- Катаболизм водных организмов. Лекция № 6

Содержание
- 2. Катаболизм или энергетический обмен метаболический распад на более простые вещества окисление протекает с высвобождением энергии Примеры
- 3. Интенсивность катаболических процессов и преобладание тех или иных катаболических процессов в качестве источников энергии в клетках
- 5. Катаболизм глюкозы - основной источник энергии Окисление глюкозы до СО2 и Н2О (аэробный распад). С6Н12О6 +
- 6. Аэробный гликолиз процесс окисления глюкозы до пировиноградной кислоты протекает в присутствии О2 Ферменты, катализирующие реакции этого
- 7. Этапы аэробного гликолиза глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с
- 8. Этапы аэробного гликолиза Этап синтеза АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват.
- 9. Схема аэробного гликолиза Глюкоза Глюкозо-6-фосфат фруктозо-6-фосфат (глюкозофосфатизомераза) фруктозо-1,6-бисфосфат (фосфофруктокиназа) реакция необратима наиболее медленная 2 триозофосфата фруктозобисфосфатальдолаза
- 10. Субстратное фосфорилирование Окисление триозофосфата восстановлению NAD*Н образование с участием Н3РО4 высокоэнергетической связи фосфат передаётся на АДФ
- 11. Итог аэробного гликолиза Распад 1 6С молекулы (глюкоза) до 2 3С молекул (пируват) Синтезируется 2 АТФ
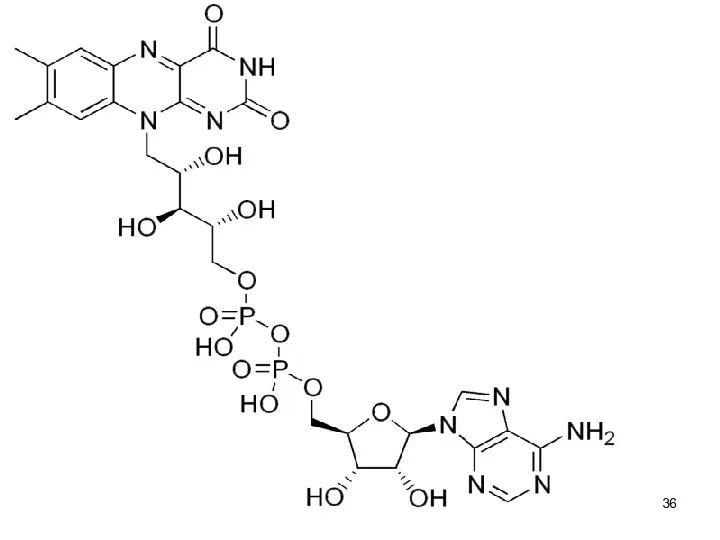
- 13. NAD
- 14. Стадии гликолиза
- 15. Стадии гликолиза
- 16. Стадии гликолиза
- 17. Стадии гликолиза
- 18. Стадии гликолиза
- 19. Стадии гликолиза
- 20. Стадии гликолиза
- 21. Стадии гликолиза
- 22. II.Образование Ацетил-КоА Углеводы, жирные кислоты и аминокислоты окисляются через цикл Кребса до СО2 и Н2О. превращаются
- 23. Образование Ацетил-КоА Окислительное декарбоксилирование пирувата Пируват (3С) - пируватдегидрогеназный комплекс (3 фермента) - СО2 и ацетальдегид
- 25. III.Цикл Кребса — это ключевой этап дыхания всех клеток, использующих кислород пересечение множества метаболических путей в
- 26. У эукариот все реакции цикла Кребса протекают внутри митохондрий ферменты - в свободном состоянии в митохондриальном
- 30. 1 Оксалоацетат (щавелевоуксусная кислота)+ Ацетил-КoA + H2O Цитратсинтетаза 4С + 2С - 6С Лимонная кислота +
- 31. 2 Цитрат (лимонная кислота) аконитаза 6С – 6С Изоцитрат (Изолимонная кислота)
- 32. 3 Изоцитрат + NAD+ Изоцитратдегидрогена-за декарбоксилирующая 6С – 5С α-кетоглутарат+ CO2 + NADH + H+
- 33. 4 α-кетоглутарат+ NAD+ + КoA α-кетоглутарат-дегидрогеназа 5С – 4С сукцинил-КoA + (янтарная к-та) + NADH +
- 34. 5 сукцинил-КoA сукцинилкофермент А синтетаза 4С – 4С сукцинат + КoA + АТФ – субстратное фосфорилирование
- 35. 6 Сукцинат + FAD Сукцинатдегидроге-наза Фумарат + FAD*Н2
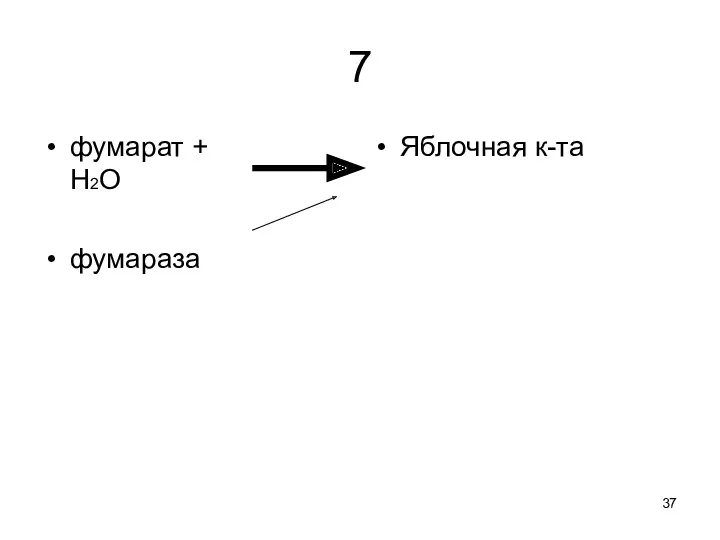
- 37. 7 фумарат + H2O фумараза Яблочная к-та
- 38. 8 Яблочная к-та + NAD+ Дегидрогеназа яблочной кислоты Оксалоацетат (щавелевоуксусная к-та) + NADH + H+
- 39. Запомнить Целый Ананас И Кусочек Суфле Сегодня Фактически Мой Обед, что соответствует ряду — цитрат, (цис-)аконитат,
- 40. Регуляция Цикл Кребса регулируется «по механизму отрицательной обратной связи» при наличии большого количества субстратов (ацетил-КоА, оксалоацетат),
- 41. Функции Интегративная функция — связующее звено между реакциями анаболизма и катаболизма. Катаболическая функция — превращение различных
- 42. Функции Анаболическая функция — использование субстратов цикла на синтез органических веществ: Оксалацетат — глюкоза, Асп, Асн.
- 43. Итог В процессе ферментативного окисления орг. в-в освобождается энергия (аэробный гликолиз, обр. Ац-СоА) электроны и протоны
- 44. IV.Дыхательная цепь Электроны - обладают высоким энергетическим потенциалом - NAD*H и FAD*H2 Электроны от восстановленных коферментов
- 45. Дыхательная цепь цепь переноса электронов (ЦПЭ) - дыхательная цепь. Восстановление молекулы О2 происходит в результате переноса
- 47. Дыхательная цепь Электроны, поступающие в ЦПЭ, по мере их продвижения от одного переносчика к другому теряют
- 48. Процессы перераспределения свободной энергии перенос электронов от восстановленных коферментов НАД*H и ФАД*H2 через ЦПЭ на кислород
- 49. фосфорилирование АДФ, или синтез АТФ АДФ + Н3РО4+7,3 ккал/моль (30,5 кДж/моль) = АТФ + Н2О.
- 50. ЦПЭ ЦПЭ – ферментный комплекс, окисляющий НАД*H и ФАД*H2 и использует энергию окисления (передачи электронов) для
- 51. ЦПЭ Комплекс III (Цитохром bc1 комплекс) переносит электроны с убихинола на два водорастворимых цитохрома с, расположенных
- 52. Окислительное фосфорилирование Процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — протонной АТФ-синтазой Поток протонов
- 54. Полифенолазы и пероксидазы в энергетическом обмене Полифенолазы и пероксидазы катализируют окисление фенольных соединений. В присутствии этих
- 55. Пероксидаза — фермент, катализирующий окисление полифенолов и некоторых ароматических аминов при помощи кислорода, перекиси водорода или
- 56. Полифенолоксидаза и пероксидаза являются конечными оксидазами и характеризуют интенсивность заключительной фазы дыхания. Согласно имеющимся представлениям, оксидазная
- 57. пероксидаза — чувствительный индикатор, регистрирующий изменения в газообмене, и может быть использована для экспресс-диагностики отравления рыб
- 59. Скачать презентацию

Катаболизм
или энергетический обмен
метаболический распад на более простые вещества
окисление
протекает с высвобождением энергии
Примеры катаболизма
превращение этанола через стадии ацетальдегида (этаналя)
Катаболизм
или энергетический обмен
метаболический распад на более простые вещества
окисление
протекает с высвобождением энергии
Примеры катаболизма
превращение этанола через стадии ацетальдегида (этаналя)

Интенсивность катаболических процессов
и преобладание тех или иных катаболических процессов в качестве
Интенсивность катаболических процессов
и преобладание тех или иных катаболических процессов в качестве


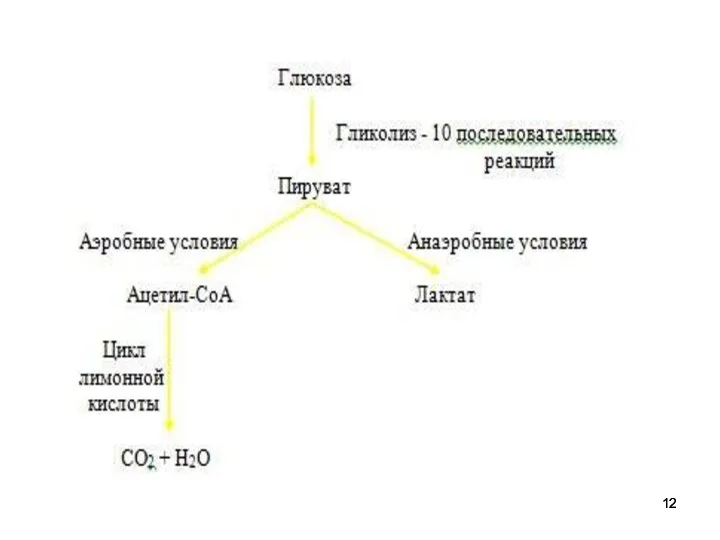
Катаболизм глюкозы - основной источник энергии
Окисление глюкозы до СО2 и Н2О
Катаболизм глюкозы - основной источник энергии
Окисление глюкозы до СО2 и Н2О

Аэробный гликолиз
процесс окисления глюкозы до пировиноградной кислоты
протекает в присутствии О2
Аэробный гликолиз
процесс окисления глюкозы до пировиноградной кислоты
протекает в присутствии О2
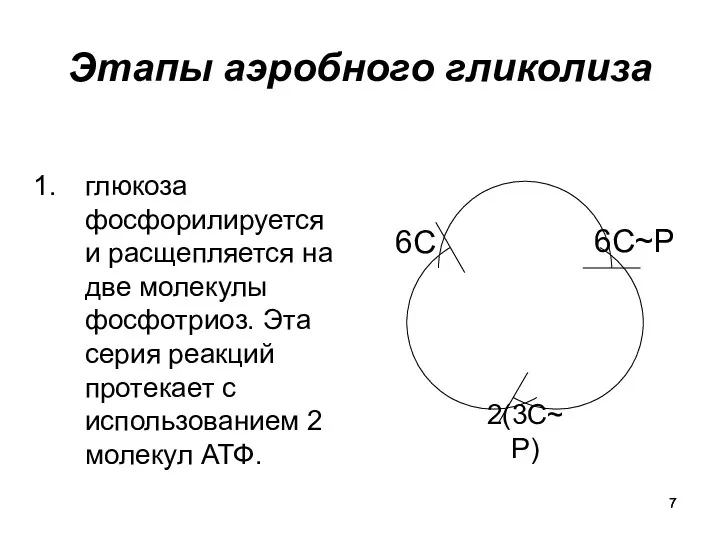
Этапы аэробного гликолиза
глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта
Этапы аэробного гликолиза
глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта

Этапы аэробного гликолиза
Этап синтеза АТФ. В результате этой серии реакций фосфотриозы
Этапы аэробного гликолиза
Этап синтеза АТФ. В результате этой серии реакций фосфотриозы

Схема аэробного гликолиза
Глюкоза
Глюкозо-6-фосфат
фруктозо-6-фосфат (глюкозофосфатизомераза)
фруктозо-1,6-бисфосфат (фосфофруктокиназа) реакция необратима наиболее медленная
2
Схема аэробного гликолиза
Глюкоза
Глюкозо-6-фосфат
фруктозо-6-фосфат (глюкозофосфатизомераза)
фруктозо-1,6-бисфосфат (фосфофруктокиназа) реакция необратима наиболее медленная
2

Субстратное фосфорилирование
Окисление триозофосфата
восстановлению NAD*Н
образование с участием Н3РО4 высокоэнергетической связи
фосфат
Субстратное фосфорилирование
Окисление триозофосфата
восстановлению NAD*Н
образование с участием Н3РО4 высокоэнергетической связи
фосфат

Итог аэробного гликолиза
Распад 1 6С молекулы (глюкоза) до 2 3С молекул
Итог аэробного гликолиза
Распад 1 6С молекулы (глюкоза) до 2 3С молекул
NAD
NAD
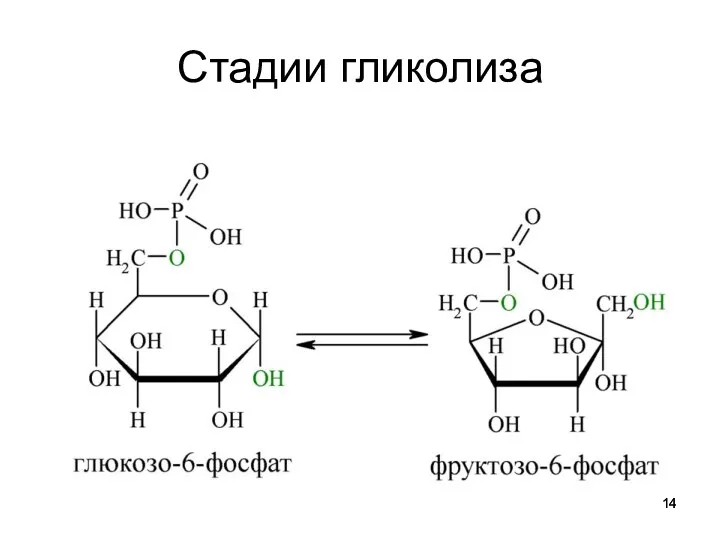
Стадии гликолиза
Стадии гликолиза
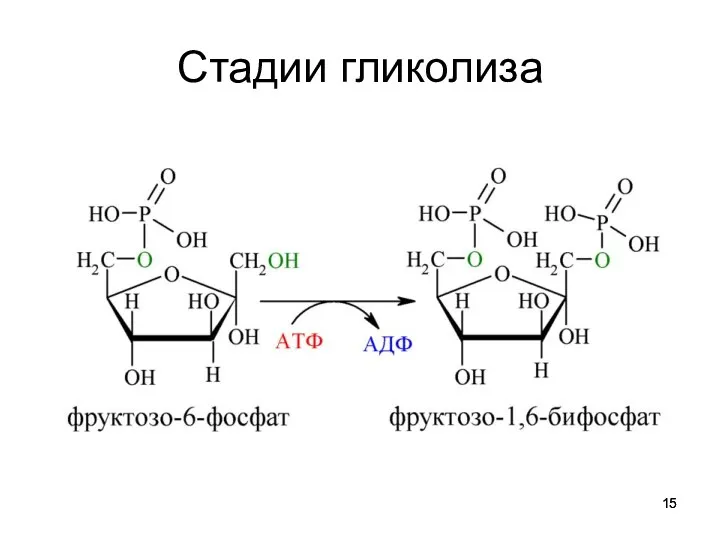
Стадии гликолиза
Стадии гликолиза
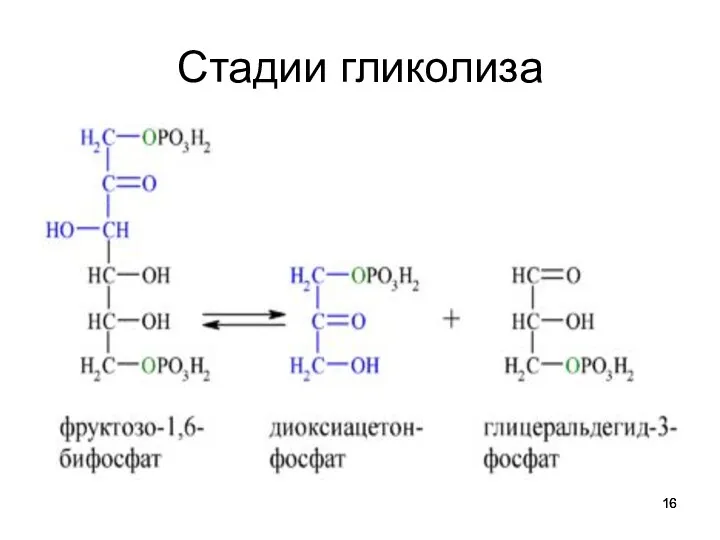
Стадии гликолиза
Стадии гликолиза
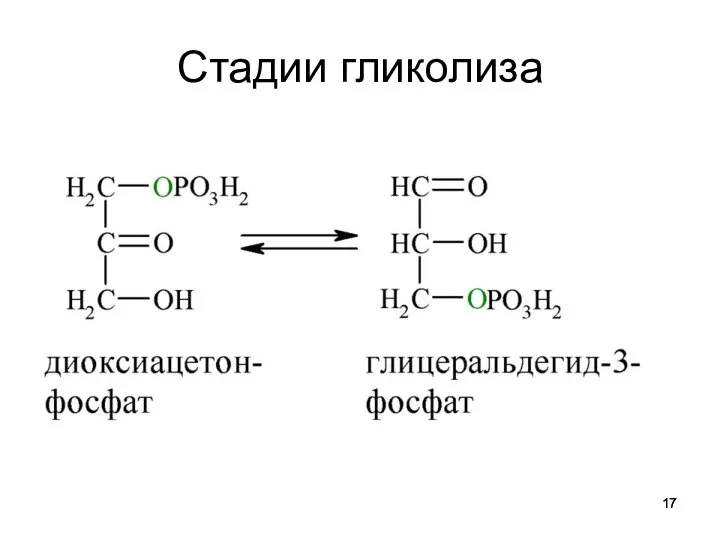
Стадии гликолиза
Стадии гликолиза
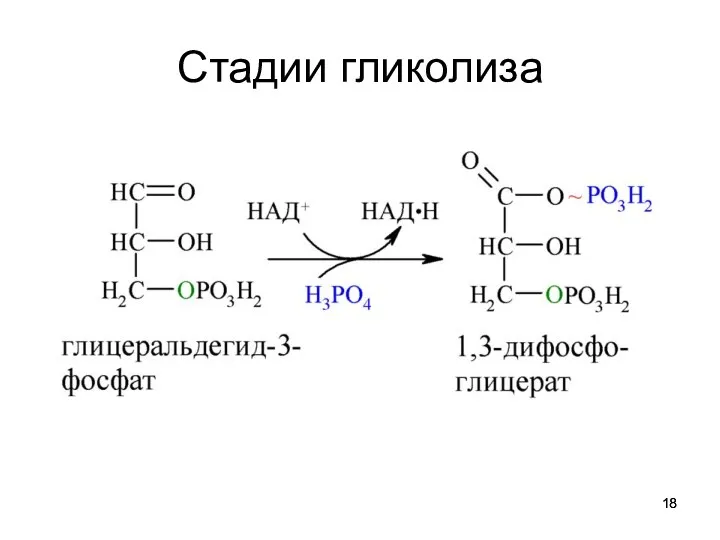
Стадии гликолиза
Стадии гликолиза
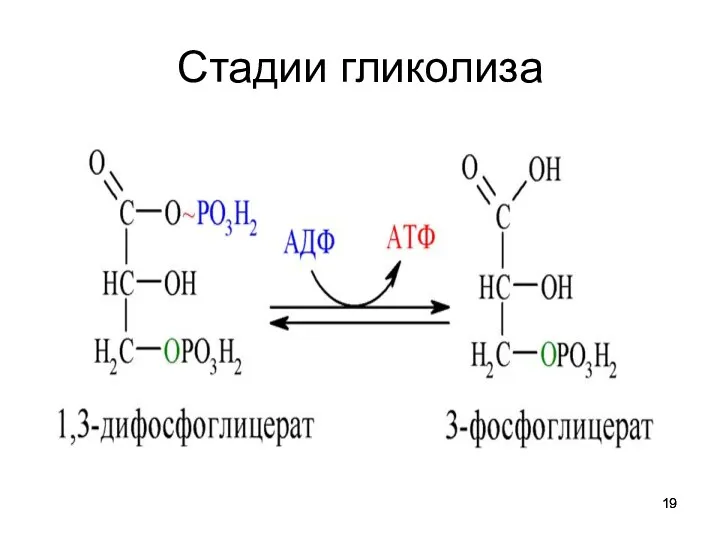
Стадии гликолиза
Стадии гликолиза
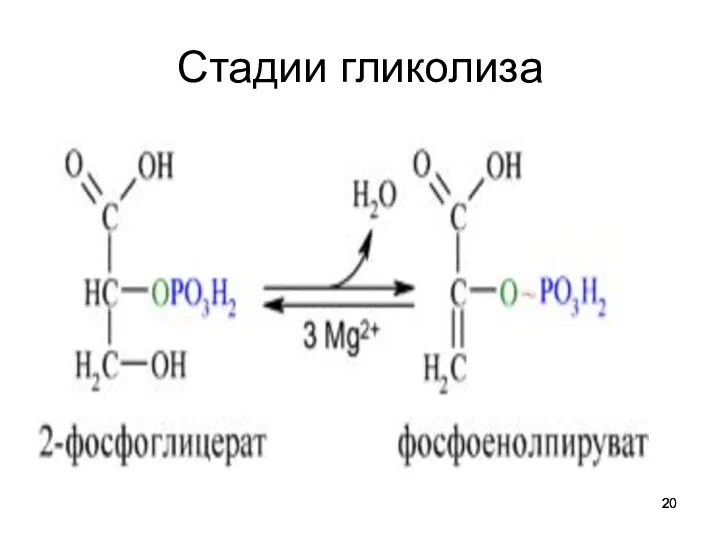
Стадии гликолиза
Стадии гликолиза
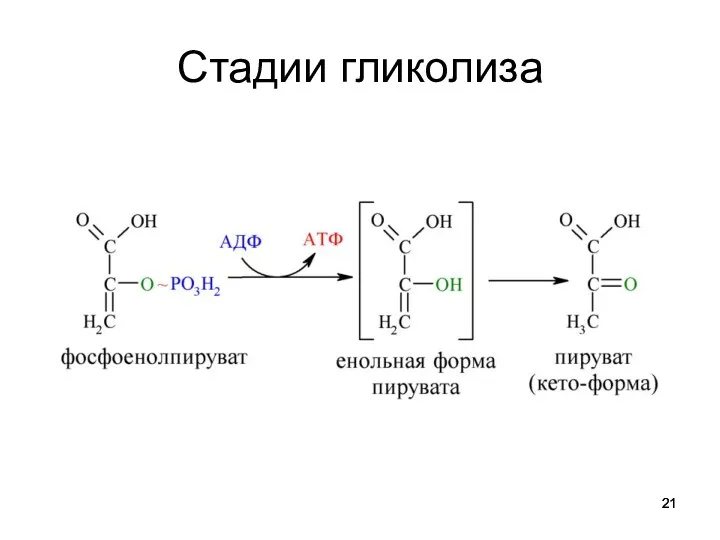
Стадии гликолиза
Стадии гликолиза

II.Образование Ацетил-КоА
Углеводы, жирные кислоты и аминокислоты окисляются через цикл Кребса до
II.Образование Ацетил-КоА
Углеводы, жирные кислоты и аминокислоты окисляются через цикл Кребса до

Образование Ацетил-КоА
Окислительное декарбоксилирование пирувата
Пируват (3С) - пируватдегидрогеназный комплекс (3 фермента) - СО2
Образование Ацетил-КоА
Окислительное декарбоксилирование пирувата
Пируват (3С) - пируватдегидрогеназный комплекс (3 фермента) - СО2
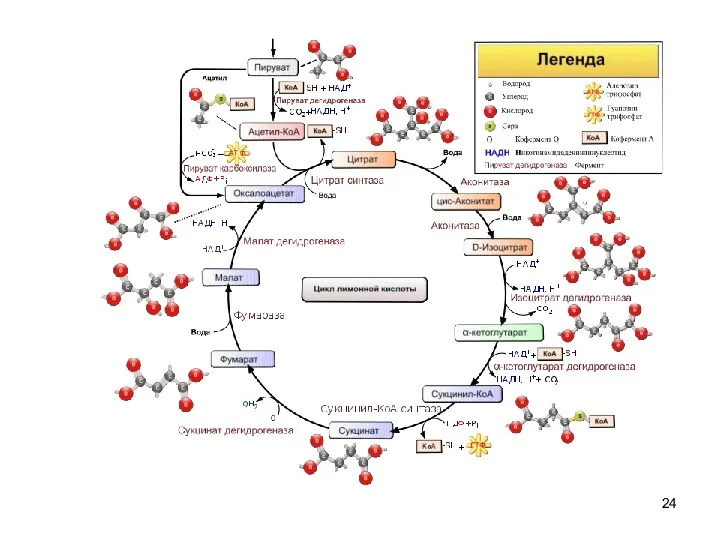

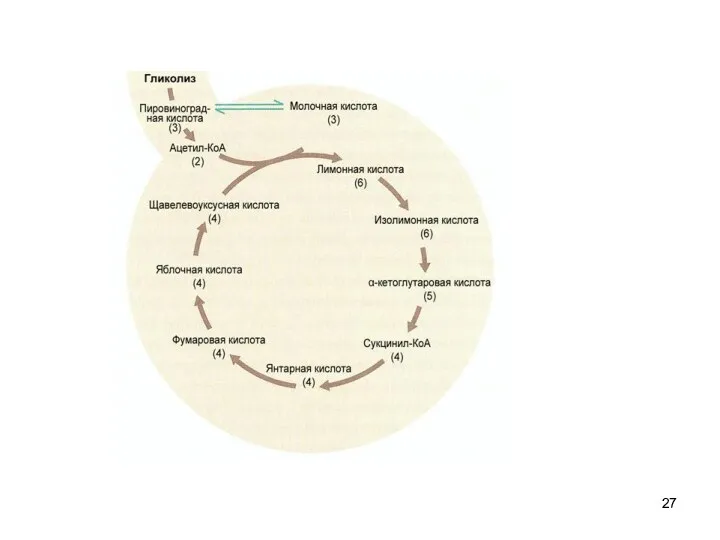
III.Цикл Кребса
— это ключевой этап дыхания всех клеток, использующих кислород
пересечение множества метаболических
III.Цикл Кребса
— это ключевой этап дыхания всех клеток, использующих кислород
пересечение множества метаболических

У эукариот
все реакции цикла Кребса протекают внутри митохондрий
ферменты - в свободном состоянии в
У эукариот
все реакции цикла Кребса протекают внутри митохондрий
ферменты - в свободном состоянии в

1
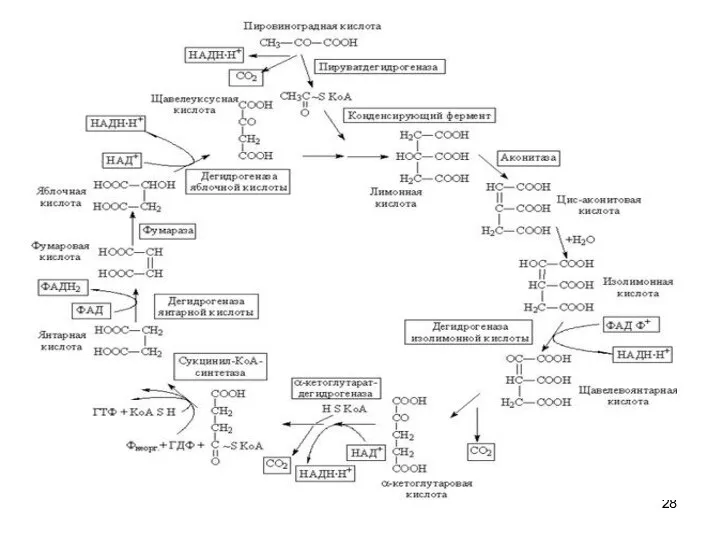
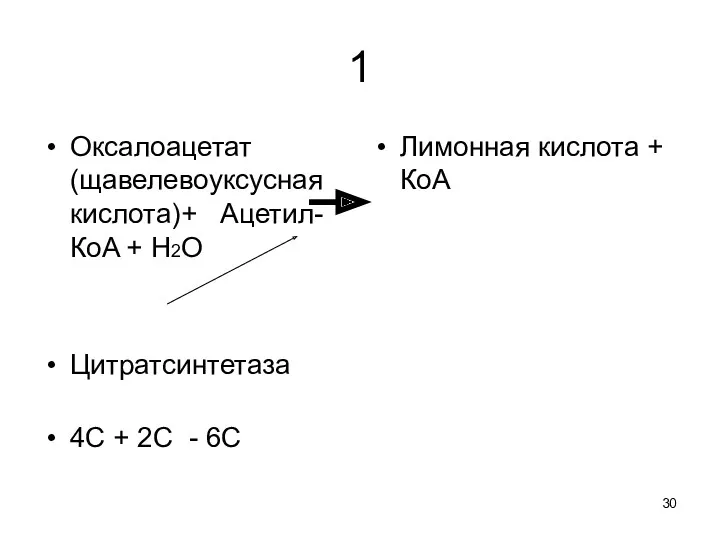
Оксалоацетат (щавелевоуксусная кислота)+ Ацетил-КoA + H2O
Цитратсинтетаза
4С + 2С -
1
Оксалоацетат (щавелевоуксусная кислота)+ Ацетил-КoA + H2O
Цитратсинтетаза
4С + 2С -
2
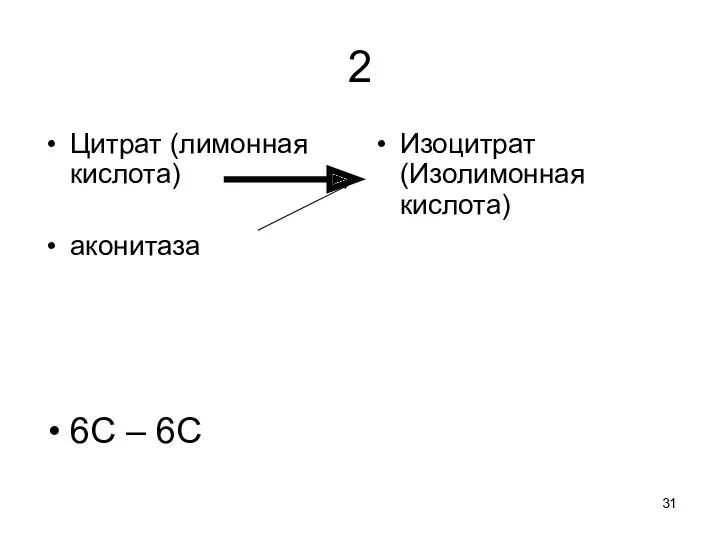
Цитрат (лимонная кислота)
аконитаза
6С – 6С
Изоцитрат (Изолимонная кислота)
2
Цитрат (лимонная кислота)
аконитаза
6С – 6С
Изоцитрат (Изолимонная кислота)
3
Изоцитрат +
NAD+
Изоцитратдегидрогена-за декарбоксилирующая
6С – 5С
α-кетоглутарат+
CO2 +
NADH + H+
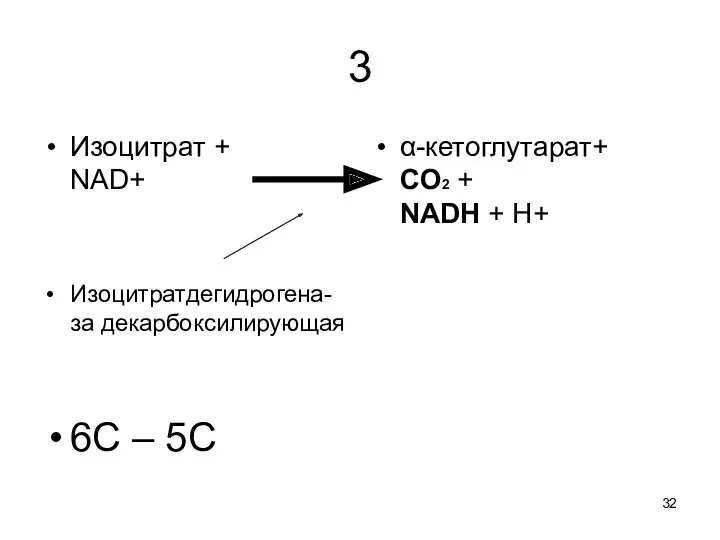
3
Изоцитрат +
NAD+
Изоцитратдегидрогена-за декарбоксилирующая
6С – 5С
α-кетоглутарат+
CO2 +
NADH + H+
4
α-кетоглутарат+
NAD+ +
КoA
α-кетоглутарат-дегидрогеназа
5С – 4С
сукцинил-КoA +
(янтарная к-та) + NADH + H+ +
CO2
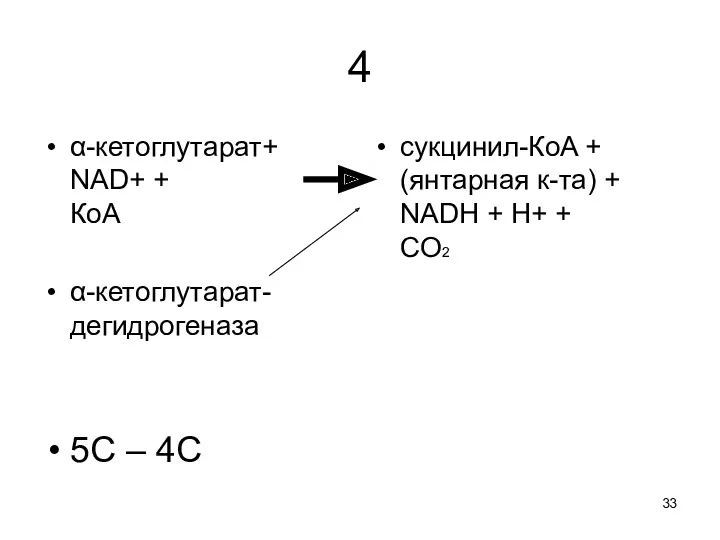
4
α-кетоглутарат+
NAD+ +
КoA
α-кетоглутарат-дегидрогеназа
5С – 4С
сукцинил-КoA +
(янтарная к-та) + NADH + H+ +
CO2
5
сукцинил-КoA
сукцинилкофермент А синтетаза
4С – 4С
сукцинат +
КoA +
АТФ – субстратное фосфорилирование
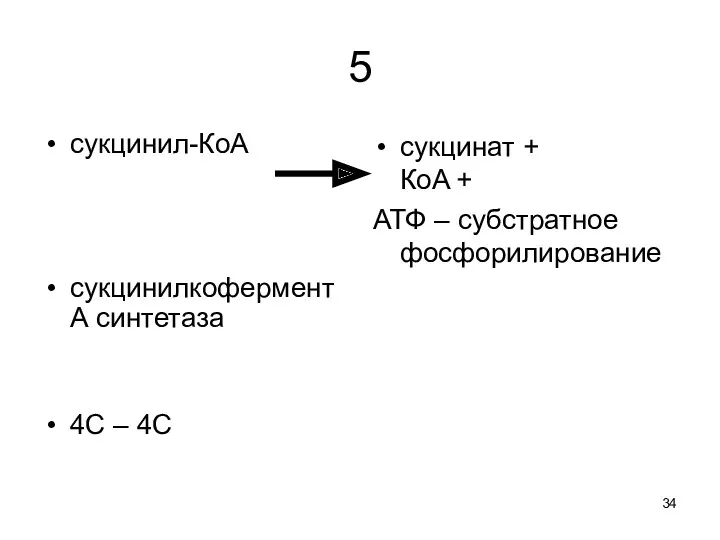
5
сукцинил-КoA
сукцинилкофермент А синтетаза
4С – 4С
сукцинат +
КoA +
АТФ – субстратное фосфорилирование
6
Сукцинат + FAD
Сукцинатдегидроге-наза
Фумарат + FAD*Н2
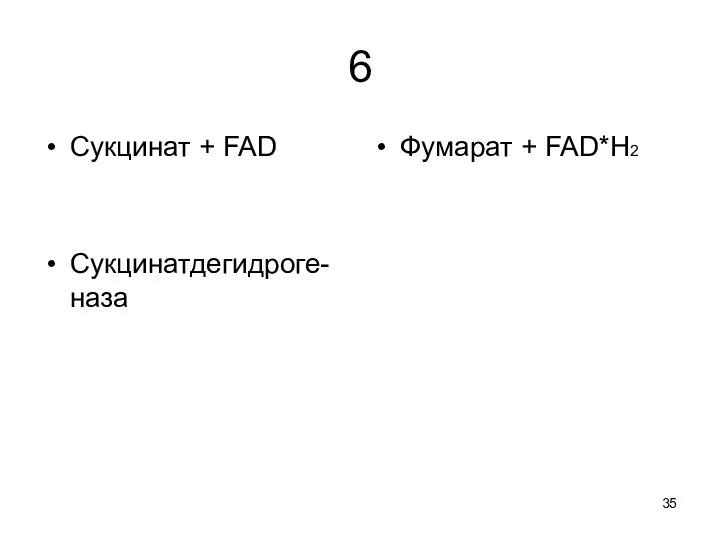
6
Сукцинат + FAD
Сукцинатдегидроге-наза
Фумарат + FAD*Н2
7
фумарат +
H2O
фумараза
Яблочная к-та
7
фумарат +
H2O
фумараза
Яблочная к-та
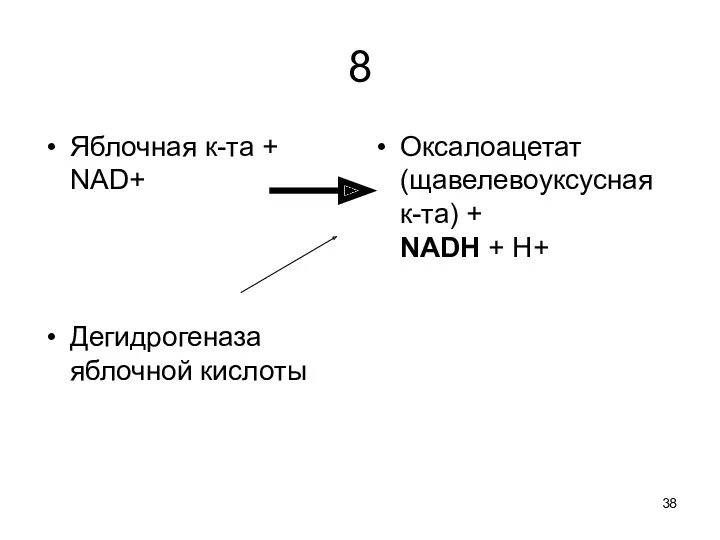
8
Яблочная к-та +
NAD+
Дегидрогеназа яблочной кислоты
Оксалоацетат (щавелевоуксусная к-та) +
NADH + H+
8
Яблочная к-та +
NAD+
Дегидрогеназа яблочной кислоты
Оксалоацетат (щавелевоуксусная к-та) + NADH + H+

Запомнить
Целый Ананас И Кусочек Суфле Сегодня Фактически Мой Обед, что соответствует
Запомнить
Целый Ананас И Кусочек Суфле Сегодня Фактически Мой Обед, что соответствует

Регуляция
Цикл Кребса регулируется «по механизму отрицательной обратной связи»
при наличии большого количества субстратов
Регуляция
Цикл Кребса регулируется «по механизму отрицательной обратной связи»
при наличии большого количества субстратов

Функции
Интегративная функция — связующее звено между реакциями анаболизма и катаболизма.
Катаболическая функция — превращение
Функции
Интегративная функция — связующее звено между реакциями анаболизма и катаболизма.
Катаболическая функция — превращение

Функции
Анаболическая функция — использование субстратов цикла на синтез органических веществ:
Оксалацетат — глюкоза, Асп, Асн.
Сукцинил-КоА —
Функции
Анаболическая функция — использование субстратов цикла на синтез органических веществ:
Оксалацетат — глюкоза, Асп, Асн.
Сукцинил-КоА —

Итог
В процессе ферментативного окисления орг. в-в освобождается энергия (аэробный гликолиз, обр.
Итог
В процессе ферментативного окисления орг. в-в освобождается энергия (аэробный гликолиз, обр.

IV.Дыхательная цепь
Электроны - обладают высоким энергетическим потенциалом - NAD*H и FAD*H2
Электроны
IV.Дыхательная цепь
Электроны - обладают высоким энергетическим потенциалом - NAD*H и FAD*H2
Электроны

Дыхательная цепь
цепь переноса электронов (ЦПЭ) - дыхательная цепь.
Восстановление молекулы О2
Дыхательная цепь
цепь переноса электронов (ЦПЭ) - дыхательная цепь.
Восстановление молекулы О2
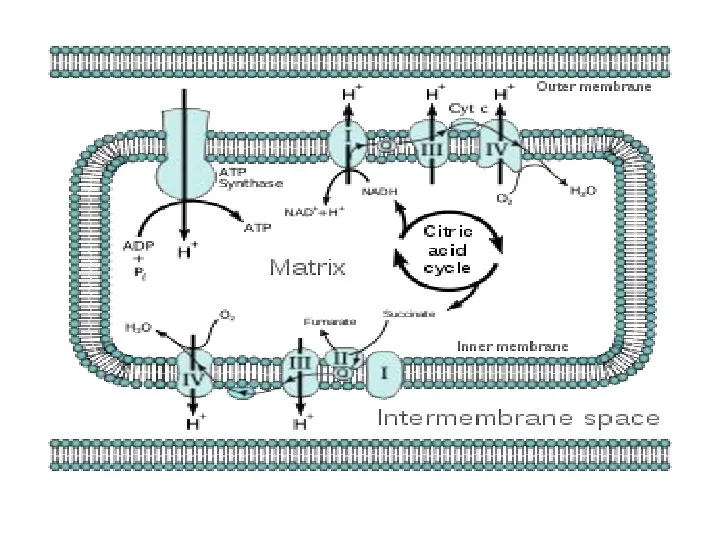

Дыхательная цепь
Электроны, поступающие в ЦПЭ, по мере их продвижения от одного
Дыхательная цепь
Электроны, поступающие в ЦПЭ, по мере их продвижения от одного

Процессы перераспределения свободной энергии
перенос электронов от восстановленных коферментов НАД*H и ФАД*H2
Процессы перераспределения свободной энергии
перенос электронов от восстановленных коферментов НАД*H и ФАД*H2

фосфорилирование АДФ, или синтез АТФ
АДФ + Н3РО4+7,3 ккал/моль (30,5 кДж/моль) =
фосфорилирование АДФ, или синтез АТФ
АДФ + Н3РО4+7,3 ккал/моль (30,5 кДж/моль) =

ЦПЭ
ЦПЭ – ферментный комплекс, окисляющий НАД*H и ФАД*H2 и использует энергию
ЦПЭ
ЦПЭ – ферментный комплекс, окисляющий НАД*H и ФАД*H2 и использует энергию

ЦПЭ
Комплекс III (Цитохром bc1 комплекс) переносит электроны с убихинола на два водорастворимых цитохрома
ЦПЭ
Комплекс III (Цитохром bc1 комплекс) переносит электроны с убихинола на два водорастворимых цитохрома

Окислительное фосфорилирование
Процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — протонной АТФ-синтазой
Окислительное фосфорилирование
Процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий — протонной АТФ-синтазой
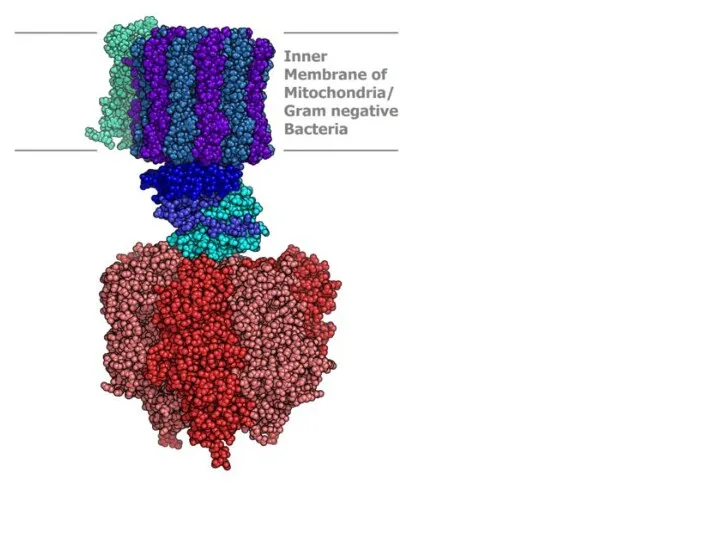

Полифенолазы и пероксидазы в энергетическом обмене
Полифенолазы и пероксидазы катализируют окисление фенольных
Полифенолазы и пероксидазы в энергетическом обмене
Полифенолазы и пероксидазы катализируют окисление фенольных

Пероксидаза — фермент, катализирующий окисление полифенолов и некоторых ароматических аминов при
Пероксидаза — фермент, катализирующий окисление полифенолов и некоторых ароматических аминов при

Полифенолоксидаза и пероксидаза являются конечными оксидазами и характеризуют интенсивность заключительной фазы
Полифенолоксидаза и пероксидаза являются конечными оксидазами и характеризуют интенсивность заключительной фазы

пероксидаза — чувствительный индикатор, регистрирующий изменения в газообмене, и может быть
пероксидаза — чувствительный индикатор, регистрирующий изменения в газообмене, и может быть
 Отряд Хищные
Отряд Хищные Животные Санкт-Петербурга и Ленинградской области
Животные Санкт-Петербурга и Ленинградской области Наземные беспозвоночные
Наземные беспозвоночные Факты о генетике
Факты о генетике Физиология микроорганизмов
Физиология микроорганизмов Происхождение и одомашнивание кошки
Происхождение и одомашнивание кошки Авторская методическая разработка по теме Слуховой анализатор.Строение уха.
Авторская методическая разработка по теме Слуховой анализатор.Строение уха. Биогенные элементы. Классификация. Топография биогенных элементов в организме человека
Биогенные элементы. Классификация. Топография биогенных элементов в организме человека Периферическая нервная система. Строение и функции спинномозговых нервов
Периферическая нервная система. Строение и функции спинномозговых нервов Род арония
Род арония Регуляция внутренних органов: роль отделов ЦНС, вегетативной нервной системы
Регуляция внутренних органов: роль отделов ЦНС, вегетативной нервной системы Экзотические животные Африки
Экзотические животные Африки Решение генетических задач
Решение генетических задач Бионика
Бионика Царства живой природы
Царства живой природы Теория эволюции Ч. Дарвина. Формы естественного отбора
Теория эволюции Ч. Дарвина. Формы естественного отбора Чарлз Дарвин об эволюции органического мира
Чарлз Дарвин об эволюции органического мира Основные элементы биотехнологических процессов
Основные элементы биотехнологических процессов Сохраним хвойные деревья в Советском лесничестве урочище Угланов лес
Сохраним хвойные деревья в Советском лесничестве урочище Угланов лес Основы генетики. 10 класс
Основы генетики. 10 класс Подкласс Розиды
Подкласс Розиды Окружающий мир 3 класс. Человек
Окружающий мир 3 класс. Человек Взаимодействие света с веществом. (Лекция 12)
Взаимодействие света с веществом. (Лекция 12) Бесхлорофильный фотосинтез
Бесхлорофильный фотосинтез Свойства живого вещества
Свойства живого вещества Биологические основы применения гербицидов
Биологические основы применения гербицидов Генетика – это наука о закономерностях наследования признаков у организмов
Генетика – это наука о закономерностях наследования признаков у организмов Паразитические высшие растения
Паразитические высшие растения