- Классификация, морфология и биология вирусов

Содержание
- 2. ОТКРЫТИЕ ВИРУСОВ 80-е годы XIX века: Луи Пастер – термин вирус (от лат. virus – яд).
- 3. Открытие вирусов 1892 – Дмитрий Иосифович Ивановский выделил возбудитель мозаичной болезни табака (фильтрующийся агент).
- 4. ПРИОРИТЕТ Дмитрий Иосифович Ивановский: 1892 – открыл «фильтрующийся вирус», но не опубликовал результатов исследований). Мартинус Виллем
- 5. ЭТАПЫ РАЗВИТИЯ ВИРУСОЛОГИИ До 1930 методы изучения вирусов основывались на фильтруемости и заражении различных чувствительных организмов
- 6. Достижения вирусологии Роль нуклеиновых кислот в наследственности (А. Д. Херши и М. Чейз, 1952; А. Гирер
- 7. Достижения вирусологии К началу XXI века описано 6 000 вирусов, изучена их структура, биология, химический состав
- 8. «…Сумеречная зона жизни, на полпути между живым и неживым, и есть зона вируса. Если бы вирус
- 9. Вирус – неклеточная форма жизни, обладающая геномом (РНК или ДНК), но лишенная собственного синтезирующего аппарата и
- 10. СВОЙСТВА ВИРУСОВ Размеры вириона: от 20–30 нм (пикорна-, парвовирусы) до 150–250 нм (герпес-, рабдовирусы) и даже
- 11. СТРОЕНИЕ ВИРИОНА Белковая оболочка – капсид (от греч. сapsa – футляр). Нуклеокапсид, капсомеры. Типы симметрии: ●
- 12. СТРОЕНИЕ ВИРИОНА безоболочечный вирус с икасаэдрическим типом симметрии; оболочечный вирус с икасаэдрическим типом симметрии; безоболочечный вирус
- 14. ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА ДНК вирусов: ∙ одноцепочечная линейная ДНК (парвовирусы: «+» и «-» нити находится в
- 15. ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА РНК вирусов – одноцепочечные молекулы (исключая реовирусы, обладающие двуцепочечной РНК), могут быть сегментированные
- 16. ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА Структурные и неструктурные белки. Капсидные белки: ∙ NP-белки (нуклеокапсидные); ∙ собственно капсидные (коровские)
- 17. ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА Вирусные ферменты: ● ферменты, участвующие в репликации и транскрипции; ● ферменты, обеспечивающие проникновение
- 18. ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА Фосфо- и гликолипиды клеточного происхождения (суперкапсид). Стабилизация вириона; суперкапсидных белков; защита; участвуют в
- 19. НЕКАНОНИЧЕСКИЕ ВИРУСЫ Вироиды – небольшие молекулы кольцевой, суперспирализованной РНК, не содержащей белка и вызывающие заболевание растений.
- 20. Прионовые болезни Карлтон Гайдучек – американский педиатр. 1953г.: Куру – «хохочущая смерть». Экзотическое заболевание среди аборигенов
- 21. Прионовые болезни Стенли Прусинен – теория инфекционных белков «прионов». Нобелевская премия 1997 г. Болезнь Крейтцфельдта-Якоба, синдром
- 22. ПРОИСХОЖДЕНИЕ ВИРУСОВ I гипотеза: вирусы – это потомки бактерий или других одноклеточных микроорганизмов, появившиеся в результате
- 23. СИСТЕМАТИКА ВИРУСОВ 1966 г. – создан Международный комитет по номенклатуре вирусов. 1971 г. опубликован первый его
- 24. Критерии таксономической классификации вирусов гомология нуклеиновых кислот; морфология, размеры, форма; наличие или отсутствие суперкапсида; тип симметрии
- 25. КЛАССИФИКАЦИЯ ВИРУСОВ Царство Vira делится на два подцарства: дезоксивирусы (ДНК-вирусы) и рибовирусы (РНК-вирусы). Подцарства подразделяются на
- 26. Классификация вирусов, патогенных для человека
- 27. Классификация вирусов, патогенных для человека
- 28. Классификация вирусов, патогенных для человека
- 29. Классификация вирусов, патогенных для человека
- 30. Классификация вирусов, патогенных для человека
- 31. Таксономия вирусов и новые вирусы (Международный таксономический комитет) 1999 год 3 порядка, 56 семейств, 9 субсемейств,
- 32. Физиология вирусов Вирус является облигатным внутриклеточным паразитом и для размножения ему требуется живая клетка. Размножение вируса
- 33. Типы взаимодействия вируса с клеткой продуктивный, или цитоцидный тип – в зараженных клетках образуется новое поколение
- 34. Продуктивный тип взаимодействия вируса с клеткой осуществляется в результате размножения, т.е. репродукции вируса (от англ. reproduce
- 35. Стадии репродукции вируса адсорбция вирионов на клетке; проникновение вирусов в клетку; депротеинизация или «раздевание» вирусов и
- 36. Адсорбция вируса на клетке Взаимодействие поверхностных белков вируса со специфическими рецепторами чувствительных клеток. Тропизм вируса (греч.
- 37. Проникновение вируса в клетку Виропексис (рецепторный эндоцитоз). Слияние оболочки вируса с клеточной мембраной (при наличии белка
- 38. Депротеинизация При участии клеточных ферментов происходит депротеинизация вириона (удаление поверхностных структур, высвобождение внутреннего компонента). После депротеинезации
- 39. Биосинтез вирусных компонентов Биосинтез вирусных компонентов – дизъюнктивный (от лат. disjunctus – разобщенный). Транскрипция → трансляция
- 40. Стратегия генома и репродукция вирусов 1 компонент: ДНК→транскрипция→мРНК→ трансляция→белок 2 компонент: ДНК→репликация→ДНК
- 41. Репликация вирусных геномов Двунитевая ДНК – полуконсервативная репликация. Однонитевая ДНК: вначале образуется репликативная форма («-» нить
- 42. Однонитевая РНК: вРНК→кРНК→вРНК, катализируется репликазой. Репликация вирусных геномов +РНК –РНК
- 43. Однонитевая РНК ретровирусов: РНК→ДНК→РНК, катализируется обратной транскриптазой. Репликация вирусных геномов
- 44. Двунитевая ДНК вируса гепатита В: ДНК→РНК→ДНК→РНК, катализируется клеточной РНК-полимеразой и обратной транскриптазой. Репликация вирусных геномов
- 45. Особенности репродукции вирусов +РНК вирусы: трансляция → репликация → сборка вириона. Остальные вирусы: транскрипция → трансляция
- 46. Формирование вирионов Самосборка: составные части вириона транспортируются в места сборки вируса в ядре или цитоплазме. Сборка
- 47. Выход вирионов из клетки Взрывной: из погибающей клетки одновременно выходит большое количество вирионов. Простые вирусы.
- 48. Выход вирионов из клетки Почкование: нуклеокапсид транспортируется к клеточным мембранам, в которые уже встроены вирусные белки
- 50. Интегративный тип взаимодействия вируса с клеткой Вирогения – интеграция (встраивание) нуклеиновой кислоты вируса в геном клетки,
- 51. Интегративный тип взаимодействия вируса с клеткой Физические или химические факторы→провирус→ автономное состояние→продуктивный тип взаимодействия с клеткой→латентные
- 52. Основные положения концепции происхождения опухолей Л.А.Зильбера «...роль вируса в развитии опухолевого процесса сводится к тому, что
- 53. ОНКОГЕННЫЕ ВИРУСЫ ЧЕЛОВЕКА
- 54. Вакцинация против папилломавируса В июне 2006 года в США была одобрена вакцина Гардасил. Помимо онкогенных типов
- 55. Методы культивирования вирусов в организме лабораторных животных; в развивающихся куриных эмбрионах; в культурах клеток.
- 56. Культивирование вирусов в организме лабораторных животных Выбор экспериментальных животных определяется целью работы и видовой чувствительностью к
- 57. Культивирование вирусов в организме лабораторных животных Если при первичном заражении животные не заболевают → последовательные «пассажи».
- 58. Реакция вирусной гемагглютинации (РГА)
- 59. Культивирование вирусов в куриных эмбрионах Используют эмбрионы в возрасте от 8 до 14 дней в зависимости
- 60. Строение куриного эмбриона Хорионаллантоисная оболочка Амниотическая полость Белок Желточный мешок Скорлупа Воздушный мешок Аллантоисная полость
- 61. Способы заражения куриного эмбриона
- 62. Способы заражения куриного эмбриона Заражение на хорионаллантоисную оболочку (вирусы вакцины, натуральной оспы, простого герпеса). На зараженной
- 63. Способы заражения куриного эмбриона Заражение в аллантоисную полость. Вирус размножается в эндодермальных клетках, переходит затем в
- 64. Культивирование вирусов в культуре ткани
- 65. Классификация клеточных культур В зависимости от техники приготовления: однослойные – клетки, способные прикрепляться и размножаться на
- 66. Классификация клеточных культур По числу жизнеспособных генераций: первичные, способные размножаться только на первых генерациях, т.е. в
- 67. Культуральная посуда
- 68. Приготовление клеточных культур Чаще используют ткани эмбрионов или почечную ткань взрослых животных. Трипсинизирование: разрушение межклеточных связей
- 69. Индикация вирусов в культуре клеток Цитопатическое действие (ЦПД) – видимые под микроскопом морфологические изменения клеток в
- 70. Образование симпластов ЦПД вируса кори на клетки Vero: образование гигантских многоядерных клеток - симпластов, с последующей
- 71. Индикация вирусов в культуре клеток Вирусные включения – скопление вирусных частиц или отдельных компонентов вирусов в
- 72. Вирусные включения
- 73. Индикация вирусов в культуре клеток Бляшки, или негативные колонии – ограниченные участки, состоящие из дегенеративных клеток,
- 74. Индикация вирусов в культуре клеток Гемадсорбция – способность культур клеток, инфицированных вирусами, адсорбировать на своей поверхности
- 75. Индикация вирусов в культуре клеток «Цветная» проба. Если вирусы не размножаются в культуре клеток, то живые
- 76. Бактериофаги
- 77. История открытия бактериофагов 1898 – Н.Ф. Гамалея обнаружил спонтанное разрушение палочек сибирской язвы. 1915 – Фредерик
- 78. Выделение бактериофагов Выделить бактериофаг можно из естественного места обитания бактерии-хозяина. Фаги можно увидеть и на плотной
- 79. Классификация бактериофагов Нитевидные фаги; фаги с длинным отростком и сокращающимся чехлом; фаги с длинным отростком без
- 80. Строение бактериофага
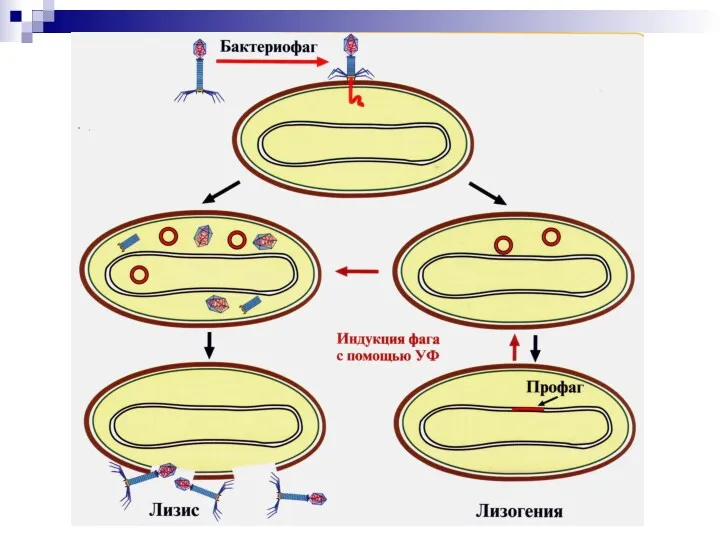
- 82. Взаимодействие фага с бактериальной клеткой Вирулентные бактериофаги Продуктивный тип. Литический цикл. Умеренные бактериофаги Интегративный тип. Лизогения.
- 84. Применение бактериофагов Лечебные бактериофаги: моновалентные, поливалентные (пиобактериофаг, интестифаг) Диагностические бактериофаги: видовые, типовые.
- 85. Лечебные бактериофаги Препарат бактериофага: фильтрат бульонной культуры, зараженной соответствующим фагом. Препараты назначаются с целью лечения и
- 87. Скачать презентацию

ОТКРЫТИЕ ВИРУСОВ
80-е годы XIX века: Луи Пастер –
термин вирус
ОТКРЫТИЕ ВИРУСОВ
80-е годы XIX века: Луи Пастер –
термин вирус

Открытие вирусов
1892 – Дмитрий Иосифович Ивановский выделил возбудитель мозаичной болезни табака
Открытие вирусов
1892 – Дмитрий Иосифович Ивановский выделил возбудитель мозаичной болезни табака

ПРИОРИТЕТ
Дмитрий Иосифович Ивановский: 1892 – открыл «фильтрующийся вирус», но не опубликовал
ПРИОРИТЕТ
Дмитрий Иосифович Ивановский: 1892 – открыл «фильтрующийся вирус», но не опубликовал

ЭТАПЫ РАЗВИТИЯ ВИРУСОЛОГИИ
До 1930 методы изучения вирусов основывались на фильтруемости и
ЭТАПЫ РАЗВИТИЯ ВИРУСОЛОГИИ
До 1930 методы изучения вирусов основывались на фильтруемости и

Достижения вирусологии
Роль нуклеиновых кислот в наследственности (А. Д. Херши и М.
Достижения вирусологии
Роль нуклеиновых кислот в наследственности (А. Д. Херши и М.

Достижения вирусологии
К началу XXI века описано 6 000 вирусов, изучена их
Достижения вирусологии
К началу XXI века описано 6 000 вирусов, изучена их

«…Сумеречная зона жизни, на полпути между живым и неживым, и
«…Сумеречная зона жизни, на полпути между живым и неживым, и
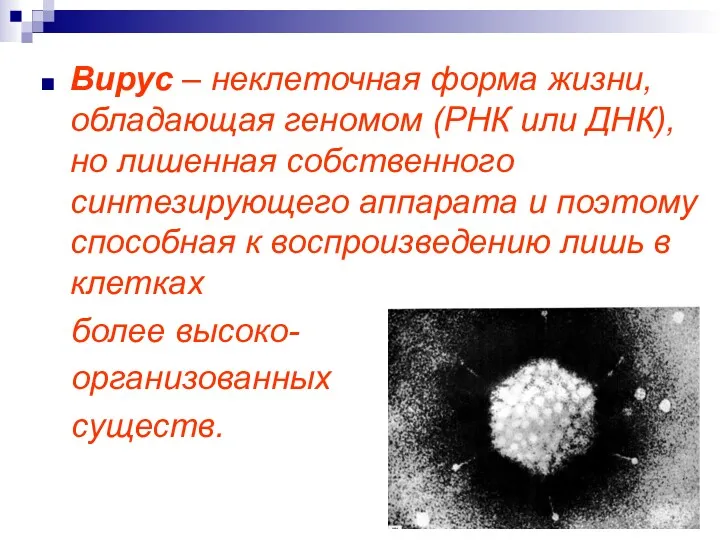
Вирус – неклеточная форма жизни, обладающая геномом (РНК или ДНК), но
Вирус – неклеточная форма жизни, обладающая геномом (РНК или ДНК), но
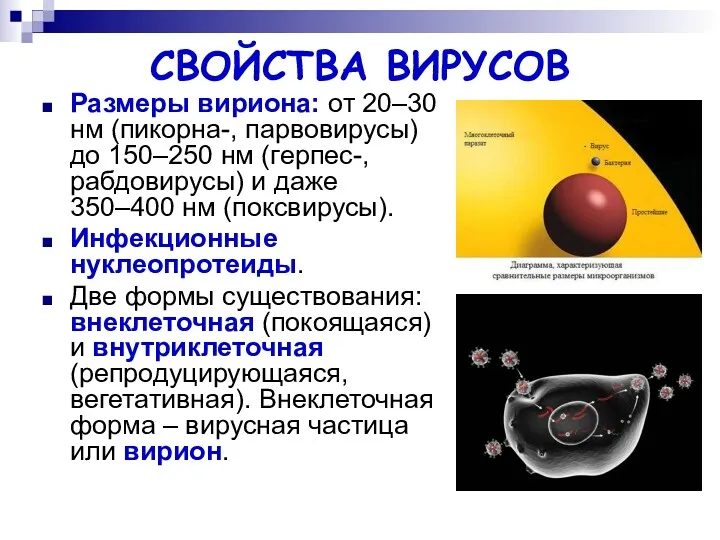
СВОЙСТВА ВИРУСОВ
Размеры вириона: от 20–30 нм (пикорна-, парвовирусы) до 150–250 нм
СВОЙСТВА ВИРУСОВ
Размеры вириона: от 20–30 нм (пикорна-, парвовирусы) до 150–250 нм
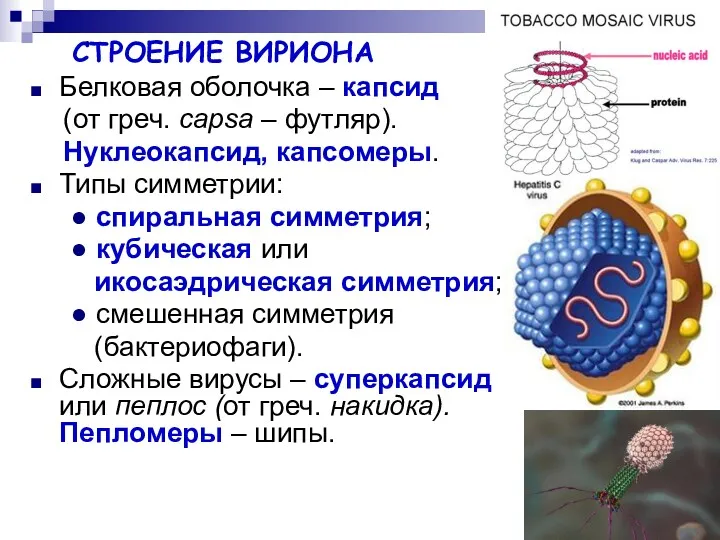
СТРОЕНИЕ ВИРИОНА
Белковая оболочка – капсид
(от греч. сapsa – футляр).
СТРОЕНИЕ ВИРИОНА
Белковая оболочка – капсид
(от греч. сapsa – футляр).
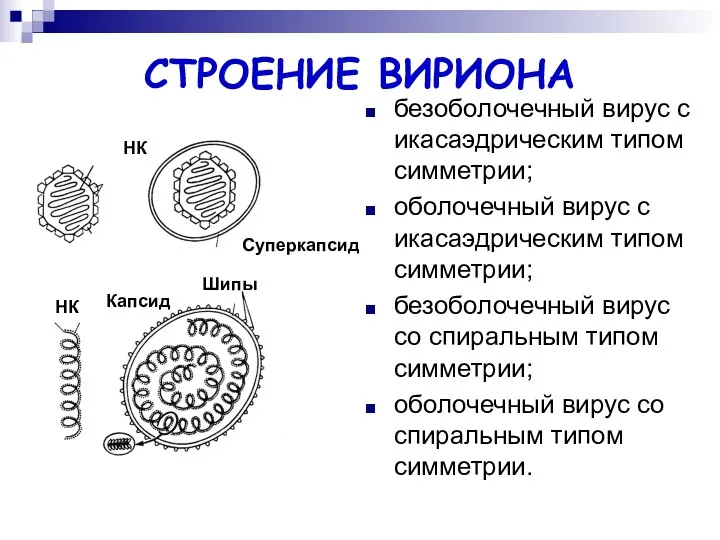
СТРОЕНИЕ ВИРИОНА
безоболочечный вирус с икасаэдрическим типом симметрии;
оболочечный вирус с икасаэдрическим типом
СТРОЕНИЕ ВИРИОНА
безоболочечный вирус с икасаэдрическим типом симметрии;
оболочечный вирус с икасаэдрическим типом

ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
ДНК вирусов:
∙ одноцепочечная линейная ДНК (парвовирусы: «+» и
ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
ДНК вирусов:
∙ одноцепочечная линейная ДНК (парвовирусы: «+» и

ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
РНК вирусов – одноцепочечные молекулы (исключая реовирусы, обладающие двуцепочечной
ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
РНК вирусов – одноцепочечные молекулы (исключая реовирусы, обладающие двуцепочечной

ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
Структурные и неструктурные белки.
Капсидные белки:
∙ NP-белки (нуклеокапсидные);
∙
ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
Структурные и неструктурные белки.
Капсидные белки:
∙ NP-белки (нуклеокапсидные);
∙

ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
Вирусные ферменты:
● ферменты, участвующие в репликации и
ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
Вирусные ферменты:
● ферменты, участвующие в репликации и

ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
Фосфо- и гликолипиды клеточного происхождения (суперкапсид). Стабилизация вириона; суперкапсидных
ХИМИЧЕСКИЙ СОСТАВ ВИРИОНА
Фосфо- и гликолипиды клеточного происхождения (суперкапсид). Стабилизация вириона; суперкапсидных

НЕКАНОНИЧЕСКИЕ ВИРУСЫ
Вироиды – небольшие молекулы кольцевой, суперспирализованной РНК, не содержащей белка
НЕКАНОНИЧЕСКИЕ ВИРУСЫ
Вироиды – небольшие молекулы кольцевой, суперспирализованной РНК, не содержащей белка

Прионовые болезни
Карлтон Гайдучек – американский педиатр.
1953г.: Куру – «хохочущая смерть».
Экзотическое заболевание
Прионовые болезни
Карлтон Гайдучек – американский педиатр.
1953г.: Куру – «хохочущая смерть».
Экзотическое заболевание
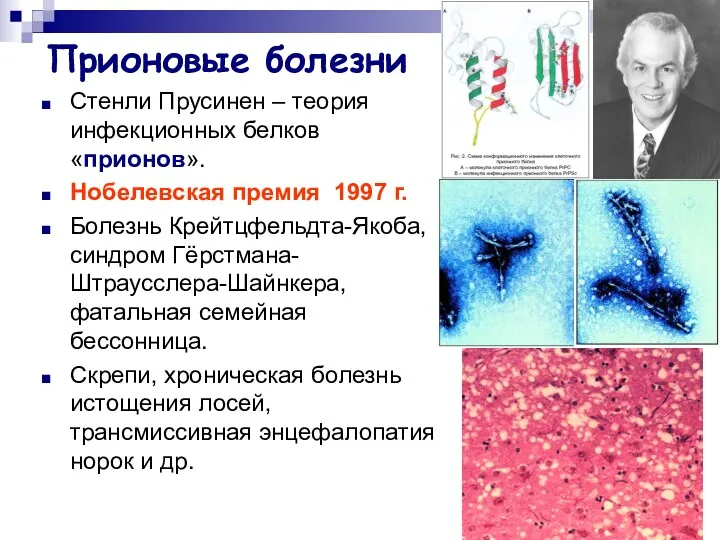
Прионовые болезни
Стенли Прусинен – теория инфекционных белков «прионов».
Нобелевская премия 1997 г.
Болезнь
Прионовые болезни
Стенли Прусинен – теория инфекционных белков «прионов».
Нобелевская премия 1997 г.
Болезнь

ПРОИСХОЖДЕНИЕ ВИРУСОВ
I гипотеза: вирусы – это потомки бактерий или других одноклеточных
ПРОИСХОЖДЕНИЕ ВИРУСОВ
I гипотеза: вирусы – это потомки бактерий или других одноклеточных

СИСТЕМАТИКА ВИРУСОВ
1966 г. – создан Международный комитет по номенклатуре вирусов.
1971 г.
СИСТЕМАТИКА ВИРУСОВ
1966 г. – создан Международный комитет по номенклатуре вирусов.
1971 г.

Критерии таксономической классификации вирусов
гомология нуклеиновых кислот;
морфология, размеры, форма;
наличие или отсутствие суперкапсида;
тип
Критерии таксономической классификации вирусов
гомология нуклеиновых кислот;
морфология, размеры, форма;
наличие или отсутствие суперкапсида;
тип

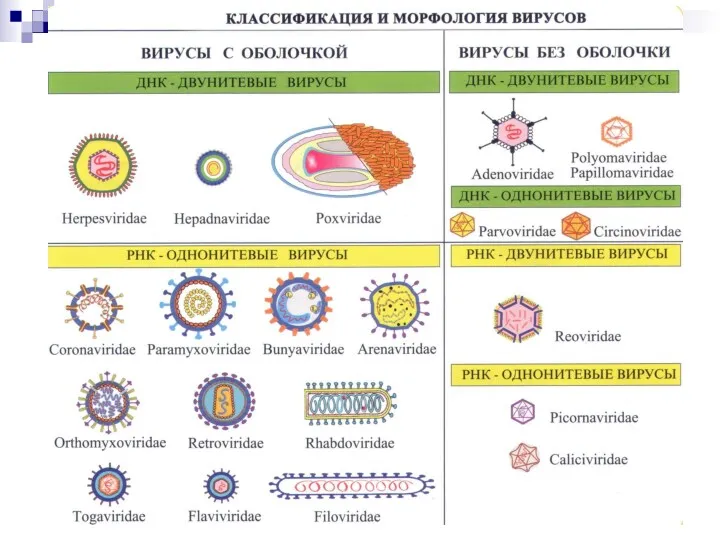
КЛАССИФИКАЦИЯ ВИРУСОВ
Царство Vira делится на два подцарства: дезоксивирусы (ДНК-вирусы) и рибовирусы
КЛАССИФИКАЦИЯ ВИРУСОВ
Царство Vira делится на два подцарства: дезоксивирусы (ДНК-вирусы) и рибовирусы
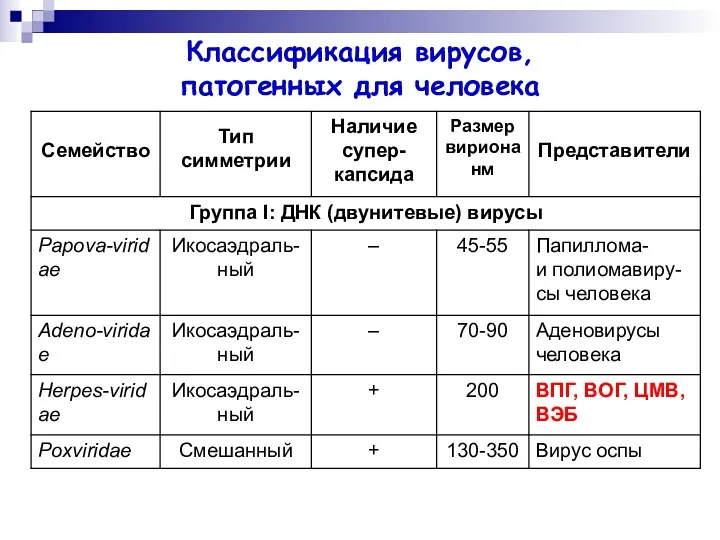
Классификация вирусов,
патогенных для человека
Классификация вирусов,
патогенных для человека
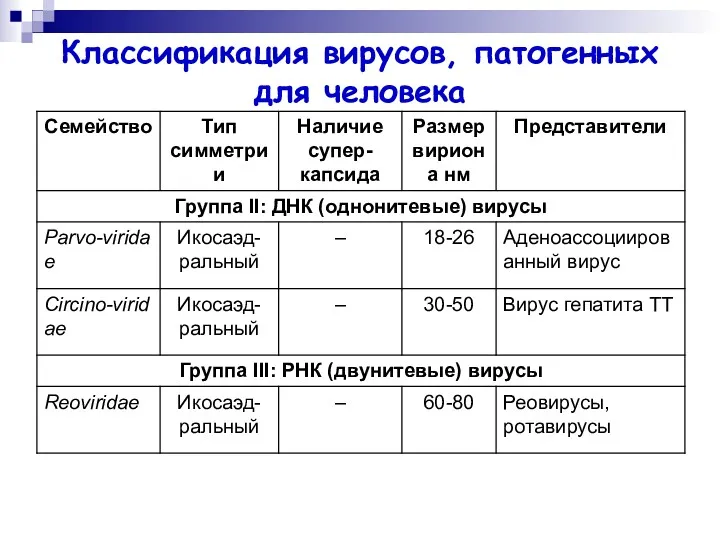
Классификация вирусов, патогенных для человека
Классификация вирусов, патогенных для человека
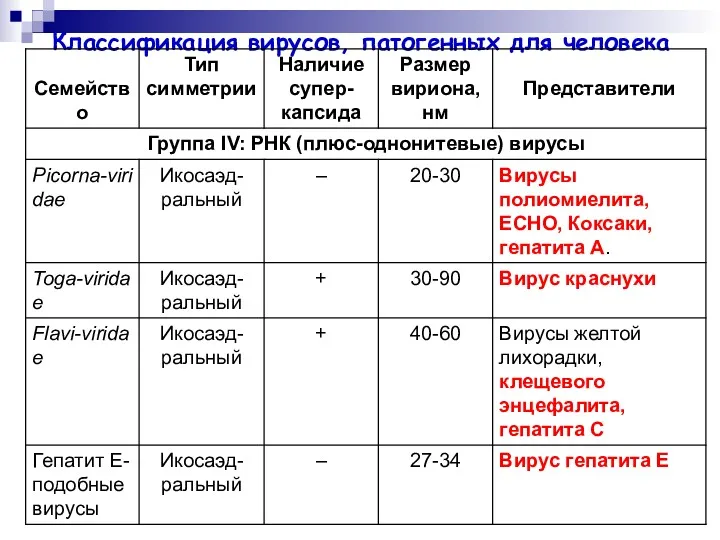
Классификация вирусов, патогенных для человека
Классификация вирусов, патогенных для человека
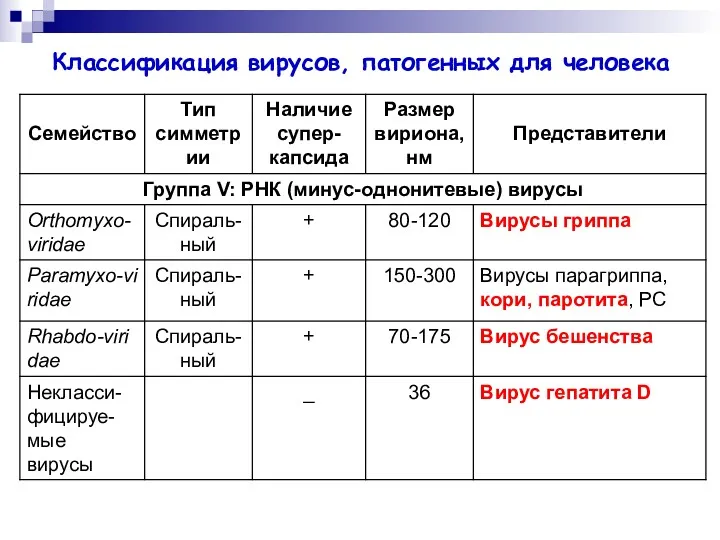
Классификация вирусов, патогенных для человека
Классификация вирусов, патогенных для человека
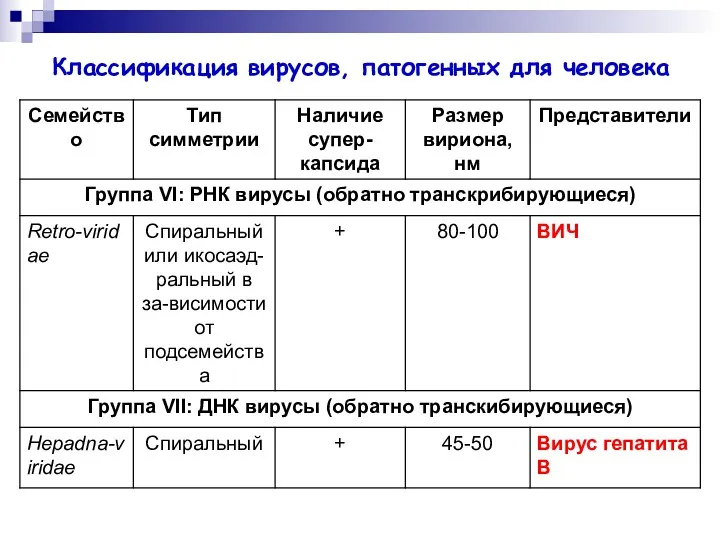
Классификация вирусов, патогенных для человека
Классификация вирусов, патогенных для человека

Таксономия вирусов и новые вирусы
(Международный таксономический комитет)
1999 год
3 порядка,
56 семейств,
Таксономия вирусов и новые вирусы
(Международный таксономический комитет)
1999 год
3 порядка,
56 семейств,

Физиология вирусов
Вирус является облигатным внутриклеточным паразитом и для размножения ему
Физиология вирусов
Вирус является облигатным внутриклеточным паразитом и для размножения ему

Типы взаимодействия вируса с клеткой
продуктивный, или цитоцидный тип – в зараженных
Типы взаимодействия вируса с клеткой
продуктивный, или цитоцидный тип – в зараженных

Продуктивный тип взаимодействия вируса с клеткой
осуществляется в результате размножения, т.е. репродукции
Продуктивный тип взаимодействия вируса с клеткой
осуществляется в результате размножения, т.е. репродукции
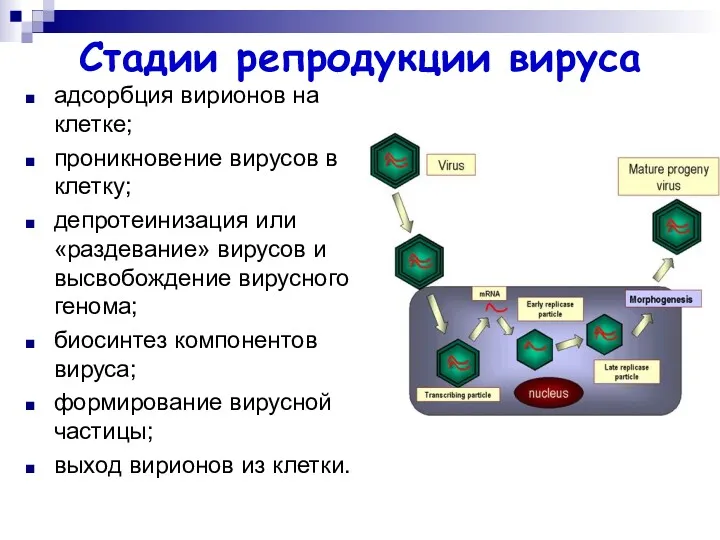
Стадии репродукции вируса
адсорбция вирионов на клетке;
проникновение вирусов в клетку;
депротеинизация или «раздевание»
Стадии репродукции вируса
адсорбция вирионов на клетке;
проникновение вирусов в клетку;
депротеинизация или «раздевание»

Адсорбция вируса на клетке
Взаимодействие поверхностных белков вируса со специфическими рецепторами чувствительных
Адсорбция вируса на клетке
Взаимодействие поверхностных белков вируса со специфическими рецепторами чувствительных
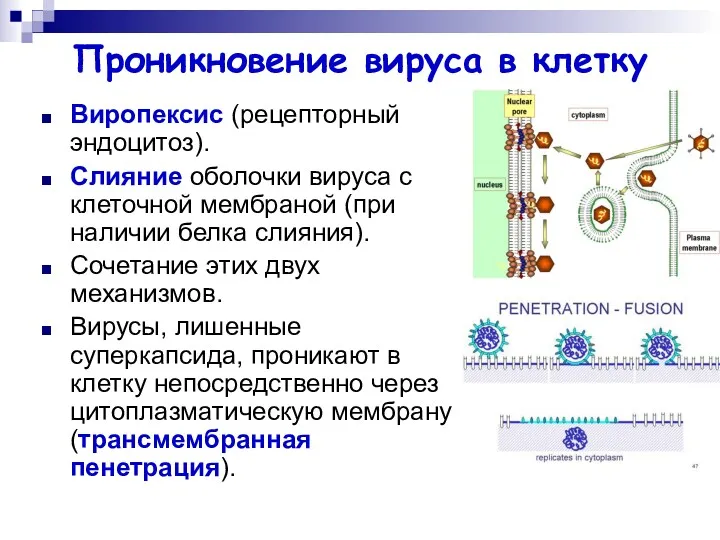
Проникновение вируса в клетку
Виропексис (рецепторный эндоцитоз).
Слияние оболочки вируса с клеточной мембраной
Проникновение вируса в клетку
Виропексис (рецепторный эндоцитоз).
Слияние оболочки вируса с клеточной мембраной
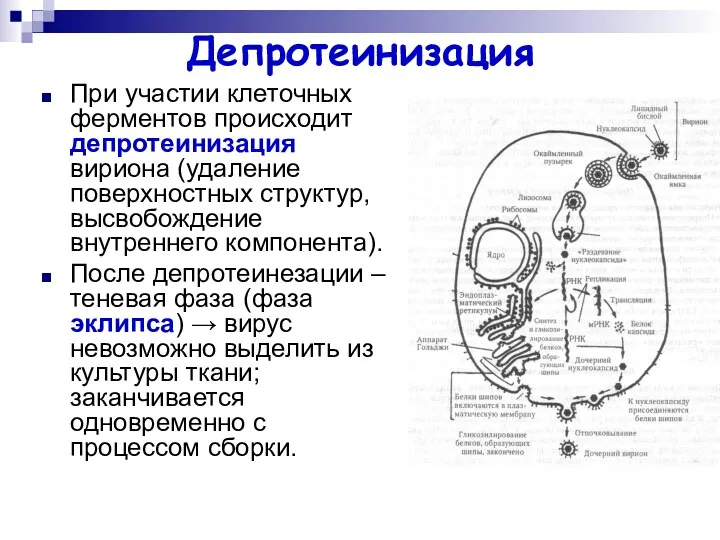
Депротеинизация
При участии клеточных ферментов происходит депротеинизация вириона (удаление поверхностных структур, высвобождение
Депротеинизация
При участии клеточных ферментов происходит депротеинизация вириона (удаление поверхностных структур, высвобождение

Биосинтез вирусных компонентов
Биосинтез вирусных компонентов – дизъюнктивный (от лат. disjunctus –
Биосинтез вирусных компонентов
Биосинтез вирусных компонентов – дизъюнктивный (от лат. disjunctus –

Стратегия генома и репродукция вирусов
1 компонент:
ДНК→транскрипция→мРНК→
трансляция→белок
2 компонент:
ДНК→репликация→ДНК
Стратегия генома и репродукция вирусов
1 компонент:
ДНК→транскрипция→мРНК→
трансляция→белок
2 компонент:
ДНК→репликация→ДНК
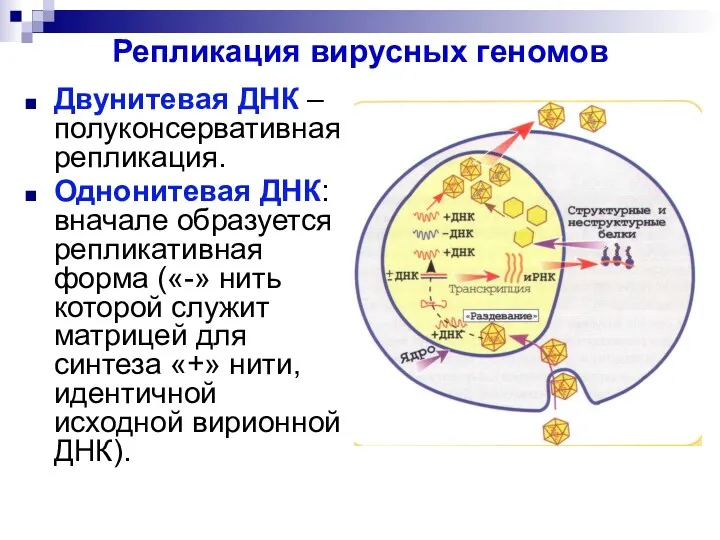
Репликация вирусных геномов
Двунитевая ДНК – полуконсервативная репликация.
Однонитевая ДНК: вначале образуется репликативная
Репликация вирусных геномов
Двунитевая ДНК – полуконсервативная репликация.
Однонитевая ДНК: вначале образуется репликативная
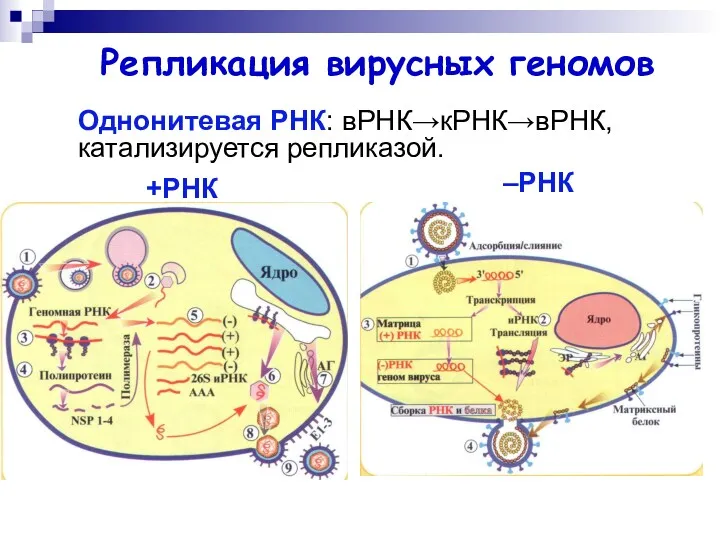
Однонитевая РНК: вРНК→кРНК→вРНК, катализируется репликазой.
Репликация вирусных геномов
+РНК
–РНК
Однонитевая РНК: вРНК→кРНК→вРНК, катализируется репликазой.
Репликация вирусных геномов
+РНК
–РНК
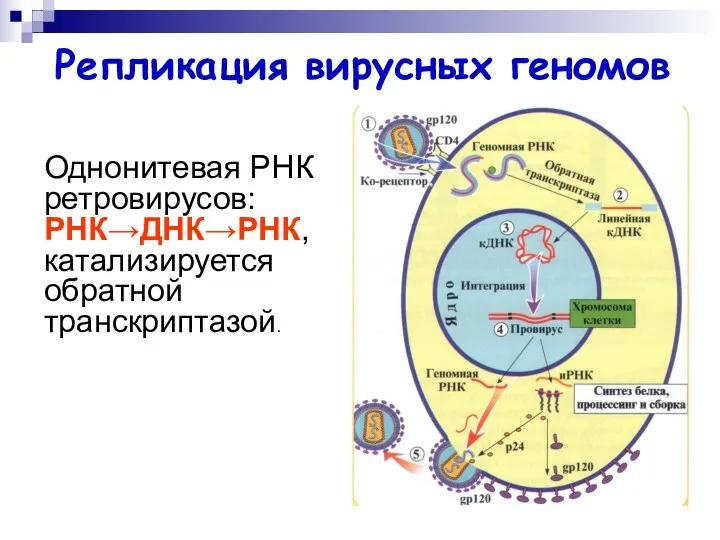
Однонитевая РНК ретровирусов: РНК→ДНК→РНК, катализируется обратной транскриптазой.
Репликация вирусных геномов
Однонитевая РНК ретровирусов: РНК→ДНК→РНК, катализируется обратной транскриптазой.
Репликация вирусных геномов
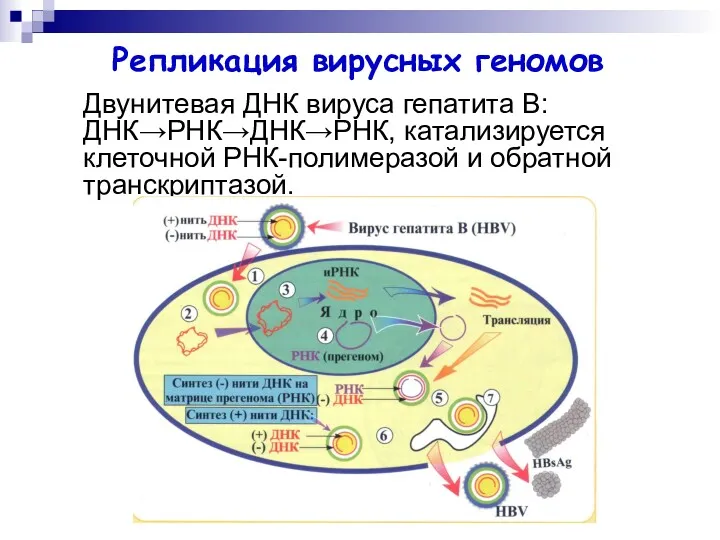
Двунитевая ДНК вируса гепатита В: ДНК→РНК→ДНК→РНК, катализируется клеточной РНК-полимеразой и обратной
Двунитевая ДНК вируса гепатита В: ДНК→РНК→ДНК→РНК, катализируется клеточной РНК-полимеразой и обратной

Особенности репродукции вирусов
+РНК вирусы: трансляция → репликация → сборка вириона.
Остальные
Особенности репродукции вирусов
+РНК вирусы: трансляция → репликация → сборка вириона.
Остальные

Формирование вирионов
Самосборка: составные части вириона транспортируются в места сборки вируса в
Формирование вирионов
Самосборка: составные части вириона транспортируются в места сборки вируса в
Выход вирионов из клетки
Взрывной: из погибающей клетки одновременно выходит большое количество
Выход вирионов из клетки
Взрывной: из погибающей клетки одновременно выходит большое количество

Выход вирионов из клетки
Почкование: нуклеокапсид транспортируется к клеточным мембранам, в которые
Выход вирионов из клетки
Почкование: нуклеокапсид транспортируется к клеточным мембранам, в которые

Интегративный тип взаимодействия вируса с клеткой
Вирогения – интеграция (встраивание) нуклеиновой кислоты
Интегративный тип взаимодействия вируса с клеткой
Вирогения – интеграция (встраивание) нуклеиновой кислоты

Интегративный тип взаимодействия вируса с клеткой
Физические или химические факторы→провирус→ автономное состояние→продуктивный
Интегративный тип взаимодействия вируса с клеткой
Физические или химические факторы→провирус→ автономное состояние→продуктивный

Основные положения концепции происхождения опухолей Л.А.Зильбера
«...роль вируса в развитии опухолевого
Основные положения концепции происхождения опухолей Л.А.Зильбера
«...роль вируса в развитии опухолевого
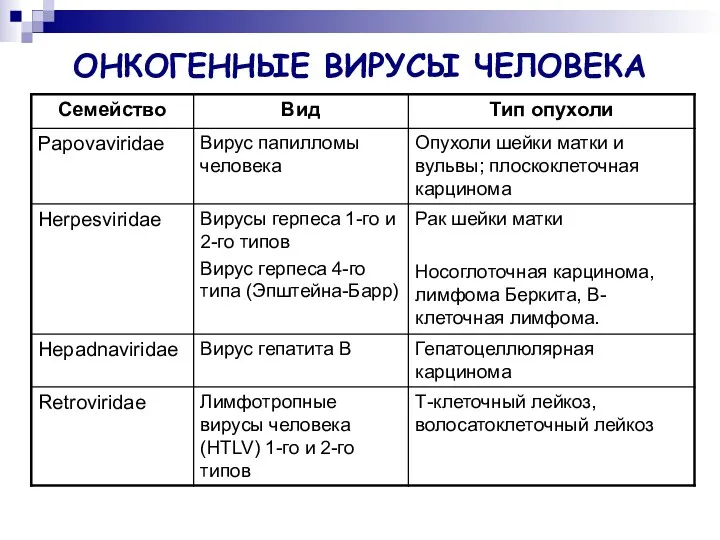
ОНКОГЕННЫЕ ВИРУСЫ ЧЕЛОВЕКА
ОНКОГЕННЫЕ ВИРУСЫ ЧЕЛОВЕКА

Вакцинация против папилломавируса
В июне 2006 года в США была одобрена
Вакцинация против папилломавируса
В июне 2006 года в США была одобрена
Методы культивирования вирусов
в организме лабораторных животных;
в развивающихся куриных эмбрионах;
в культурах клеток.
Методы культивирования вирусов
в организме лабораторных животных;
в развивающихся куриных эмбрионах;
в культурах клеток.

Культивирование вирусов в организме лабораторных животных
Выбор экспериментальных животных
определяется целью
Культивирование вирусов в организме лабораторных животных
Выбор экспериментальных животных
определяется целью

Культивирование вирусов в организме лабораторных животных
Если при первичном заражении животные не
Культивирование вирусов в организме лабораторных животных
Если при первичном заражении животные не
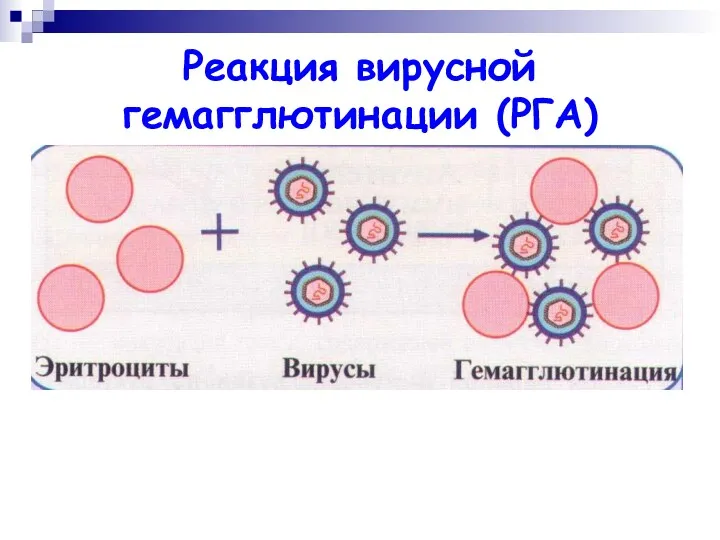
Реакция вирусной гемагглютинации (РГА)
Реакция вирусной гемагглютинации (РГА)

Культивирование вирусов в куриных эмбрионах
Используют эмбрионы в возрасте от 8 до
Культивирование вирусов в куриных эмбрионах
Используют эмбрионы в возрасте от 8 до
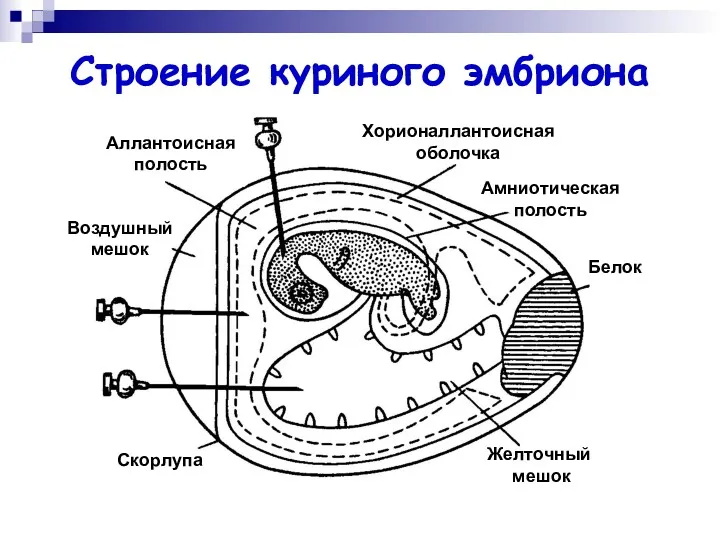
Строение куриного эмбриона
Хорионаллантоисная
оболочка
Амниотическая
полость
Белок
Желточный
мешок
Скорлупа
Воздушный
мешок
Аллантоисная
полость
Строение куриного эмбриона
Хорионаллантоисная
оболочка
Амниотическая
полость
Белок
Желточный
мешок
Скорлупа
Воздушный
мешок
Аллантоисная
полость

Способы заражения куриного эмбриона
Способы заражения куриного эмбриона
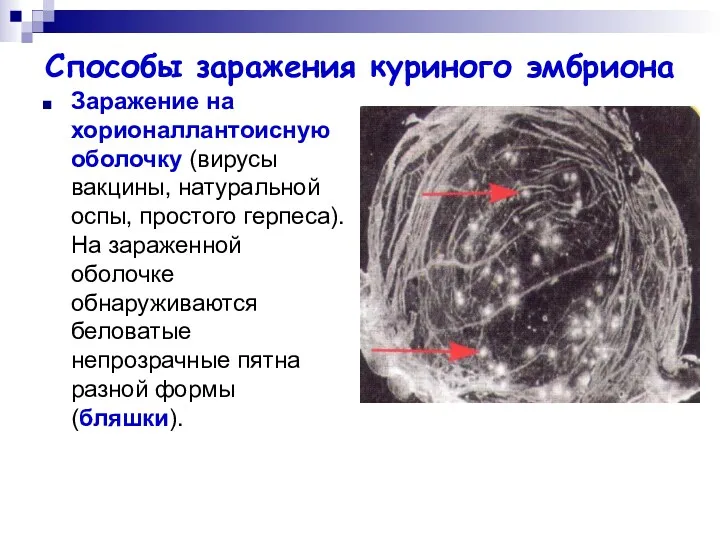
Способы заражения куриного эмбриона
Заражение на хорионаллантоисную оболочку (вирусы вакцины, натуральной оспы,
Способы заражения куриного эмбриона
Заражение на хорионаллантоисную оболочку (вирусы вакцины, натуральной оспы,

Способы заражения куриного эмбриона
Заражение в аллантоисную полость. Вирус размножается в эндодермальных
Способы заражения куриного эмбриона
Заражение в аллантоисную полость. Вирус размножается в эндодермальных

Культивирование вирусов в культуре ткани
Культивирование вирусов в культуре ткани

Классификация клеточных культур
В зависимости от техники приготовления:
однослойные – клетки, способные
Классификация клеточных культур
В зависимости от техники приготовления:
однослойные – клетки, способные

Классификация клеточных культур
По числу жизнеспособных генераций:
первичные, способные размножаться только на первых
Классификация клеточных культур
По числу жизнеспособных генераций:
первичные, способные размножаться только на первых

Культуральная посуда
Культуральная посуда
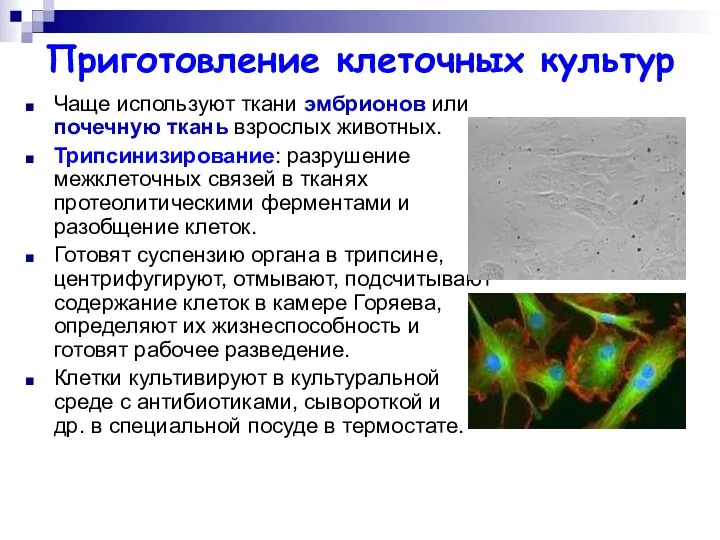
Приготовление клеточных культур
Чаще используют ткани эмбрионов или почечную ткань взрослых животных.
Трипсинизирование:
Приготовление клеточных культур
Чаще используют ткани эмбрионов или почечную ткань взрослых животных.
Трипсинизирование:
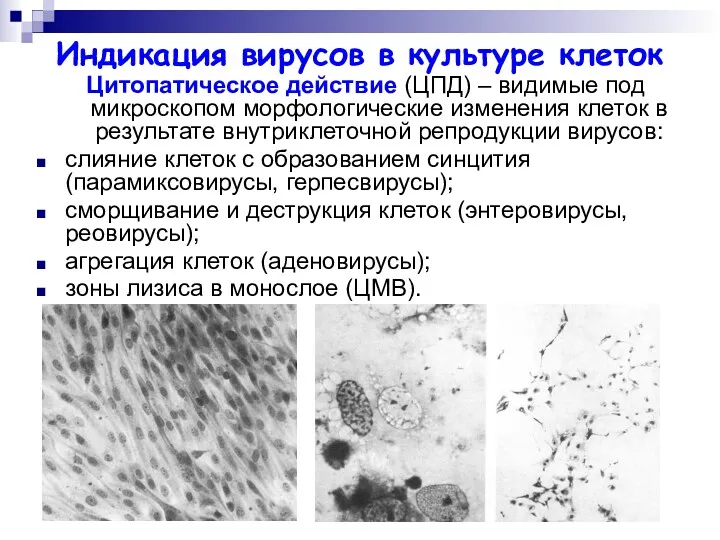
Индикация вирусов в культуре клеток
Цитопатическое действие (ЦПД) – видимые под микроскопом
Индикация вирусов в культуре клеток
Цитопатическое действие (ЦПД) – видимые под микроскопом
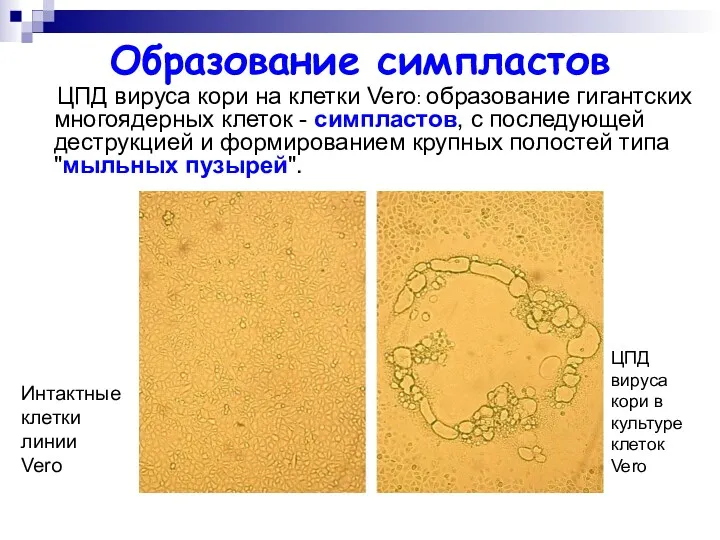
Образование симпластов
ЦПД вируса кори на клетки Vero: образование гигантских многоядерных
Образование симпластов
ЦПД вируса кори на клетки Vero: образование гигантских многоядерных
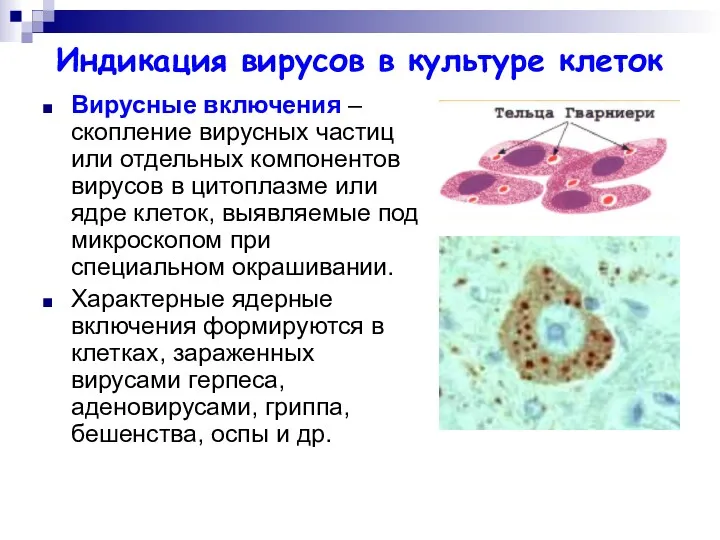
Индикация вирусов в культуре клеток
Вирусные включения – скопление вирусных частиц или
Индикация вирусов в культуре клеток
Вирусные включения – скопление вирусных частиц или
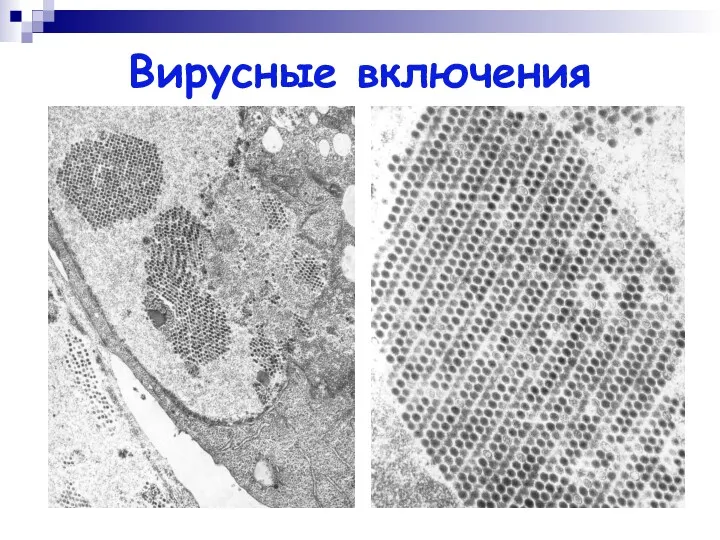
Вирусные включения
Вирусные включения

Индикация вирусов в культуре клеток
Бляшки, или негативные колонии – ограниченные участки,
Индикация вирусов в культуре клеток
Бляшки, или негативные колонии – ограниченные участки,
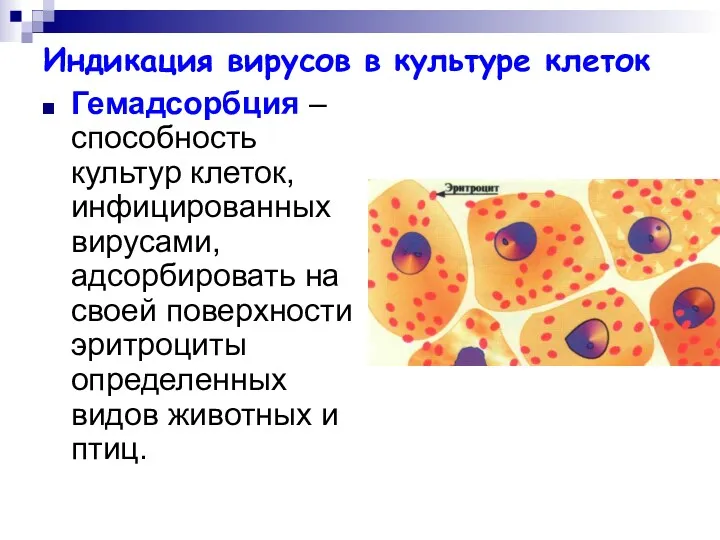
Индикация вирусов в культуре клеток
Гемадсорбция – способность культур клеток, инфицированных вирусами,
Индикация вирусов в культуре клеток
Гемадсорбция – способность культур клеток, инфицированных вирусами,

Индикация вирусов в культуре клеток
«Цветная» проба. Если вирусы не размножаются в
Индикация вирусов в культуре клеток
«Цветная» проба. Если вирусы не размножаются в

Бактериофаги
Бактериофаги

История открытия бактериофагов
1898 – Н.Ф. Гамалея обнаружил спонтанное разрушение палочек сибирской
История открытия бактериофагов
1898 – Н.Ф. Гамалея обнаружил спонтанное разрушение палочек сибирской

Выделение бактериофагов
Выделить бактериофаг можно из естественного места обитания бактерии-хозяина.
Фаги можно
Выделение бактериофагов
Выделить бактериофаг можно из естественного места обитания бактерии-хозяина.
Фаги можно
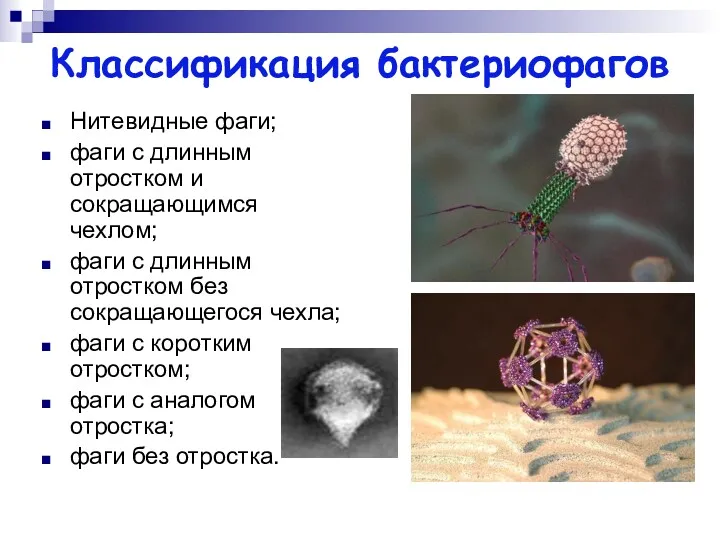
Классификация бактериофагов
Нитевидные фаги;
фаги с длинным отростком и сокращающимся чехлом;
фаги с длинным
Классификация бактериофагов
Нитевидные фаги;
фаги с длинным отростком и сокращающимся чехлом;
фаги с длинным
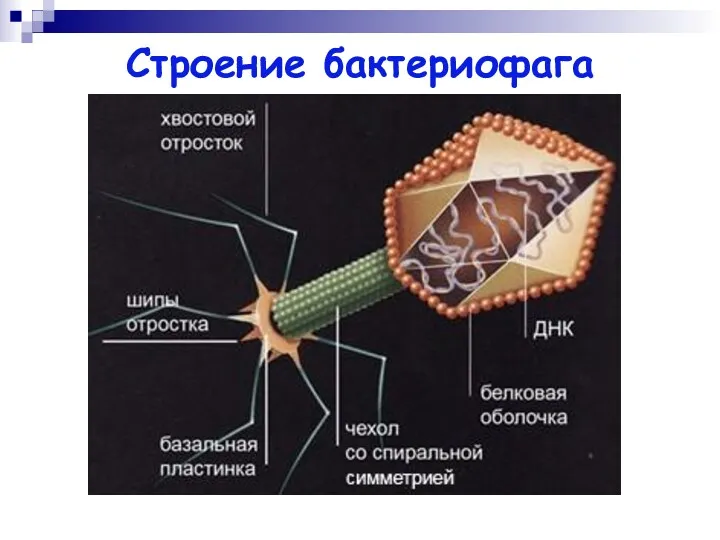
Строение бактериофага
Строение бактериофага
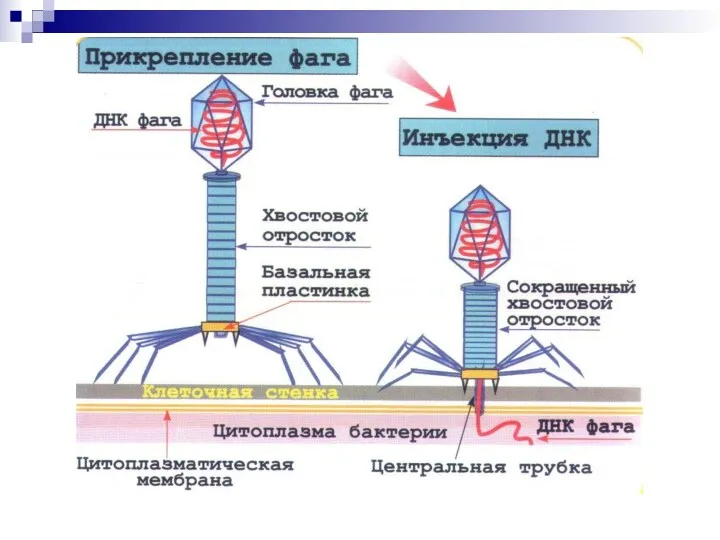

Взаимодействие фага с бактериальной клеткой
Вирулентные бактериофаги
Продуктивный тип. Литический цикл.
Умеренные
Взаимодействие фага с бактериальной клеткой
Вирулентные бактериофаги
Продуктивный тип. Литический цикл.
Умеренные

Применение бактериофагов
Лечебные бактериофаги: моновалентные, поливалентные (пиобактериофаг, интестифаг)
Диагностические бактериофаги: видовые, типовые.
Применение бактериофагов
Лечебные бактериофаги: моновалентные, поливалентные (пиобактериофаг, интестифаг)
Диагностические бактериофаги: видовые, типовые.

Лечебные бактериофаги
Препарат бактериофага: фильтрат бульонной культуры, зараженной соответствующим фагом.
Препараты назначаются с
Лечебные бактериофаги
Препарат бактериофага: фильтрат бульонной культуры, зараженной соответствующим фагом.
Препараты назначаются с
 Белки. Переваривание и всасывание
Белки. Переваривание и всасывание Бактерии и их виды (часть 1)
Бактерии и их виды (часть 1) Презентация к уроку биологии в 8 классе транспортные системы организма
Презентация к уроку биологии в 8 классе транспортные системы организма Антропогенез. Человеческие расы
Антропогенез. Человеческие расы Утворення перлин
Утворення перлин Характеристика семейства Розоцветные
Характеристика семейства Розоцветные Презентация к уроку 5 класса ФГОС
Презентация к уроку 5 класса ФГОС Соцветия. Виды соцветий
Соцветия. Виды соцветий Мир как система. Системный подход в современной науке
Мир как система. Системный подход в современной науке Which statement about the properties of life is false?
Which statement about the properties of life is false? Цитоплазма. Химический состав
Цитоплазма. Химический состав Выборка вещи. Контрастный метод
Выборка вещи. Контрастный метод Изучение строения скелета человека
Изучение строения скелета человека Презентация к уроку биологии 8 класс Кожа. Строение и функции
Презентация к уроку биологии 8 класс Кожа. Строение и функции Язык животных
Язык животных Вода в жизни человека
Вода в жизни человека Тип Плоские черви
Тип Плоские черви Видоизменения побегов
Видоизменения побегов Клетка. Клеточная теория. Строение и функции клетки, органоиды клетки
Клетка. Клеточная теория. Строение и функции клетки, органоиды клетки Энергетический и пластический обмен
Энергетический и пластический обмен Класс насекомые
Класс насекомые Самые красивые цветы. Творческое название: красивейшие цветы. 6 класс
Самые красивые цветы. Творческое название: красивейшие цветы. 6 класс Закономерности наследования, установленные Г. Менделем
Закономерности наследования, установленные Г. Менделем Терминология и символика, используемая в генетике.
Терминология и символика, используемая в генетике. Вирусы
Вирусы Закономерности формирования и наследования признаков. Изменчивость
Закономерности формирования и наследования признаков. Изменчивость Соцветия. Виды соцветий
Соцветия. Виды соцветий Биофизика цветного зрения
Биофизика цветного зрения