- Гены и аллели. Аллельные взаимодействия

Содержание
- 2. МЕНДЕЛЕВСКАЯ ГЕНЕТИКА Мендель: наследуемость отдельных признаков у гороха (Pisum sativum). цитологические наблюдения за хромосомами подтвердили менделевские
- 3. Горох размножается самоопылением: растения устроены таким образом, что пыльца обычно попадает на рыльце пестика того же
- 5. Моногибридное скрещивание одна пара контрастных признаков Моногибридное скрещивание родители – Р1 (parental generation) потомство – F1
- 6. моногибридное скрещивание растений с длинным и коротким стеблем F1 – только высокие растения. При опылении низких
- 7. В F1 было зарегистрировано лишь одно из пары альтернативных проявлений каждого признака, названное доминантным. Первый закон
- 8. Анализируя результаты скрещиваний по другим парам контрастных признаков, Мендель получил такую же картину.
- 9. Мендель предположил существование единиц наследственности, отвечающих за каждый признак. Наследственные признаки контролируются единицами, представленными у каждого
- 10. Если один организм несет два наследственных фактора, отвечающих за разные контрастные признаки, то одна единица наследственности
- 11. Родительские формы (Р) были константны каждый из них содержал задатки только одного типа гомозиготны по задаткам
- 12. Гетерозиготным называется организм, произошедший от слияния гамет, несущих различные наследственные задатки. Гибриды F1 образуются в результате
- 13. решетка Пеннета для моногибридного скрещивания
- 14. Г. Мендель: выдвинул математически обоснованную и проверяемую гипотезу комбинирования наследственных задатков, с помощью которой можно предсказать
- 15. Современная терминология Наследственные единичные формы гена называют аллелями. Единицы или факторы, которые постулировал Мендель, обусловливают внешние
- 16. Анализирующее скрещивание если действительно гетерозиготы F1 образуют с равной вероятностью гаметы, несущие доминантные и рецессивные задатки
- 17. Скрещивание формы с доминантным признаком и формы – гомозиготного рецессива получило название анализирующего скрещивания. скрещивая гибриды
- 18. рецессивные задатки не исчезают в гетерозиготном организме, а остаются неизменными и вновь проявляются при встрече с
- 19. Позднее У. Бэтсон, исходя из этого феномена, сформулировал правило чистоты гамет, согласно которому явление расщепления основано
- 20. Генотип – это совокупность наследственных задатков организма; фенотип – совокупность признаков организма.
- 21. Закон независимого наследования признаков Г. Мендель: две формы гороха, различающийся одновременно по форме и по окраске
- 22. Частоты фенотипов: правило произведения вероятностей: если два независимых события происходят одновременно, то вероятность двух исходов равна
- 24. Контрольное скрещивание: два признака 31 круглых желтых (AaBb) 26 круглых зеленых (Aabb) 27 морщинистых желтых (aaBb)
- 25. Тригибридное скрещивание Результаты тригибридного, или трехфактороного скрещивания выглядят более сложными, чем при дигибридном скрещивании.
- 26. Закон независимого наследования признаков С учетом независимого комбинирования аллелей можно определить эти соотношения для любого числа
- 27. Повторное открытие законов Менделя Выводы Менделя не соответствовали представлениям о причинах изменчивости организмов. Считалось, что изменчивость
- 28. В конце XIX в. Вальтер Флемминг, исследуя клетки саламандры, впервые описал клеточное ядро и хромосомы. 1879
- 29. начало ХХ в. Гуго де Фриз, Карл Корренс и Эрих Чермак, независимо друг от друга, провели
- 30. 1902 г. Вальтер Саттон в США и Теодор Бовери в Германии предположили, что гены находятся в
- 31. Моногенное наследование Вальсирующие мыши (в F1 нормальное поведение; в F2 – ¾ нормальных : ¼ вальсирующих)
- 32. В 1902 г. А. Гаррод начал публикации о врожденных аномалиях метаболизма у человека. известно более 100
- 33. Аутосомно-доминантный тип наследования Брахидактилия
- 34. Родословная с брахидактилией
- 36. Отклонение от пропорций Менделя
- 37. Генотип и фенотип В случае полного доминирования расщепление по генотипу 1АА : 2Аа : 1 аа
- 38. неполное доминирование – наблюдается промежуточное выражение у гетерозигот при сравнении с обеими гомозиготными родительскими формами красная
- 40. Синдром Тея-Сакса аа – тяжелое нарушение липидного обмена смерть в течение первых трех лет жизни нарушения
- 41. Кодоминирование Совместная экспрессия двух аллелей в гетерозиготном фенотипе называется кодоминированием. наследование группы крови MN у человека.
- 42. Другие взаимодействия аллелей множественный аллелизм Группы крови АВ0 описанны в 1900-х годах Карлом Ландштайнером ген I
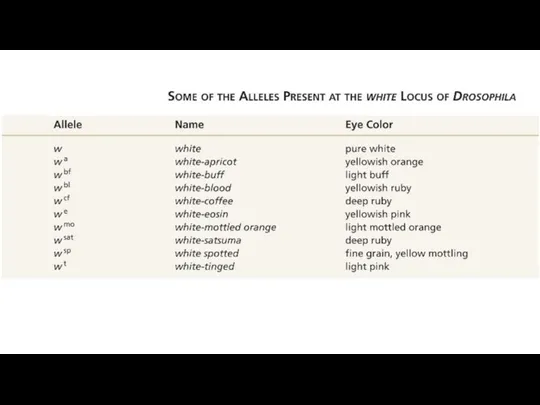
- 44. Локус white у Drosophila 1921 г. Томас Хант Морган и Кальвин Бриджес рецессивная мутация белоглазости у
- 46. Плейотропия. один ген оказывает влияние на несколько признаков. Мендель отмечал, что один из изучаемых им генов
- 47. У высших растений гены, обуславливающие красную (антоциановую) окраску цветков, одновременно контролируют красную окраску стебля. У человека
- 48. Дрозофилы, гомозиготные по аллелю vg, имеют зачаточные крылья. изменены жужальца одна из пар дорзальных щетинок расположена
- 49. У человека – рецессивный ген определяющий фенилкетонурию – болезнь, приводящую к серьезным умственным нарушениям. Нормальный ген
- 50. Люди, гомозиготные по этому гену и не подвергавшиеся лечению, отличаются от нормальных по уровню содержания фенилаланина
- 51. Летальные аллели если мутации относятся к летальным, то гомозиготные носители рецессивных леталей не выживают.
- 52. Платиновая окраска шерсти у лисиц доминантный ген, который существует только в гетерозиготе, поскольку обладает рецессивным летальным
- 53. Гены Х-хромосомы Гены, передающиеся в составе Х-хромосомы называются Х-сцепленными.
- 55. Ограниченное полом и зависящее от пола наследование признаков Самки и самцы у домашних кур резко отличаются
- 56. У кур, независимо от того, несут они доминантный (Н) или рецессивный аллель (h) этого гена, оперенье
- 57. Куры породы Леггорн генотип hh, поэтому петухи сильно отличаются по своему оперенью от кур. Все особи
- 58. Продукция молока у крупного рогатого скота. Независимо от генотипа, который влияет на количество молока, оно вырабатывается
- 59. Частичное облысение у человека рога у некоторых пород овец (например, у Дорсеттской)
- 60. Пенетрантность и экспрессивность Доля особей, у которых, в той или иной мере, проявляется мутантный фенотип, указывает
- 61. Пенетрантность гена –доля особей, у которых проявляется ожидаемый фенотип. Экспрессивность –степень выраженности фенотипа (у тех особей,
- 62. влияние внешней среды и (или) генотипа. Сальмонелла, несущая мутацию гена, кодирующего ИГФ-синтетазу, неспособна расти на минимальной
- 63. Способность генотипа так или иначе проявляться в различных условиях среды отражает норму его реакции – способность
- 64. Д.К. Беляев с сотрудниками показал можно добиться рождения живых щенков, гомозиготных по доминантной аллели платиновой окраски,
- 65. Родословная с ретинобластомой в случае неполной пенетрантности. Непораженная женщина II. 4 должна быть гетерозиготой, поскольку ее
- 66. Проявление экспрессии генов Хорея Гентингтона – аутосомно доминантный тип Поражение переднего отдела и коры головного мозга
- 67. Болезнь Гентингтона (первоначально хорея Гентингтона) у человека наследуется по доминантно-аутосомному типу. Поражение переднего отдела и коры
- 68. неполная пенетрантности и варьирующая экспрессивность – проявление доминантного гена, вызывающего хорею Гентингтона у человека. непроизвольные подергивания
- 69. Распределение 802 случаев хореи Гентингтона по возрасту начала заболевания
- 70. Генетическая антисипация Генетические болезни, симптомы которых появляются в раннем возрасте и усиливаются в каждом последующем поколении
- 71. Мышечная дистрофия взрослых – миотонической дистрофии (МД). Аутосомно-доминантный тип, варьирование по тяжести. Средняя тяжесть: катаракта, небольшая
- 72. Участок гена МД представлен в виде вариабельного числа повторов. У здоровых людей около пяти копий этого
- 74. Геномный (родительский) импринтинг Зависит от происхождения хромосомы несущей определенный ген. Отдельные участки хромосом у некоторых видов
- 75. Инактивация одной их Х-хромосом у самок млекопитающих. У мышей на ранних стадиях развития зародыша во всех
- 76. Дифференциальный импринтинг человеческой хромосомы 15 обусловил два наследственных заболевания. происходит одинаковая по размеру делеция участка одной
- 77. Другие взаимодействия аллелей У дрожжей S. cerevisidae есть формы, наследственно различающиеся по окраске колоний: красные и
- 78. генетический блок в биосинтезе пуринов: отсутствует активность фермента фосфорибозиламиноимидазолкарбоксилазы (мутант ade2), и поэтому дрожжи для своего
- 79. Если исследовать гомозиготы по рецессивным аллелям ade 2, то в этом случае неактивная молекула белка состоит
- 81. Каждый функциональный центр обычно представлен отдельным доменом – полуавтономным участком специфически уложенной полипептидной цепи
- 82. В некоторых случаях при объединении в гибриде двух разных аллелей независимого происхождения, рецессивных по отношению к
- 83. Это явление описано для многих объектов: дрозофилы, мышей, зеленой водоросли Chlamydomonas, многих грибов и т.д. Примерно
- 85. Гены аллельные Лежат в одинаковых локусах гомологичных хромосом и отвечают за один признак Обозначаются (обычно) одинаковыми
- 86. Неаллельное взаимодействие генов
- 87. Генетический анализ при взаимодействии генов Один из первых примеров взаимодействия генов был обнаружен в начале ХХ
- 88. Розовидный х гороховидный Р Aabb aaBB F1 AaBa Ореховидный
- 89. 1. Присутствие доминантных аллелей А и В у 9/16 кур второго поколения ведет к образованию нового
- 90. взаимодействие доминантных генов А и В изменяет форму гребня. расщепление дигибридного скрещивания нарушается, НО: общее расщепление
- 91. При взаимодействии генов в случае дигибридных скрещиваний расщепление в F2 по фенотипу может быть различным: 9
- 92. Новые фенотипы Наследование плода у тыквы Cucurbita pepo. При скрещивании растений с дисковидной формой плодов (ААВВ)
- 93. комплементарность, или комплементарного (взаимно дополнительного) действия, когда доминантные аллели одного гена обусловили нормальный (или дикий) фенотип
- 94. Анализ комплементации аллелей Две линии дрозофил с недоразвитыми крыльями Чтобы определить являются ли эти мутации аллелями
- 95. 1. Все потомство имеет нормальные крылья. Интерпретация: Две рецессивные мутации не являются аллельными и локализованы в
- 96. Анализ комплементации, разработанный генетиком Эдвардом Л. Льюисом, часто называют цис-транс тестом. цис – означает положение аллелей
- 97. Комплементарность. У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое число форм, наследственно различающихся по
- 98. F1 все мухи имеют темно-красные глаза (дикий тип) F2: четыре класса расщепления: мухи с темно-красными ярко-красными
- 99. формально-генетический анализ полностью абстрагируются от механизмов действия исследуемых генов.
- 100. Биохимический механизм взаимодействия генов st и bw исследован достаточно подробно.
- 101. Эпистаз Гомозиготное состояние двух рецессивных аллелей первого локуса перекрывает экспрессию других аллелей и/или прекращает их экспрессию.
- 102. Наследование цвета шерсти у мышей. Окраска дикого типа обусловлена чередованием кольцевых полос пигмента по длине каждого
- 103. В присутствии аллеля А из молекулы предшественника образуется черный пигмент. В присутствии аллеля В черный пигмент
- 104. Другой тип эпистаза: доминантный аллель одного локуса маскирует экспрессию аллелей другого локуса. Доминантный аллель W обуславливает
- 105. Фенотип крови Бомбей Антигены A и B происходят из одного предшественника, Н-вещества, к молекулам которого затем
- 106. Дефектное Н-вещество не связывается с ферментом, добавляющим концевые углеводные остатки. приводит к фенотипу Бомбей. этот дефект
- 107. Полимерия. Пастушья сумка с треугольными и овальными плодами некумулятивная полимерия.
- 108. Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии. шведский генетик Г. Нильсон-Эле в
- 109. Гены-модификаторы. Природа генов-модификаторов до сих пор вызывает споры: в частности, не ясно, существуют ли специальные модификаторы,
- 110. Модификатор – это ген, влияющий на фенотипическое проявление гена другого локуса. У мышей ген пятнистой окраски
- 111. Количественные признаки Открытие Менделем основных законов наследственности оказалось возможным благодаря тому, что он анализировал контрастирующие признаки,
- 112. Непрерывная изменчивость Для некоторых признаков характерна дискретная, или прерывистая, изменчивость. признаки, изученные Менделем, А, В, АВ
- 113. Непрерывная изменчивость Признаки проявляющие непрерывную изменчивость, называют количественными, или метрическими, поскольку разница между особями невелика и
- 114. Наследование количественных признаков в основе беспрерывной изменчивости лежат наследственные факторы, для количественных признаков характерно полигенное наследование.
- 115. Аддитивное действие генов – основа непрерывной изменчивости 1. признаки с непрерывной изменчивостью обычно можно измерить (взвесить,
- 116. Определение числа генов, детерминирующих признак
- 117. Если число генов мало, то для определения количества ожидаемых фенотипических классов проще пользоваться формулой (2n +
- 118. Значение полигенного наследования признаков Полигенные признаки: рост, вес, экстерьер животных, высота и урожайность зерна у растений,
- 119. У человека: пигментация кожи, степень развития интеллекта, формы поведения, полнота, предрасположенность к определенным заболеваниям. полнота и
- 120. Анализ полигенных признаков Распределение частот встречаемости фенотипов обычно имеет характер нормального распределения. Для оценки экспериментальных данных
- 121. Для анализа признаков с нормальным распределением величины используют: Среднюю величину Дисперсию Стандартное отклонение Стандартную ошибку средней
- 122. Наследуемость Имея методы измерения количественных признаков, можно оценить в какой степени данный признак определяется генотипом и
- 123. Наследуемость в широком смысле Один из способов оценки изменчивости количественных признаков состоит в использовании инбредных линий.
- 124. Эксперимент по искусственному отбору – сельскохозяйственная лаборатория штата Иллинойс (США) С 1896 г. Отбирают растения с
- 125. Обычно наследуемость значимых для организма признаков мала, поскольку, в основном, она обусловлена генетически и за время
- 126. Близнецовый метод у человека Для определения влияния генетических и средовых факторов на фенотип человека исследуются монозиготные,
- 127. Полученные результаты сравнивают с результатами анализа дизиготных, или не идентичных близнецов, которые развиваются из двух яйцеклеток,
- 128. Другой подход состоит в определении величины конкордантности фенотипов у пар близнецов выросших вместе. Если у одного
- 129. Если у монозиготных близнецов конкордантность по признаку выше чем у дизиготных, то этот признак, в основном,
- 130. Во многих случаях признаки могут считаться полигенными, но полигенность наследования трудно доказать при существенном влиянии на
- 132. Скачать презентацию

МЕНДЕЛЕВСКАЯ ГЕНЕТИКА
Мендель: наследуемость отдельных признаков у гороха (Pisum sativum).
цитологические наблюдения за
МЕНДЕЛЕВСКАЯ ГЕНЕТИКА
Мендель: наследуемость отдельных признаков у гороха (Pisum sativum).
цитологические наблюдения за
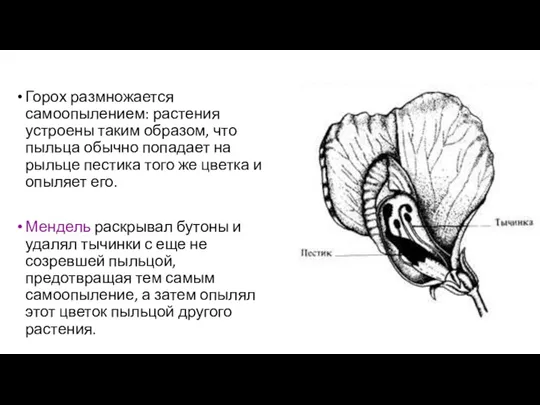
Горох размножается самоопылением: растения устроены таким образом, что пыльца обычно попадает
Горох размножается самоопылением: растения устроены таким образом, что пыльца обычно попадает


Моногибридное скрещивание
одна пара контрастных признаков
Моногибридное скрещивание
родители – Р1 (parental generation)
Моногибридное скрещивание
одна пара контрастных признаков
Моногибридное скрещивание
родители – Р1 (parental generation)
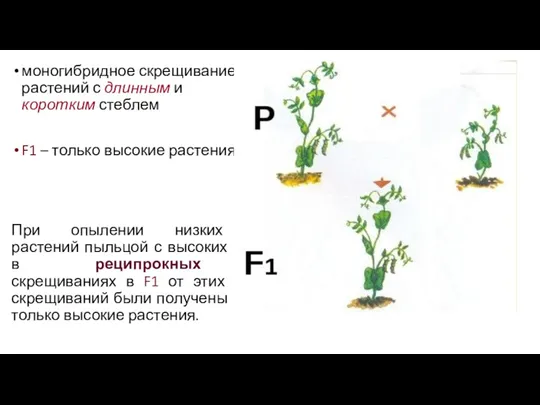
моногибридное скрещивание растений с длинным и коротким стеблем
F1 – только
моногибридное скрещивание растений с длинным и коротким стеблем
F1 – только

В F1 было зарегистрировано лишь одно из пары альтернативных проявлений каждого

Анализируя результаты скрещиваний по другим парам контрастных признаков, Мендель получил такую
Анализируя результаты скрещиваний по другим парам контрастных признаков, Мендель получил такую
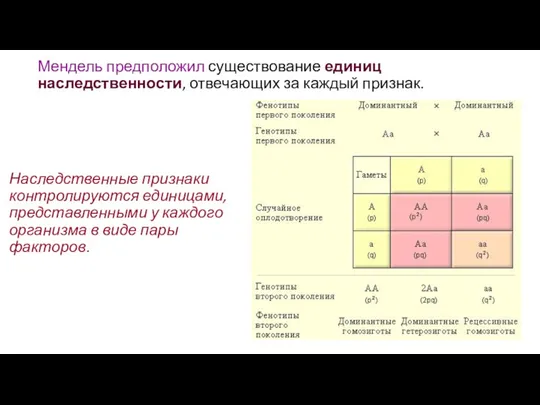
Мендель предположил существование единиц наследственности, отвечающих за каждый признак.
Наследственные признаки контролируются
Мендель предположил существование единиц наследственности, отвечающих за каждый признак.
Наследственные признаки контролируются
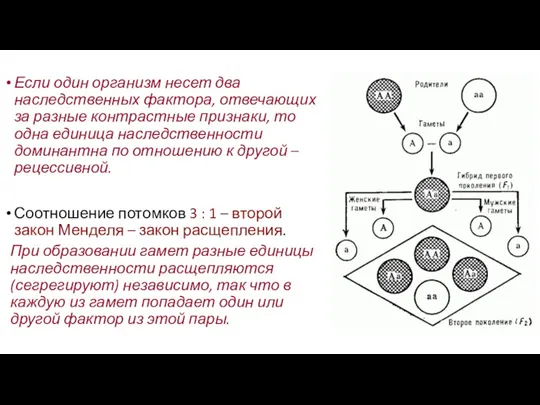
Если один организм несет два наследственных фактора, отвечающих за разные контрастные
Если один организм несет два наследственных фактора, отвечающих за разные контрастные

Родительские формы (Р) были константны
каждый из них содержал задатки только одного
Родительские формы (Р) были константны
каждый из них содержал задатки только одного
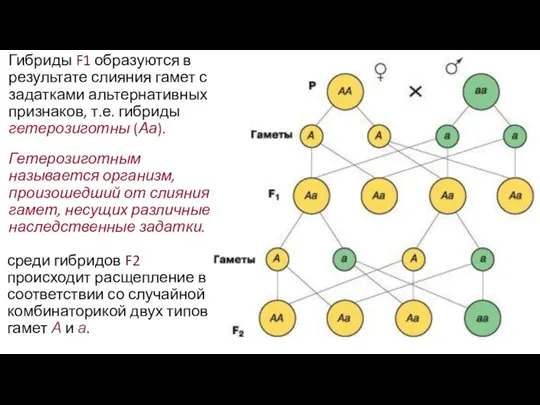
Гетерозиготным называется организм, произошедший от слияния гамет, несущих различные наследственные задатки.
Гибриды
Гетерозиготным называется организм, произошедший от слияния гамет, несущих различные наследственные задатки.
Гибриды

решетка Пеннета для моногибридного скрещивания
решетка Пеннета для моногибридного скрещивания

Г. Мендель:
выдвинул математически обоснованную и проверяемую гипотезу комбинирования наследственных задатков, с
Г. Мендель:
выдвинул математически обоснованную и проверяемую гипотезу комбинирования наследственных задатков, с

Современная терминология
Наследственные единичные формы гена называют аллелями.
Единицы или факторы, которые постулировал
Современная терминология
Наследственные единичные формы гена называют аллелями.
Единицы или факторы, которые постулировал
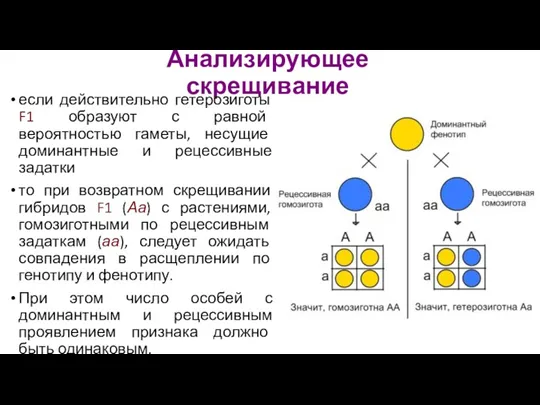
Анализирующее скрещивание
если действительно гетерозиготы F1 образуют с равной вероятностью гаметы, несущие
Анализирующее скрещивание
если действительно гетерозиготы F1 образуют с равной вероятностью гаметы, несущие
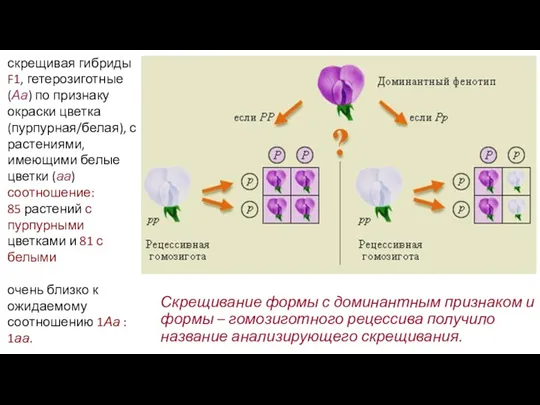
Скрещивание формы с доминантным признаком и формы – гомозиготного рецессива получило
Скрещивание формы с доминантным признаком и формы – гомозиготного рецессива получило

рецессивные задатки не исчезают в гетерозиготном организме, а остаются неизменными и
рецессивные задатки не исчезают в гетерозиготном организме, а остаются неизменными и

Позднее У. Бэтсон, исходя из этого феномена, сформулировал правило чистоты гамет,
Позднее У. Бэтсон, исходя из этого феномена, сформулировал правило чистоты гамет,
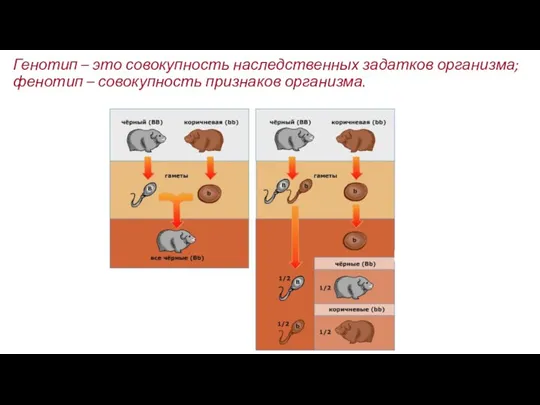
Генотип – это совокупность наследственных задатков организма; фенотип – совокупность признаков
Генотип – это совокупность наследственных задатков организма; фенотип – совокупность признаков
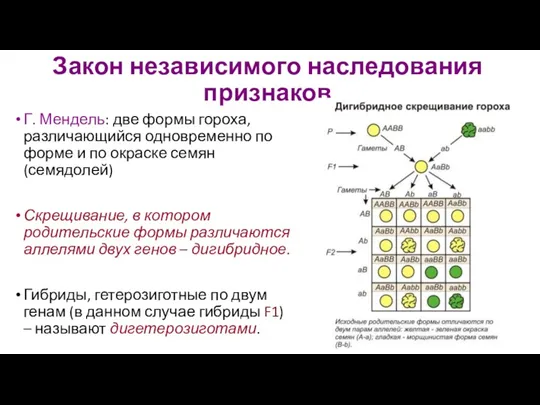
Закон независимого наследования признаков
Г. Мендель: две формы гороха, различающийся одновременно по
Закон независимого наследования признаков
Г. Мендель: две формы гороха, различающийся одновременно по
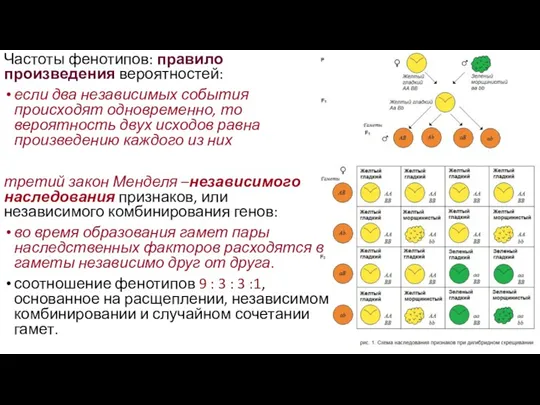
Частоты фенотипов: правило произведения вероятностей:
если два независимых события происходят одновременно,
Частоты фенотипов: правило произведения вероятностей:
если два независимых события происходят одновременно,
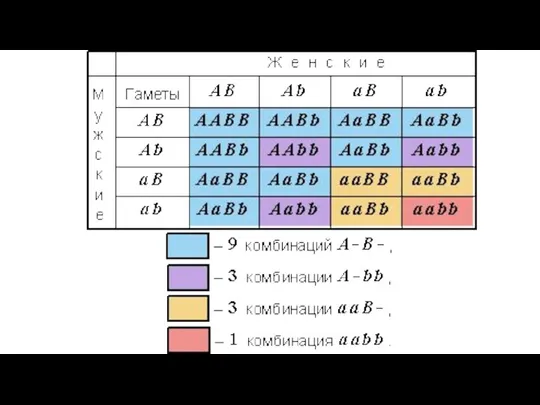

Контрольное скрещивание: два признака
31 круглых желтых (AaBb)
26 круглых зеленых (Aabb)
27 морщинистых
Контрольное скрещивание: два признака
31 круглых желтых (AaBb)
26 круглых зеленых (Aabb)
27 морщинистых

Тригибридное скрещивание
Результаты тригибридного, или трехфактороного скрещивания выглядят более сложными, чем при
Тригибридное скрещивание
Результаты тригибридного, или трехфактороного скрещивания выглядят более сложными, чем при
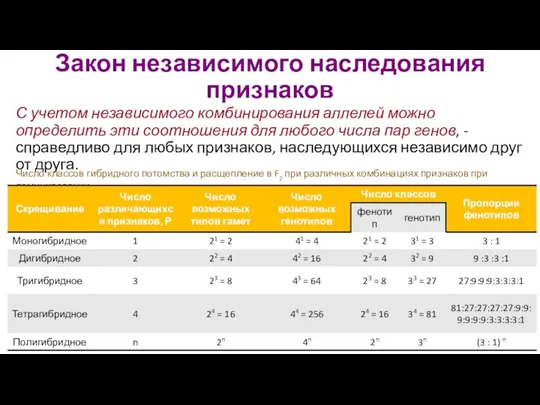
Закон независимого наследования признаков
С учетом независимого комбинирования аллелей можно определить эти
Закон независимого наследования признаков
С учетом независимого комбинирования аллелей можно определить эти

Повторное открытие законов Менделя
Выводы Менделя не соответствовали представлениям о причинах изменчивости
Повторное открытие законов Менделя
Выводы Менделя не соответствовали представлениям о причинах изменчивости

В конце XIX в. Вальтер Флемминг, исследуя клетки саламандры, впервые описал
В конце XIX в. Вальтер Флемминг, исследуя клетки саламандры, впервые описал
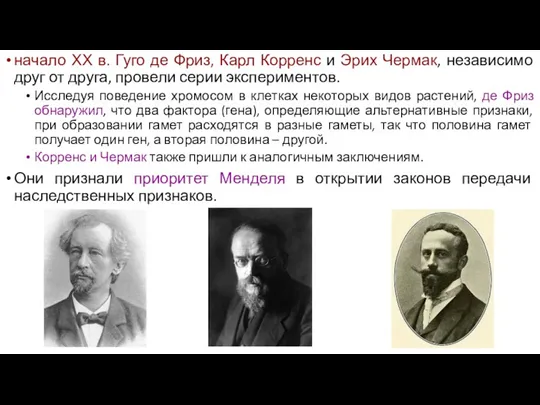
начало ХХ в. Гуго де Фриз, Карл Корренс и Эрих Чермак,
начало ХХ в. Гуго де Фриз, Карл Корренс и Эрих Чермак,

1902 г. Вальтер Саттон в США и Теодор Бовери в Германии
1902 г. Вальтер Саттон в США и Теодор Бовери в Германии

Моногенное наследование
Вальсирующие мыши (в F1 нормальное поведение; в F2 – ¾
Моногенное наследование
Вальсирующие мыши (в F1 нормальное поведение; в F2 – ¾
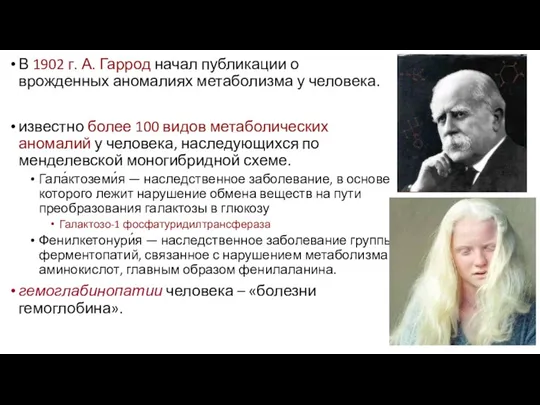
В 1902 г. А. Гаррод начал публикации о врожденных аномалиях метаболизма
В 1902 г. А. Гаррод начал публикации о врожденных аномалиях метаболизма

Аутосомно-доминантный тип наследования
Брахидактилия
Аутосомно-доминантный тип наследования
Брахидактилия

Родословная с брахидактилией
Родословная с брахидактилией

Отклонение от пропорций Менделя
Отклонение от пропорций Менделя

Генотип и фенотип
В случае полного доминирования расщепление по генотипу
1АА :
Генотип и фенотип
В случае полного доминирования расщепление по генотипу
1АА :

неполное доминирование – наблюдается промежуточное выражение у гетерозигот при сравнении с
неполное доминирование – наблюдается промежуточное выражение у гетерозигот при сравнении с


Синдром Тея-Сакса
аа – тяжелое нарушение липидного обмена
смерть в течение первых
Синдром Тея-Сакса
аа – тяжелое нарушение липидного обмена
смерть в течение первых

Кодоминирование
Совместная экспрессия двух аллелей в гетерозиготном фенотипе называется кодоминированием.
наследование группы крови
Кодоминирование
Совместная экспрессия двух аллелей в гетерозиготном фенотипе называется кодоминированием.
наследование группы крови

Другие взаимодействия аллелей
множественный аллелизм
Группы крови АВ0 описанны в 1900-х годах Карлом
Другие взаимодействия аллелей
множественный аллелизм
Группы крови АВ0 описанны в 1900-х годах Карлом


Локус white у Drosophila
1921 г. Томас Хант Морган и Кальвин Бриджес
Локус white у Drosophila
1921 г. Томас Хант Морган и Кальвин Бриджес

Плейотропия.
один ген оказывает влияние на несколько признаков.
Мендель отмечал, что один из
Плейотропия.
один ген оказывает влияние на несколько признаков.
Мендель отмечал, что один из

У высших растений гены, обуславливающие красную (антоциановую) окраску цветков, одновременно контролируют
У высших растений гены, обуславливающие красную (антоциановую) окраску цветков, одновременно контролируют

Дрозофилы, гомозиготные по аллелю vg, имеют зачаточные крылья.
изменены жужальца
одна из пар
Дрозофилы, гомозиготные по аллелю vg, имеют зачаточные крылья.
изменены жужальца
одна из пар

У человека – рецессивный ген определяющий фенилкетонурию – болезнь, приводящую к
У человека – рецессивный ген определяющий фенилкетонурию – болезнь, приводящую к

Люди, гомозиготные по этому гену и не подвергавшиеся лечению, отличаются от
Люди, гомозиготные по этому гену и не подвергавшиеся лечению, отличаются от

Летальные аллели
если мутации относятся к летальным, то гомозиготные носители рецессивных леталей
Летальные аллели
если мутации относятся к летальным, то гомозиготные носители рецессивных леталей

Платиновая окраска шерсти у лисиц
доминантный ген, который существует только в
Платиновая окраска шерсти у лисиц
доминантный ген, который существует только в

Гены Х-хромосомы
Гены, передающиеся в составе Х-хромосомы называются Х-сцепленными.
Гены Х-хромосомы
Гены, передающиеся в составе Х-хромосомы называются Х-сцепленными.

Ограниченное полом и зависящее от пола наследование признаков
Самки и самцы у
Ограниченное полом и зависящее от пола наследование признаков
Самки и самцы у

У кур, независимо от того, несут они доминантный (Н) или рецессивный
У кур, независимо от того, несут они доминантный (Н) или рецессивный

Куры породы Леггорн генотип hh, поэтому петухи сильно отличаются по своему
Куры породы Леггорн генотип hh, поэтому петухи сильно отличаются по своему

Продукция молока у крупного рогатого скота.
Независимо от генотипа, который влияет на
Продукция молока у крупного рогатого скота.
Независимо от генотипа, который влияет на

Частичное облысение у человека
рога у некоторых пород овец (например, у
Частичное облысение у человека
рога у некоторых пород овец (например, у

Пенетрантность и экспрессивность
Доля особей, у которых, в той или иной мере,
Пенетрантность и экспрессивность
Доля особей, у которых, в той или иной мере,

Пенетрантность гена –доля особей, у которых проявляется ожидаемый фенотип.
Экспрессивность –степень выраженности
Пенетрантность гена –доля особей, у которых проявляется ожидаемый фенотип.
Экспрессивность –степень выраженности
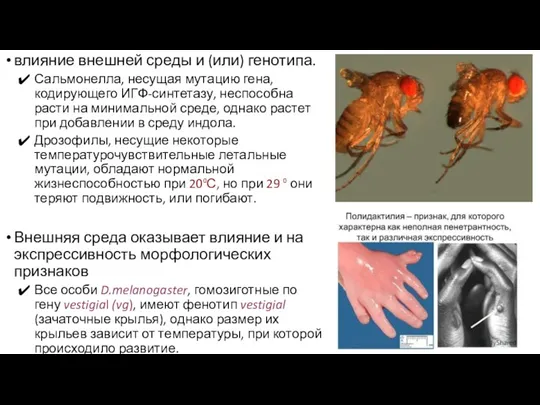
влияние внешней среды и (или) генотипа.
Сальмонелла, несущая мутацию гена, кодирующего ИГФ-синтетазу,
влияние внешней среды и (или) генотипа.
Сальмонелла, несущая мутацию гена, кодирующего ИГФ-синтетазу,
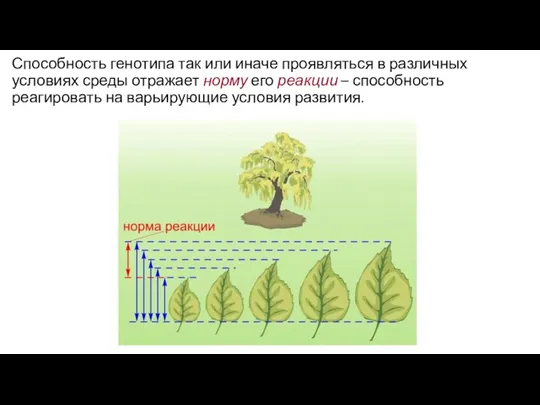
Способность генотипа так или иначе проявляться в различных условиях среды отражает
Способность генотипа так или иначе проявляться в различных условиях среды отражает

Д.К. Беляев с сотрудниками показал
можно добиться рождения живых щенков, гомозиготных по
Д.К. Беляев с сотрудниками показал
можно добиться рождения живых щенков, гомозиготных по
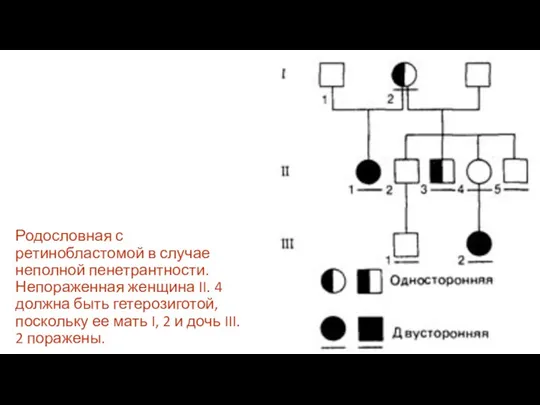
Родословная с ретинобластомой в случае неполной пенетрантности. Непораженная женщина II. 4
Родословная с ретинобластомой в случае неполной пенетрантности. Непораженная женщина II. 4

Проявление экспрессии генов
Хорея Гентингтона – аутосомно доминантный тип
Поражение переднего отдела и
Проявление экспрессии генов
Хорея Гентингтона – аутосомно доминантный тип
Поражение переднего отдела и

Болезнь Гентингтона (первоначально хорея Гентингтона) у человека наследуется по доминантно-аутосомному типу.
Поражение
Болезнь Гентингтона (первоначально хорея Гентингтона) у человека наследуется по доминантно-аутосомному типу.
Поражение

неполная пенетрантности и варьирующая экспрессивность – проявление доминантного гена, вызывающего хорею
неполная пенетрантности и варьирующая экспрессивность – проявление доминантного гена, вызывающего хорею
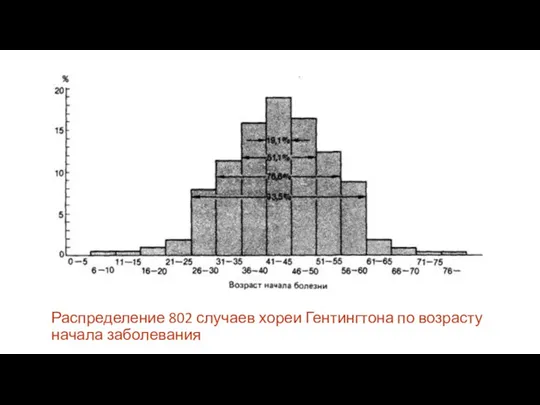
Распределение 802 случаев хореи Гентингтона по возрасту начала заболевания
Распределение 802 случаев хореи Гентингтона по возрасту начала заболевания

Генетическая антисипация
Генетические болезни, симптомы которых появляются в раннем возрасте и усиливаются
Генетическая антисипация
Генетические болезни, симптомы которых появляются в раннем возрасте и усиливаются

Мышечная дистрофия взрослых – миотонической дистрофии (МД).
Аутосомно-доминантный тип, варьирование по тяжести.
Средняя
Мышечная дистрофия взрослых – миотонической дистрофии (МД).
Аутосомно-доминантный тип, варьирование по тяжести.
Средняя

Участок гена МД представлен в виде вариабельного числа повторов.
У здоровых людей
Участок гена МД представлен в виде вариабельного числа повторов.
У здоровых людей


Геномный (родительский) импринтинг
Зависит от происхождения хромосомы несущей определенный ген.
Отдельные участки хромосом
Геномный (родительский) импринтинг
Зависит от происхождения хромосомы несущей определенный ген.
Отдельные участки хромосом

Инактивация одной их Х-хромосом у самок млекопитающих.
У мышей на ранних стадиях
Инактивация одной их Х-хромосом у самок млекопитающих.
У мышей на ранних стадиях

Дифференциальный импринтинг человеческой хромосомы 15 обусловил два наследственных заболевания.
происходит одинаковая по
Дифференциальный импринтинг человеческой хромосомы 15 обусловил два наследственных заболевания.
происходит одинаковая по

Другие взаимодействия аллелей
У дрожжей S. cerevisidae есть формы, наследственно различающиеся по
Другие взаимодействия аллелей
У дрожжей S. cerevisidae есть формы, наследственно различающиеся по

генетический блок в биосинтезе пуринов: отсутствует активность фермента фосфорибозиламиноимидазолкарбоксилазы (мутант ade2),
генетический блок в биосинтезе пуринов: отсутствует активность фермента фосфорибозиламиноимидазолкарбоксилазы (мутант ade2),

Если исследовать гомозиготы по рецессивным аллелям ade 2, то в этом
Если исследовать гомозиготы по рецессивным аллелям ade 2, то в этом

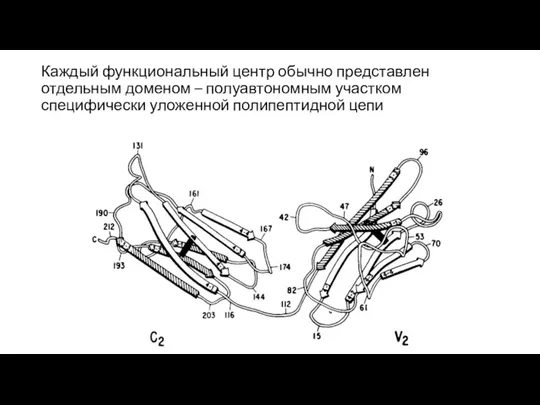
Каждый функциональный центр обычно представлен отдельным доменом – полуавтономным участком специфически
Каждый функциональный центр обычно представлен отдельным доменом – полуавтономным участком специфически

В некоторых случаях при объединении в гибриде двух разных аллелей независимого
В некоторых случаях при объединении в гибриде двух разных аллелей независимого

Это явление описано для многих объектов: дрозофилы, мышей, зеленой водоросли Chlamydomonas,
Это явление описано для многих объектов: дрозофилы, мышей, зеленой водоросли Chlamydomonas,


Гены
аллельные
Лежат в одинаковых локусах гомологичных хромосом и отвечают за один
Гены
аллельные
Лежат в одинаковых локусах гомологичных хромосом и отвечают за один
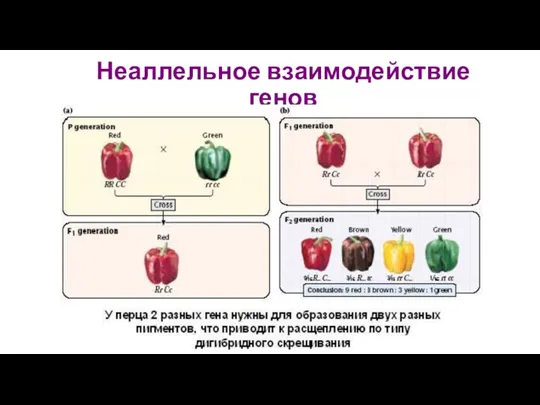
Неаллельное взаимодействие генов
Неаллельное взаимодействие генов

Генетический анализ при взаимодействии генов
Один из первых примеров взаимодействия генов был
Генетический анализ при взаимодействии генов
Один из первых примеров взаимодействия генов был
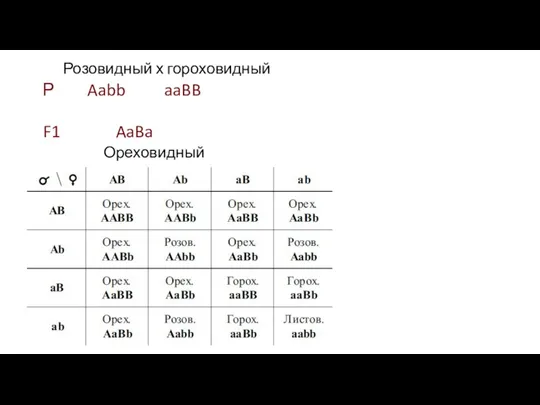
Розовидный х гороховидный
Р Aabb aaBB
F1 AaBa
Ореховидный
Розовидный х гороховидный
Р Aabb aaBB
F1 AaBa
Ореховидный

1. Присутствие доминантных аллелей А и В у 9/16 кур второго
1. Присутствие доминантных аллелей А и В у 9/16 кур второго
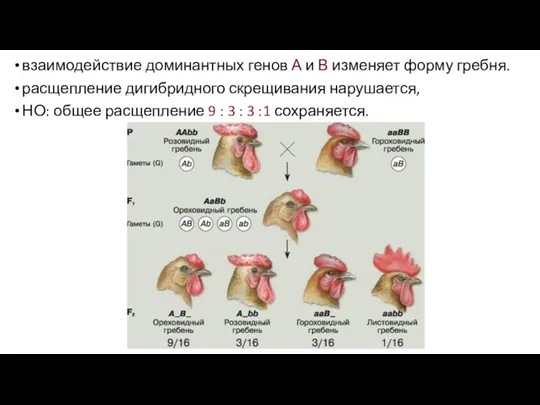
взаимодействие доминантных генов А и В изменяет форму гребня.
расщепление дигибридного скрещивания
взаимодействие доминантных генов А и В изменяет форму гребня.
расщепление дигибридного скрещивания

При взаимодействии генов в случае дигибридных скрещиваний расщепление в F2 по
При взаимодействии генов в случае дигибридных скрещиваний расщепление в F2 по

Новые фенотипы
Наследование плода у тыквы Cucurbita pepo.
При скрещивании растений с дисковидной
Новые фенотипы
Наследование плода у тыквы Cucurbita pepo.
При скрещивании растений с дисковидной

комплементарность, или комплементарного (взаимно дополнительного) действия, когда доминантные аллели одного гена
комплементарность, или комплементарного (взаимно дополнительного) действия, когда доминантные аллели одного гена

Анализ комплементации аллелей
Две линии дрозофил с недоразвитыми крыльями
Чтобы определить являются ли
Анализ комплементации аллелей
Две линии дрозофил с недоразвитыми крыльями
Чтобы определить являются ли

1. Все потомство имеет нормальные крылья.
Интерпретация: Две рецессивные мутации не являются
1. Все потомство имеет нормальные крылья.
Интерпретация: Две рецессивные мутации не являются

Анализ комплементации, разработанный генетиком Эдвардом Л. Льюисом, часто называют цис-транс тестом.
цис
Анализ комплементации, разработанный генетиком Эдвардом Л. Льюисом, часто называют цис-транс тестом.
цис

Комплементарность.
У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое
Комплементарность.
У популярного генетического объекта плодовой мушки Drosophila melanogaster имеется большое

F1 все мухи имеют темно-красные глаза (дикий тип)
F2: четыре класса расщепления:
F1 все мухи имеют темно-красные глаза (дикий тип)
F2: четыре класса расщепления:

формально-генетический анализ
полностью абстрагируются от механизмов действия исследуемых генов.
формально-генетический анализ
полностью абстрагируются от механизмов действия исследуемых генов.

Биохимический механизм взаимодействия генов st и bw исследован достаточно подробно.
Биохимический механизм взаимодействия генов st и bw исследован достаточно подробно.

Эпистаз
Гомозиготное состояние двух рецессивных аллелей первого локуса перекрывает экспрессию других аллелей
Эпистаз
Гомозиготное состояние двух рецессивных аллелей первого локуса перекрывает экспрессию других аллелей

Наследование цвета шерсти у мышей.
Окраска дикого типа обусловлена чередованием кольцевых полос
Наследование цвета шерсти у мышей.
Окраска дикого типа обусловлена чередованием кольцевых полос

В присутствии аллеля А из молекулы предшественника образуется черный пигмент.
В присутствии
В присутствии аллеля А из молекулы предшественника образуется черный пигмент.
В присутствии
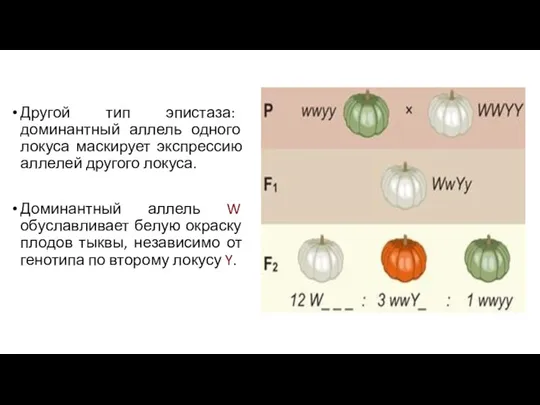
Другой тип эпистаза: доминантный аллель одного локуса маскирует экспрессию аллелей другого
Другой тип эпистаза: доминантный аллель одного локуса маскирует экспрессию аллелей другого

Фенотип крови Бомбей
Антигены A и B происходят из одного предшественника, Н-вещества,
Фенотип крови Бомбей
Антигены A и B происходят из одного предшественника, Н-вещества,

Дефектное Н-вещество не связывается с ферментом, добавляющим концевые углеводные остатки.
приводит к
Дефектное Н-вещество не связывается с ферментом, добавляющим концевые углеводные остатки.
приводит к

Полимерия.
Пастушья сумка с треугольными и овальными плодами
некумулятивная полимерия.
Полимерия.
Пастушья сумка с треугольными и овальными плодами
некумулятивная полимерия.

Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии.
шведский
Однозначные, или полимерные, гены могут действовать и по типу кумулятивной полимерии.
шведский

Гены-модификаторы.
Природа генов-модификаторов до сих пор вызывает споры: в частности, не ясно,
Гены-модификаторы.
Природа генов-модификаторов до сих пор вызывает споры: в частности, не ясно,

Модификатор – это ген, влияющий на фенотипическое проявление гена другого локуса.
У
Модификатор – это ген, влияющий на фенотипическое проявление гена другого локуса.
У

Количественные признаки
Открытие Менделем основных законов наследственности оказалось возможным благодаря тому, что
Количественные признаки
Открытие Менделем основных законов наследственности оказалось возможным благодаря тому, что
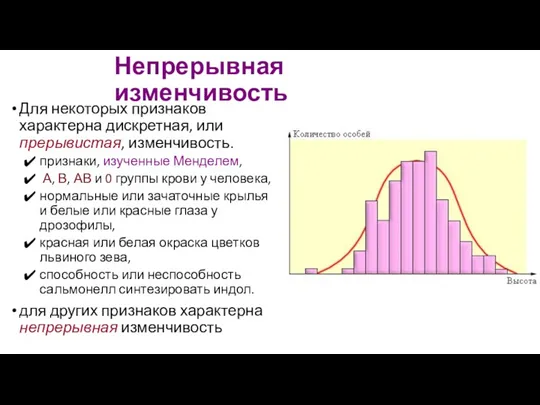
Непрерывная изменчивость
Для некоторых признаков характерна дискретная, или прерывистая, изменчивость.
признаки, изученные Менделем,
Непрерывная изменчивость
Для некоторых признаков характерна дискретная, или прерывистая, изменчивость.
признаки, изученные Менделем,

Непрерывная изменчивость
Признаки проявляющие непрерывную изменчивость, называют количественными, или метрическими, поскольку разница
Непрерывная изменчивость
Признаки проявляющие непрерывную изменчивость, называют количественными, или метрическими, поскольку разница

Наследование количественных признаков
в основе беспрерывной изменчивости лежат наследственные факторы,
для количественных
Наследование количественных признаков
в основе беспрерывной изменчивости лежат наследственные факторы,
для количественных

Аддитивное действие генов – основа непрерывной изменчивости
1. признаки с непрерывной изменчивостью
Аддитивное действие генов – основа непрерывной изменчивости
1. признаки с непрерывной изменчивостью

Определение числа генов, детерминирующих признак
Определение числа генов, детерминирующих признак

Если число генов мало, то для определения количества ожидаемых фенотипических классов
Если число генов мало, то для определения количества ожидаемых фенотипических классов

Значение полигенного наследования признаков
Полигенные признаки:
рост,
вес,
экстерьер животных,
высота и
Значение полигенного наследования признаков
Полигенные признаки:
рост,
вес,
экстерьер животных,
высота и

У человека:
пигментация кожи,
степень развития интеллекта,
формы поведения,
полнота,
предрасположенность
У человека:
пигментация кожи,
степень развития интеллекта,
формы поведения,
полнота,
предрасположенность
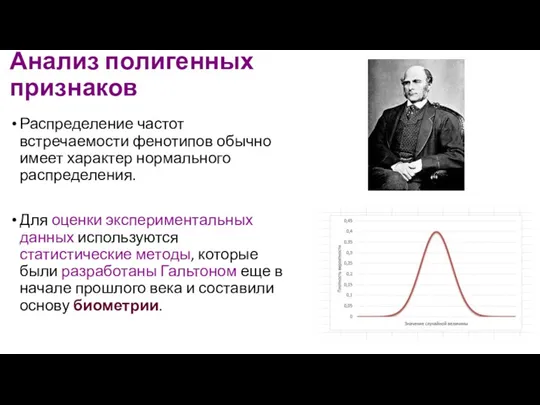
Анализ полигенных признаков
Распределение частот встречаемости фенотипов обычно имеет характер нормального распределения.
Для
Анализ полигенных признаков
Распределение частот встречаемости фенотипов обычно имеет характер нормального распределения.
Для

Для анализа признаков с нормальным распределением величины используют:
Среднюю величину
Дисперсию
Стандартное отклонение
Стандартную ошибку
Для анализа признаков с нормальным распределением величины используют:
Среднюю величину
Дисперсию
Стандартное отклонение
Стандартную ошибку

Наследуемость
Имея методы измерения количественных признаков, можно оценить в какой степени данный
Наследуемость
Имея методы измерения количественных признаков, можно оценить в какой степени данный

Наследуемость в широком смысле
Один из способов оценки изменчивости количественных признаков состоит
Наследуемость в широком смысле
Один из способов оценки изменчивости количественных признаков состоит

Эксперимент по искусственному отбору – сельскохозяйственная лаборатория штата Иллинойс (США)
С 1896
Эксперимент по искусственному отбору – сельскохозяйственная лаборатория штата Иллинойс (США)
С 1896

Обычно наследуемость значимых для организма признаков мала, поскольку, в основном, она
Обычно наследуемость значимых для организма признаков мала, поскольку, в основном, она

Близнецовый метод у человека
Для определения влияния генетических и средовых факторов на
Близнецовый метод у человека
Для определения влияния генетических и средовых факторов на

Полученные результаты сравнивают с результатами анализа дизиготных, или не идентичных близнецов,
Полученные результаты сравнивают с результатами анализа дизиготных, или не идентичных близнецов,

Другой подход состоит в определении величины конкордантности фенотипов у пар близнецов
Другой подход состоит в определении величины конкордантности фенотипов у пар близнецов
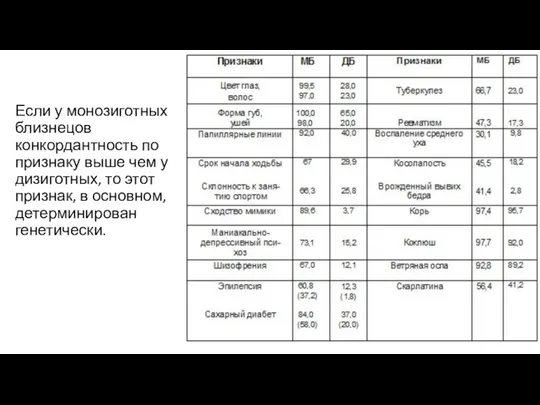
Если у монозиготных близнецов конкордантность по признаку выше чем у дизиготных,
Если у монозиготных близнецов конкордантность по признаку выше чем у дизиготных,

Во многих случаях признаки могут считаться полигенными, но полигенность наследования трудно
Во многих случаях признаки могут считаться полигенными, но полигенность наследования трудно
 Дикие животные
Дикие животные Анатомо-физиологические особенности органа зрения
Анатомо-физиологические особенности органа зрения Естественный отбор
Естественный отбор Строение и функции клетки
Строение и функции клетки Органические вещества клетки – нуклеиновые кислоты ДНК и РНК
Органические вещества клетки – нуклеиновые кислоты ДНК и РНК Қол қаңқасының сүйектер
Қол қаңқасының сүйектер ОПРЕДЕЛЕНИЕ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ МОЕЙ СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ (ССС). УРОК-ПРАКТИКУМ для учащихся 8- классов Учитель Фурса М.Н. ГБОУ гимназия № 209
ОПРЕДЕЛЕНИЕ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ МОЕЙ СЕРДЕЧНО-СОСУДИСТОЙ СИСТЕМЫ (ССС). УРОК-ПРАКТИКУМ для учащихся 8- классов Учитель Фурса М.Н. ГБОУ гимназия № 209 Сущность жизни и свойства живого. Уровни организации живой материи
Сущность жизни и свойства живого. Уровни организации живой материи Особенности двудольных растений. Обобщение игра. 7 класс
Особенности двудольных растений. Обобщение игра. 7 класс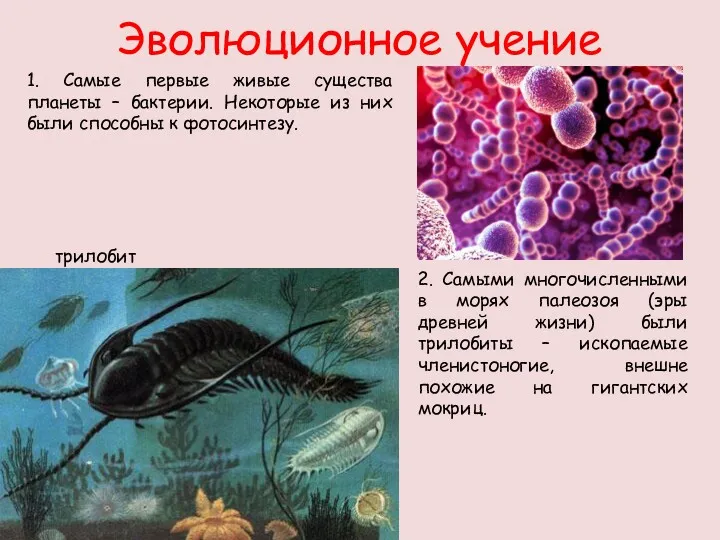 Эволюционное учение
Эволюционное учение Любимые сад и огород. В.Н. Пастарнаков. Цикл: для начинающих. Часть 1. Посадка рассады
Любимые сад и огород. В.Н. Пастарнаков. Цикл: для начинающих. Часть 1. Посадка рассады Биология полорогих: дзерена, сайгака, горала
Биология полорогих: дзерена, сайгака, горала Кормление лошадей
Кормление лошадей Методы микробиологической диагностики вирусных инфекций. Профилактика вирусных инфекций
Методы микробиологической диагностики вирусных инфекций. Профилактика вирусных инфекций Общее учение о нервной системе (неврология). Развитие нервной системы. Спинной мозг
Общее учение о нервной системе (неврология). Развитие нервной системы. Спинной мозг Знакомство с внешним строением цветкового растения
Знакомство с внешним строением цветкового растения Скопа - птица 2018 года
Скопа - птица 2018 года Культура горох посевной
Культура горох посевной Моллюски
Моллюски Селекция и её достижения
Селекция и её достижения Развитие растения. Семя, плод, зародыш, прорастание
Развитие растения. Семя, плод, зародыш, прорастание Кислородсодержащие органические соединения. Органическая химия
Кислородсодержащие органические соединения. Органическая химия Высшие или плацентарные звери
Высшие или плацентарные звери Содержание кислорода в атмосфере
Содержание кислорода в атмосфере Дикие животные
Дикие животные Радиочувствительность тканей, органов, организма. Радиационные синдромы
Радиочувствительность тканей, органов, организма. Радиационные синдромы Презентация урока биологии Насекомые
Презентация урока биологии Насекомые Общий покров. Кожа и её производные
Общий покров. Кожа и её производные