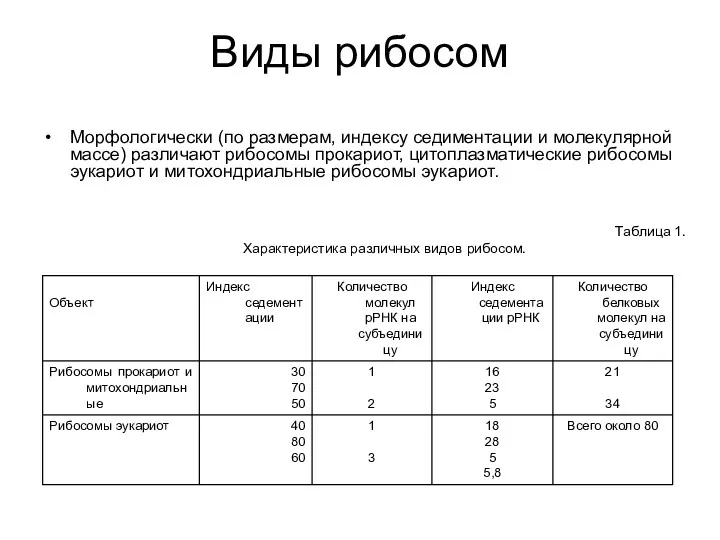
Виды рибосом
Морфологически (по размерам, индексу седиментации и молекулярной массе) различают рибосомы
Виды рибосом
Морфологически (по размерам, индексу седиментации и молекулярной массе) различают рибосомы
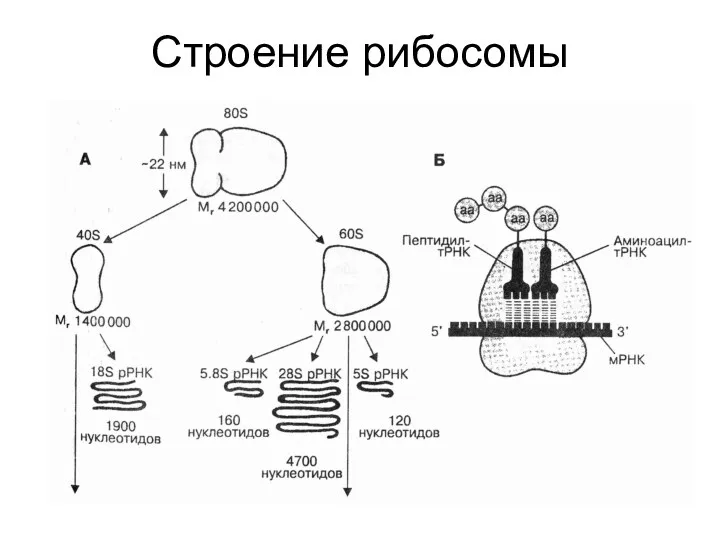
Строение рибосомы
Строение рибосомы

Строение
рибосомы
Схема РНК-связывающих участков рибосомы. Буквами обозначены участки связывания тРНК. А —
Строение
рибосомы
Схема РНК-связывающих участков рибосомы. Буквами обозначены участки связывания тРНК. А —

Виды РНК
матричные РНК являются матрицей для синтеза белка в рибосомах;
транспортные
Виды РНК
матричные РНК являются матрицей для синтеза белка в рибосомах;
транспортные

Нуклеотиды — фосфатные эфиры нуклеозидов. Нуклеозиды — N–гликозильные производные (N–гликозиды) разных азотистых оснований
Нуклеотиды — фосфатные эфиры нуклеозидов. Нуклеозиды — N–гликозильные производные (N–гликозиды) разных азотистых оснований
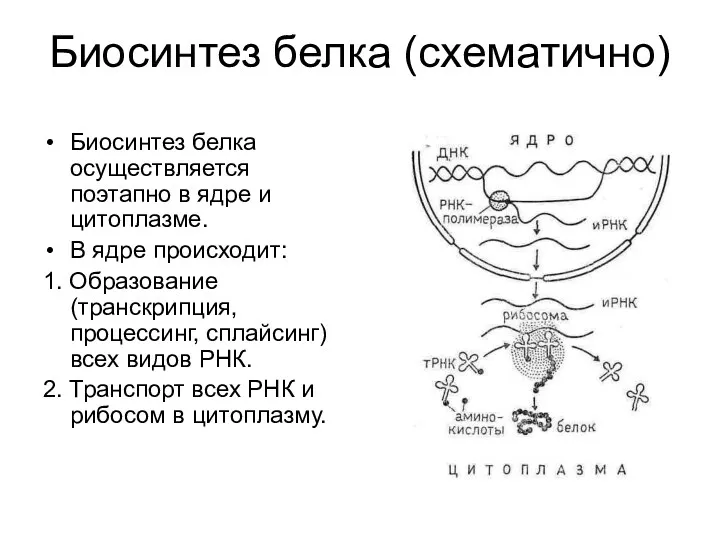
Биосинтез белка (схематично)
Биосинтез белка осуществляется поэтапно в ядре и цитоплазме.
В ядре
Биосинтез белка (схематично)
Биосинтез белка осуществляется поэтапно в ядре и цитоплазме.
В ядре

Биосинтез белка (схематично)
В цитоплазме идет трансляция (синтез) на свободных полисомах или
Биосинтез белка (схематично)
В цитоплазме идет трансляция (синтез) на свободных полисомах или

Биосинтез белка (схематично)
Во время инициации последовательно происходит образование комплекса полной рибосомы
Биосинтез белка (схематично)
Во время инициации последовательно происходит образование комплекса полной рибосомы
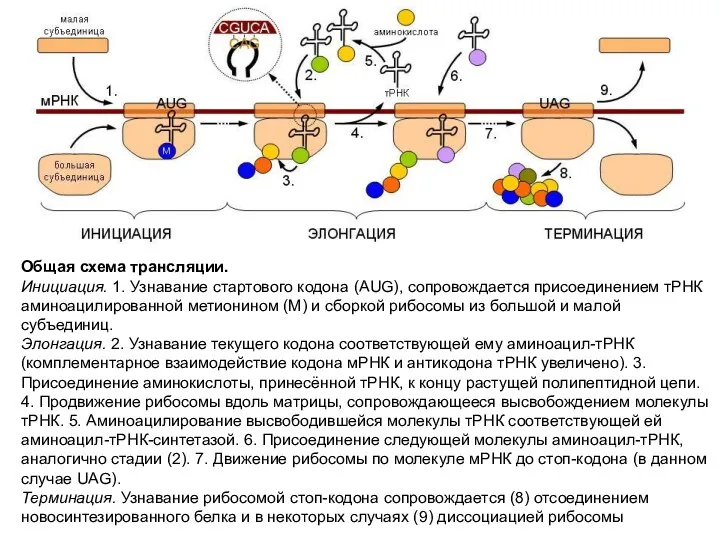
Общая схема трансляции.
Инициация. 1. Узнавание стартового кодона (AUG), сопровождается присоединением тРНК
Общая схема трансляции. Инициация. 1. Узнавание стартового кодона (AUG), сопровождается присоединением тРНК
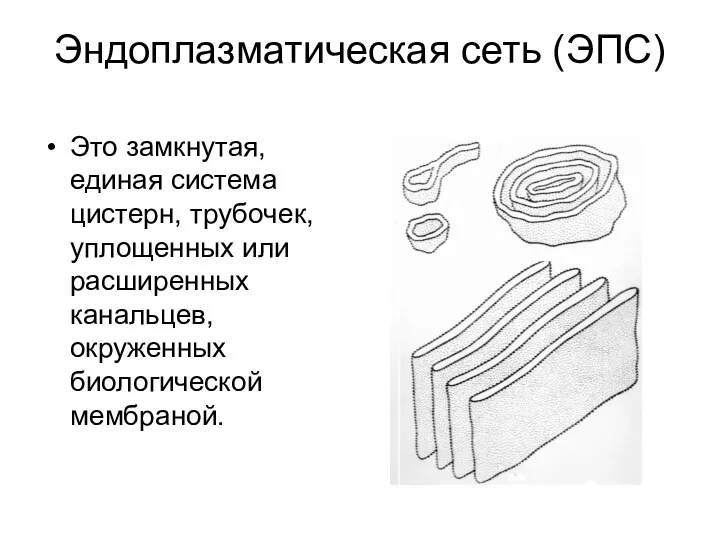
Эндоплазматическая сеть (ЭПС)
Это замкнутая, единая система цистерн, трубочек, уплощенных или расширенных
Эндоплазматическая сеть (ЭПС)
Это замкнутая, единая система цистерн, трубочек, уплощенных или расширенных

Эндоплазматическая сеть (ЭПС)
Выделяют:
гранулярную или шероховатую ЭПС (грЭПС, шЭПС),
агранулярную или гладкую
Эндоплазматическая сеть (ЭПС)
Выделяют:
гранулярную или шероховатую ЭПС (грЭПС, шЭПС),
агранулярную или гладкую
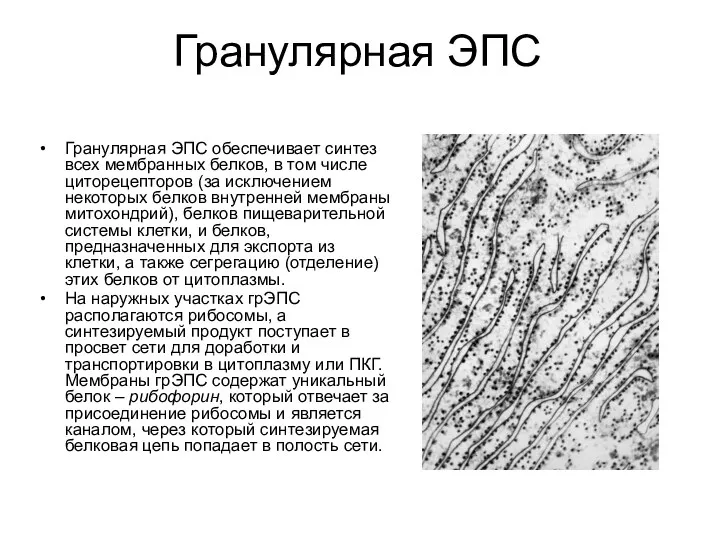
Гранулярная ЭПС
Гранулярная ЭПС обеспечивает синтез всех мембранных белков, в том числе
Гранулярная ЭПС
Гранулярная ЭПС обеспечивает синтез всех мембранных белков, в том числе
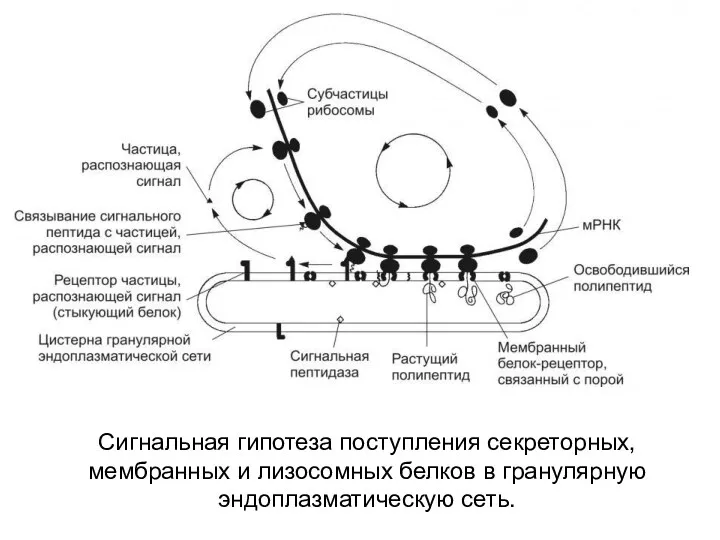
Сигнальная гипотеза поступления секреторных, мембранных и лизосомных белков в гранулярную эндоплазматическую
Сигнальная гипотеза поступления секреторных, мембранных и лизосомных белков в гранулярную эндоплазматическую

Функции грЭПС
Обеспечивает синтез белков, липидов и углеводов, а также начальное посттрансляционное
Функции грЭПС
Обеспечивает синтез белков, липидов и углеводов, а также начальное посттрансляционное

Агранулярная ЭПС
Это участки эндоплазматической сети, на которых отсутствуют рибосомы и осуществляет:
Агранулярная ЭПС
Это участки эндоплазматической сети, на которых отсутствуют рибосомы и осуществляет:
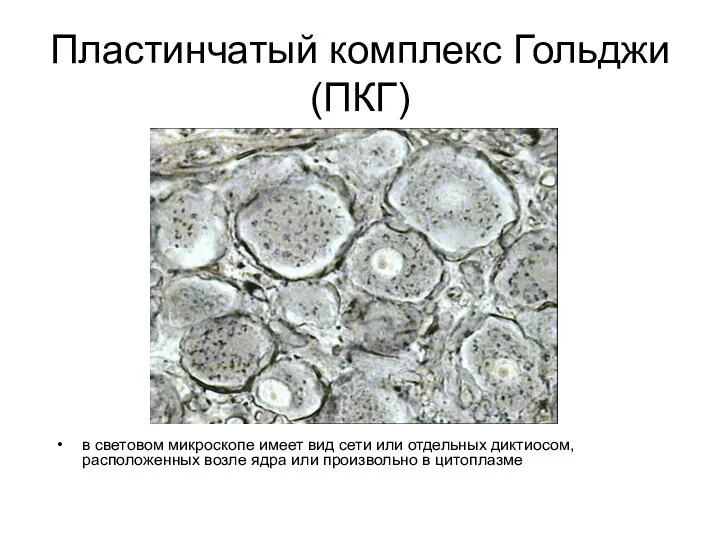
Пластинчатый комплекс Гольджи (ПКГ)
в световом микроскопе имеет вид сети или
Пластинчатый комплекс Гольджи (ПКГ)
в световом микроскопе имеет вид сети или
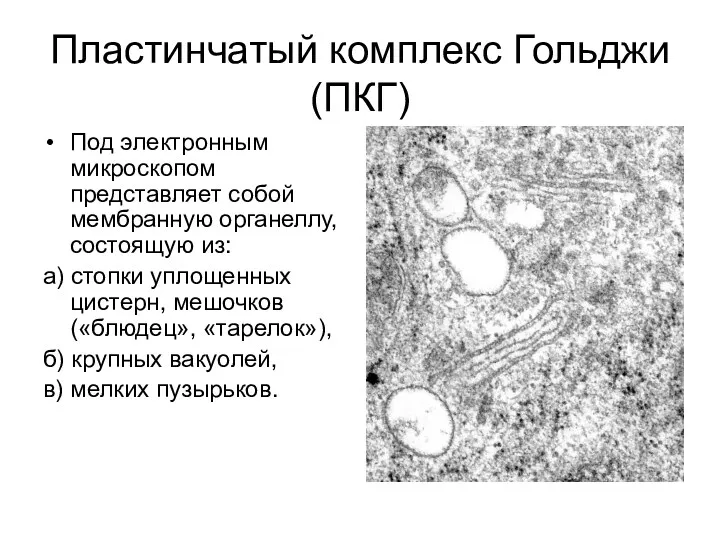
Пластинчатый комплекс Гольджи (ПКГ)
Под электронным микроскопом представляет собой мембранную органеллу, состоящую
Пластинчатый комплекс Гольджи (ПКГ)
Под электронным микроскопом представляет собой мембранную органеллу, состоящую
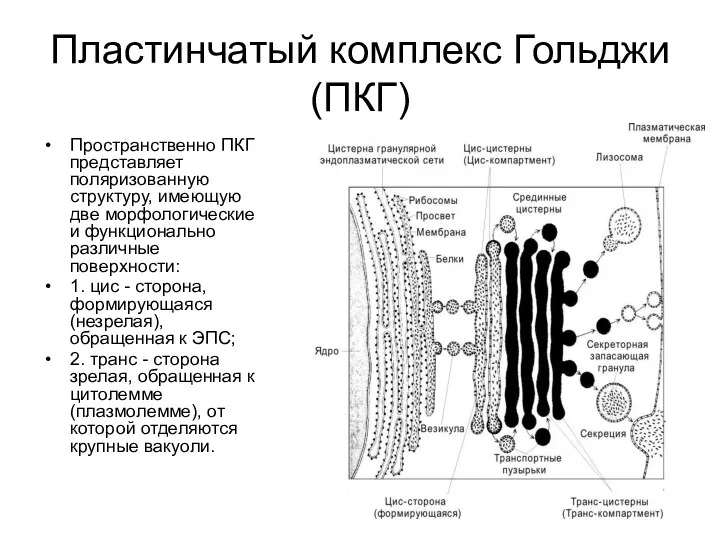
Пластинчатый комплекс Гольджи (ПКГ)
Пространственно ПКГ представляет поляризованную структуру, имеющую две морфологические
Пластинчатый комплекс Гольджи (ПКГ)
Пространственно ПКГ представляет поляризованную структуру, имеющую две морфологические

Пластинчатый комплекс Гольджи (ПКГ)
Цистерны имеют вид изогнутых дисков, диаметром 0,5 –
Пластинчатый комплекс Гольджи (ПКГ)
Цистерны имеют вид изогнутых дисков, диаметром 0,5 –


Функции ПКГ
Доработка (процессинг) продуктов, поступивших из ЭПС (включение углеводных или липидных
Функции ПКГ
Доработка (процессинг) продуктов, поступивших из ЭПС (включение углеводных или липидных
 Фізика в житті тварин. Трение в жизни растений и животных
Фізика в житті тварин. Трение в жизни растений и животных Дихання рослин і тварин. Значення дихання для організмів. Обмін речовин і енергії
Дихання рослин і тварин. Значення дихання для організмів. Обмін речовин і енергії Карантинные сорняки
Карантинные сорняки Этапы развития жизни на Земли
Этапы развития жизни на Земли Полезные и вредные комнатные растения
Полезные и вредные комнатные растения Презентация к уроку по биологии Цветок 6 класс
Презентация к уроку по биологии Цветок 6 класс Мочевыделительная система
Мочевыделительная система жизнь в морях и океанах(1)
жизнь в морях и океанах(1) Семя, его строение и условия прорастания семян
Семя, его строение и условия прорастания семян Нуклеин қышқылдарының құрылысы және биосинтезі
Нуклеин қышқылдарының құрылысы және биосинтезі Репликация и транскрипция
Репликация и транскрипция Кедры России. Проект
Кедры России. Проект Флора и растительность Челябинской области
Флора и растительность Челябинской области Моё домашнее животное - хомяк
Моё домашнее животное - хомяк Классификация и строение микроорганизмов. Микробиология
Классификация и строение микроорганизмов. Микробиология презентация к уроку по теме Кровь и кровообращение. 8 класс
презентация к уроку по теме Кровь и кровообращение. 8 класс Пресмыкающиеся или рептилии
Пресмыкающиеся или рептилии Микробы - друзья и враги человека
Микробы - друзья и враги человека Урок-презентация В бобовом царстве, мотыльковом государстве
Урок-презентация В бобовом царстве, мотыльковом государстве Урок биологии Головной мозг, 8 класс
Урок биологии Головной мозг, 8 класс Презентация к уроку биологии в 6 классе по программе VIII вида Гранит и его свойства
Презентация к уроку биологии в 6 классе по программе VIII вида Гранит и его свойства Ученые биологи и их вклад в науку
Ученые биологи и их вклад в науку Нуклеиновые кислоты и синтез белка в организме
Нуклеиновые кислоты и синтез белка в организме Оплодотворение. Способы оплодотворения
Оплодотворение. Способы оплодотворения Тест по теме Лишайники
Тест по теме Лишайники Птахи нашої місцевості
Птахи нашої місцевості конспект урока по биологии
конспект урока по биологии Класс Насекомые
Класс Насекомые