- Химизм фотосинтеза

Содержание
- 2. I. История изучения 1779 г. - Ингенхауз показал значение света в процессе исправления испорченного воздуха 1772
- 3. Работы К.А.Тимирязева Выводы К.А.Тимирязева СПХ= СДФ 2 max хлорофилла = 2 max фотосинтеза. Фотосинтез в красных
- 4. Характеристика участков спектра
- 5. Работы К.А. Тимирязева Роль работ К.А. Тимирязева: 1) Заложил основы изучения энергетики фотосинтеза. 2) Экспериментально доказал,
- 6. II. Открытия, лежащие в основе современной схемы фотосинтеза Происхождение кислорода при фотосинтезе. Открытие световых и темновых
- 7. Происхождение кислорода при фотосинтезе Изучение бактериального фотосинтеза. Опыты Т. В. Энгельмана (1883), К.Б. Ван-Ниля (1937- 41);
- 8. Опыты Р. Хилла (1937) Реакция Хилла: процесс разложения воды хлоропластами с выделением кислорода в присутствии акцепторов
- 9. Происхождение кислорода при фотосинтезе Исследования с радиоактивными изотопами С. Рубен и М. Камен - 1940 г.
- 10. Открытие световых и темновых реакций Опыты Ф. Блэкмэна (1905) по зависимости фотосинтеза от температуры Опыты с
- 11. Открытие фотосинтетического фосфорилирования 1954 г. - Д. Арнон, М. Аллен и Ф. Уотли НAДФ+ (хлоропласты, свет)
- 12. Открытие двух фотосистем Эффект Р. Эмерсона (1957). Σ ИФ680-700+ИФ650-660>>ИФ680-700+ ИФ650-660 Фотосистема: светособирающий (пигментный, антенный) комплекс (ССК)
- 13. Фотосистемы I и II Фотосистема I (ФС I): ССК +ФРЦ I (П700)+молекулы переносчики П700 - димер
- 14. Распределение фотосистем а-схематическое изображение хлоропласта в разрезе б и в -распределение фотосистемы I и фотосистемы II
- 15. III. Современная схема фотосинтеза
- 16. Световые и темновые реакции Световые реакции Совокупность процессов, в результате которых за счет энергии света синтезируется
- 17. IV. Световые реакции фотосинтеза Фотофизический этап фотосинтеза Реакции поглощения энергии пигментами, запасания ее в виде электронного
- 18. Энергетические состояния молекулы хлорофилла
- 20. So - основное энергетическое (синглетное) состояние: молекула находится в тепловом равновесии со средой, все электроны попарно
- 21. Фотосистема: ССК + ФРЦ + молекулы – переносчики Фотосистема I (ФС I): ССК +ФРЦ I (П700)+молекулы
- 22. Схема миграции энергии возбуждения по пигментной матрице Пигменты- сборщики света Пигменты-ловушки
- 23. Назовите, какие факторы влияют на строение ССК. У каких растений больше размер ССК: -теневыносливых или растущих
- 24. Передача энергии между молекулами пигментов Резонансный путь Каротин(400-550) →хлорофилл b(650) → хлорофилл a (660-675) → П700
- 25. Фотофизический этап фотосинтеза 1. Возбуждение молекулы хлорофилла: Хл + hv → Хл* 2. Передача энергии возбужденного
- 26. Фотохимический этап фотосинтеза
- 27. Первичное разделение зарядов хлорофилл→ хлорофилл * → ē + Хл +
- 30. Расположение комплексов в тилакоидной мембране
- 31. Путь переноса ē от фотосистемы II к фотосистеме I П680 →феофитин→пластохиноны → → b/f-комплекс (цитb →FeS→цитf)
- 32. Фотоокисление воды 2Н2О + 4 hν → 4Н+ + 4 ē + О2 2Mn 4+ +
- 33. Схема строения цепи электронного транспорта в хлоропластах
- 34. Пластохинон Окислительно-восстановительные превращения пластохинона Предполагаемая схема расположения пластохинона в мембране РХ + 2 ē → РХ-2
- 35. Фотофосфорилирование Циклическое АДФ + Н3РО4 + hv → АТФ + Н2О. Нециклическое 2 НАДФ + 2Н2О
- 36. Циклическое фотофосфорилирование
- 37. Нециклическое фотофосфорилирование
- 38. Фотосинтетическое фосфорилирование
- 39. Хемиосмотический механизм синтеза АТФ Хемиосмотическая теория разработана в 1961-1966 гг. В 1967 г. Андреа Ягендорф использовал
- 40. Сущность теории Митчелла Фосфорилирование происходит на мембранах, которые непроницаемы для Н+ и имеют ЭТЦ. Энергия, высвобождаемая
- 41. Хемиосмотическое сопряжение как механизм фосфорилирования
- 42. Свет→ протонный потенциал (∆μΗ+) → АТФ
- 44. АТРсинтазный комплекс СF0 – интегральный гидрофобный белок мембраны, протонный канал, 4 типа пептидов (a, b, b',
- 45. АТФсинтаза
- 46. Энергозависимые изменения состояния каталитических центров Три этапа синтеза АТФ : Присоединение АДФ и Фн к активному
- 47. Модель АТФсинтазы P. Boyer, J. Walker Нобелевская премия, 1997 Ротационный механизм работы ферментного комплекса: Энергия, освобождаемая
- 48. Схема, иллюстрирующая вращение γ субъединицы АТФсинтазы
- 50. Скачать презентацию

I. История изучения
1779 г. - Ингенхауз показал значение света в
I. История изучения
1779 г. - Ингенхауз показал значение света в
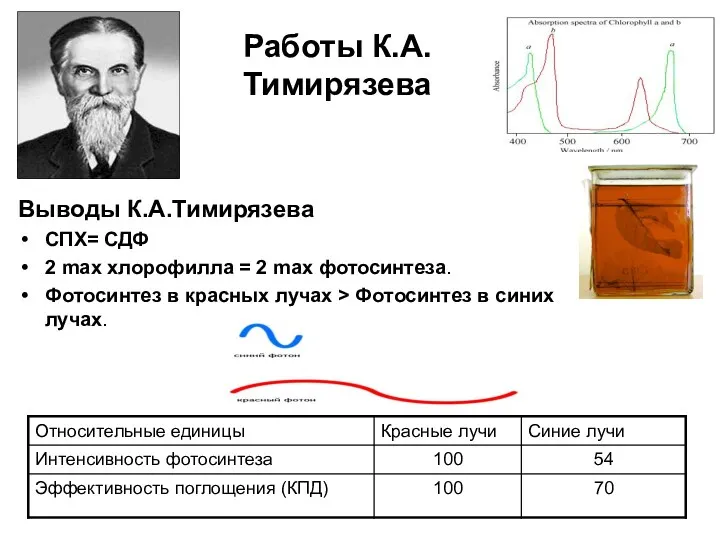
Работы К.А.Тимирязева
Выводы К.А.Тимирязева
СПХ= СДФ
2 max хлорофилла = 2 max фотосинтеза.
Фотосинтез
Работы К.А.Тимирязева
Выводы К.А.Тимирязева
СПХ= СДФ
2 max хлорофилла = 2 max фотосинтеза.
Фотосинтез
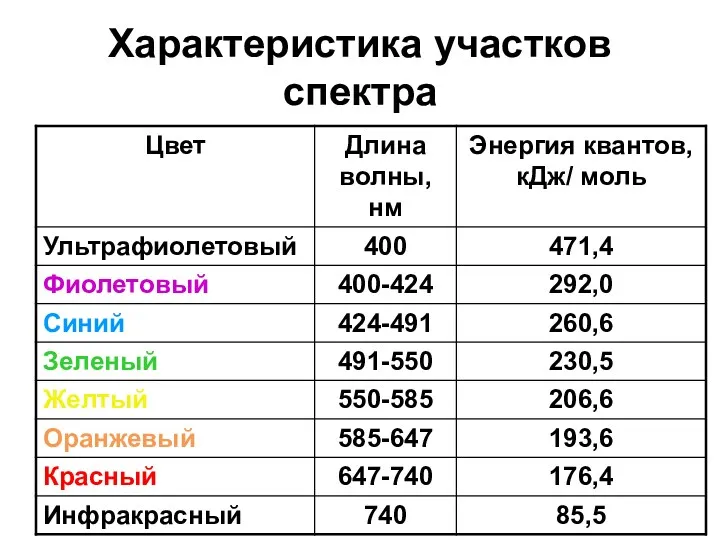
Характеристика участков спектра
Характеристика участков спектра

Работы К.А. Тимирязева
Роль работ К.А. Тимирязева:
1) Заложил основы изучения энергетики фотосинтеза.
2)
Работы К.А. Тимирязева
Роль работ К.А. Тимирязева:
1) Заложил основы изучения энергетики фотосинтеза.
2)

II. Открытия, лежащие в основе современной схемы фотосинтеза
Происхождение кислорода при фотосинтезе.
Открытие
II. Открытия, лежащие в основе современной схемы фотосинтеза
Происхождение кислорода при фотосинтезе.
Открытие

Происхождение кислорода при фотосинтезе
Изучение бактериального фотосинтеза.
Опыты Т. В. Энгельмана (1883),
Происхождение кислорода при фотосинтезе
Изучение бактериального фотосинтеза.
Опыты Т. В. Энгельмана (1883),
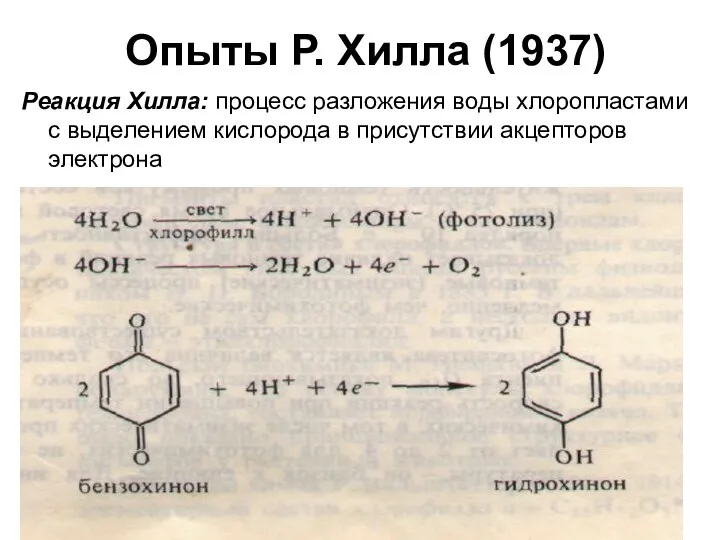
Опыты Р. Хилла (1937)
Реакция Хилла: процесс разложения воды хлоропластами с выделением
Опыты Р. Хилла (1937)
Реакция Хилла: процесс разложения воды хлоропластами с выделением
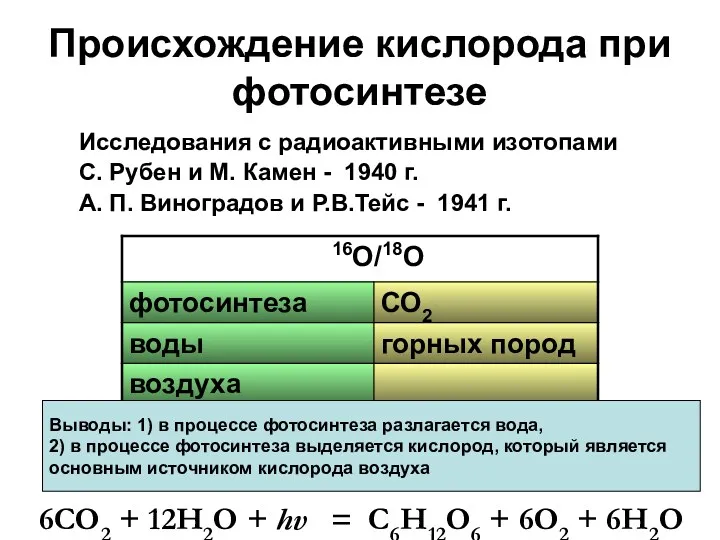
Происхождение кислорода при фотосинтезе
Исследования с радиоактивными изотопами
С. Рубен и М. Камен
Происхождение кислорода при фотосинтезе
Исследования с радиоактивными изотопами
С. Рубен и М. Камен

Открытие световых и темновых реакций
Опыты Ф. Блэкмэна (1905) по зависимости фотосинтеза
Открытие световых и темновых реакций
Опыты Ф. Блэкмэна (1905) по зависимости фотосинтеза

Открытие фотосинтетического фосфорилирования
1954 г. - Д. Арнон, М. Аллен и
Открытие фотосинтетического фосфорилирования
1954 г. - Д. Арнон, М. Аллен и

Открытие двух фотосистем
Эффект Р. Эмерсона (1957).
Σ ИФ680-700+ИФ650-660>>ИФ680-700+ ИФ650-660
Фотосистема: светособирающий (пигментный,
Открытие двух фотосистем
Эффект Р. Эмерсона (1957).
Σ ИФ680-700+ИФ650-660>>ИФ680-700+ ИФ650-660
Фотосистема: светособирающий (пигментный,

Фотосистемы I и II
Фотосистема I (ФС I):
ССК +ФРЦ I (П700)+молекулы
Фотосистемы I и II
Фотосистема I (ФС I):
ССК +ФРЦ I (П700)+молекулы
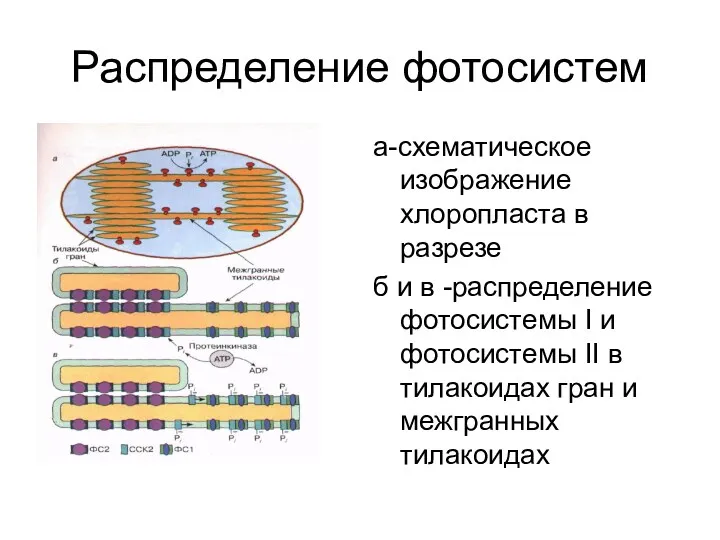
Распределение фотосистем
а-схематическое изображение хлоропласта в разрезе
б и в -распределение фотосистемы I
Распределение фотосистем
а-схематическое изображение хлоропласта в разрезе
б и в -распределение фотосистемы I

III. Современная схема фотосинтеза
III. Современная схема фотосинтеза

Световые и темновые реакции
Световые реакции
Совокупность процессов, в результате которых за
Световые и темновые реакции
Световые реакции
Совокупность процессов, в результате которых за

IV. Световые реакции фотосинтеза
Фотофизический этап фотосинтеза
Реакции поглощения энергии пигментами, запасания
IV. Световые реакции фотосинтеза
Фотофизический этап фотосинтеза
Реакции поглощения энергии пигментами, запасания
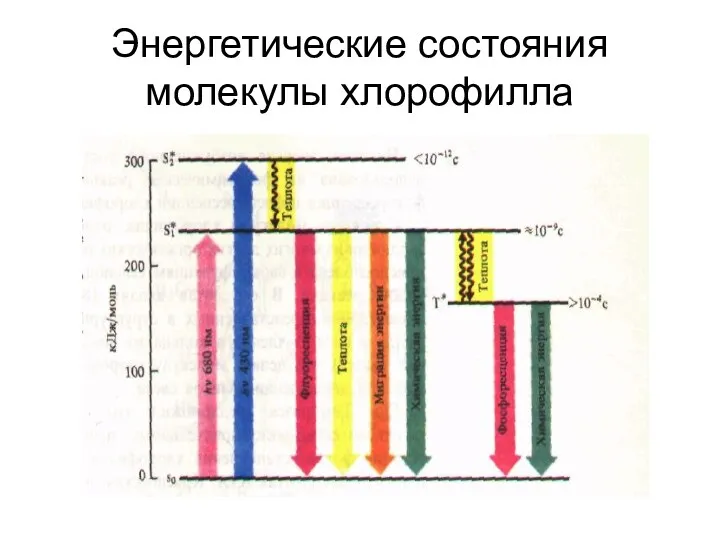
Энергетические состояния молекулы хлорофилла
Энергетические состояния молекулы хлорофилла
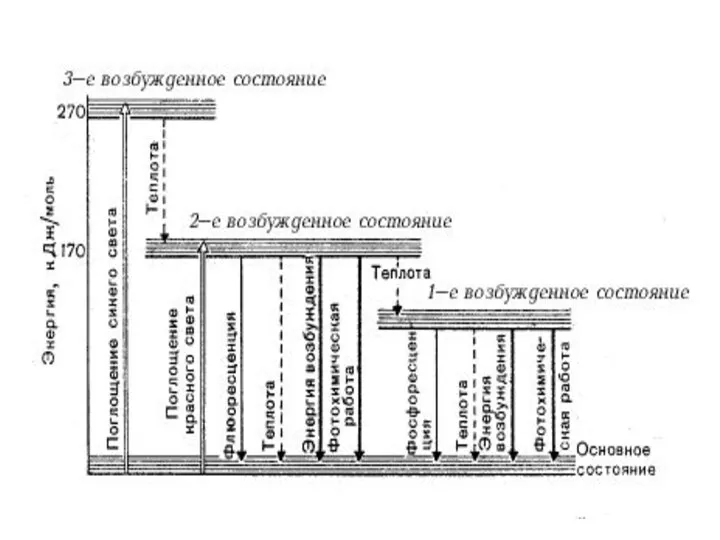

So - основное энергетическое (синглетное) состояние: молекула находится в тепловом равновесии
So - основное энергетическое (синглетное) состояние: молекула находится в тепловом равновесии

Фотосистема:
ССК + ФРЦ + молекулы – переносчики
Фотосистема I (ФС I):
Фотосистема:
ССК + ФРЦ + молекулы – переносчики
Фотосистема I (ФС I):
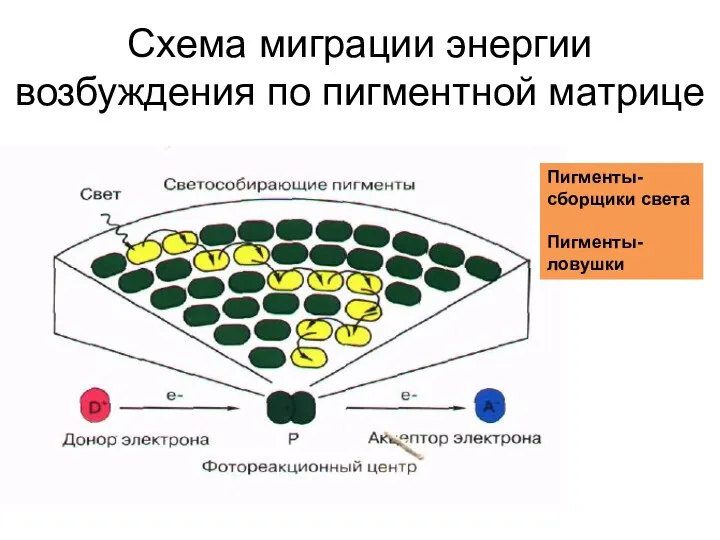
Схема миграции энергии возбуждения по пигментной матрице
Пигменты- сборщики света
Пигменты-ловушки
Схема миграции энергии возбуждения по пигментной матрице
Пигменты- сборщики света
Пигменты-ловушки

Назовите, какие факторы влияют на строение ССК.
У каких растений больше размер
Назовите, какие факторы влияют на строение ССК.
У каких растений больше размер

Передача энергии между молекулами пигментов
Резонансный путь
Каротин(400-550) →хлорофилл b(650) → хлорофилл
Передача энергии между молекулами пигментов
Резонансный путь
Каротин(400-550) →хлорофилл b(650) → хлорофилл

Фотофизический этап фотосинтеза
1. Возбуждение молекулы хлорофилла:
Хл + hv → Хл*
2.
Фотофизический этап фотосинтеза
1. Возбуждение молекулы хлорофилла:
Хл + hv → Хл*
2.
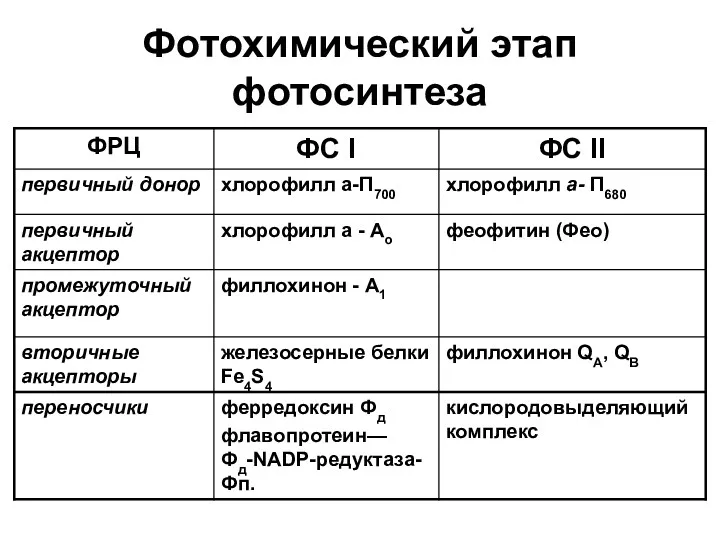
Фотохимический этап фотосинтеза
Фотохимический этап фотосинтеза

Первичное разделение зарядов
хлорофилл→ хлорофилл * → ē + Хл +
Первичное разделение зарядов
хлорофилл→ хлорофилл * → ē + Хл +
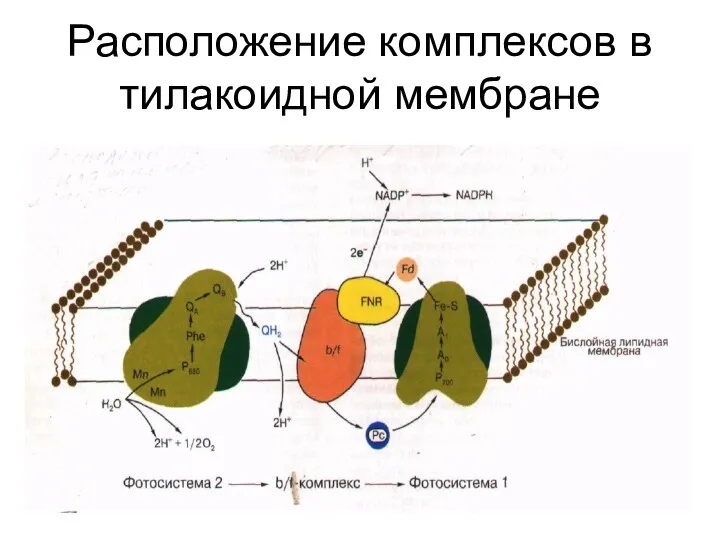
Расположение комплексов в тилакоидной мембране
Расположение комплексов в тилакоидной мембране

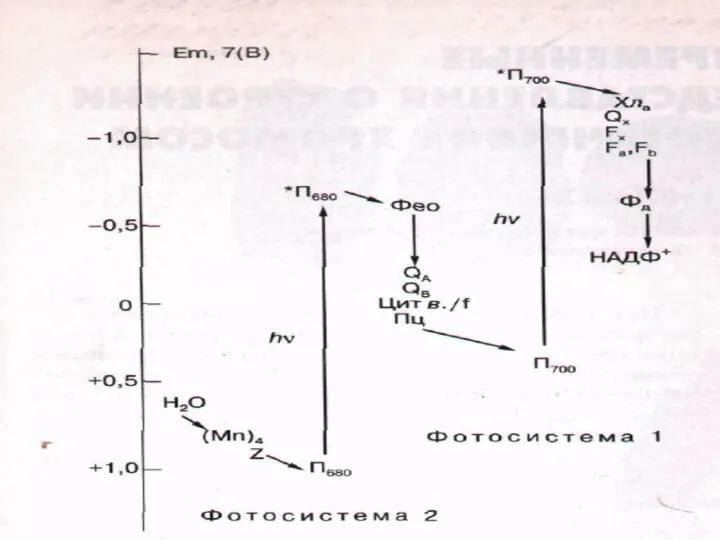
Путь переноса ē от фотосистемы II к фотосистеме I
П680 →феофитин→пластохиноны →
Путь переноса ē от фотосистемы II к фотосистеме I
П680 →феофитин→пластохиноны →
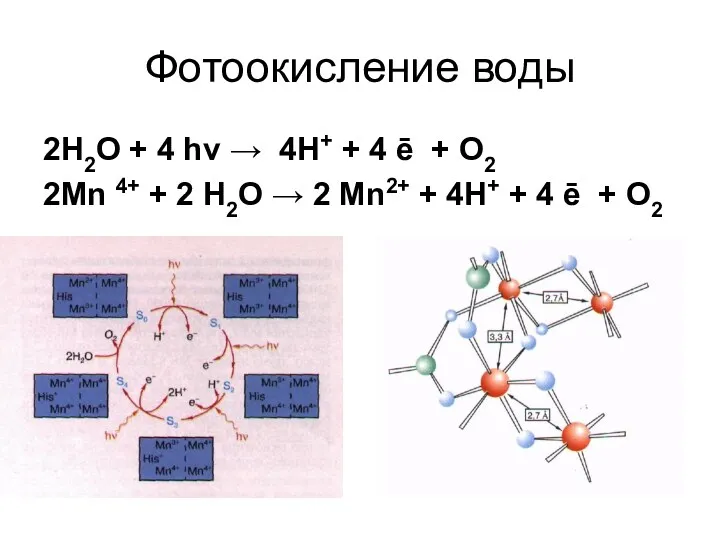
Фотоокисление воды
2Н2О + 4 hν → 4Н+ + 4 ē
Фотоокисление воды
2Н2О + 4 hν → 4Н+ + 4 ē
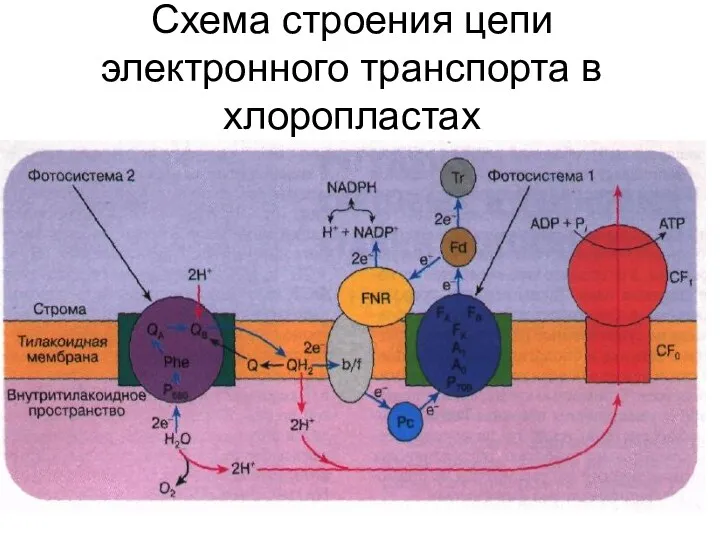
Схема строения цепи электронного транспорта в хлоропластах
Схема строения цепи электронного транспорта в хлоропластах
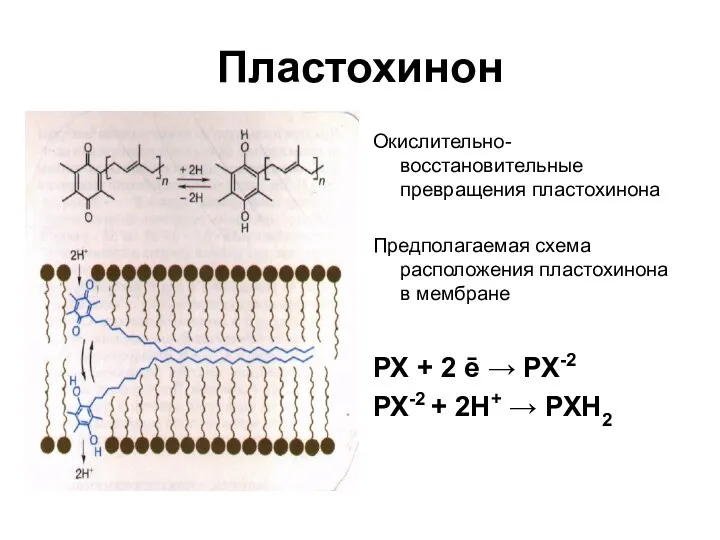
Пластохинон
Окислительно-восстановительные превращения пластохинона
Предполагаемая схема расположения пластохинона в мембране
РХ + 2
Пластохинон
Окислительно-восстановительные превращения пластохинона
Предполагаемая схема расположения пластохинона в мембране
РХ + 2

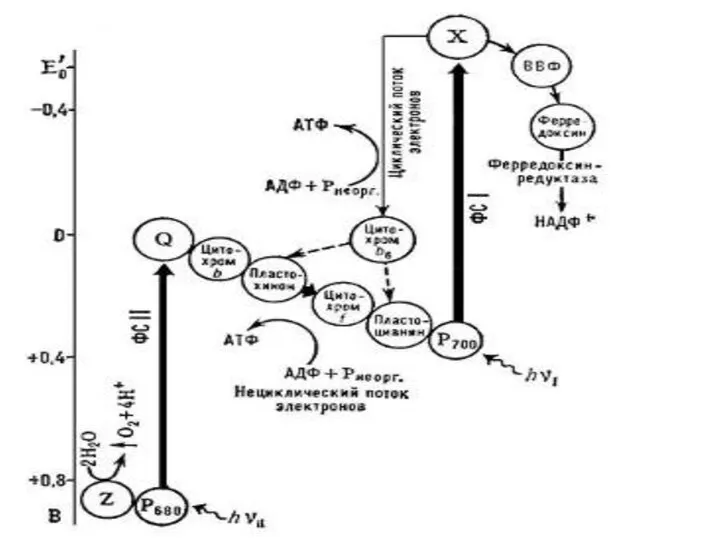
Фотофосфорилирование
Циклическое
АДФ + Н3РО4 + hv → АТФ + Н2О.
Нециклическое
2 НАДФ
Фотофосфорилирование
Циклическое
АДФ + Н3РО4 + hv → АТФ + Н2О.
Нециклическое
2 НАДФ
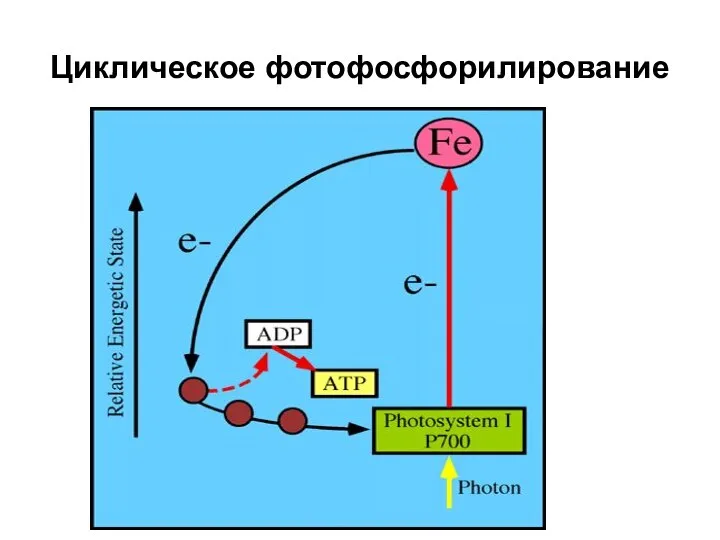
Циклическое фотофосфорилирование
Циклическое фотофосфорилирование
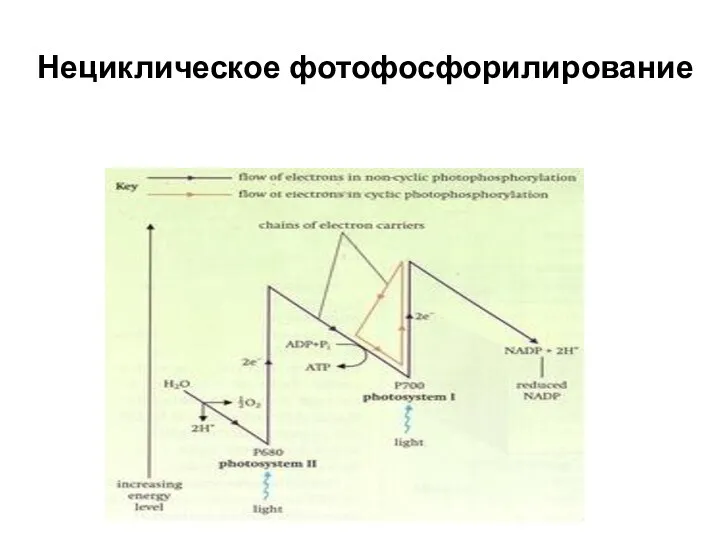
Нециклическое фотофосфорилирование
Нециклическое фотофосфорилирование

Фотосинтетическое фосфорилирование
Фотосинтетическое фосфорилирование

Хемиосмотический механизм синтеза АТФ
Хемиосмотическая теория разработана в 1961-1966 гг.
В
Хемиосмотический механизм синтеза АТФ
Хемиосмотическая теория разработана в 1961-1966 гг.
В

Сущность теории Митчелла
Фосфорилирование происходит на мембранах, которые непроницаемы для Н+
Сущность теории Митчелла
Фосфорилирование происходит на мембранах, которые непроницаемы для Н+
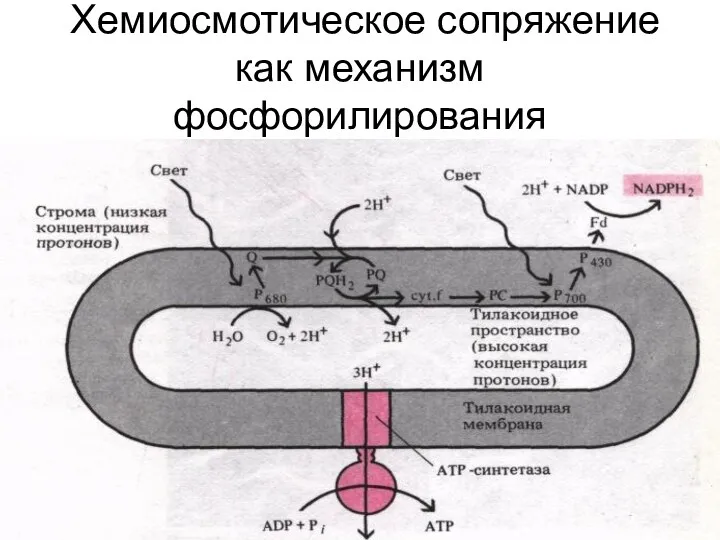
Хемиосмотическое сопряжение как механизм фосфорилирования
Хемиосмотическое сопряжение как механизм фосфорилирования
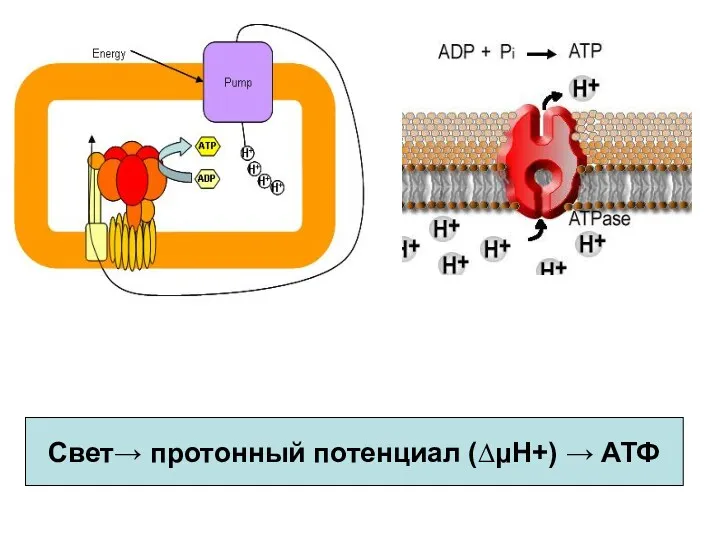
Свет→ протонный потенциал (∆μΗ+) → АТФ
Свет→ протонный потенциал (∆μΗ+) → АТФ
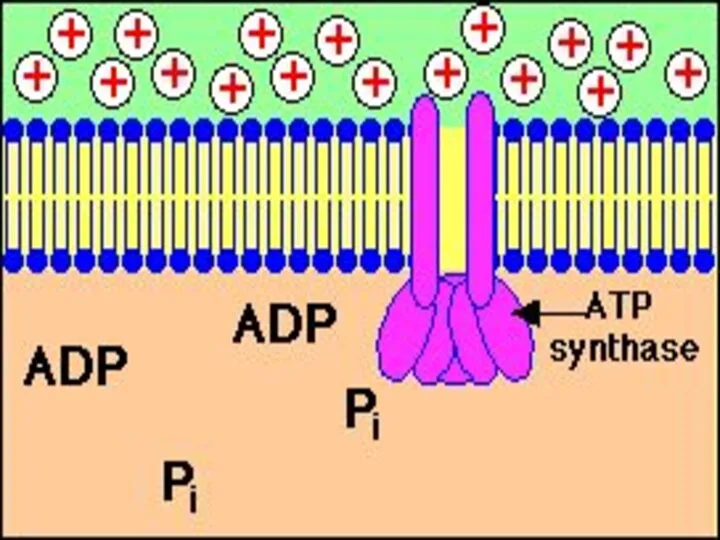
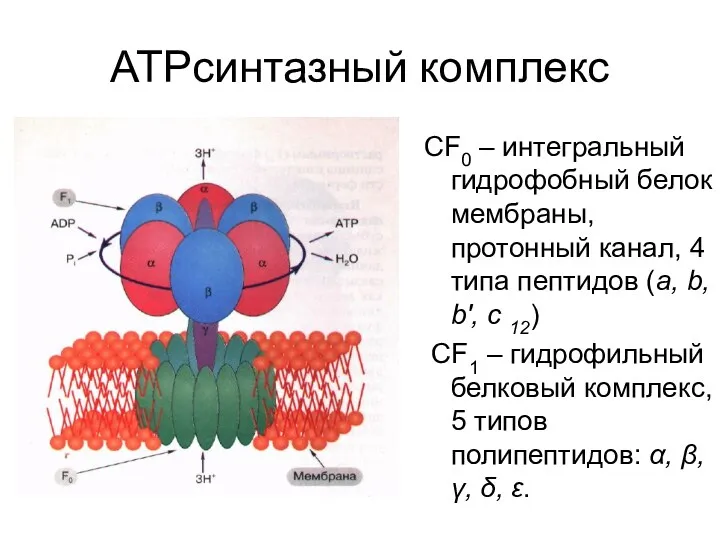
АТРсинтазный комплекс
СF0 – интегральный гидрофобный белок мембраны, протонный канал, 4 типа
АТРсинтазный комплекс
СF0 – интегральный гидрофобный белок мембраны, протонный канал, 4 типа
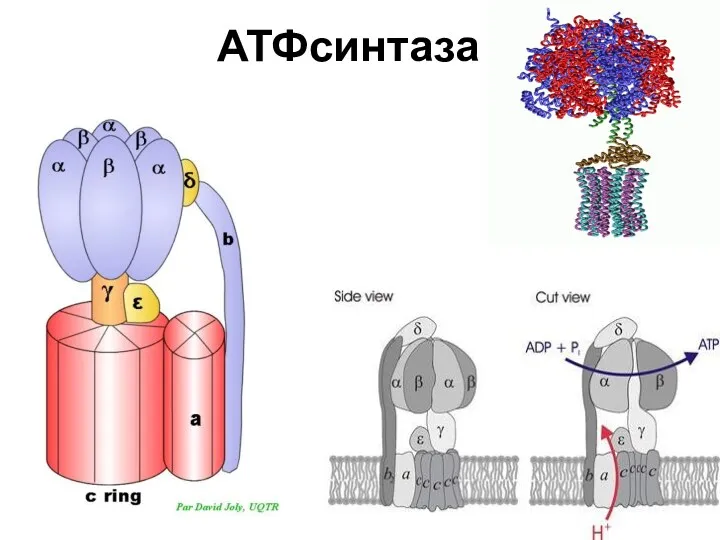
АТФсинтаза
АТФсинтаза

Энергозависимые изменения состояния каталитических центров
Три этапа синтеза АТФ :
Присоединение АДФ и
Энергозависимые изменения состояния каталитических центров
Три этапа синтеза АТФ :
Присоединение АДФ и

Модель АТФсинтазы
P. Boyer, J. Walker
Нобелевская премия, 1997
Ротационный механизм работы ферментного комплекса:
Энергия,
Модель АТФсинтазы
P. Boyer, J. Walker
Нобелевская премия, 1997
Ротационный механизм работы ферментного комплекса:
Энергия,
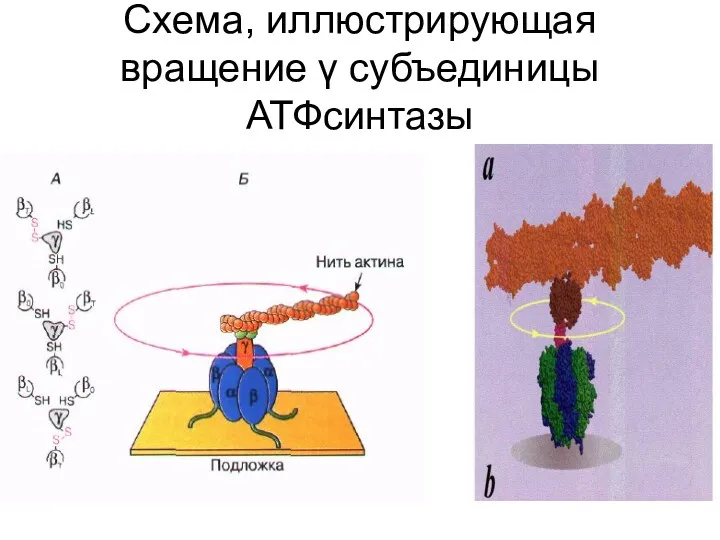
Схема, иллюстрирующая вращение γ субъединицы АТФсинтазы
Схема, иллюстрирующая вращение γ субъединицы АТФсинтазы
 Строение периферического отдела: скелет верхних и нижних конечностей
Строение периферического отдела: скелет верхних и нижних конечностей Жүйке жасушасының құрылысы
Жүйке жасушасының құрылысы Кроссворд Газообмен
Кроссворд Газообмен Спинний мозок. Будова і функції
Спинний мозок. Будова і функції Проверка знаний по теме Передвижение веществ в растении
Проверка знаний по теме Передвижение веществ в растении Пингвины. Виды пингвинов
Пингвины. Виды пингвинов Синапсы
Синапсы Обрезка и формировка деревьев и кустарников
Обрезка и формировка деревьев и кустарников Электрқозушылық түсінігі. Биопотенциалдар. Кардиомиоцит пен жүйке талшығының әсер потенциалы
Электрқозушылық түсінігі. Биопотенциалдар. Кардиомиоцит пен жүйке талшығының әсер потенциалы Эволюция. Доказательства эволюции
Эволюция. Доказательства эволюции Обмен веществ. Нормы и режим питания
Обмен веществ. Нормы и режим питания Органы пищеварения и обмен веществ
Органы пищеварения и обмен веществ Удивительные овощи. Фото-отчет по выставке
Удивительные овощи. Фото-отчет по выставке Growth and Development
Growth and Development Биосинтез холестерина, жирных кислот. Липопротеины. Регуляция и патология липидного обмена
Биосинтез холестерина, жирных кислот. Липопротеины. Регуляция и патология липидного обмена Углеводы. Состав. Классификация
Углеводы. Состав. Классификация Плоды, семена
Плоды, семена Интегрированный урок по биологии-химии Лекарства и их влияние на организм человека
Интегрированный урок по биологии-химии Лекарства и их влияние на организм человека Особенности Подцарства Простейших. Инфузория туфелька и эвглена зелёная
Особенности Подцарства Простейших. Инфузория туфелька и эвглена зелёная Мезозойская эра
Мезозойская эра ДЕРАТИЗАЦИЯ
ДЕРАТИЗАЦИЯ Генетически модифицированные организмы (ГМО)
Генетически модифицированные организмы (ГМО) Презентация о воде
Презентация о воде Разнообразие организмов и их классификация
Разнообразие организмов и их классификация Шеміршекті балықтар класы
Шеміршекті балықтар класы Путешествие в царство цветов и бабочек. Почему нельзя рвать цветы и ловить бабочек
Путешествие в царство цветов и бабочек. Почему нельзя рвать цветы и ловить бабочек Причины разнообразия жизни на Земле
Причины разнообразия жизни на Земле Скалярия обыкновенная
Скалярия обыкновенная