- Инженерия антител

Содержание
- 2. Генно-инженерная технология революционизировала иммунологию, сформировав направление, получившее название "инженерия антител", целью которого является получение на основе
- 3. Сайт-направленный мутагенез. Синтезируют пару праймеров, несущих мутацию, и пару праймеров, комплементарных концам нужного фрагмента ДНК. В
- 4. Следующим этапом развития гибридомной технологии после получения гибридом стало получение " квадром " - клеточных линий,
- 5. В последние годы применение моноклональных антител в научно-исследовательской и клинической практике сильно возросло, во многом благодаря
- 6. Трудности использования моноклональных антител в медицине. Первая и наиглавнейшая из них – «анти-мышиный» антительный ответ человеческой
- 7. Химерные антитела первого поколения. Ген вариабельного домена мышиного антитела клонируется и экспрессируется в системе вместе с
- 8. 2. Гиперхимерные антитела второго поколения. В этом подходе используются лишь минимальные фрагменты мышиного антитела, которые определяют
- 9. 3. Получение биспецифических гуманизированных антител. Биспецифичные антитела получают из моноклональных антител, специфичных к совершенно разным антигенам.
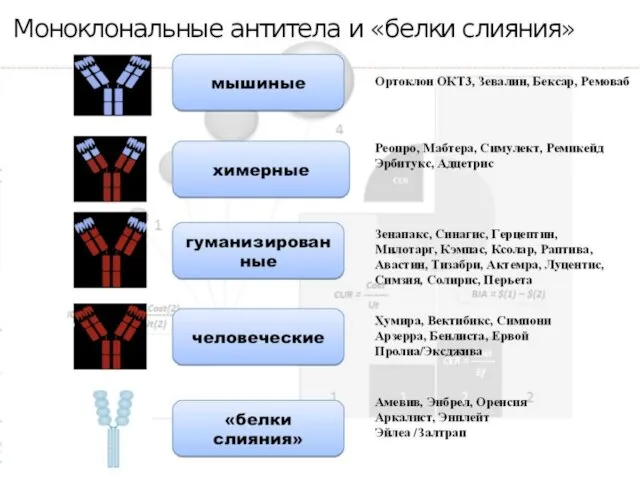
- 12. Ортоклон, Муромонаб-CD3 Мононуклеальные антитела против CD3 лимфоцитов человека, обладают иммунодепрессивными свойствами в отношении антигена Т3 (CD3)
- 13. Этанерцепт — препарат, сочетающий в своей структуре два белка— препарат, сочетающий в своей структуре два белка,
- 14. Другим перспективным подходом для решения проблем, обусловленных применением мышиных антител в организме человека может стать использование
- 15. Fv-фрагменты - нековалентно ассоциированные гетеродимеры V(H) и V(L) -доменов. Они менее стабильны, чем sc-Fv-фрагменты. Промежуточное положение
- 16. Технология получения рекомбинантных фаговых антител. Fab- и Fv-фрагменты антител имеют те же характеристики связывания с антигеном,
- 17. В 1990 году McCafferty и соавт. предложили методику, с помощью которой удалось добиться экспрессии scFv-фрагментов на
- 18. Часто используют ген белка рVIII, однако из-за того, что этот белок присутствует на поверхности фага в
- 19. Используют такие компоненты антител в качестве лабораторных реагентов, заменяющих обычные антитела, например, в проточной цитометрии или
- 20. Иммунотоксины и иммуноферменты рекомбинантные Рекомбинантные иммунотоксины и иммуноферменты получают в результате экспрессии иммуноглобулиновых генов, соединенных с
- 21. Абзимы, каталитические антитела — моноклональные антитела, обладающие каталитической активностью. Существуют как природные абзимы (в молоке, в
- 22. Иммуноадгезины (immunoadhesins) обычно состоят из Fc-фрагмента молекулы антитела и лиганда, специфичного к определенному рецептору, например CD4
- 23. Для улучшения направленной доставки противоопухолевых препаратов было разработано новое поколение лекарственных препаратов — иммунолипосомы. Иммунолипосомы представляют
- 24. В настоящее время различают три типа иммунолипосом: А, В и C. Иммунолипосомы типа А — моноклональные
- 25. Иммунополимеры Биоконъюгаты лекарственных препаратов с биодеградируемыми, растворимыми в воде полимерами, например, гидроксипропилметакриамином (ГПМА), начинают исследоваться в
- 26. галактозомин-направленный ГПМА-полимерный доксурубицин
- 27. ПГК-паклитаксел (конъюгат паклитаксела с полиглутаминовой кислотой, XYOTAX) и ПЭГ-каптотецина (PROTHECAN)
- 29. Скачать презентацию

Генно-инженерная технология революционизировала иммунологию, сформировав направление, получившее название "инженерия антител", целью
Генно-инженерная технология революционизировала иммунологию, сформировав направление, получившее название "инженерия антител", целью
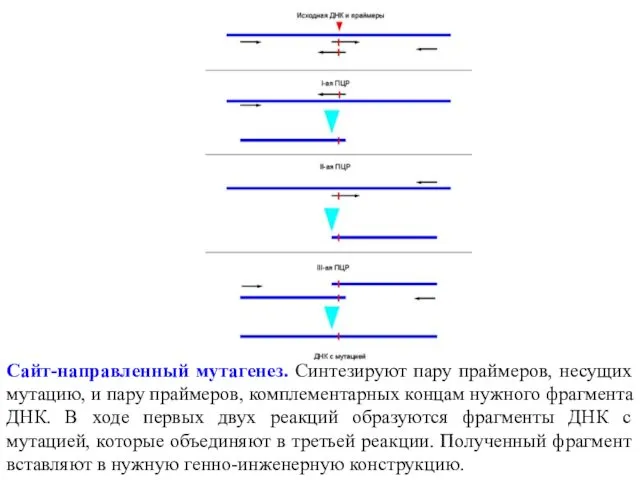
Сайт-направленный мутагенез. Синтезируют пару праймеров, несущих мутацию, и пару праймеров, комплементарных
Сайт-направленный мутагенез. Синтезируют пару праймеров, несущих мутацию, и пару праймеров, комплементарных

Следующим этапом развития гибридомной технологии после получения гибридом стало получение "
Следующим этапом развития гибридомной технологии после получения гибридом стало получение "

В последние годы применение моноклональных антител в научно-исследовательской и клинической практике
В последние годы применение моноклональных антител в научно-исследовательской и клинической практике

Трудности использования моноклональных антител в медицине.
Первая и наиглавнейшая из них –
Трудности использования моноклональных антител в медицине.
Первая и наиглавнейшая из них –
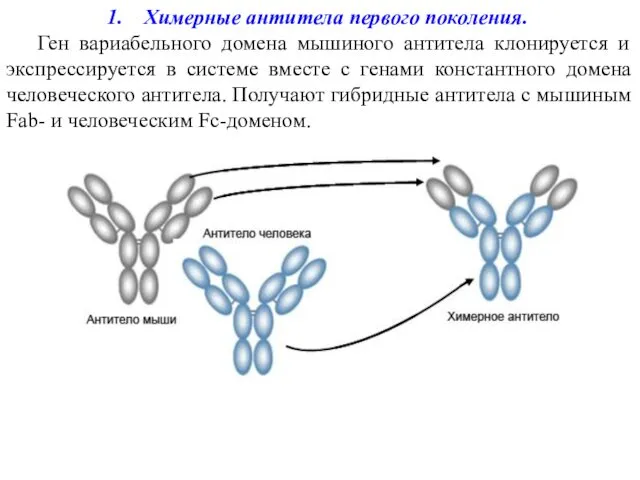
Химерные антитела первого поколения.
Ген вариабельного домена мышиного антитела клонируется и
Химерные антитела первого поколения.
Ген вариабельного домена мышиного антитела клонируется и

2. Гиперхимерные антитела второго поколения.
В этом подходе используются лишь минимальные
2. Гиперхимерные антитела второго поколения.
В этом подходе используются лишь минимальные
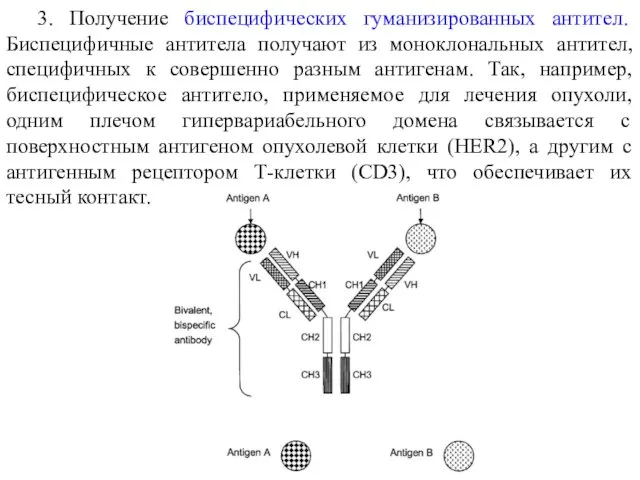
3. Получение биспецифических гуманизированных антител. Биспецифичные антитела получают из моноклональных антител,
3. Получение биспецифических гуманизированных антител. Биспецифичные антитела получают из моноклональных антител,
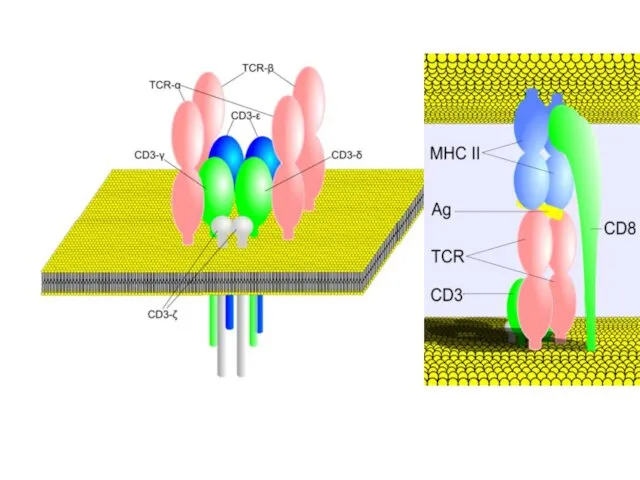

Ортоклон, Муромонаб-CD3
Мононуклеальные антитела против CD3 лимфоцитов человека, обладают иммунодепрессивными свойствами в
Ортоклон, Муромонаб-CD3
Мононуклеальные антитела против CD3 лимфоцитов человека, обладают иммунодепрессивными свойствами в

Этанерцепт — препарат, сочетающий в своей структуре два белка— препарат, сочетающий
Этанерцепт — препарат, сочетающий в своей структуре два белка— препарат, сочетающий

Другим перспективным подходом для решения проблем, обусловленных применением мышиных антител в
Другим перспективным подходом для решения проблем, обусловленных применением мышиных антител в

Fv-фрагменты - нековалентно ассоциированные гетеродимеры V(H) и V(L) -доменов. Они менее
Fv-фрагменты - нековалентно ассоциированные гетеродимеры V(H) и V(L) -доменов. Они менее
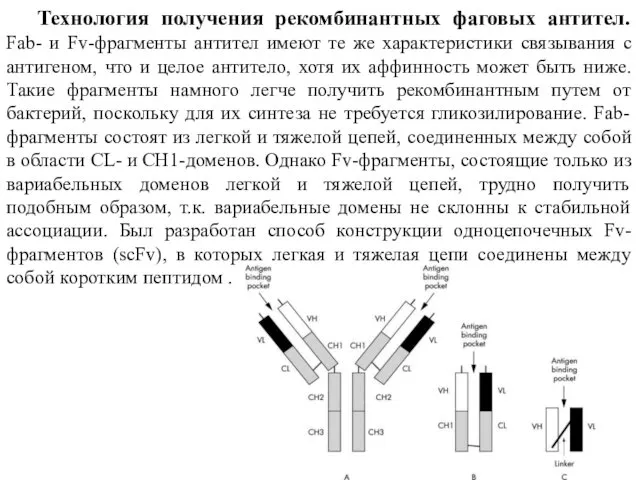
Технология получения рекомбинантных фаговых антител. Fab- и Fv-фрагменты антител имеют те
Технология получения рекомбинантных фаговых антител. Fab- и Fv-фрагменты антител имеют те

В 1990 году McCafferty и соавт. предложили методику, с помощью которой
В 1990 году McCafferty и соавт. предложили методику, с помощью которой

Часто используют ген белка рVIII, однако из-за того, что этот белок
Часто используют ген белка рVIII, однако из-за того, что этот белок
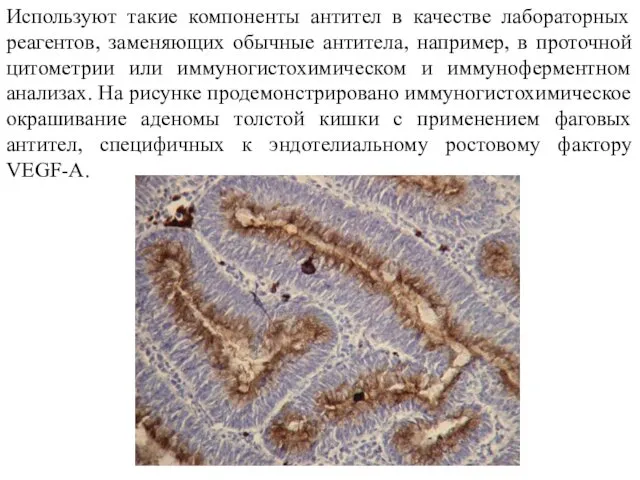
Используют такие компоненты антител в качестве лабораторных реагентов, заменяющих обычные антитела,
Используют такие компоненты антител в качестве лабораторных реагентов, заменяющих обычные антитела,
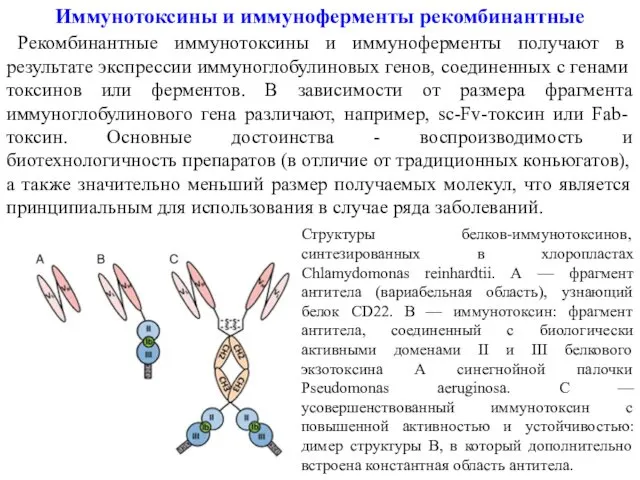
Иммунотоксины и иммуноферменты рекомбинантные
Рекомбинантные иммунотоксины и иммуноферменты получают в результате
Иммунотоксины и иммуноферменты рекомбинантные
Рекомбинантные иммунотоксины и иммуноферменты получают в результате

Абзимы, каталитические антитела — моноклональные антитела, обладающие каталитической активностью. Существуют как
Абзимы, каталитические антитела — моноклональные антитела, обладающие каталитической активностью. Существуют как

Иммуноадгезины (immunoadhesins) обычно состоят из Fc-фрагмента молекулы антитела и лиганда, специфичного
Иммуноадгезины (immunoadhesins) обычно состоят из Fc-фрагмента молекулы антитела и лиганда, специфичного
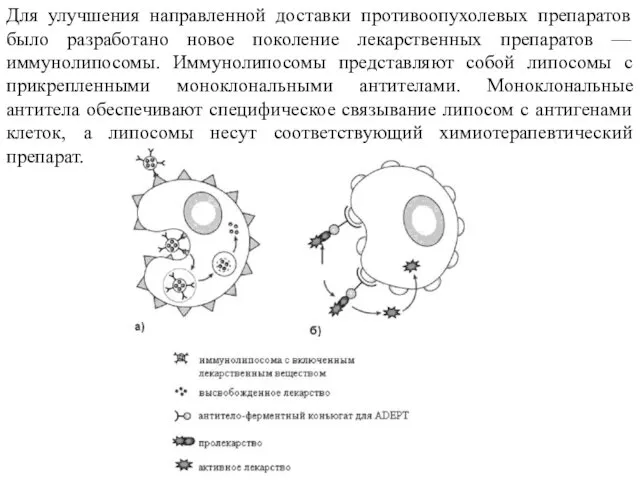
Для улучшения направленной доставки противоопухолевых препаратов было разработано новое поколение лекарственных
Для улучшения направленной доставки противоопухолевых препаратов было разработано новое поколение лекарственных

В настоящее время различают три типа иммунолипосом: А, В и C.
В настоящее время различают три типа иммунолипосом: А, В и C.

Иммунополимеры
Биоконъюгаты лекарственных препаратов с биодеградируемыми, растворимыми в воде полимерами, например,
Иммунополимеры
Биоконъюгаты лекарственных препаратов с биодеградируемыми, растворимыми в воде полимерами, например,
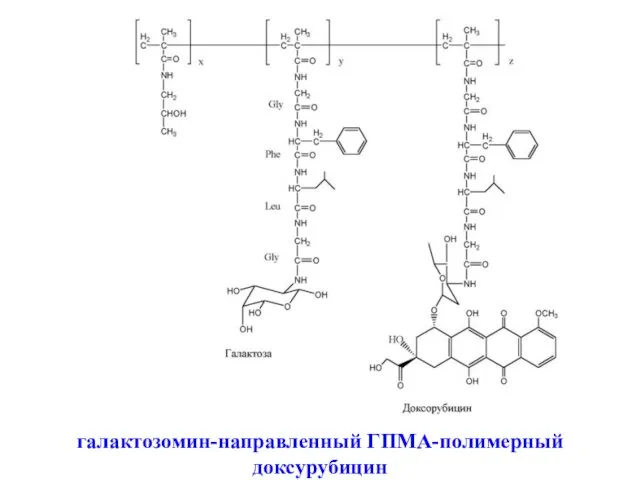
галактозомин-направленный ГПМА-полимерный доксурубицин
галактозомин-направленный ГПМА-полимерный доксурубицин
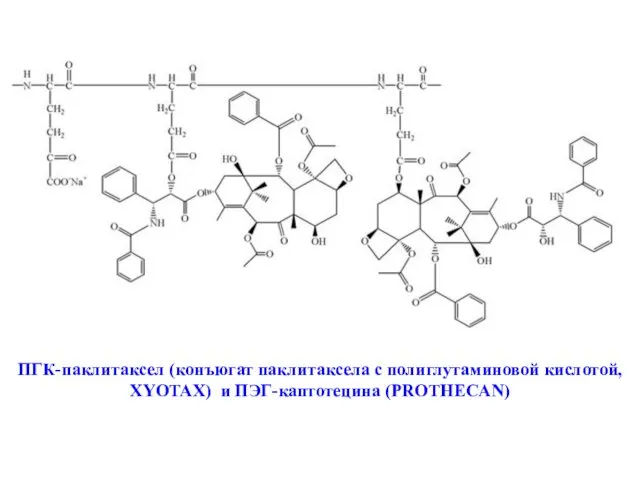
ПГК-паклитаксел (конъюгат паклитаксела с полиглутаминовой кислотой, XYOTAX) и ПЭГ-каптотецина (PROTHECAN)
ПГК-паклитаксел (конъюгат паклитаксела с полиглутаминовой кислотой, XYOTAX) и ПЭГ-каптотецина (PROTHECAN)
 10 самых необычных растений мира
10 самых необычных растений мира Химический состав клетки. Неорганические вещества
Химический состав клетки. Неорганические вещества Гипоталамус - гипофиз - бүйрекүсті бездер жүйес
Гипоталамус - гипофиз - бүйрекүсті бездер жүйес Викторина В мире животных
Викторина В мире животных 20231031_gametogenez_oplodotvorenie
20231031_gametogenez_oplodotvorenie СПИД - сведи вероятность к нулю(презентация)
СПИД - сведи вероятность к нулю(презентация) Царство грибы. Общая характеристика
Царство грибы. Общая характеристика Эпителиальные ткани
Эпителиальные ткани Грибные болезни насекомых
Грибные болезни насекомых Дыхательная система
Дыхательная система Алколоиды
Алколоиды Оптическое устройство глаза
Оптическое устройство глаза Урок-игра Органы цветкового растения
Урок-игра Органы цветкового растения Отдел мохообразные. Общая характеристика
Отдел мохообразные. Общая характеристика Биохимия соединительной ткани
Биохимия соединительной ткани Cell structure function
Cell structure function Interesting facts about the tiger
Interesting facts about the tiger Отличия прокариот от эукариот
Отличия прокариот от эукариот Культивирование бурых водорослей
Культивирование бурых водорослей Получение продуцентов с помощью генной инженерии
Получение продуцентов с помощью генной инженерии Частная физиология нервной системы
Частная физиология нервной системы Тема: Классификация овощей
Тема: Классификация овощей Биосинтез белка
Биосинтез белка Відчуття та сприймання
Відчуття та сприймання Совместная жизнь видов в биогеоценозе
Совместная жизнь видов в биогеоценозе Интерактивный кроссворд к теме Сенсорные системы
Интерактивный кроссворд к теме Сенсорные системы Методы внесения химических средств защиты растений. (Лекция 5)
Методы внесения химических средств защиты растений. (Лекция 5) презентация на тему жизнь в морях и океанах
презентация на тему жизнь в морях и океанах