Ионные каналы, строение, классификация, способ активации, прикладные аспекты клетки. (Лекция 2) презентация
- Ионные каналы, строение, классификация, способ активации, прикладные аспекты клетки. (Лекция 2)

Содержание
- 2. А- - органические анионы 10 30 15 20 000 Активный транспорт ионов вызывает различия в ионном
- 3. Зачем нужен концентрационный градиент? Электрические токи, возникающие в клетке, обеспечиваются пассивным движением ионов через мембрану Для
- 4. Виды ионного транспорта Активный - с затратой энергии АТФ, против концентрационного и/или электрического градиента Первичный Вторичный
- 5. Для того, чтобы ионы могли двигаться через мембрану, необходимо иметь мембранные структуры, сообщающие вне- и внутриклеточную
- 6. Как выглядит ионный канал? Центральная водная пора Устья канала Ворота
- 7. Два основных типа ионных каналов В зависимости от роли в нейрональной сигнализации, различают 2 основных типа
- 8. Работа канала Покой – канал закрыт, но может открыться под действием адекватного стимула Активация- открытие канала
- 9. Модель потенциал-зависимого ионного канала
- 10. Классификация ионных каналов По избирательности Неселективные (никотиновый холинорецептор) Селективные (Na+, K+, Ca++, Cl-) По механизму активации
- 11. Избирательность (селективность) каналов Селективные (Na+, К+, Са 2+, Cl- каналы ). Селективность определяется размерами поры и
- 12. Неселективный ионный канал Н-холинорецептор
- 13. Способы открытия (активации) управляемых ионных каналов Активация физическими изменениями Потенциал-управляемые каналы Каналы, активирующиеся растяжением Активация химическими
- 14. Калиевые каналы
- 15. Кальциевые каналы
- 16. Открытое и закрытое состояние ионных каналов Переход из закрытого в открытое состояние происходит моментально. Канал открывается
- 17. Молекулярные механизмы активации и инактивации каналов
- 18. Проводимость и проницаемость каналов Величина тока, проходящего через канал, связана со скоростью движения ионов через него
- 19. Строение ионного канала Цитоплазма Аминокислоты Спиральные сегменты Домены Субъединицы Канал Методы исследования Выделение белков каналов Аминокислотная
- 20. Потенциал-управляемые селективные ионные каналы 4 сенсора потенциала
- 21. Структура основных потенциал-активируемых ионных каналов Порообразующая α-субъединица потенциал-активируемых натриевых и кальциевых каналов представляет собой одну белковую
- 22. Работа отдельного канала пэтч-кламп (patch-clamp) Преимущества Возможность исследовать отдельный канал Возможность менять потенциал на мембране Возможность
- 23. Нобелевская премия 1991 года в области физиологии и медицины Эрвин Нейер и Берт Сакманн «за открытия
- 24. Что заставляет ионы двигаться через открытые каналы?
- 25. Движение ионов через каналы Движение иона через канал управляется двумя силами: 1) химической движущей силой, которая
- 26. Равновесные потенциалы(Е) итоговая движущая сила (V- Е) Cl- -89 - 47 Cl-каналы i = gV, i
- 27. Расчет равновесного потенциала Равновесный потенциал для какого-либо иона Х можно рассчитать из уравнения, полученного в 1888
- 29. Скачать презентацию
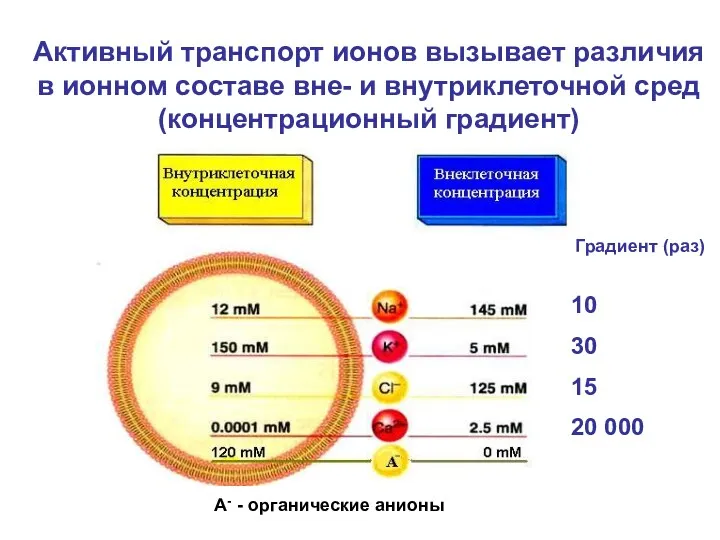
А- - органические анионы
10
30
15
20 000
Активный транспорт ионов вызывает различия
в ионном
А- - органические анионы
10
30
15
20 000
Активный транспорт ионов вызывает различия
в ионном

Зачем нужен концентрационный градиент?
Электрические токи, возникающие в клетке, обеспечиваются пассивным движением
Зачем нужен концентрационный градиент?
Электрические токи, возникающие в клетке, обеспечиваются пассивным движением

Виды ионного транспорта
Активный - с затратой энергии АТФ, против концентрационного и/или
Виды ионного транспорта
Активный - с затратой энергии АТФ, против концентрационного и/или
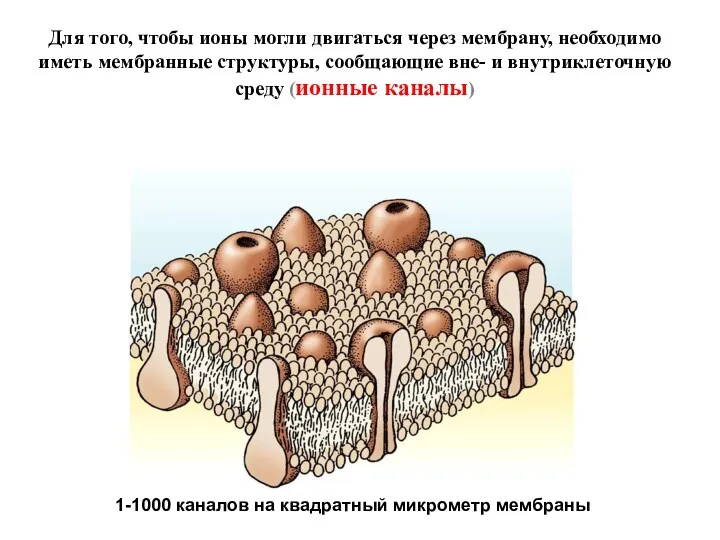
Для того, чтобы ионы могли двигаться через мембрану, необходимо иметь мембранные
Для того, чтобы ионы могли двигаться через мембрану, необходимо иметь мембранные
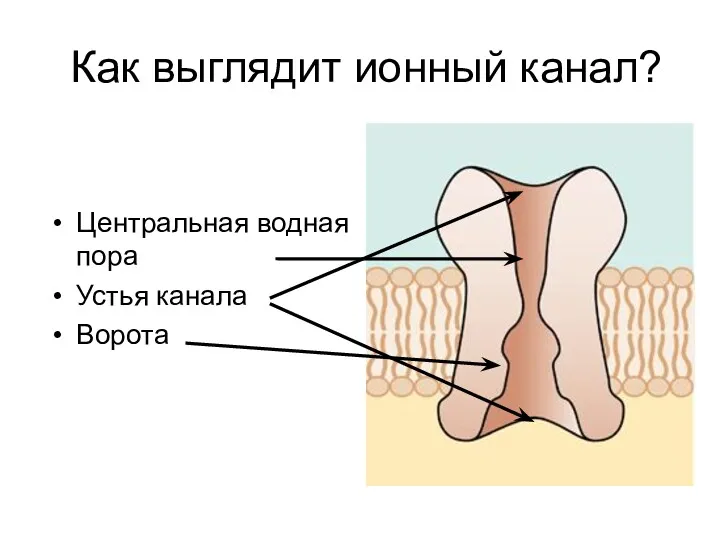
Как выглядит ионный канал?
Центральная водная пора
Устья канала
Ворота
Как выглядит ионный канал?
Центральная водная пора
Устья канала
Ворота
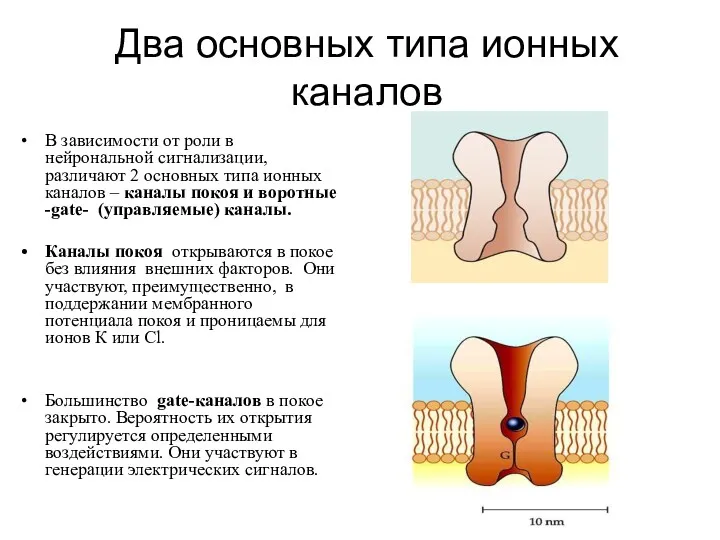
Два основных типа ионных каналов
В зависимости от роли в нейрональной сигнализации,
Два основных типа ионных каналов
В зависимости от роли в нейрональной сигнализации,

Работа канала
Покой – канал закрыт, но может открыться под действием адекватного
Работа канала
Покой – канал закрыт, но может открыться под действием адекватного
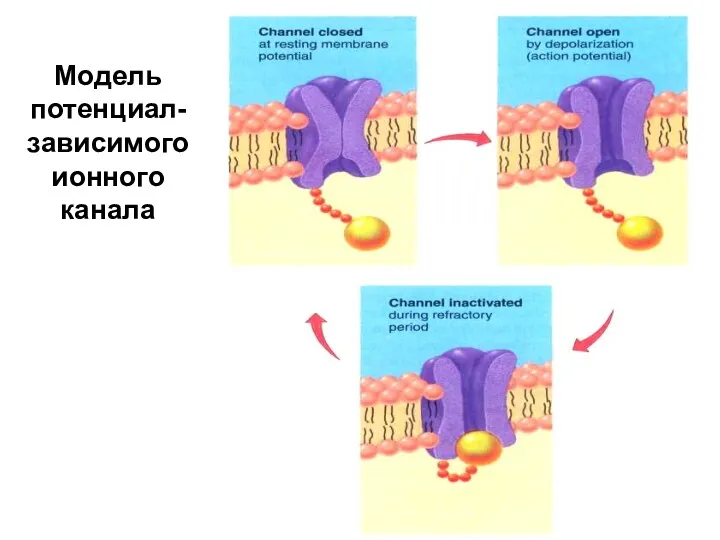
Модель потенциал-зависимого ионного канала
Модель потенциал-зависимого ионного канала

Классификация ионных каналов
По избирательности
Неселективные (никотиновый холинорецептор)
Селективные (Na+, K+, Ca++, Cl-)
По
Классификация ионных каналов
По избирательности
Неселективные (никотиновый холинорецептор)
Селективные (Na+, K+, Ca++, Cl-)
По
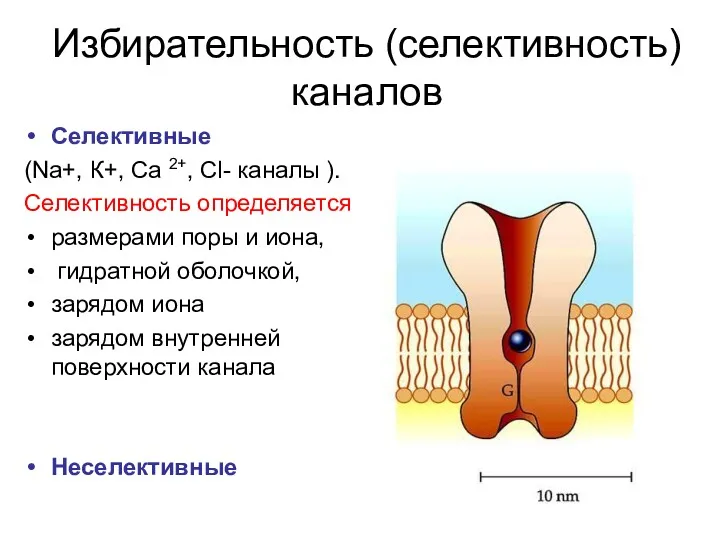
Избирательность (селективность) каналов
Селективные
(Na+, К+, Са 2+, Cl- каналы ).
Избирательность (селективность) каналов
Селективные
(Na+, К+, Са 2+, Cl- каналы ).
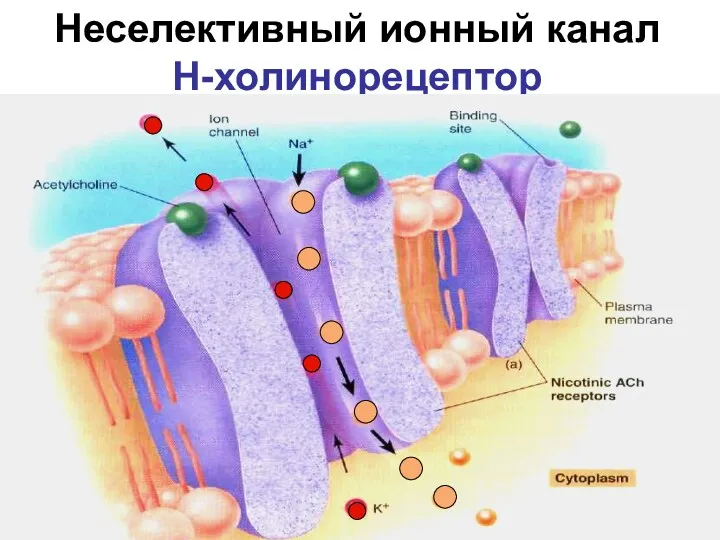
Неселективный ионный канал
Н-холинорецептор
Неселективный ионный канал
Н-холинорецептор
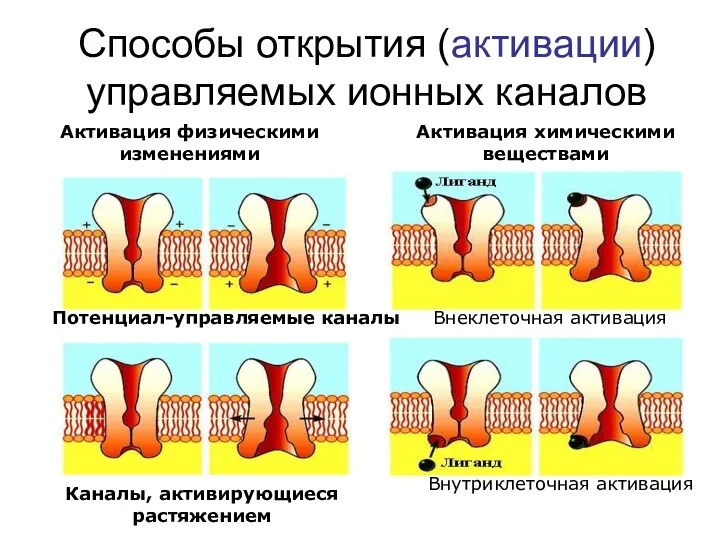
Способы открытия (активации) управляемых ионных каналов
Активация физическими изменениями
Потенциал-управляемые каналы
Каналы, активирующиеся растяжением
Активация
Способы открытия (активации) управляемых ионных каналов
Активация физическими изменениями
Потенциал-управляемые каналы
Каналы, активирующиеся растяжением
Активация
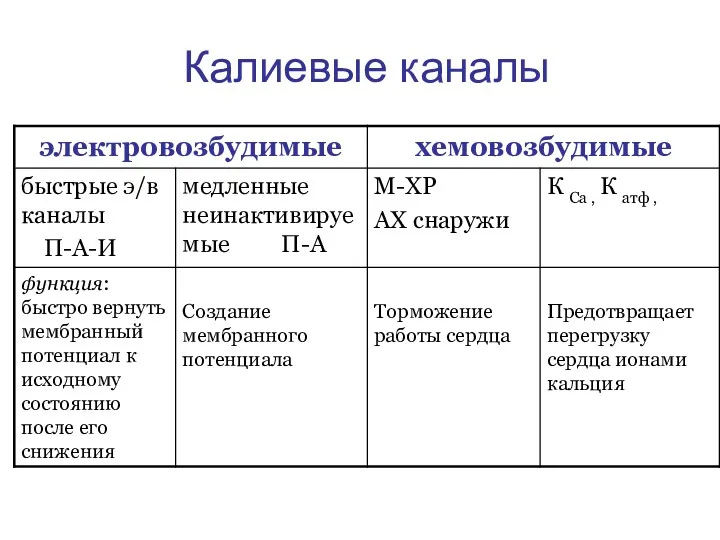
Калиевые каналы
Калиевые каналы
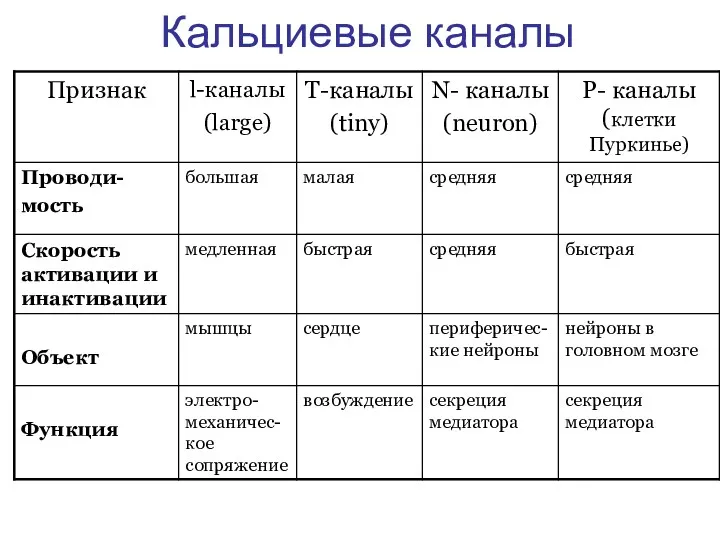
Кальциевые каналы
Кальциевые каналы

Открытое и закрытое состояние ионных каналов
Переход из закрытого в открытое
Открытое и закрытое состояние ионных каналов
Переход из закрытого в открытое
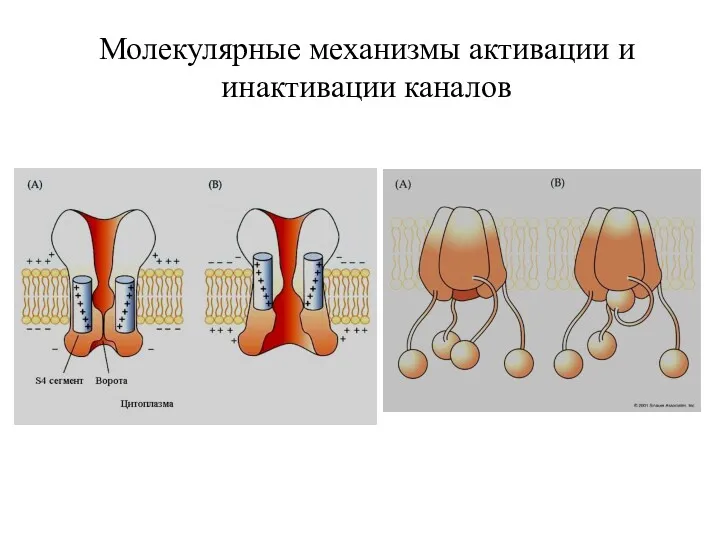
Молекулярные механизмы активации и инактивации каналов
Молекулярные механизмы активации и инактивации каналов

Проводимость и проницаемость каналов
Величина тока, проходящего через канал, связана со скоростью
Проводимость и проницаемость каналов
Величина тока, проходящего через канал, связана со скоростью
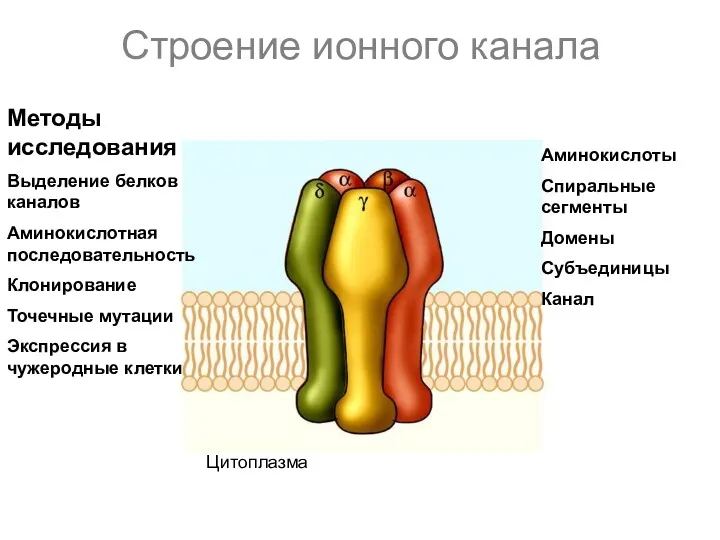
Строение ионного канала
Цитоплазма
Аминокислоты
Спиральные сегменты
Домены
Субъединицы
Канал
Методы исследования
Выделение белков каналов
Аминокислотная последовательность
Клонирование
Точечные мутации
Экспрессия в чужеродные
Строение ионного канала
Цитоплазма
Аминокислоты
Спиральные сегменты
Домены
Субъединицы
Канал
Методы исследования
Выделение белков каналов
Аминокислотная последовательность
Клонирование
Точечные мутации
Экспрессия в чужеродные
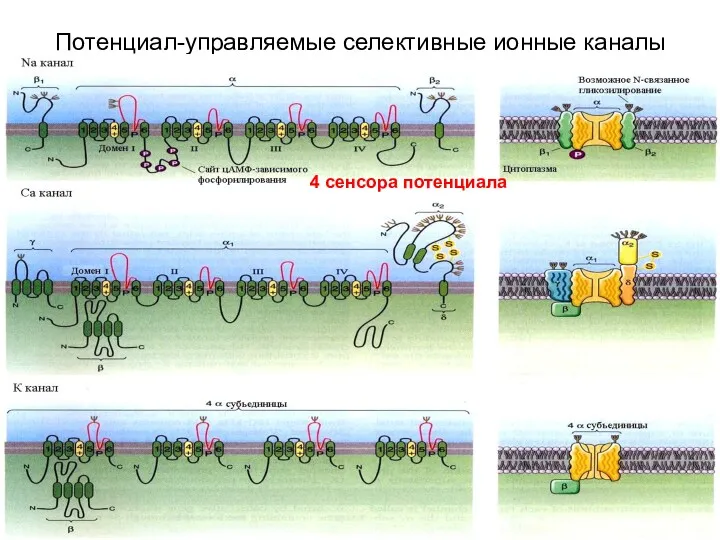
Потенциал-управляемые селективные ионные каналы
4 сенсора потенциала
Потенциал-управляемые селективные ионные каналы
4 сенсора потенциала
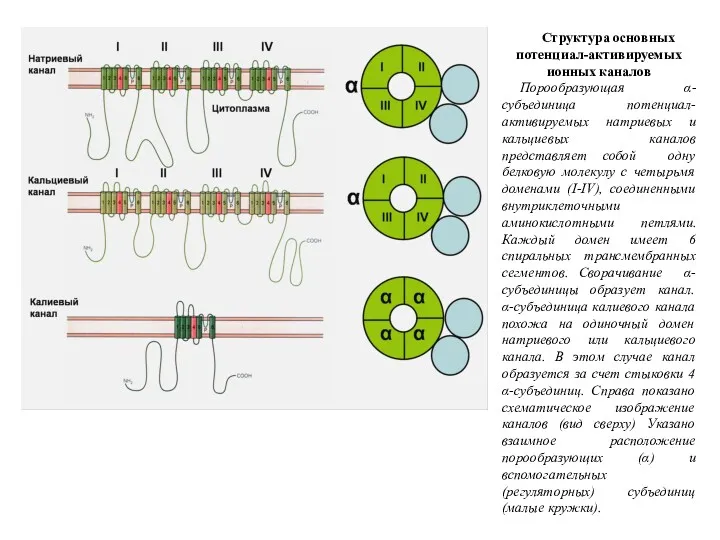
Структура основных потенциал-активируемых ионных каналов
Порообразующая α-субъединица потенциал-активируемых натриевых и кальциевых каналов
Структура основных потенциал-активируемых ионных каналов
Порообразующая α-субъединица потенциал-активируемых натриевых и кальциевых каналов
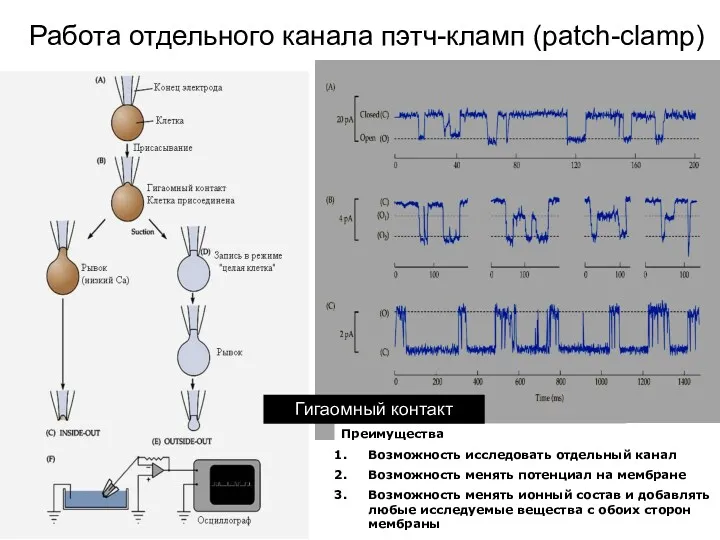
Работа отдельного канала пэтч-кламп (patch-clamp)
Преимущества
Возможность исследовать отдельный канал
Возможность менять потенциал на
Работа отдельного канала пэтч-кламп (patch-clamp)
Преимущества
Возможность исследовать отдельный канал
Возможность менять потенциал на

Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейер и
Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейер и

Что заставляет ионы двигаться через открытые каналы?
Что заставляет ионы двигаться через открытые каналы?
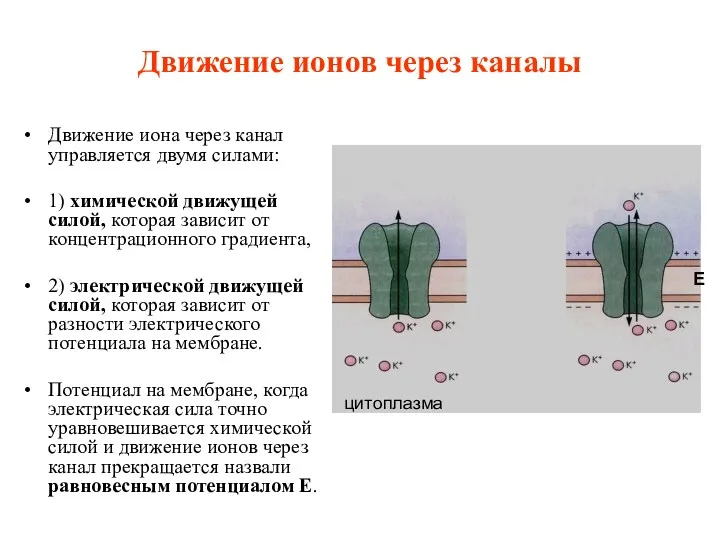
Движение ионов через каналы
Движение иона через канал управляется двумя силами:
1)
Движение ионов через каналы
Движение иона через канал управляется двумя силами:
1)

Равновесные потенциалы(Е) итоговая движущая сила (V- Е)
Cl-
-89 - 47
Cl-каналы
i = gV,
i
Равновесные потенциалы(Е) итоговая движущая сила (V- Е)
Cl-
-89 - 47
Cl-каналы
i = gV,
i

Расчет равновесного потенциала
Равновесный потенциал для какого-либо иона Х можно рассчитать из
Расчет равновесного потенциала
Равновесный потенциал для какого-либо иона Х можно рассчитать из
 Механическая обработка молока
Механическая обработка молока Чудо-огірок
Чудо-огірок Жыныс жасушаларының құрылысы
Жыныс жасушаларының құрылысы Строение и жизненные циклы червей класса cestoda
Строение и жизненные циклы червей класса cestoda Чому мігрують тварини. 7 клас
Чому мігрують тварини. 7 клас Жизнь леса
Жизнь леса Yon-Ka. Препараты для мужчин
Yon-Ka. Препараты для мужчин Катархей. Архей
Катархей. Архей Зимующие птицы
Зимующие птицы Биологическая безопасность в биотехнологии
Биологическая безопасность в биотехнологии Предмет, задачи и история развития радиобиологии, ее роль в решении задач агропромышленного комплекса
Предмет, задачи и история развития радиобиологии, ее роль в решении задач агропромышленного комплекса Хвостатая семейка. Интересные факты и истории из жизни
Хвостатая семейка. Интересные факты и истории из жизни Общая характеристика нематод. Оксиуроз лошадей. Пассалуроз кроликов. Гетеракиоз кур
Общая характеристика нематод. Оксиуроз лошадей. Пассалуроз кроликов. Гетеракиоз кур Рост и развитие волос
Рост и развитие волос Различия в строении клеток эукариот и прокариот. Строение клеток прокариот
Различия в строении клеток эукариот и прокариот. Строение клеток прокариот Проект Лук - зеленый друг. 3 класс
Проект Лук - зеленый друг. 3 класс Отряд Жуки, или Жёсткокрылые
Отряд Жуки, или Жёсткокрылые Задача 2. Яблоки. Механизм потемнения среза яблок
Задача 2. Яблоки. Механизм потемнения среза яблок Слухова сенсорна система
Слухова сенсорна система Видообразование. Пути и способы видообразования
Видообразование. Пути и способы видообразования Гусеобразные Мелеузовского района: земляные и речные утки. Часть 2
Гусеобразные Мелеузовского района: земляные и речные утки. Часть 2 Кіт Лісовий
Кіт Лісовий Общая морфология клеток прокариот
Общая морфология клеток прокариот Интерактивная игра Загадки
Интерактивная игра Загадки Радиационная биофизика. Кислородный эффект в радиобиологии. (Лекция 4)
Радиационная биофизика. Кислородный эффект в радиобиологии. (Лекция 4) Беловежская пуща
Беловежская пуща Митоз — это деление клетки
Митоз — это деление клетки Класс двудольные. Семейство Крестоцветные
Класс двудольные. Семейство Крестоцветные