- Изменения морфологии неопластических клеток. Лекция 6

Содержание
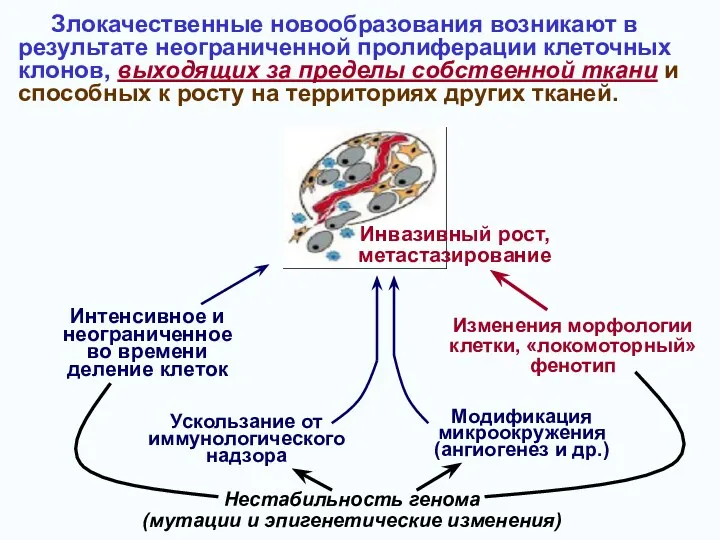
- 2. Злокачественные новообразования возникают в результате неограниченной пролиферации клеточных клонов, выходящих за пределы собственной ткани и способных
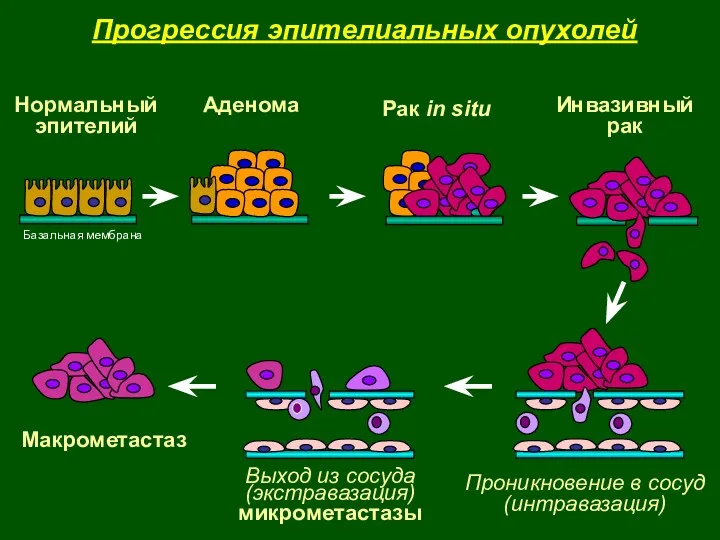
- 3. Нормальный эпителий Аденома Рак in situ Инвазивный рак Базальная мембрана Проникновение в сосуд (интравазация) Выход из
- 4. Инвазия клеток рака молочной железы Нормальная ткань молочной железы Рак in situ (внутрипротоковый рак) Инвазивный рак
- 5. Прижизненная мультифотонная микроскопия: Опухолевые клетки в кровеносном сосуде Кровеносный сосуд Опухолевые клетки Внеклеточный матрикс
- 6. Как клетки приобретают повышенную локомоторную активность и способность к инвазии? Видеосъемка А. Александровой и М. Ломакиной
- 7. А т т р а к т а н т Протрузия Ретракция Направленное движение клетки:
- 8. А т т р а к т а н т Протрузия Ретракция Направленное движение клетки: Протрузия
- 9. циклы «протрузия-ретракция» Фибробласты и другие типы мезенхимальных клеток Вид сбоку Вид сверху Ведущий край Хвост Типы
- 10. Локомоция Активность псевдоподий и перестройка актинового цитоскелета, обеспечивающая протрузии и ретракции Разрушение контактов клеток друг с
- 11. Амебоидное движение (формирование блебов и «перетекание») Фибробластоподобное движение (циклы «протрузия-ретракция») Фибробласты и многие другие типы клеток
- 12. Амебоидная миграция клеток карциносаркомы Уокера под агарозой Bergert et al., Proc Natl Acad Sci U S
- 13. Разные типы клеток исходно обладают разной способностью к миграции: клетки крови, фибробласты – высокой эпителиальные клетки
- 14. Различия морфологической организации эпителиоцитов и фибробластов Эпителиальная клетка Фибробласт Межклеточные контакты (Е-кадгерин) кольцевой актиновый пучок прямые
- 15. Организация актина на ведущем крае клетки при фибробластоподобном движении Протрузия обеспечивается полимеризацией и ветвлением актиновых фибрилл
- 16. Rho Rac1 MLC-P Rho-киназа WAVE Arp2/3 Rac1 Функция Rac1 обеспечивает формирование протрузий (полимеризация актина ламеллоподии), функция
- 17. Croft and Olson, Cancer Cell, 2008, 14:349-351 Регуляция фибробластоподобного и амебоидного движения в клетках меланомы ?
- 18. Изменение типа клеточной подвижности при переключении активности внутриклеточных сигнальных путей (ламеллиподии и блебы) НТ1080 НТ1080 +
- 19. 3D cancer cell invasion. Multicellular spheroids of HT-1080 fibrosarcoma cells were embedded within 3D gels of
- 20. Инвадоподии (подосомы) Yamaguchi et al., Current Opinion in Cell Biology, 2005 Revach OY, Geiger B, Cell
- 21. Изменения формы клеток и организации актинового цитоскелета при неопластической трансформации Эпителиоцит Фибробласты Ras актин винкулин IAR2-C4
- 22. Эпителиальная клетка Фибробласт Трансформированные клетки Неопластическая трансформация морфологии
- 23. N-Ras Rac1/GTP Rho/GTP P-cofilin Клетки REF52 Rho Ras Rac1 LIMK кофилин Образование «рафлов» Разрушение актиновых фибрилл
- 24. Нормальный фибробласт человека Фибробласт человека, трансформированный SV40 Препараты и видеосъемка А. Александровой и М. Ломакиной
- 25. Изменение характера движения трансформированных фибробластов (без аттрактанта) А. Александрова и М. Ломакина, 2007 0ч-2ч-4ч 6ч-8ч Нормальные
- 26. Тесты на миграционную и инвазивную способность клеток in vitro: миграция через фильтры (8 μM), покрытые матригелем,
- 27. Е-кадгерин (зеленый) Виментин (красный) F-актин (красный) Локомоторный фенотип эпителиальных неопластических клеток часто связан с эпителиально-мезенхимальным переходом
- 28. Эпителиально-мезенхимальный переход Утрата эпителиальной полярности Разделение на отдельные клетки Дисперсия при приобретении клеточной подвижности Признаки ЭМП:
- 29. Shibue and Weinberg, Nat Rev Clin Oncol, 2017 14(10): 611–629
- 30. На ранних этапах ЭМП происходит разрушение краевого актинового пучка и перестройка стабильных тангенциальных Е-кадхерин-содержащих контактов между
- 31. Физиологические механизмы ЭМП (эмбриогенез, заживление ран) Активация рецепторов факторов роста (HGF/SF, TGFβ и др.) Активация Notch*
- 32. Репрессия HES, HRT и др. Notch HES, HRT и др. CSL MAML PS p300 CoR MAML
- 33. TGFβ-зависимая сигнализация Gudey et al., Future Oncol. 2014, 10(11):1853-61
- 34. Регулируемый транс-мембранный протеолиз – общий механизм активации внутриклеточной сигнализации в опухолях
- 35. Переключение маркеров эпителиальной и мезенхимальной дифференцировок при экспрессии мутантного онкобелка Ras в эпителиальных клетках молочной железы
- 36. ЭМП, локомоторный фенотип Реорганизация цитоскелета и контактов с матриксом Активность псевдоподий и стресс-фибрилл Разрушение межклеточных контактов
- 37. Механизмы ЭМП Гиперметилирование, инактивирующие мутации гена Е-кадгерина Активация рецепторов HGF/SF, TGFβ и др. цитокинов Активирующие мутации
- 38. Амебоидное движение Фибробластоподобное движение ЭМП может вызывать фибробластоподобное и амебоидное движение клеток Вид сбоку Вид сверху
- 39. ЭМП сопровождается приобретением ряда характеристик/маркеров стволовых опухолевых клеток (CD44+/CD24- и др.) Mani et al., Cell, 2008,
- 40. Транскрипционные факторы Рецепторы митогенов/мотогенов ГТФ-азы сем. Rho (Rac, Rho, Cdc42) Миграция Реорганизация цитоскелета и контактов с
- 41. Wicki et al., Cancer Cell, 2006, 9:261-272 Подопланин (красный) и Е-кадгерин (коричневый) ко-экспрессируются на инвазивном фронте
- 42. Adapted from Yamazaki et al.,Cancer Sci, 96, pp. 379–386, 2005 Стратегии миграции опухолевых клеток Podoplanin, MMPs
- 43. Shibue and Weinberg, Nat Rev Clin Oncol, 2017 14(10): 611–629 ЭМП и МЭП при прогрессии опухолей
- 44. Rho-ГТФазы (Rac, Rho, Cdc42) Реорганизация цитоскелета и контактов с матриксом Образование псевдоподий и стресс-фибрилл Разрушение межклеточных
- 45. Откуда приходят сигналы, индуцирующие ЭМП? Мутации рецепторов и других компонентов сигнальных путей в опухолевой клетке, активация
- 46. Злокачественные новообразования возникают в результате неограниченной пролиферации клеточных клонов, выходящих за пределы собственной ткани и способных
- 48. Скачать презентацию
Злокачественные новообразования возникают в
результате неограниченной пролиферации клеточных клонов, выходящих за
Злокачественные новообразования возникают в
результате неограниченной пролиферации клеточных клонов, выходящих за
Нормальный
эпителий
Аденома
Рак in situ
Инвазивный
рак
Базальная мембрана
Проникновение в сосуд
(интравазация)
Выход из сосуда
(экстравазация)
микрометастазы
Макрометастаз
Прогрессия эпителиальных опухолей
Нормальный
эпителий
Аденома
Рак in situ
Инвазивный
рак
Базальная мембрана
Проникновение в сосуд
(интравазация)
Выход из сосуда
(экстравазация)
микрометастазы
Макрометастаз
Прогрессия эпителиальных опухолей
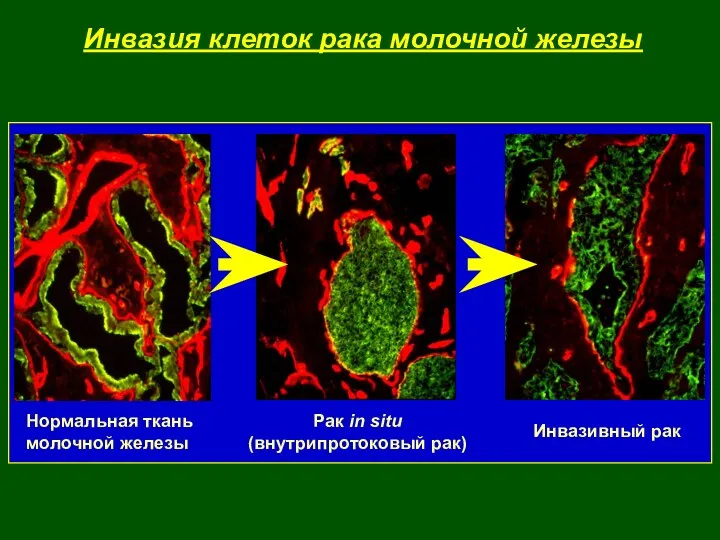
Инвазия клеток рака молочной железы
Нормальная ткань
молочной железы
Рак in situ
(внутрипротоковый рак)
Инвазивный рак
Инвазия клеток рака молочной железы
Нормальная ткань
молочной железы
Рак in situ
(внутрипротоковый рак)
Инвазивный рак
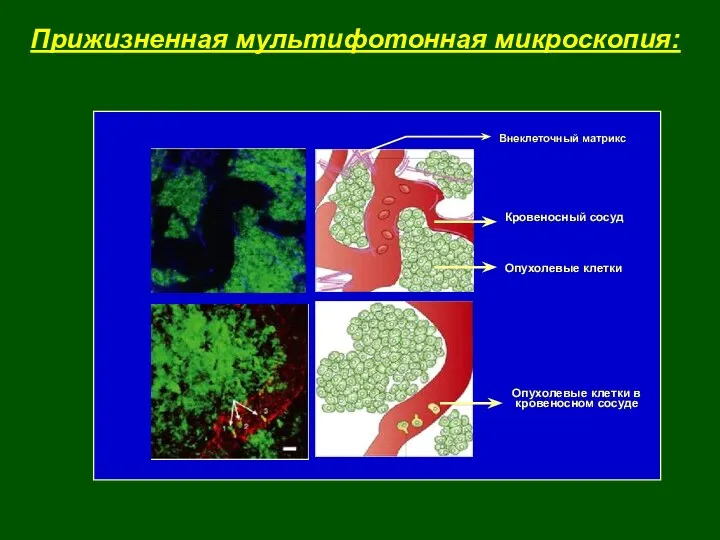
Прижизненная мультифотонная микроскопия:
Опухолевые клетки в
кровеносном сосуде
Кровеносный сосуд
Опухолевые клетки
Внеклеточный матрикс
Прижизненная мультифотонная микроскопия:
Опухолевые клетки в
кровеносном сосуде
Кровеносный сосуд
Опухолевые клетки
Внеклеточный матрикс
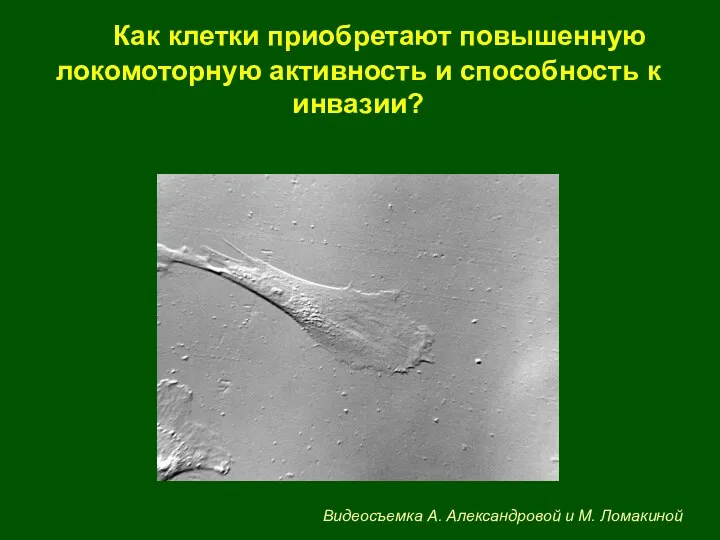
Как клетки приобретают повышенную локомоторную активность и способность к инвазии?
Видеосъемка
Как клетки приобретают повышенную локомоторную активность и способность к инвазии?
Видеосъемка
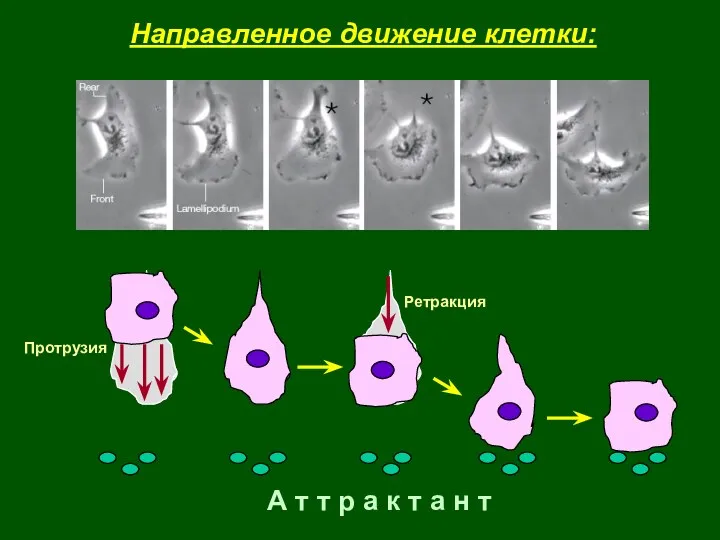
А т т р а к т а н т
Протрузия
Ретракция
Направленное движение
А т т р а к т а н т
Протрузия
Ретракция
Направленное движение
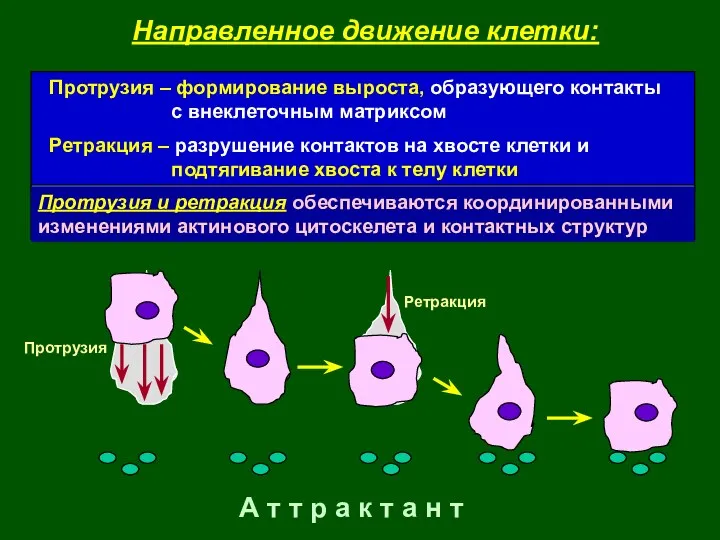
А т т р а к т а н т
Протрузия
Ретракция
Направленное движение
А т т р а к т а н т
Протрузия
Ретракция
Направленное движение
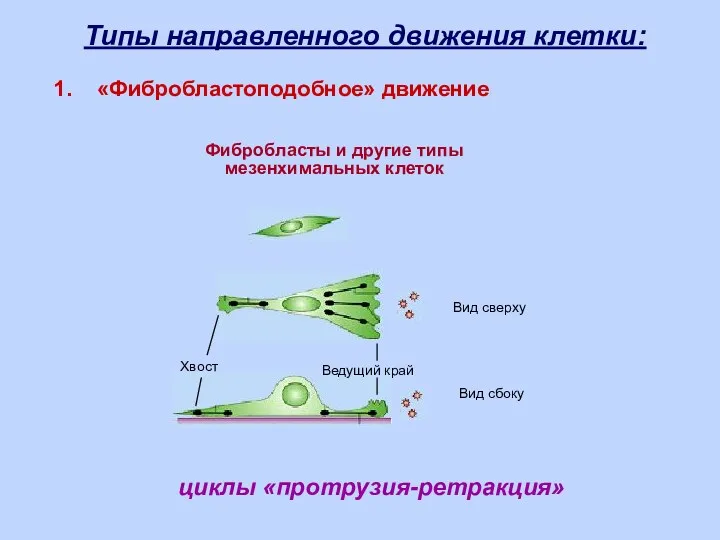
циклы «протрузия-ретракция»
Фибробласты и другие типы
мезенхимальных клеток
Вид сбоку
Вид сверху
Ведущий край
Хвост
Типы направленного движения
циклы «протрузия-ретракция»
Фибробласты и другие типы
мезенхимальных клеток
Вид сбоку
Вид сверху
Ведущий край
Хвост
Типы направленного движения
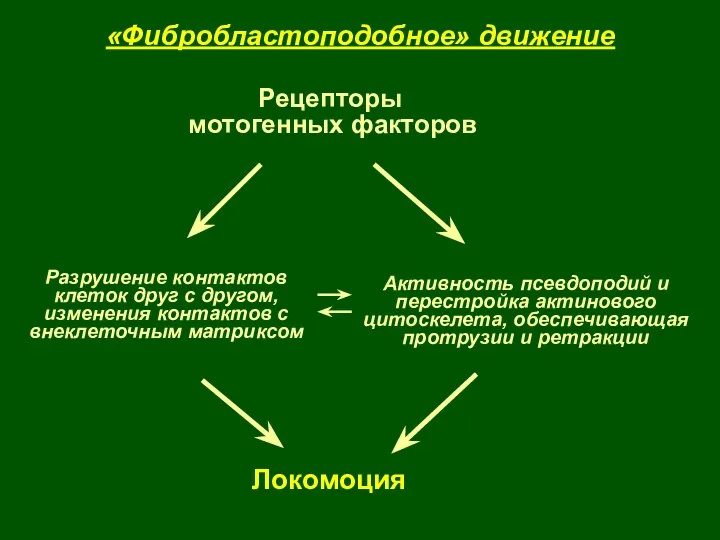
Локомоция
Активность псевдоподий и перестройка актинового цитоскелета, обеспечивающая протрузии и ретракции
Разрушение контактов
клеток
Локомоция
Активность псевдоподий и перестройка актинового цитоскелета, обеспечивающая протрузии и ретракции
Разрушение контактов
клеток
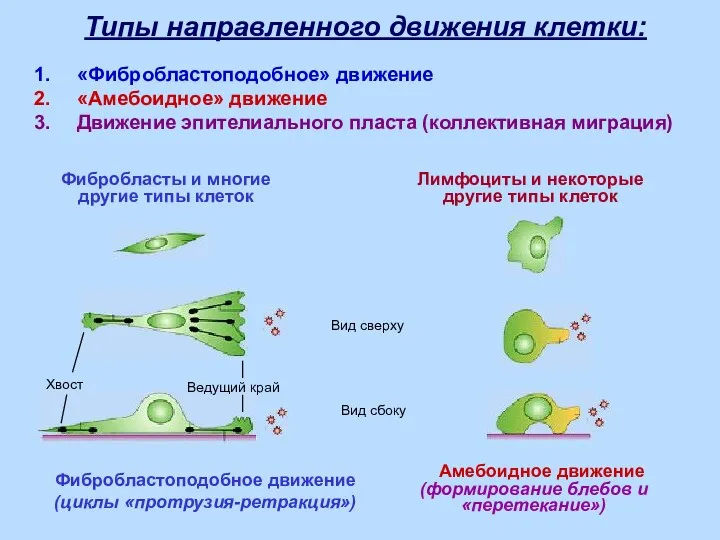
Амебоидное движение
(формирование блебов и «перетекание»)
Фибробластоподобное движение
(циклы «протрузия-ретракция»)
Фибробласты и многие
другие
Амебоидное движение
(формирование блебов и «перетекание»)
Фибробластоподобное движение
(циклы «протрузия-ретракция»)
Фибробласты и многие
другие
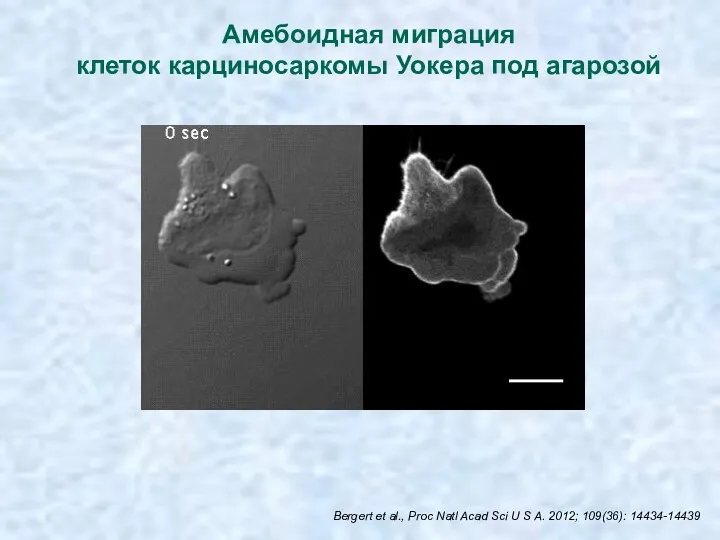
Амебоидная миграция
клеток карциносаркомы Уокера под агарозой
Bergert et al., Proc Natl
Амебоидная миграция
клеток карциносаркомы Уокера под агарозой
Bergert et al., Proc Natl

Разные типы клеток исходно обладают
разной способностью к миграции:
клетки крови, фибробласты –
Разные типы клеток исходно обладают
разной способностью к миграции:
клетки крови, фибробласты –
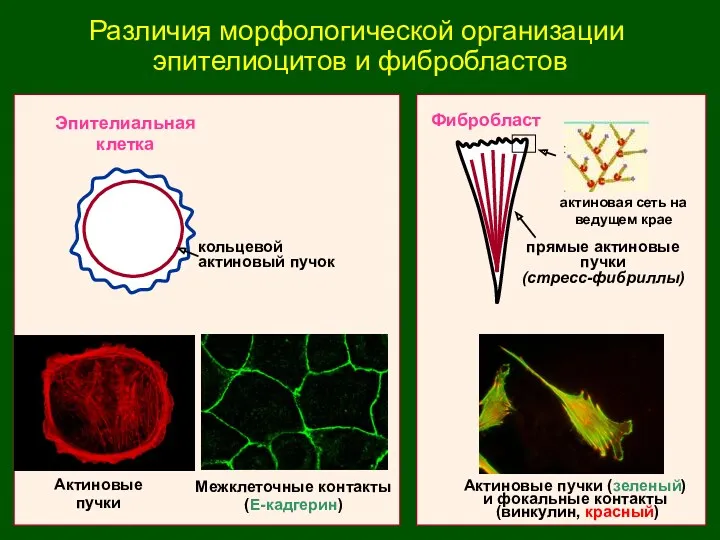
Различия морфологической организации
эпителиоцитов и фибробластов
Эпителиальная
клетка
Фибробласт
Межклеточные контакты
(Е-кадгерин)
кольцевой
актиновый пучок
прямые актиновые
пучки
Различия морфологической организации
эпителиоцитов и фибробластов
Эпителиальная
клетка
Фибробласт
Межклеточные контакты
(Е-кадгерин)
кольцевой
актиновый пучок
прямые актиновые
пучки
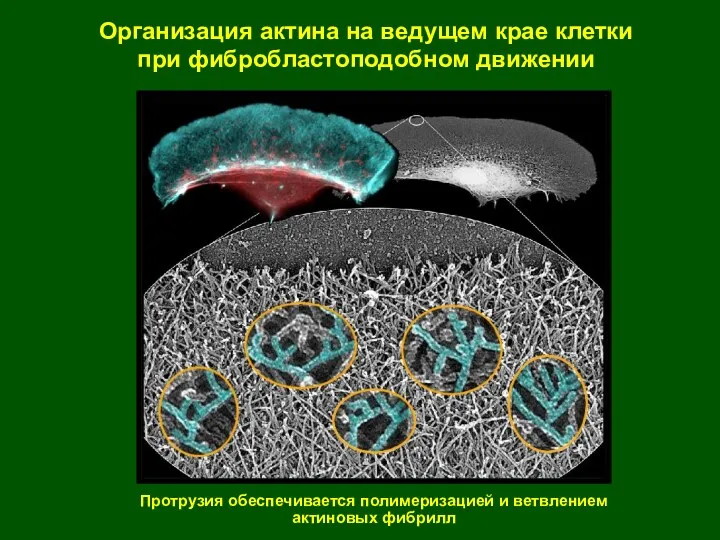
Организация актина на ведущем крае клетки
при фибробластоподобном движении
Протрузия обеспечивается полимеризацией
Организация актина на ведущем крае клетки
при фибробластоподобном движении
Протрузия обеспечивается полимеризацией
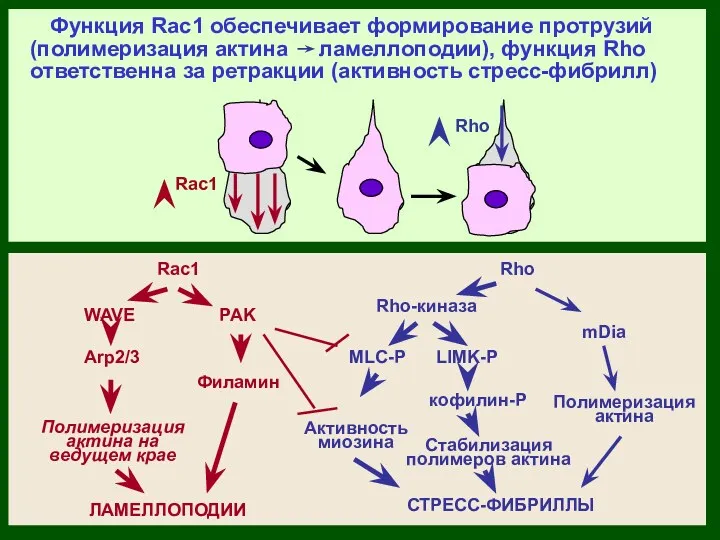
Rho
Rac1
MLC-P
Rho-киназа
WAVE
Arp2/3
Rac1
Функция Rac1 обеспечивает формирование протрузий (полимеризация актина ламеллоподии), функция Rho
Rho
Rac1
MLC-P
Rho-киназа
WAVE
Arp2/3
Rac1
Функция Rac1 обеспечивает формирование протрузий (полимеризация актина ламеллоподии), функция Rho
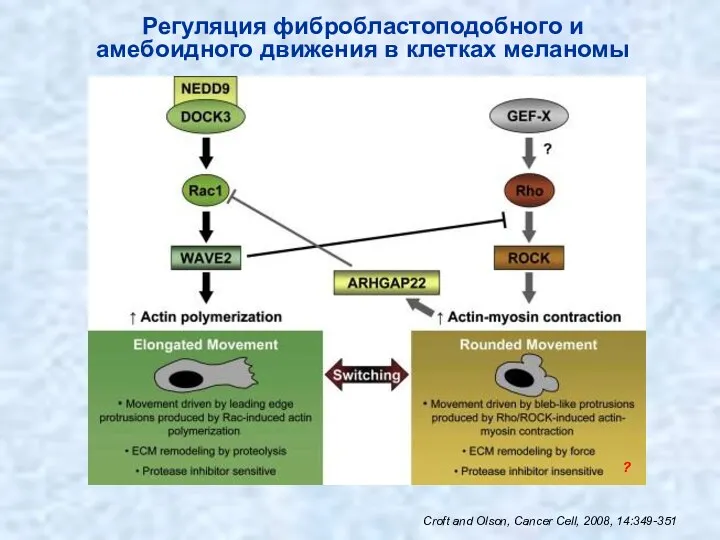
Croft and Olson, Cancer Cell, 2008, 14:349-351
Регуляция фибробластоподобного и амебоидного движения
Croft and Olson, Cancer Cell, 2008, 14:349-351
Регуляция фибробластоподобного и амебоидного движения
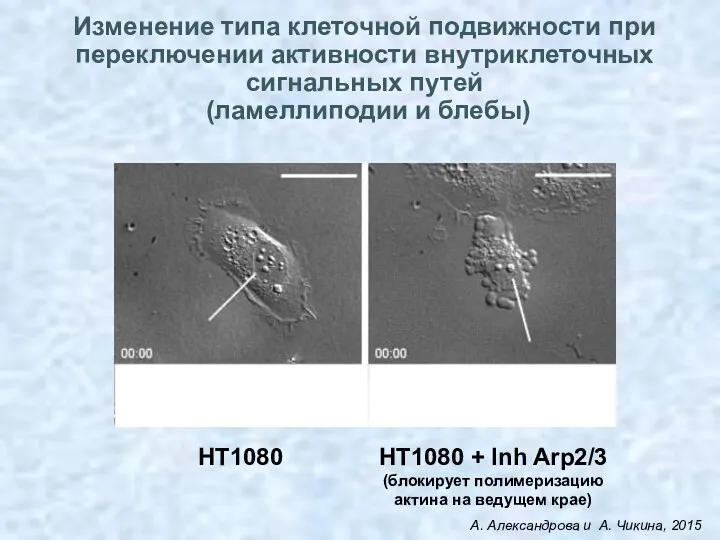
Изменение типа клеточной подвижности при переключении активности внутриклеточных сигнальных путей
(ламеллиподии
Изменение типа клеточной подвижности при переключении активности внутриклеточных сигнальных путей
(ламеллиподии
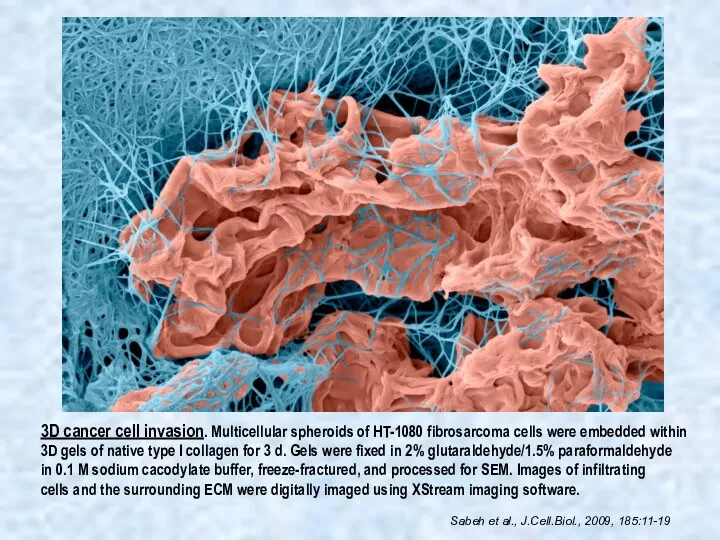
3D cancer cell invasion. Multicellular spheroids of HT-1080 fibrosarcoma cells were
3D cancer cell invasion. Multicellular spheroids of HT-1080 fibrosarcoma cells were
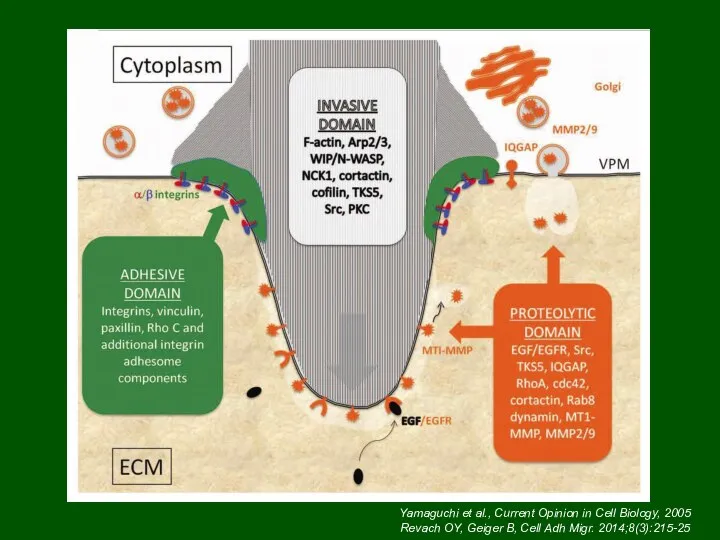
Инвадоподии (подосомы)
Yamaguchi et al., Current Opinion in Cell Biology, 2005
Revach OY,
Инвадоподии (подосомы)
Yamaguchi et al., Current Opinion in Cell Biology, 2005
Revach OY,
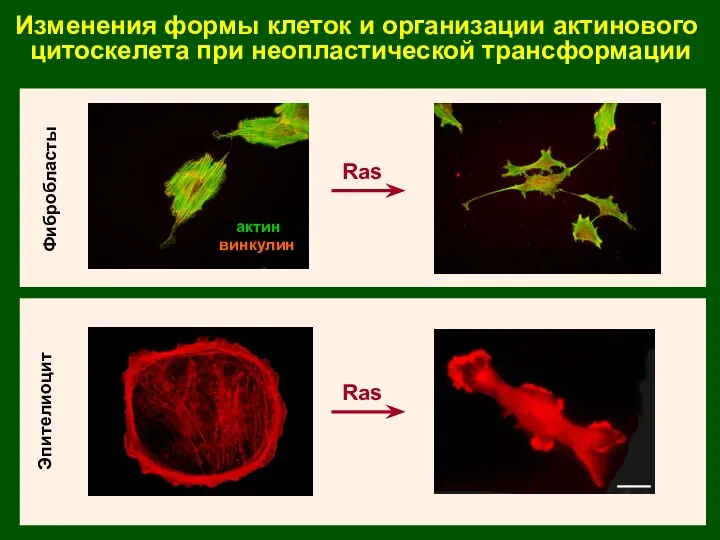
Изменения формы клеток и организации актинового
цитоскелета при неопластической трансформации
Эпителиоцит
Фибробласты
Ras
актин
винкулин
IAR2-C4
Ras
Изменения формы клеток и организации актинового
цитоскелета при неопластической трансформации
Эпителиоцит
Фибробласты
Ras
актин
винкулин
IAR2-C4
Ras
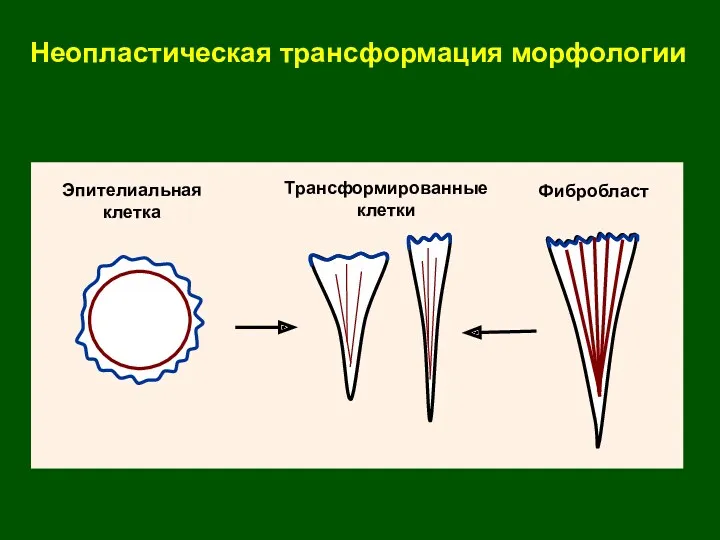
Эпителиальная
клетка
Фибробласт
Трансформированные
клетки
Неопластическая трансформация морфологии
Эпителиальная
клетка
Фибробласт
Трансформированные
клетки
Неопластическая трансформация морфологии
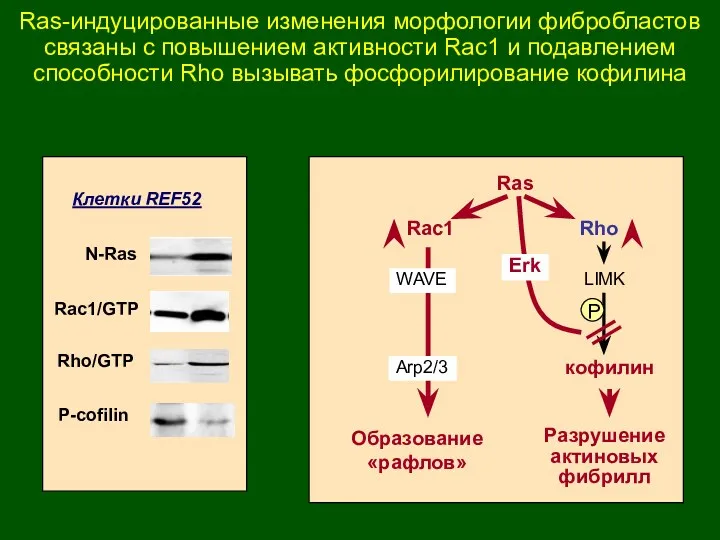
N-Ras
Rac1/GTP
Rho/GTP
P-cofilin
Клетки REF52
Rho
Ras
Rac1
LIMK
кофилин
Образование
«рафлов»
Разрушение
актиновых
фибрилл
P
Erk
WAVE
Arp2/3
Ras-индуцированные изменения морфологии фибробластов связаны с повышением активности Rac1 и
N-Ras
Rac1/GTP
Rho/GTP
P-cofilin
Клетки REF52
Rho
Ras
Rac1
LIMK
кофилин
Образование
«рафлов»
Разрушение
актиновых
фибрилл
P
Erk
WAVE
Arp2/3
Ras-индуцированные изменения морфологии фибробластов связаны с повышением активности Rac1 и
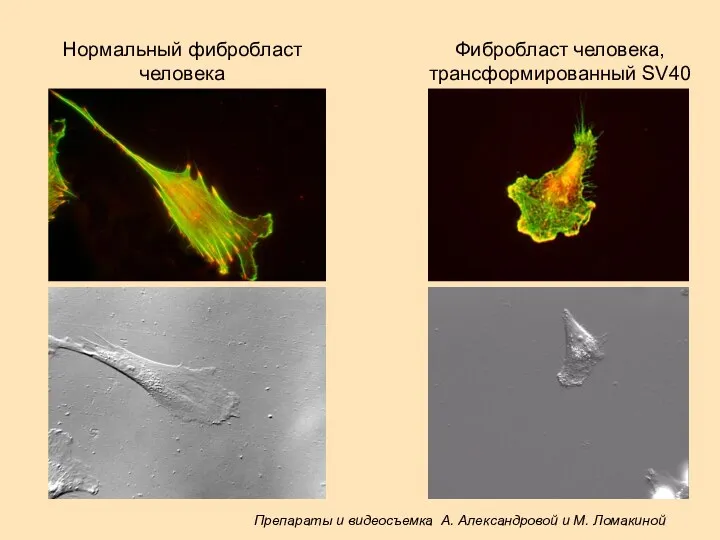
Нормальный фибробласт
человека
Фибробласт человека,
трансформированный SV40
Препараты и видеосъемка А. Александровой и М. Ломакиной
Нормальный фибробласт
человека
Фибробласт человека,
трансформированный SV40
Препараты и видеосъемка А. Александровой и М. Ломакиной
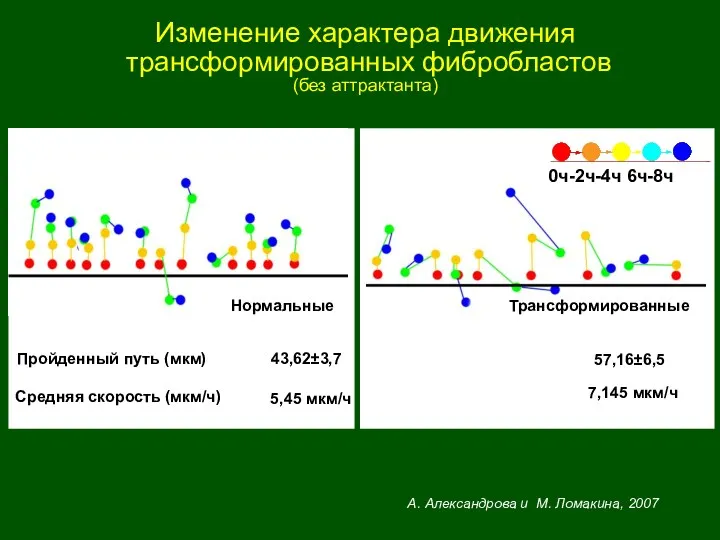
Изменение характера движения
трансформированных фибробластов
(без аттрактанта)
А. Александрова и М. Ломакина, 2007
0ч-2ч-4ч
Изменение характера движения
трансформированных фибробластов
(без аттрактанта)
А. Александрова и М. Ломакина, 2007
0ч-2ч-4ч

Тесты на миграционную и инвазивную способность клеток in vitro: миграция через
Тесты на миграционную и инвазивную способность клеток in vitro: миграция через
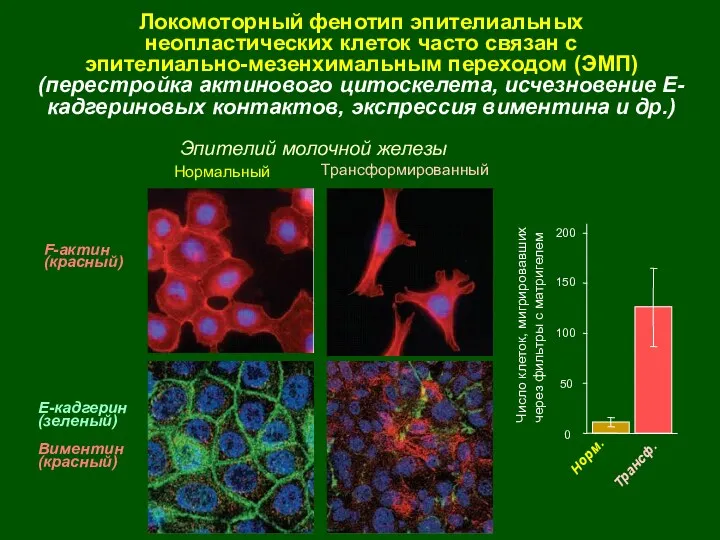
Е-кадгерин
(зеленый)
Виментин
(красный)
F-актин
(красный)
Локомоторный фенотип эпителиальных
неопластических клеток часто связан с
эпителиально-мезенхимальным
Е-кадгерин
(зеленый)
Виментин
(красный)
F-актин
(красный)
Локомоторный фенотип эпителиальных
неопластических клеток часто связан с
эпителиально-мезенхимальным
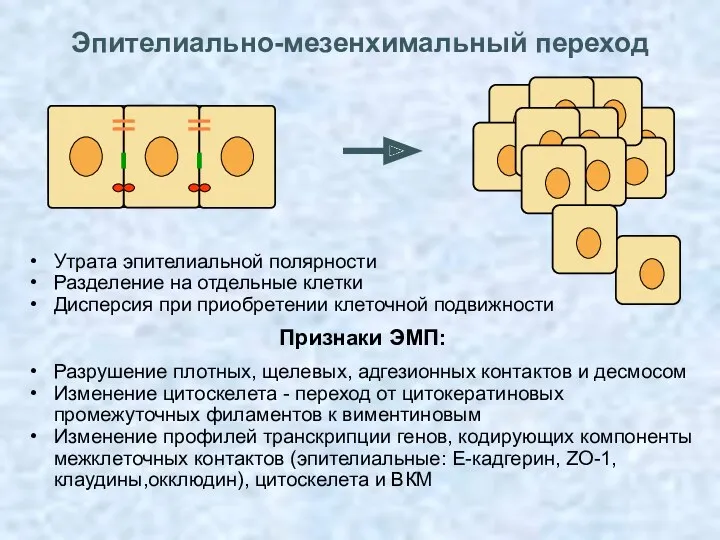
Эпителиально-мезенхимальный переход
Утрата эпителиальной полярности
Разделение на отдельные клетки
Дисперсия при приобретении клеточной
Эпителиально-мезенхимальный переход
Утрата эпителиальной полярности
Разделение на отдельные клетки
Дисперсия при приобретении клеточной
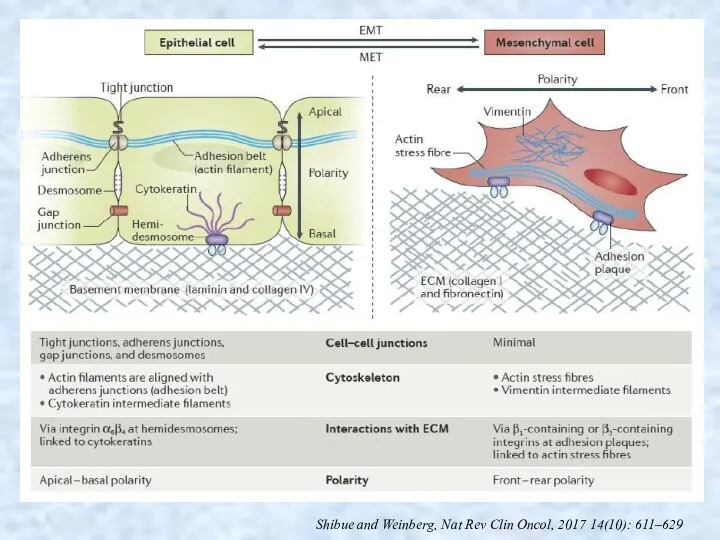
Shibue and Weinberg, Nat Rev Clin Oncol, 2017 14(10): 611–629
Shibue and Weinberg, Nat Rev Clin Oncol, 2017 14(10): 611–629
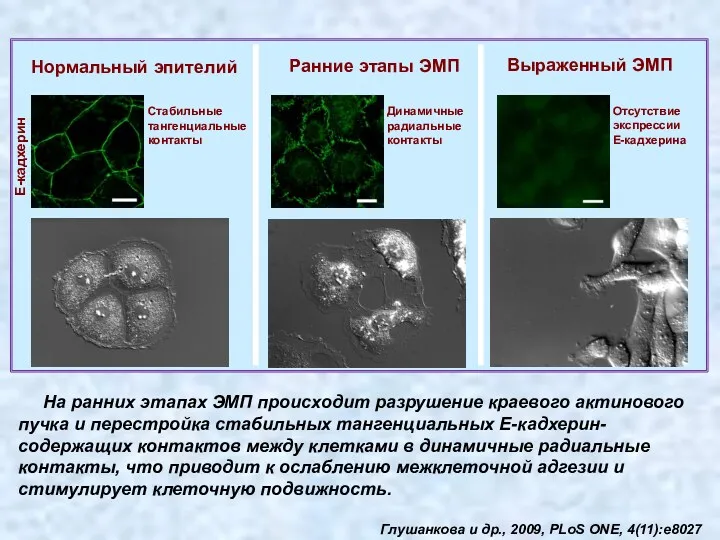
На ранних этапах ЭМП происходит разрушение краевого актинового пучка и
На ранних этапах ЭМП происходит разрушение краевого актинового пучка и
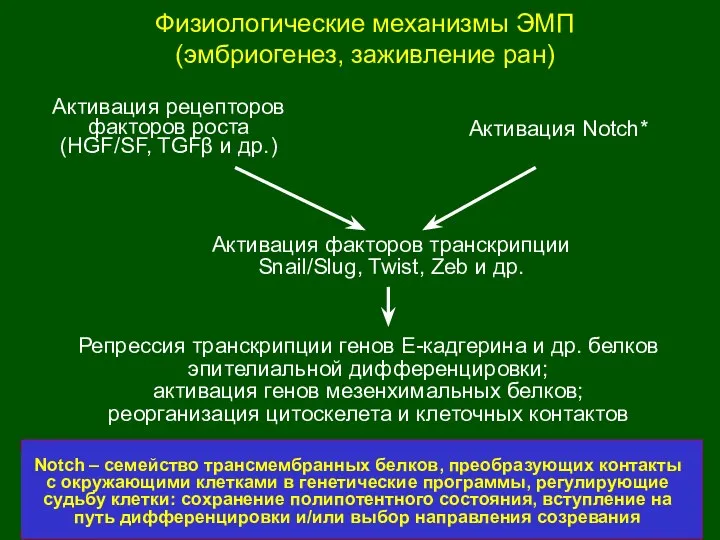
Физиологические механизмы ЭМП
(эмбриогенез, заживление ран)
Активация рецепторов
факторов роста (HGF/SF, TGFβ и др.)
Активация
Физиологические механизмы ЭМП
(эмбриогенез, заживление ран)
Активация рецепторов
факторов роста (HGF/SF, TGFβ и др.)
Активация
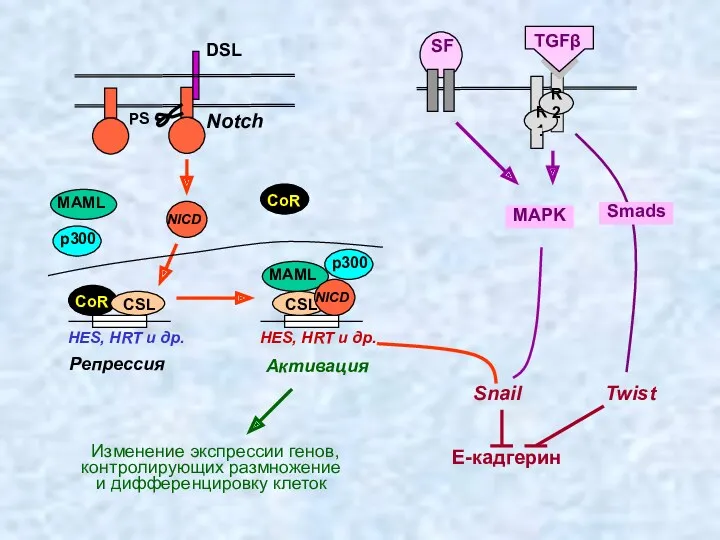
Репрессия
HES, HRT и др.
Notch
HES, HRT и др.
CSL
MAML
PS
p300
CoR
MAML
p300
CoR
CSL
Активация
Изменение экспрессии генов,
контролирующих размножение
Репрессия
HES, HRT и др.
Notch
HES, HRT и др.
CSL
MAML
PS
p300
CoR
MAML
p300
CoR
CSL
Активация
Изменение экспрессии генов,
контролирующих размножение
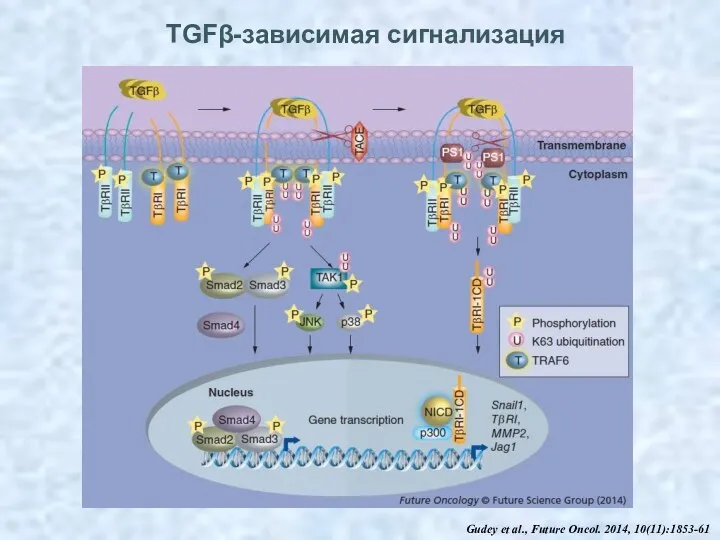
TGFβ-зависимая сигнализация
Gudey et al., Future Oncol. 2014, 10(11):1853-61
TGFβ-зависимая сигнализация
Gudey et al., Future Oncol. 2014, 10(11):1853-61
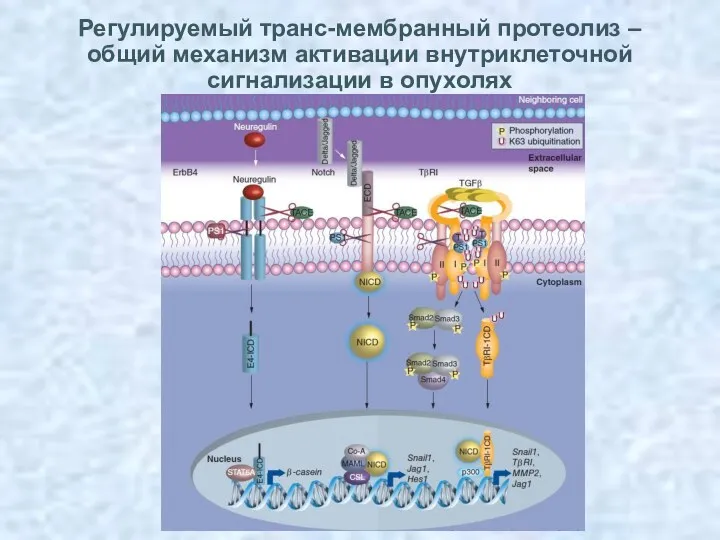
Регулируемый транс-мембранный протеолиз – общий механизм активации внутриклеточной сигнализации в опухолях
Регулируемый транс-мембранный протеолиз – общий механизм активации внутриклеточной сигнализации в опухолях
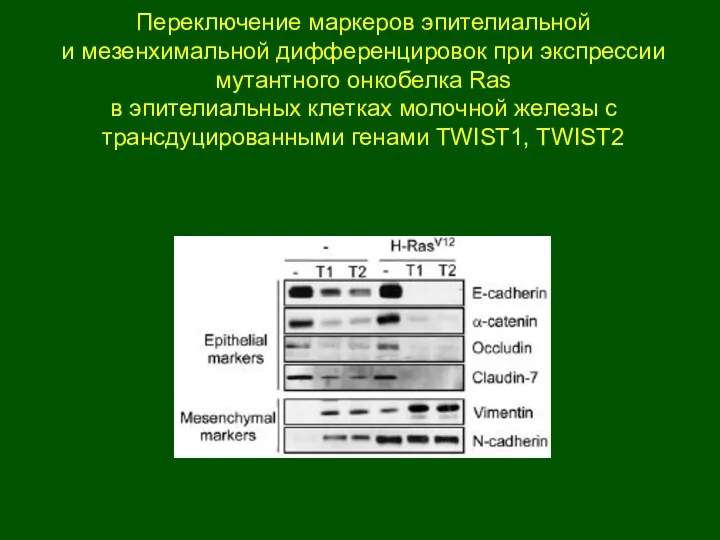
Переключение маркеров эпителиальной
и мезенхимальной дифференцировок при экспрессии мутантного онкобелка Ras
в
Переключение маркеров эпителиальной
и мезенхимальной дифференцировок при экспрессии мутантного онкобелка Ras
в
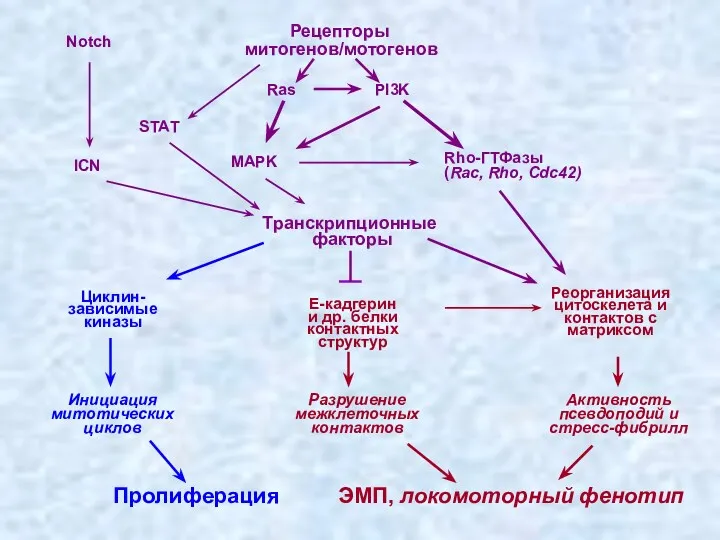
ЭМП, локомоторный фенотип
Реорганизация
цитоскелета и
контактов с
матриксом
Активность
псевдоподий и
стресс-фибрилл
Разрушение
межклеточных
контактов
Е-кадгерин
и
ЭМП, локомоторный фенотип
Реорганизация
цитоскелета и
контактов с
матриксом
Активность
псевдоподий и
стресс-фибрилл
Разрушение
межклеточных
контактов
Е-кадгерин
и

Механизмы ЭМП
Гиперметилирование, инактивирующие мутации гена Е-кадгерина
Активация рецепторов HGF/SF,
TGFβ и
Механизмы ЭМП
Гиперметилирование, инактивирующие мутации гена Е-кадгерина
Активация рецепторов HGF/SF,
TGFβ и
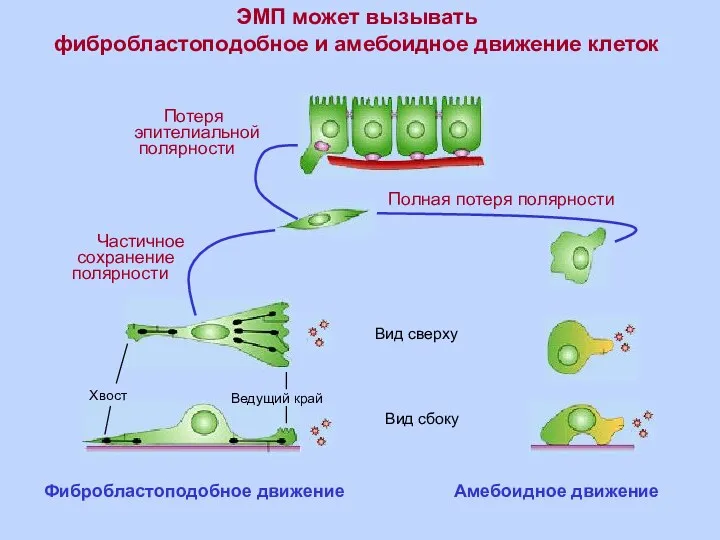
Амебоидное движение
Фибробластоподобное движение
ЭМП может вызывать
фибробластоподобное и амебоидное движение клеток
Вид
Амебоидное движение
Фибробластоподобное движение
ЭМП может вызывать
фибробластоподобное и амебоидное движение клеток
Вид
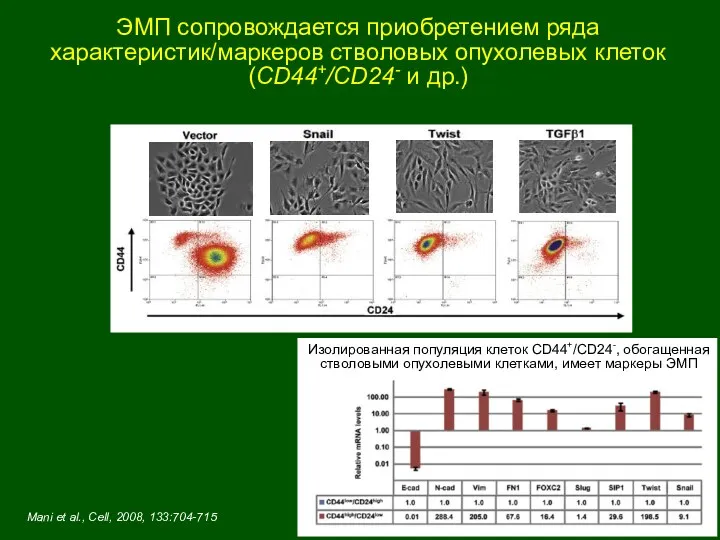
ЭМП сопровождается приобретением ряда характеристик/маркеров стволовых опухолевых клеток (CD44+/CD24- и др.)
Mani
ЭМП сопровождается приобретением ряда характеристик/маркеров стволовых опухолевых клеток (CD44+/CD24- и др.)
Mani
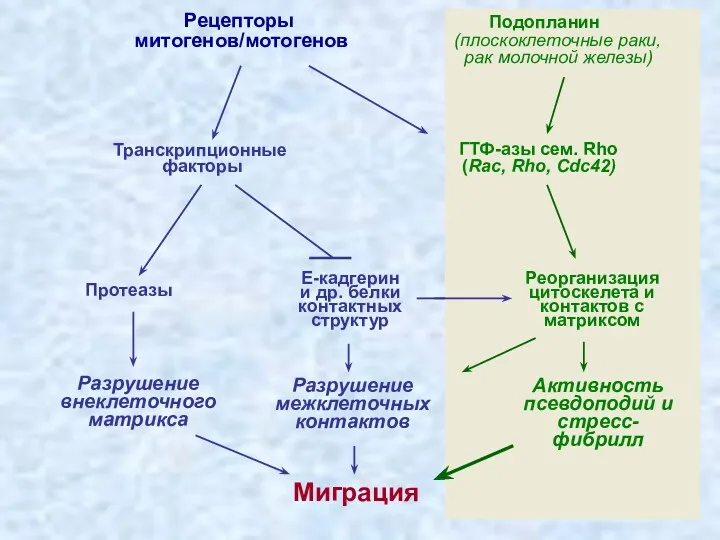
Транскрипционные
факторы
Рецепторы
митогенов/мотогенов
ГТФ-азы сем. Rho
(Rac, Rho, Cdc42)
Миграция
Реорганизация
цитоскелета и
контактов
Транскрипционные
факторы
Рецепторы
митогенов/мотогенов
ГТФ-азы сем. Rho
(Rac, Rho, Cdc42)
Миграция
Реорганизация
цитоскелета и
контактов
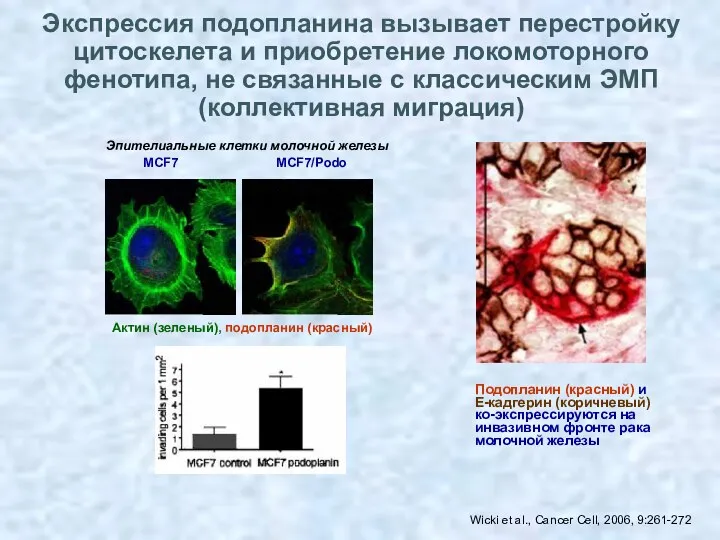
Wicki et al., Cancer Cell, 2006, 9:261-272
Подопланин (красный) и
Е-кадгерин (коричневый)
ко-экспрессируются
Wicki et al., Cancer Cell, 2006, 9:261-272
Подопланин (красный) и
Е-кадгерин (коричневый)
ко-экспрессируются
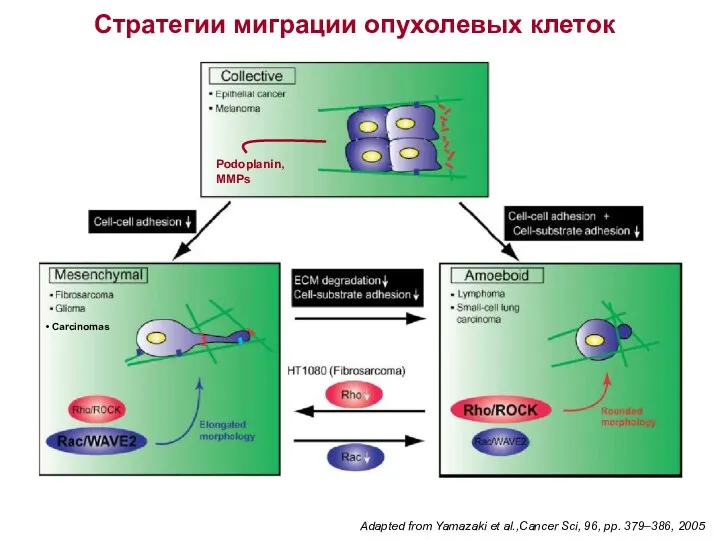
Adapted from Yamazaki et al.,Cancer Sci, 96, pp. 379–386, 2005
Стратегии
Adapted from Yamazaki et al.,Cancer Sci, 96, pp. 379–386, 2005
Стратегии
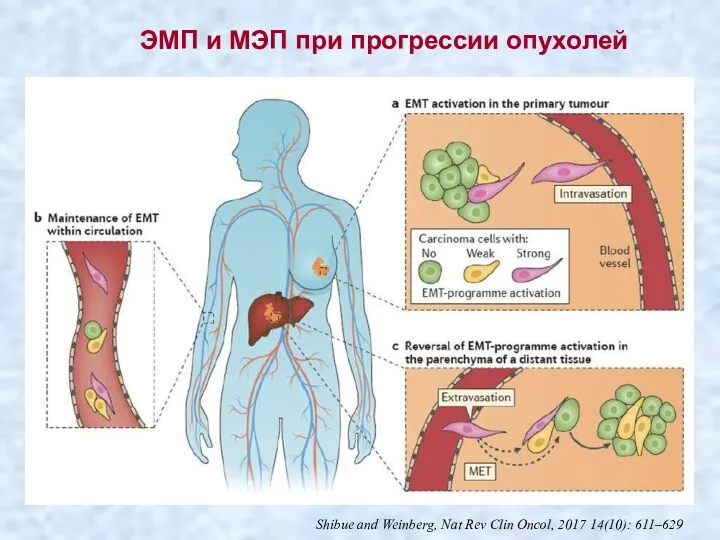
Shibue and Weinberg, Nat Rev Clin Oncol, 2017 14(10): 611–629
ЭМП
Shibue and Weinberg, Nat Rev Clin Oncol, 2017 14(10): 611–629
ЭМП
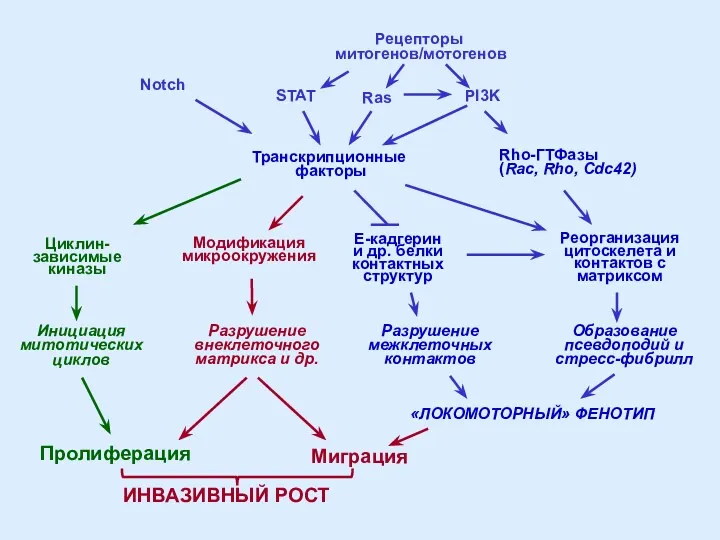
Rho-ГТФазы
(Rac, Rho, Cdc42)
Реорганизация
цитоскелета и
контактов с
матриксом
Образование
псевдоподий и
стресс-фибрилл
Разрушение
межклеточных
контактов
Транскрипционные
факторы
Rho-ГТФазы
(Rac, Rho, Cdc42)
Реорганизация
цитоскелета и
контактов с
матриксом
Образование
псевдоподий и
стресс-фибрилл
Разрушение
межклеточных
контактов
Транскрипционные
факторы
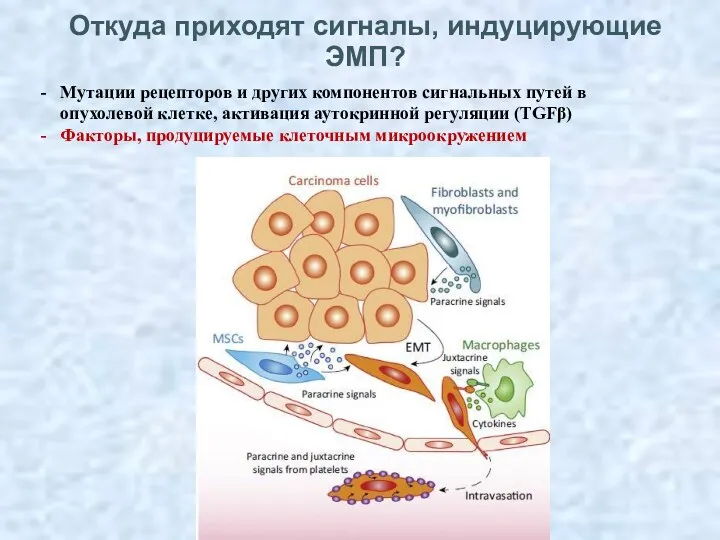
Откуда приходят сигналы, индуцирующие ЭМП?
Мутации рецепторов и других компонентов сигнальных путей
Откуда приходят сигналы, индуцирующие ЭМП?
Мутации рецепторов и других компонентов сигнальных путей
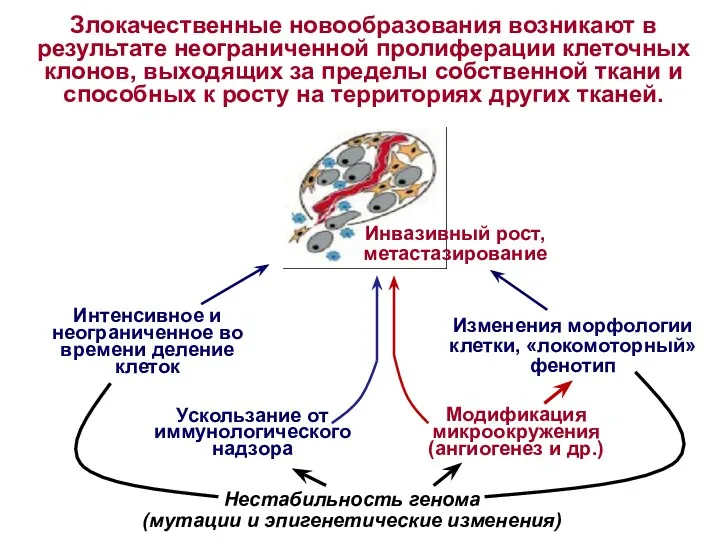
Злокачественные новообразования возникают в
результате неограниченной пролиферации клеточных клонов, выходящих за пределы
Злокачественные новообразования возникают в
результате неограниченной пролиферации клеточных клонов, выходящих за пределы
 Николай Федорович Гамалея
Николай Федорович Гамалея Cells
Cells Строение синапсов. Потенциал действия. Механизм проведения потенциала действия. Физиология боли, медиаторы боли
Строение синапсов. Потенциал действия. Механизм проведения потенциала действия. Физиология боли, медиаторы боли Всасывание, пищеварение
Всасывание, пищеварение Процесс кровообращения. Сердце, анатомия и физиология
Процесс кровообращения. Сердце, анатомия и физиология презентация к уроку биологии 7 класс
презентация к уроку биологии 7 класс Рост и развитие животных
Рост и развитие животных Строение эукариотической клетки
Строение эукариотической клетки Урок биологии в 9 классе по теме Антропогенез
Урок биологии в 9 классе по теме Антропогенез Домашние любимцы. Кошки и собаки
Домашние любимцы. Кошки и собаки Ознаки захворювання яблук
Ознаки захворювання яблук Пищевая микробиология. Молочные продукты
Пищевая микробиология. Молочные продукты Элективный курс Репродуктивное здоровье
Элективный курс Репродуктивное здоровье Отдел голосеменные
Отдел голосеменные Рыбы Байкала
Рыбы Байкала Тип Інфузорії
Тип Інфузорії Растения Дальнего Востока
Растения Дальнего Востока Полезный мед - натуральный мед
Полезный мед - натуральный мед Цитологические основы наследования признаков при моногибридном скрещивании
Цитологические основы наследования признаков при моногибридном скрещивании Моллюски осьминоги
Моллюски осьминоги Nervous sistem
Nervous sistem Домашние питомцы
Домашние питомцы Роль генетически модифицированных организмов на организм человека
Роль генетически модифицированных организмов на организм человека Молекулярно-биологические методы диагностики
Молекулярно-биологические методы диагностики Птицы
Птицы Витамины в составе фруктов и овощей
Витамины в составе фруктов и овощей Голосеменные растения
Голосеменные растения Возникновение жизни
Возникновение жизни