- Генетична карта поліомавірусів dsDNA

Содержание
- 2. Генетична карта поліомавірусів вірусу SV-40 вірусу поліоми
- 3. Genome Organization of SV40 Early unit: large T antigen (LT), small t antigen (sT), 17K T
- 4. Functional organization of SV40 large T antigen 1. Activation of cellular DNA and RNA synthesis binding
- 5. SV 40 DNA replication In the nucleus. Large T antigen binds to the SV40 origin Using
- 6. The replication of SV40 DNA
- 7. Genomic Organization of Papillomavirus Genome 7Kb circular genome Encode all proteins on the same DNA strand
- 8. Early Region E2 (Regulator of DNA replication and RNA transcription) Functional Assignments of Viral Early Proteins
- 9. Early Region Functional Assignments of Viral Early Proteins
- 10. Modulation of growth regulatory mechanisms Early Region Functional Assignments of Viral Early Proteins
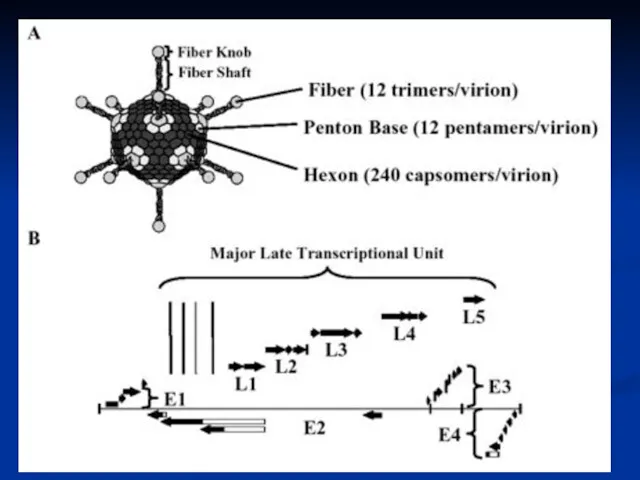
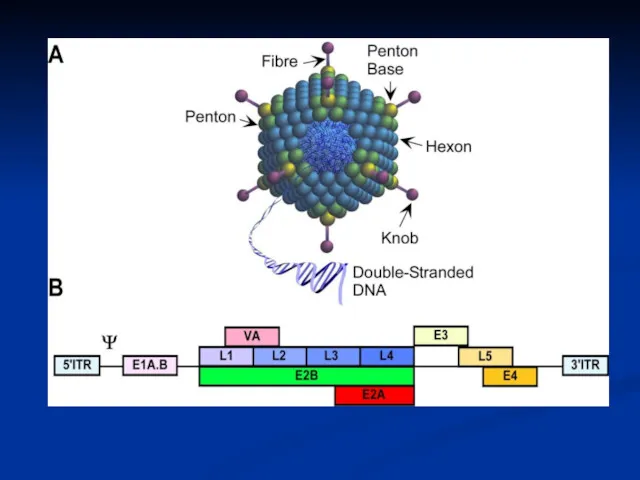
- 15. Adenovirus Replication Endosome Virion Integrins CAR Virion DNA Nuclear pore Nucleus Cytoplasm coxsackie-adenovirus receptor
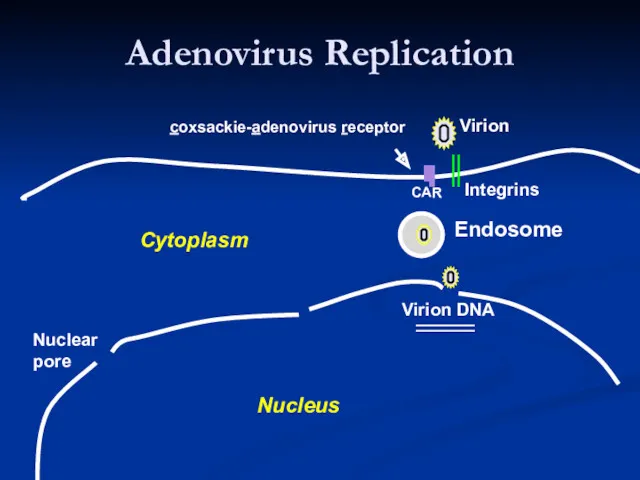
- 16. Adenovirus Replication Endosome Early mRNAs Early proteins Virion Integrins CAR Virion DNA Modulate Cell Cycle Shut
- 17. Adenovirus Replication Endosome Late mRNAs Early mRNAs Early proteins Late proteins Virion Integrins CAR Virion DNA
- 18. The early protein include: For transcription EIA is referred to as an ‘immediate early’ gene) For
- 19. Генетична карта вірусу простого герпесу
- 20. Транскрипція вірусу простого герпесу
- 21. Реплікація ДНК вірусу простого герпесу
- 22. Блочний запис генетичної інформації у герпесвірусів
- 23. Генетична карта бактеріофагу Т4
- 24. Схема розрізання конкатемеру фага Т4 Конкатемер ДНК фагу Т4 Кінцеві надлишки Розрізання Геном фагу, що пакується
- 25. Генетична карта бактеріофагу лямбда
- 26. Інтеграція геному фагу лямбда
- 27. POXVIRIDAE Figure 2 Schematic representation of the DNA of Vaccinia virus (VACV) (WR strain): (Top) Linear
- 28. Figure 3 The infectious cycle of Vaccinia virus (VACV): IMV, intracellular mature virus; EEV, extracellular enveloped
- 29. The poxvirus genome comprises a linear molecule of dsDNA with covalently closed termini; terminal hairpins constitute
- 30. Генетична карта вірусу PBCV-1
- 32. Скачать презентацию
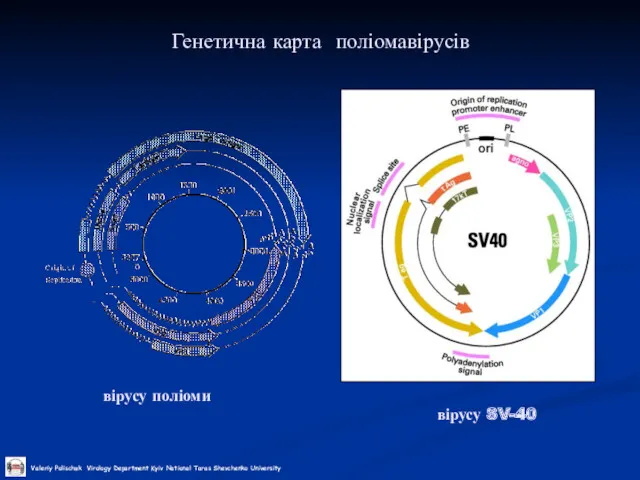
Генетична карта поліомавірусів
вірусу SV-40
вірусу поліоми
Генетична карта поліомавірусів
вірусу SV-40
вірусу поліоми
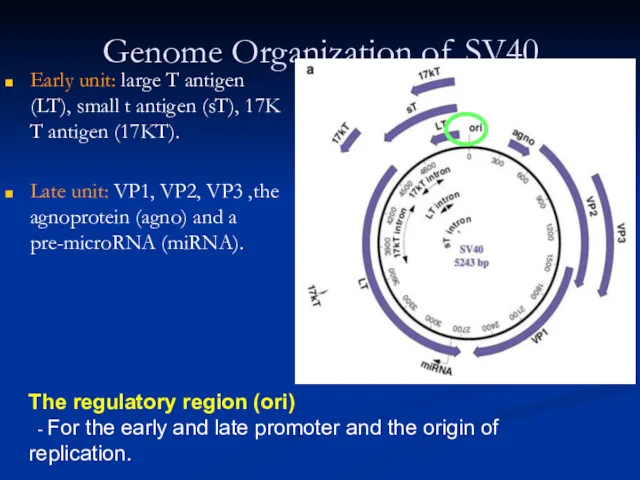
Genome Organization of SV40
Early unit: large T antigen (LT), small t
Genome Organization of SV40
Early unit: large T antigen (LT), small t

Functional organization of SV40 large T antigen
1. Activation of cellular DNA
Functional organization of SV40 large T antigen
1. Activation of cellular DNA

SV 40 DNA replication
In the nucleus.
Large T antigen binds to
SV 40 DNA replication
In the nucleus.
Large T antigen binds to
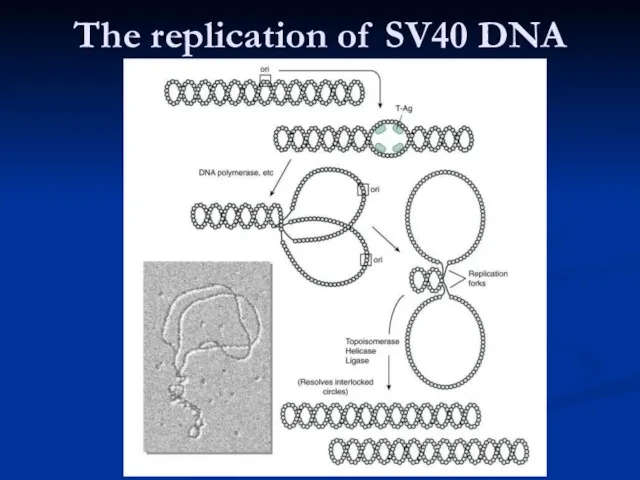
The replication of SV40 DNA
The replication of SV40 DNA

Genomic Organization of Papillomavirus
Genome
7Kb circular genome
Encode all proteins on the
Genomic Organization of Papillomavirus
Genome
7Kb circular genome
Encode all proteins on the
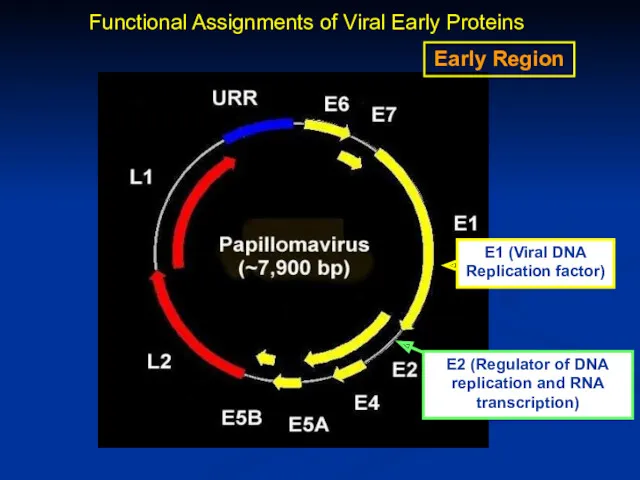
Early Region
E2 (Regulator of DNA replication and RNA transcription)
Functional Assignments of
Early Region
E2 (Regulator of DNA replication and RNA transcription)
Functional Assignments of
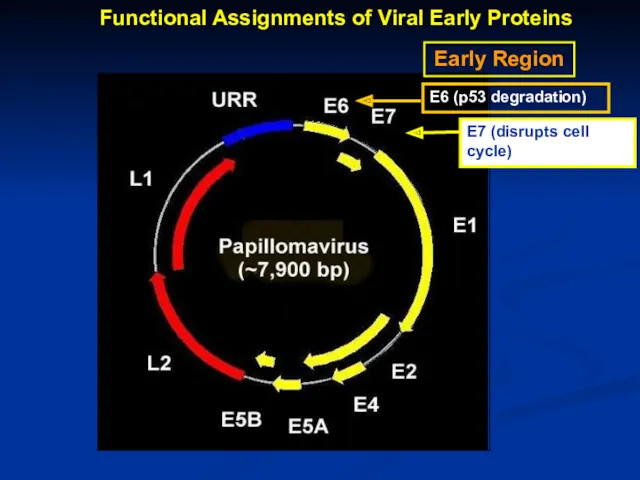
Early Region
Functional Assignments of Viral Early Proteins
Early Region
Functional Assignments of Viral Early Proteins
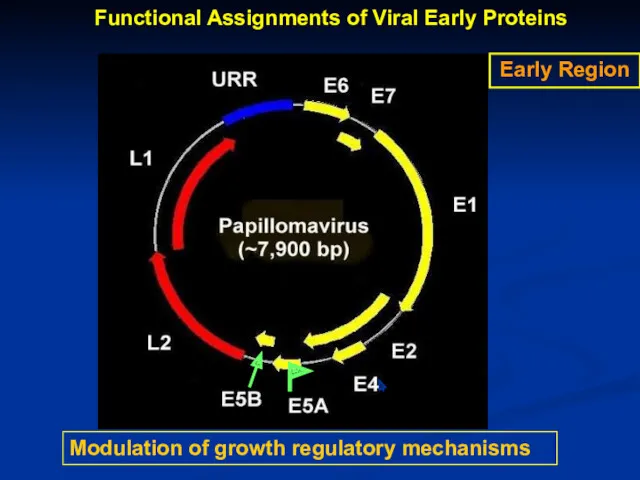
Modulation of growth regulatory mechanisms
Early Region
Functional Assignments of Viral Early Proteins
Modulation of growth regulatory mechanisms
Early Region
Functional Assignments of Viral Early Proteins

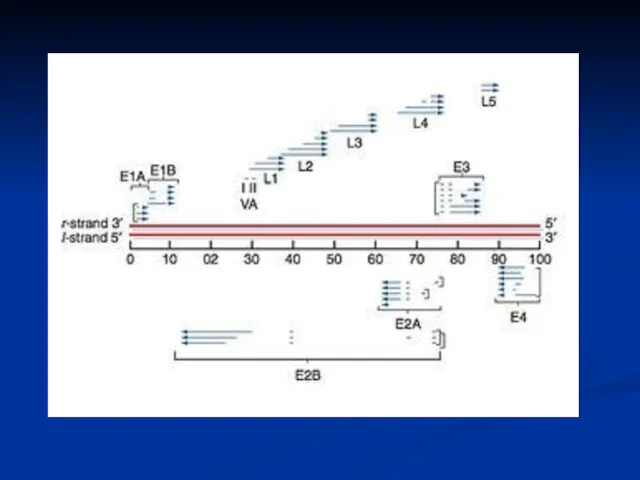
Adenovirus Replication
Endosome
Virion
Integrins
CAR
Virion DNA
Nuclear
pore
Nucleus
Cytoplasm
coxsackie-adenovirus receptor
Adenovirus Replication
Endosome
Virion
Integrins
CAR
Virion DNA
Nuclear
pore
Nucleus
Cytoplasm
coxsackie-adenovirus receptor
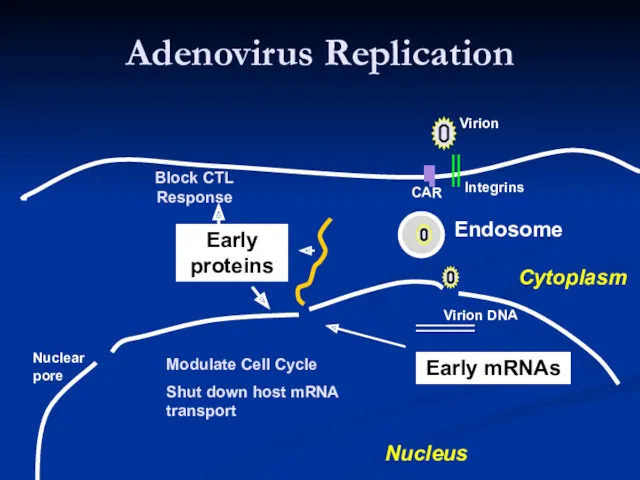
Adenovirus Replication
Endosome
Early mRNAs
Early
proteins
Virion
Integrins
CAR
Virion DNA
Modulate Cell Cycle
Shut down host mRNA transport
Block CTL
Adenovirus Replication
Endosome
Early mRNAs
Early
proteins
Virion
Integrins
CAR
Virion DNA
Modulate Cell Cycle
Shut down host mRNA transport
Block CTL
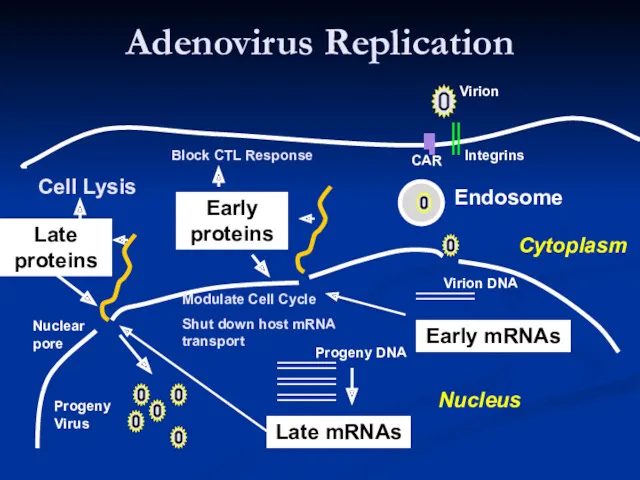
Adenovirus Replication
Endosome
Late mRNAs
Early mRNAs
Early
proteins
Late
proteins
Virion
Integrins
CAR
Virion DNA
Modulate Cell Cycle
Shut down host mRNA transport
Progeny
Adenovirus Replication
Endosome
Late mRNAs
Early mRNAs
Early
proteins
Late
proteins
Virion
Integrins
CAR
Virion DNA
Modulate Cell Cycle
Shut down host mRNA transport
Progeny

The early protein include:
For transcription
EIA is referred to as an
The early protein include:
For transcription
EIA is referred to as an

Генетична карта вірусу простого герпесу
Генетична карта вірусу простого герпесу

Транскрипція вірусу простого герпесу
Транскрипція вірусу простого герпесу

Реплікація ДНК вірусу простого герпесу
Реплікація ДНК вірусу простого герпесу
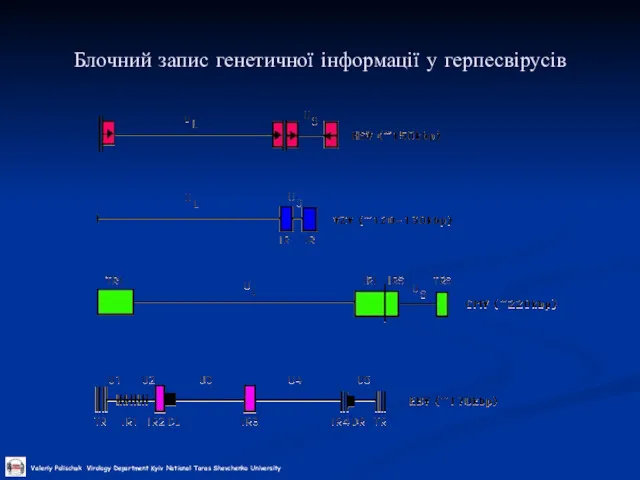
Блочний запис генетичної інформації у герпесвірусів
Блочний запис генетичної інформації у герпесвірусів
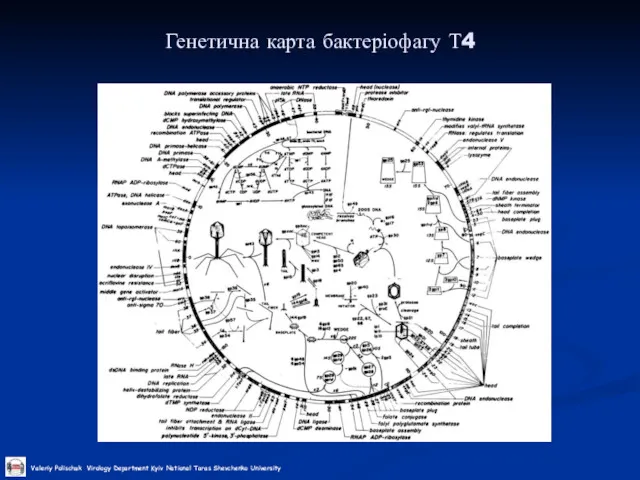
Генетична карта бактеріофагу Т4
Генетична карта бактеріофагу Т4

Схема розрізання конкатемеру фага Т4
Конкатемер ДНК фагу Т4
Кінцеві надлишки
Розрізання
Геном фагу, що
Схема розрізання конкатемеру фага Т4
Конкатемер ДНК фагу Т4
Кінцеві надлишки
Розрізання
Геном фагу, що
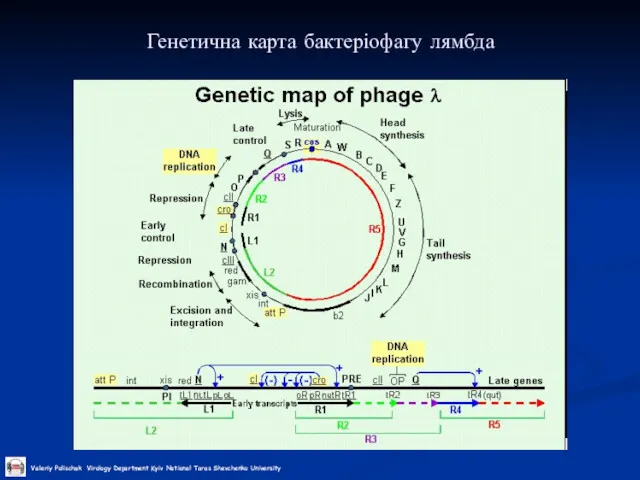
Генетична карта бактеріофагу лямбда
Генетична карта бактеріофагу лямбда
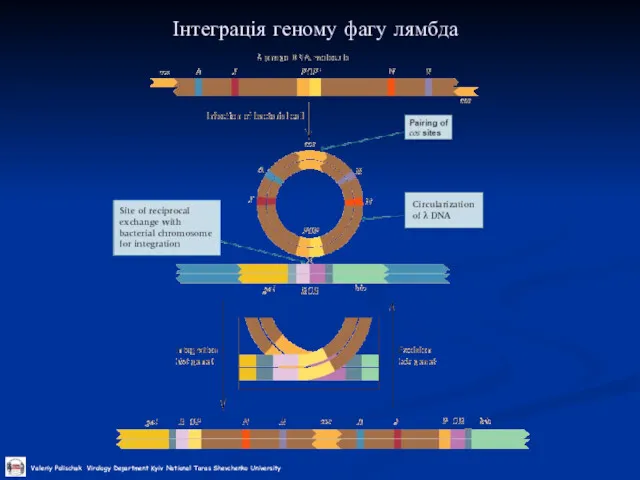
Інтеграція геному фагу лямбда
Інтеграція геному фагу лямбда
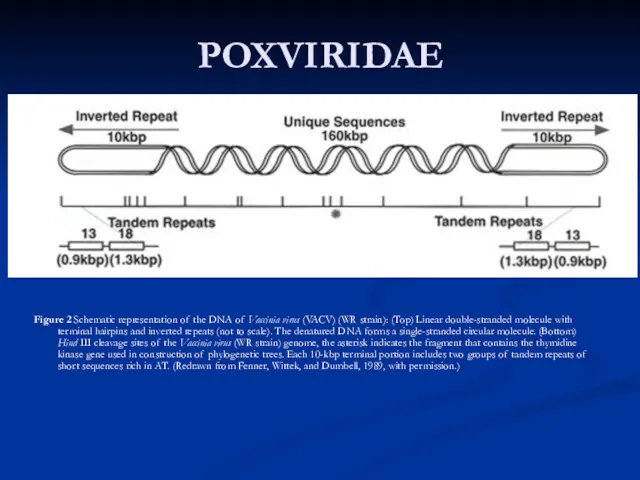
POXVIRIDAE
Figure 2 Schematic representation of the DNA of Vaccinia virus (VACV)
POXVIRIDAE
Figure 2 Schematic representation of the DNA of Vaccinia virus (VACV)
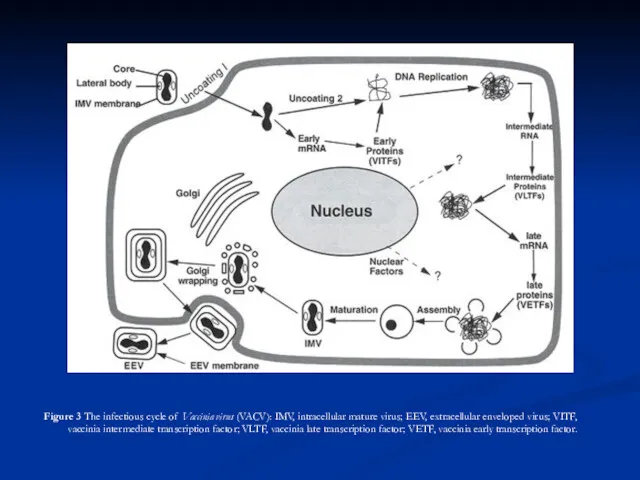
Figure 3 The infectious cycle of Vaccinia virus (VACV): IMV,
Figure 3 The infectious cycle of Vaccinia virus (VACV): IMV,

The poxvirus genome comprises a linear molecule of dsDNA with
The poxvirus genome comprises a linear molecule of dsDNA with
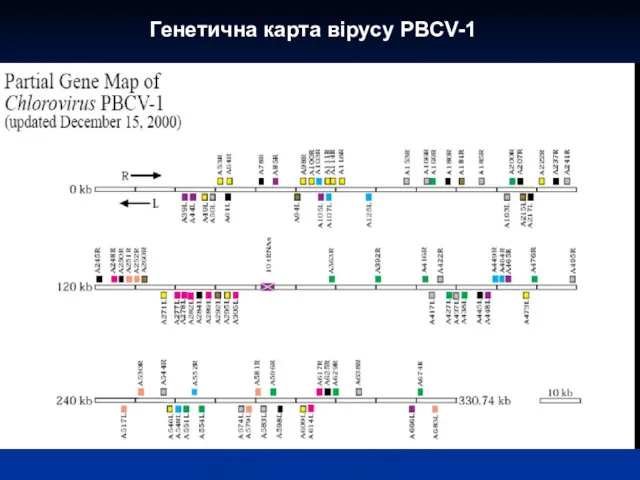
Генетична карта вірусу PBCV-1
Генетична карта вірусу PBCV-1
 Семейство лилейные, отдел цветковые или покрытосеменные
Семейство лилейные, отдел цветковые или покрытосеменные Комнатные растения (часть 2)
Комнатные растения (часть 2) Цитологические основы наследственности
Цитологические основы наследственности What is the engine of our body machine
What is the engine of our body machine Птицы
Птицы У Чёрного моря
У Чёрного моря Вредители питомников и молодняков. (Лекция 5)
Вредители питомников и молодняков. (Лекция 5) Ткани растений
Ткани растений Изучение биоритмов человека – их влияние на жизнедеятельность
Изучение биоритмов человека – их влияние на жизнедеятельность Кормовая и сахарная свёкла
Кормовая и сахарная свёкла Сана және өзіндік сана
Сана және өзіндік сана Растительный и животный мир Республики Удмуртия
Растительный и животный мир Республики Удмуртия Интересные растения
Интересные растения Жасуша ядросы
Жасуша ядросы Бидай. Бидайдың зиянкестері
Бидай. Бидайдың зиянкестері Значение бактерий в природе и жизни человека
Значение бактерий в природе и жизни человека Устройство речевого аппарата
Устройство речевого аппарата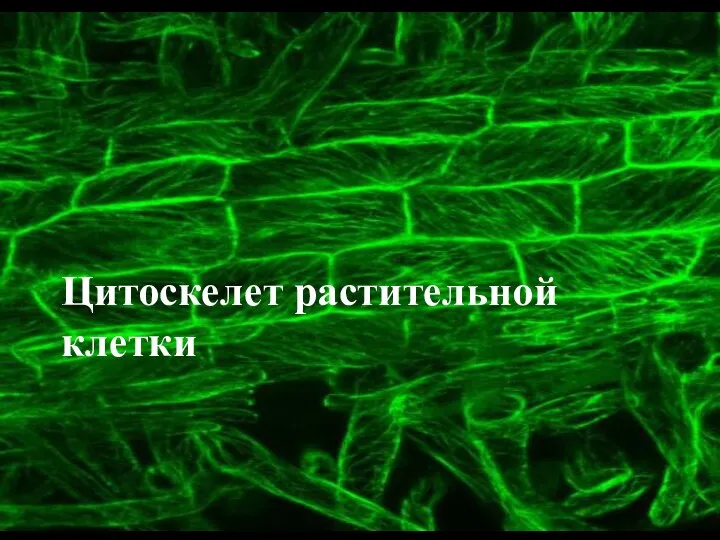 Цитоскелет растительной клетки
Цитоскелет растительной клетки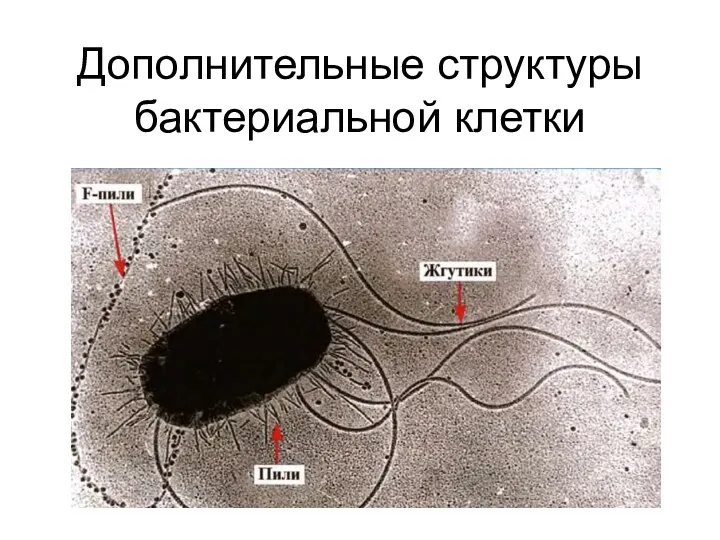 Дополнительные структуры бактериальной клетки
Дополнительные структуры бактериальной клетки Влияние факторов внешней среды на онтогенез
Влияние факторов внешней среды на онтогенез Морфология эпителиальной ткани
Морфология эпителиальной ткани Продолговатый мозг. Черепно-мозговые нервы (IX - XII)
Продолговатый мозг. Черепно-мозговые нервы (IX - XII) Сүт және сүт өнідерінің тағамдық биологиялық құндылығы мен қауіпсіздігі
Сүт және сүт өнідерінің тағамдық биологиялық құндылығы мен қауіпсіздігі Индивидуальное развитие цветковых растений
Индивидуальное развитие цветковых растений Генная инженерия: новые возможности и проблемы
Генная инженерия: новые возможности и проблемы Клонирование
Клонирование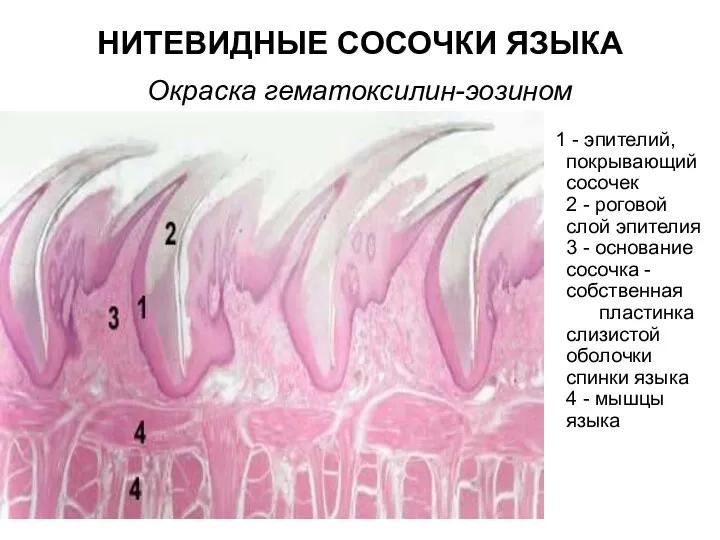 Нитевидные сосочки языка. Окраска гематоксилин-эозином
Нитевидные сосочки языка. Окраска гематоксилин-эозином Птицы Челябинской области
Птицы Челябинской области