- Гормоны. Регуляция метаболизма

Содержание
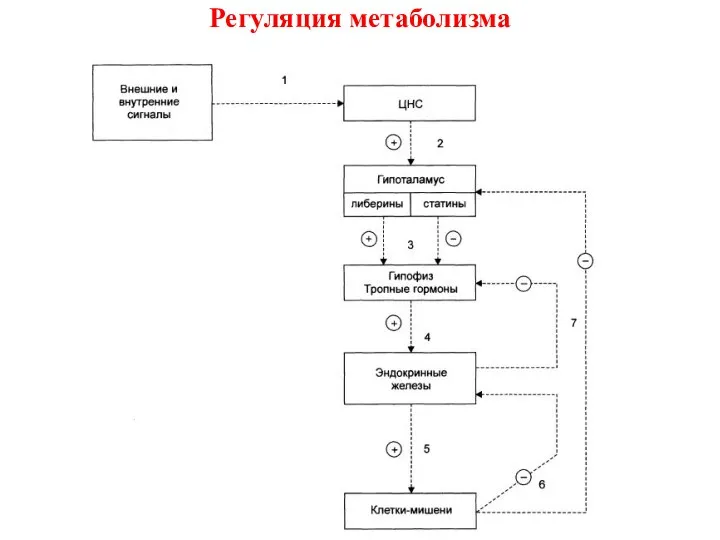
- 2. Регуляция метаболизма Система регуляции обмена веществ и функций организма образуют три иерархических уровня: 1 – ЦНС.
- 3. Регуляция метаболизма Синтез и секреция гормонов стимулируется внешними и внутренними сигналами, поступающими в ЦНС; Эти сигналы
- 4. Регуляция метаболизма
- 5. Регуляция метаболизма Не все эндокринные железы регулируются подобным образом: Гормоны задней доли гипофиза (окситоцин и вазопрессин)
- 6. Гормоны Гормоны – вещества органической природы, вырабатывающиеся в специализированных клетках желез внутренней секреции, поступающие в кровь
- 7. Гормоны 1) Пептидные и белковые гормоны включают: гормоны гипоталамуса и гипофиза (тиролиберин, соматолиберин, соматостатин, гормон роста,
- 8. Гормоны гипоталамуса Гипоталамус - место взаимодействия высших отделов ЦНС и эндокринной системы. В гипоталамусе открыто 7
- 9. Гормоны гипофиза В гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной природы, оказывающих стимулирующий эффект
- 10. Гормоны передней доли гипофиза
- 11. Гормоны задней и средней долей гипофиза Гормоны задней доли гипофиза: Окситоцин у млекопитающих связан со стимуляцией
- 12. Гормоны щитовидной железы Синтезируются гормоны –йодированные производные аминокислоты тирозина. Трийодтиронин и тироксин (тетрайодтиронин). Регулируют скорость основного
- 13. Гормоны поджелудочной железы Поджелудочная железа относится к железам со смешанной секрецией. Панкреатические островки (островки Лангерганса): α-
- 14. Гормоны поджелудочной железы Глюкагон Полипептид. Вызывает увеличение концентрации глюкозы в крови главным образом за счет распада
- 15. Гормоны надпочечников Мозговое вещество вырабатывает гормоны, которые считаются производными аминокислот. Корковое вещество секретирует гормоны стероидной природы.
- 16. Гормоны надпочечников Гормоны коркового вещества надпочечников: Глюкокортикоиды -кортикостероиды, оказывающие влияние на обмен углеводов, белков, жиров и
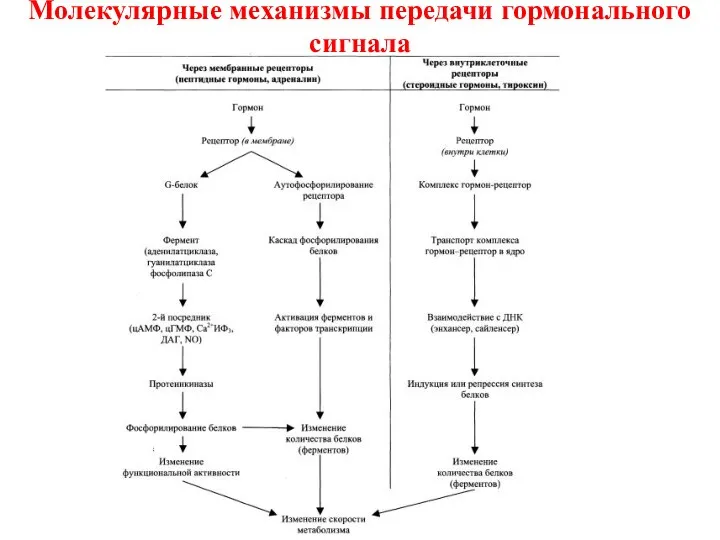
- 17. Молекулярные механизмы передачи гормонального сигнала По механизму действия гормоны можно разделить на 2 группы: Гормоны, взаимодействующие
- 18. Механизмы передачи гормонального сигнала Гормоны, взаимодействующие с клеточными рецепторами, передают сигнал на уровне клетки через вторичные
- 19. Молекулярные механизмы передачи гормонального сигнала
- 20. Молекулярные механизмы передачи гормонального сигнала Аденилатциклазная мессенджерная система: В нем задействовано мимимум пять белков: 1) рецептор
- 21. Молекулярные механизмы передачи гормонального сигнала Аденилатциклазная мессенджерная система: 1) Cвязывание гормона с β-адренергическим рецептором приводит к
- 22. Молекулярные механизмы передачи гормонального сигнала Аденилатциклазная мессенджерная система: 3) Аденилатциклаза представляет собой интегральный белок плазматических мембран,
- 23. Молекулярные механизмы передачи гормонального сигнала Аденилатциклазная мессенджерная система: 4) Протеинкиназа А– это внутриклеточный фермент, через который
- 24. Молекулярные механизмы передачи гормонального сигнала
- 25. Молекулярные механизмы передачи гормонального сигнала Ряд гормонов оказывает тормозящий эффект на аденилатциклазу, соответственно снижая уровень цАМФ
- 26. Молекулярные механизмы передачи гормонального сигнала К внутриклеточной системе мессенджеров относят также производные фосфолипидов мембран эукариотических клеток,
- 27. Молекулярные механизмы передачи гормонального сигнала
- 28. Молекулярные механизмы передачи гормонального сигнала Биологические эффекты этих вторичных мессенджеров реализуютсяпо-разному. Диацилглицерол, как и свободныt ионов
- 30. Скачать презентацию

Регуляция метаболизма
Система регуляции обмена веществ и функций организма образуют три иерархических
Регуляция метаболизма
Система регуляции обмена веществ и функций организма образуют три иерархических

Регуляция метаболизма
Синтез и секреция гормонов стимулируется внешними и внутренними сигналами, поступающими
Регуляция метаболизма
Синтез и секреция гормонов стимулируется внешними и внутренними сигналами, поступающими
Регуляция метаболизма
Регуляция метаболизма

Регуляция метаболизма
Не все эндокринные железы регулируются подобным образом:
Гормоны задней доли гипофиза
Регуляция метаболизма
Не все эндокринные железы регулируются подобным образом:
Гормоны задней доли гипофиза

Гормоны
Гормоны – вещества органической природы, вырабатывающиеся в специализированных клетках желез внутренней
Гормоны
Гормоны – вещества органической природы, вырабатывающиеся в специализированных клетках желез внутренней

Гормоны
1) Пептидные и белковые гормоны включают:
гормоны гипоталамуса и гипофиза (тиролиберин,
Гормоны
1) Пептидные и белковые гормоны включают:
гормоны гипоталамуса и гипофиза (тиролиберин,

Гормоны гипоталамуса
Гипоталамус - место взаимодействия высших отделов ЦНС и эндокринной системы.
В
Гормоны гипоталамуса
Гипоталамус - место взаимодействия высших отделов ЦНС и эндокринной системы.
В

Гормоны гипофиза
В гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной
Гормоны гипофиза
В гипофизе синтезируется ряд биологически активных гормонов белковой и пептидной

Гормоны передней доли гипофиза
Гормоны передней доли гипофиза

Гормоны задней и средней долей гипофиза
Гормоны задней доли гипофиза:
Окситоцин у млекопитающих
Гормоны задней и средней долей гипофиза
Гормоны задней доли гипофиза:
Окситоцин у млекопитающих

Гормоны щитовидной железы
Синтезируются гормоны –йодированные производные аминокислоты тирозина.
Трийодтиронин и тироксин (тетрайодтиронин).
Регулируют
Гормоны щитовидной железы
Синтезируются гормоны –йодированные производные аминокислоты тирозина.
Трийодтиронин и тироксин (тетрайодтиронин).
Регулируют

Гормоны поджелудочной железы
Поджелудочная железа относится к железам со смешанной секрецией.
Панкреатические островки
Гормоны поджелудочной железы
Поджелудочная железа относится к железам со смешанной секрецией.
Панкреатические островки

Гормоны поджелудочной железы
Глюкагон
Полипептид.
Вызывает увеличение концентрации глюкозы в крови главным образом за
Гормоны поджелудочной железы
Глюкагон
Полипептид.
Вызывает увеличение концентрации глюкозы в крови главным образом за

Гормоны надпочечников
Мозговое вещество вырабатывает гормоны, которые считаются производными аминокислот.
Корковое вещество
Гормоны надпочечников
Мозговое вещество вырабатывает гормоны, которые считаются производными аминокислот.
Корковое вещество

Гормоны надпочечников
Гормоны коркового вещества надпочечников:
Глюкокортикоиды -кортикостероиды, оказывающие влияние на обмен углеводов,
Гормоны надпочечников
Гормоны коркового вещества надпочечников:
Глюкокортикоиды -кортикостероиды, оказывающие влияние на обмен углеводов,

Молекулярные механизмы передачи гормонального сигнала
По механизму действия гормоны можно разделить на
Молекулярные механизмы передачи гормонального сигнала
По механизму действия гормоны можно разделить на

Механизмы передачи гормонального сигнала
Гормоны, взаимодействующие с клеточными рецепторами, передают сигнал на
Механизмы передачи гормонального сигнала
Гормоны, взаимодействующие с клеточными рецепторами, передают сигнал на
Молекулярные механизмы передачи гормонального сигнала
Молекулярные механизмы передачи гормонального сигнала

Молекулярные механизмы передачи гормонального сигнала
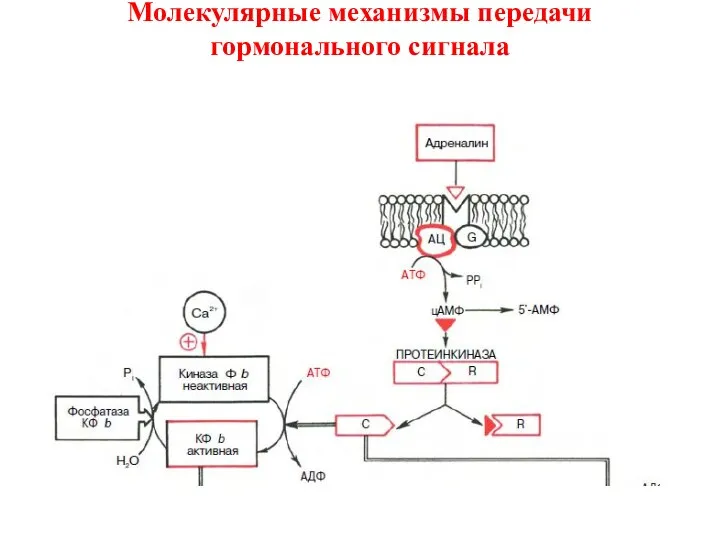
Аденилатциклазная мессенджерная система:
В нем задействовано мимимум пять
Молекулярные механизмы передачи гормонального сигнала
Аденилатциклазная мессенджерная система:
В нем задействовано мимимум пять

Молекулярные механизмы передачи гормонального сигнала
Аденилатциклазная мессенджерная система:
1) Cвязывание гормона с β-адренергическим
Молекулярные механизмы передачи гормонального сигнала
Аденилатциклазная мессенджерная система:
1) Cвязывание гормона с β-адренергическим
Молекулярные механизмы передачи гормонального сигнала
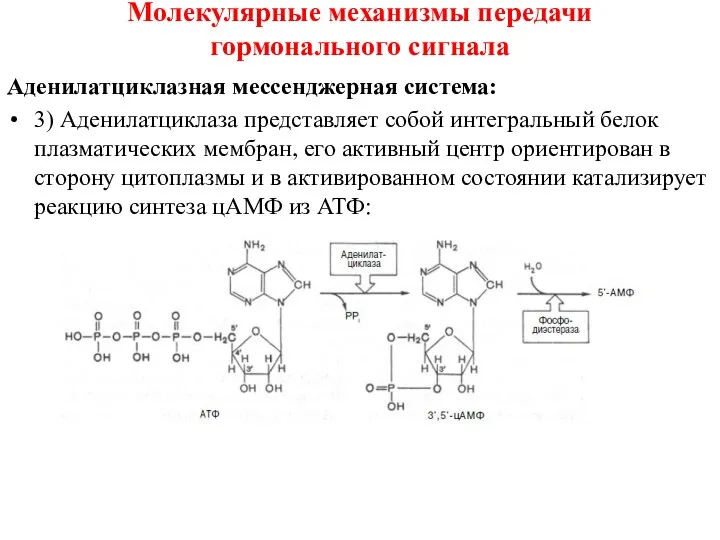
Аденилатциклазная мессенджерная система:
3) Аденилатциклаза представляет собой интегральный
Молекулярные механизмы передачи гормонального сигнала
Аденилатциклазная мессенджерная система:
3) Аденилатциклаза представляет собой интегральный

Молекулярные механизмы передачи гормонального сигнала
Аденилатциклазная мессенджерная система:
4) Протеинкиназа А– это внутриклеточный
Молекулярные механизмы передачи гормонального сигнала
Аденилатциклазная мессенджерная система:
4) Протеинкиназа А– это внутриклеточный
Молекулярные механизмы передачи гормонального сигнала
Молекулярные механизмы передачи гормонального сигнала

Молекулярные механизмы передачи гормонального сигнала
Ряд гормонов оказывает тормозящий эффект на аденилатциклазу,
Молекулярные механизмы передачи гормонального сигнала
Ряд гормонов оказывает тормозящий эффект на аденилатциклазу,

Молекулярные механизмы передачи гормонального сигнала
К внутриклеточной системе мессенджеров относят также производные
Молекулярные механизмы передачи гормонального сигнала
К внутриклеточной системе мессенджеров относят также производные
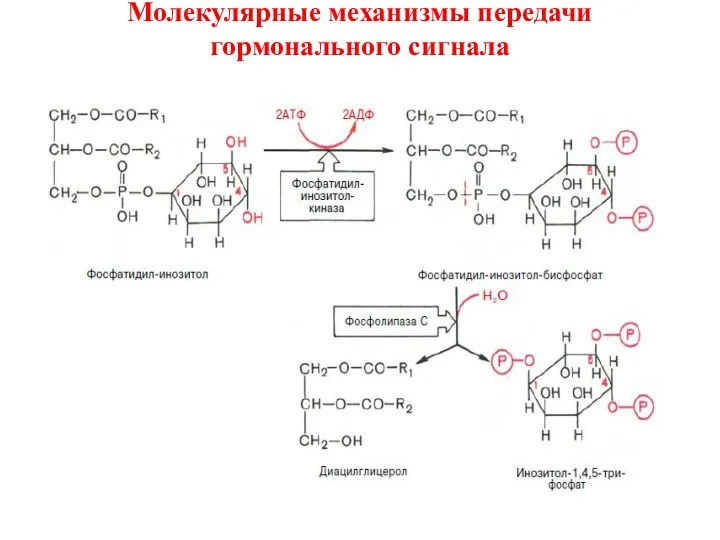
Молекулярные механизмы передачи гормонального сигнала
Молекулярные механизмы передачи гормонального сигнала

Молекулярные механизмы передачи гормонального сигнала
Биологические эффекты этих вторичных мессенджеров реализуютсяпо-разному.
Диацилглицерол,
Молекулярные механизмы передачи гормонального сигнала
Биологические эффекты этих вторичных мессенджеров реализуютсяпо-разному.
Диацилглицерол,
 Снег как индикатор окружающей среды. Диск
Снег как индикатор окружающей среды. Диск Жизнь луга
Жизнь луга Протерозойская эра
Протерозойская эра Причини вимирання динозаврів
Причини вимирання динозаврів Барабанна перетинка: будова та функції
Барабанна перетинка: будова та функції Строение клетки. Растительная, животная. Цитология
Строение клетки. Растительная, животная. Цитология Химический состав клетки. Неорганические вещества
Химический состав клетки. Неорганические вещества Вид. Критерии вида
Вид. Критерии вида Вегетативное размножение растений
Вегетативное размножение растений Введение в биологию развития. Основные закономерности эмбрионального развития. Периодизация постэмбрионального периода
Введение в биологию развития. Основные закономерности эмбрионального развития. Периодизация постэмбрионального периода ГИА_Молекулярная биология. Химический состав клетки_Назарченко И.В
ГИА_Молекулярная биология. Химический состав клетки_Назарченко И.В Тип Кольчатые черви
Тип Кольчатые черви Белый медведь
Белый медведь Мейоз. Механизм мейоза
Мейоз. Механизм мейоза Презентация к уроку биологии 8 класс по теме Витамины
Презентация к уроку биологии 8 класс по теме Витамины Пропорции выражают характер. Птицы с разным характером пропорций
Пропорции выражают характер. Птицы с разным характером пропорций Основные понятия генетики
Основные понятия генетики Экологические факторы
Экологические факторы Мейоз. Черты отличия митоза и мейоза
Мейоз. Черты отличия митоза и мейоза fМуравьи - какие они?
fМуравьи - какие они? Обмен веществ и энергии. Терморегуляция
Обмен веществ и энергии. Терморегуляция Происхождение растений. Основные этапы развития растительного мира
Происхождение растений. Основные этапы развития растительного мира Строение бактериальной клетки
Строение бактериальной клетки Биологическое окисление при участии кислорода
Биологическое окисление при участии кислорода Нуклеиновые кислоты. Строение и функции
Нуклеиновые кислоты. Строение и функции Углеводы: структура и функции
Углеводы: структура и функции Развитие генетики
Развитие генетики Птицы наши друзья и соседи
Птицы наши друзья и соседи