- Липиды: функции и обмен

Содержание
- 2. ЛИПИДЫ (от греч. lipos - жир) объединяют неоднородную группу органических соединений биологической природы, которым присуще одно
- 3. Липиды по физиологическому значению делят на Структурные: в комплексе с белками формируют биологические мембраны, определяют их
- 4. Многокомпонентные липиды А) Простые липиды: воски; ацилдиолы; ацилглицеролы. Б) Сложные или смешанные липиды: диольные фосфолипиды; глицерофосфолипиды;
- 5. Классификация жиров
- 6. Биологическая роль липидов Энергетическая: 1 г жира = 39кДж. Самые энергоемкие. Энергия окисления жиров используется во
- 7. Жирные кислоты - длинноцепочечные органические кислоты, содержат одну полярную карбоксильную группу и углеводородный радикал, в состав
- 8. Жирные кислоты: - насыщенные (масляная, пальмитиновая, стеариновая) (не содержат двойных связей) - ненасыщенные (олеиновая, линолевая, линоленовая)
- 9. Функции ЖК 1. Жирные кислоты являются строительными блоками для фосфолипидов и гликолипидов. Эти амфипатические молекулы являются
- 10. 3. Жирные кислоты являются топливными молекулами. Они запасаются в виде триацилглицеролов. При их освобождении и окислении
- 11. Функции незаменимых ЖК: 1) из них образуются биорегуляторы - эйкозаноиды; 2) необходимы для построения мембран (обеспечивают
- 12. Эйкозаноиды Эйкозаноиды - это производные эйкозаполиеновых жирных кислот, т.е. С20-жирных кислот (арахидоновой кислоты). Их делят на
- 13. Классификация эйкозаноидов Эйкозаноиды Простаноиды Лейкотриены простагландины тромбоксаны простациклины
- 14. Схема синтеза
- 15. ПРОСТАГЛАНДИНЫ Функции (используемые для терапии) : расширение сосудов, снижение артериального давления ингибиторы тромбообразования PgF2 прерывает беременность
- 16. Биологическая активность эйкозаноидов Эйкозаноиды – локальные биорегуляторы, действуют путем связывания с мембранными рецепторами в непосредственной близости
- 17. Переваривание глицерофосфолипидов Распад глицерофосфолипидов происходит в кишечнике при участии фосфолипаз, секретируемых поджелудочной железой. Известно несколько типов
- 18. Фосфолипаза А2 катализирует гидролитическое отщепление жирной кислоты во втором положении глицерофосфолипида. В результате действия фосфолипазы А2
- 19. Переваривание липидов
- 20. Таким образом, в результате действия фосфолипаз глицерофосфолипиды расщепляются до глицерола, высших жирных кислот, азотистого основания и
- 21. Транспорт липидов Жиры гидрофобны, поэтому существуют специальные механизмы их транспорта в крови. Свободные (неэстерифицированные) жирные кислоты
- 22. Строение липопротеинов Липопротеины являются молекулярными комплексами, состоящими из липидов и белков. Существует несколько классов липопротеинов (ЛП),
- 23. Структура липопротеина
- 24. Функции аполипопротеинов 1) являются структурными компонентами липопротеинов; 2) участвуют в узнавании и взаимодействии с рецепторами мембран;
- 25. Липопротеины подразделяются на 4 основные класса в зависимости от плотности (определяемой с помощью ультрацентрифугирования) и электрофоретической
- 26. Сложные липиды и стероиды
- 27. Ацилглицеролы Ацилглицеролы (ацилглицерины, нейтральные жиры) – это сложные эфиры трехатомного спирта глицерола и высших жирных кислот.
- 28. Общая формула ацилглицерола
- 29. Сложные триацилглицеролы В состав входят остатки разных ЖК, например, 1-пальмитоил-2-стеароил-3-олеилглицерол.
- 30. Значение Животные жиры и растительные масла являются важнейшими составляющими пищи человека. ЖК и глицерол при биохимическом
- 31. Воски сложные эфиры высших многоатомных спиртов и высших жирных кислот с примесью свободных жирных кислот, спиртов,
- 32. Фосфолипиды - это сложные эфиры различных многоатомных и аминоспиртов с жирными кислотами и фосфорной кислотой основные
- 33. Х= -СH2-CH2-N(+)(СН3)3 - фосфатидилхолины Х= -СН2-СН2-NH2 - фосфатидилэтаноламины Х= -СН2-СН(NH2)COOH - фосфатидилсерины Х= -СН2-СН(ОН)-СН2-ОН - фосфатидилглицерины
- 35. Гликолипиды ГЛИКОСФИНГОЛИПИДЫ отличаются от фосфолипидов: - нет остатка фосфорной кислоты - есть моносахарид или его производное
- 36. Стероиды - высокомолекулярные полициклические спирты ( неомыляемы и не способны к гидролизу)
- 37. Эфиры с жирными кислотами – стерины Роль холестерола: его производные образуют биологически активные вещества, желчные кислоты,
- 38. Желчные кислоты Это производные холановой кислоты или С24-стероиды. Они являются основными продуктами метаболизма холестерола, синтезируются в
- 40. ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЛИПИДОВ 1) гидролиз ТАГ – липолиз 2) окисление ЖК и глицерина до конечных продуктов
- 41. 1) Липолиз Мобилизация ТАГ из депо – гидролитическое расщепление липидов до СЖК и глицерина, которые используются
- 42. 2) β-окисление жирных кислот Впервые механизм изучен в 1904 г. Кнопом ЖК окисляется по β-атому С:
- 44. 2 этап – непосредственно β-окисление R-CH2-CH2-CO~SКоА R-CH=CH-CO~SКоА R-CHОН-CH2-CO~SКоА R-CО-CH2-CO~SКоА R-CО~SКоА + CH3-CO~SКоА пальмитиновая кислота 16С=14С+2С НАД
- 45. Окисление ненасыщенных жирных кислот При окислении ненасыщенных жирных кислот возникает потребность клетки в дополнительных ферментах изомеразах:
- 46. Окисление жирных кислот с нечетным числом атомов углерода Жирные кислоты с нечетным числом углеродов поступают в
- 47. РАСПАД ГЛИЦЕРОЛА до СО2 и Н2О Ферменты глицеролкиназа (- 1 АТФ) глицерофосфатдегидрогеназа (НАД+ → НАДН) (+
- 48. Предшественники для синтеза липидов ДОАФ (промежуточный продукт гликолиза) – для глицерола, если образование липидов происходит из
- 49. 3) Синтез жирных кислот При синтезе наращиваются 2С-фрагменты и затем восстанавливаются с участием НАДФН. Работает мультиферментный
- 50. 1 этап – образование малонил-КоА СН3СО~КоА + СО2 → СООН-СН2СО~КоА Фермент карбоксилаза (кофермент – биотин) использует
- 51. Синтез триацилглицеролов 2 пути синтеза ТАГ: Печень использует оба пути 1) + 2) 1) на основе
- 52. Транспорт жиров из печени В печени синтезируется большое количество различных видов эндогенного жира. Жир выводится из
- 53. 5) Синтез фосфолипидов Фосфатидная кислота – ключевой продукт синтеза До стадии фосфатидной кислоты синтез как у
- 54. СИНТЕЗ ХОЛЕСТЕРОЛА Холестерол – стероид, характерный только для животных организмов. 50% содержится в печени, 20% в
- 55. II этап – мевалоновая кислота конденсируется в сквален Сначала мевалоновая кислота теряет СО2 и превращается в
- 56. Все клетки организма способны синтезировать холестерол Синтез в организме : 50% образуется в печени, около 15%
- 57. ГМГ-редуктаза ключевой фермент синтеза холестерола, его активность регулируется: высокие концентрации ХС блокируют синтез фермента в печени
- 58. Основные виды патологии липидного обмена Желчекаменная болезнь Холестерол является компонентом желчных кислот и может находиться в
- 59. Атеросклероз При повреждении эндотелия сосудов ЛП проникают в сосудистую стенку и поглощаются фагоцитами. Все их составные
- 60. • холестериновый коэффициент атерогенности k = ХС общ – ХС ЛПВП ХС – концентрация общего холестерола,
- 62. Скачать презентацию

ЛИПИДЫ
(от греч. lipos - жир) объединяют неоднородную группу органических соединений биологической
ЛИПИДЫ
(от греч. lipos - жир) объединяют неоднородную группу органических соединений биологической

Липиды по физиологическому значению делят на
Структурные: в комплексе с белками формируют
Липиды по физиологическому значению делят на
Структурные: в комплексе с белками формируют

Многокомпонентные липиды
А) Простые липиды:
воски;
ацилдиолы;
ацилглицеролы.
Б) Сложные или смешанные липиды:
диольные фосфолипиды;
глицерофосфолипиды;
сфингофосфолипиды;
гликолипиды.
Многокомпонентные липиды
А) Простые липиды:
воски;
ацилдиолы;
ацилглицеролы.
Б) Сложные или смешанные липиды:
диольные фосфолипиды;
глицерофосфолипиды;
сфингофосфолипиды;
гликолипиды.
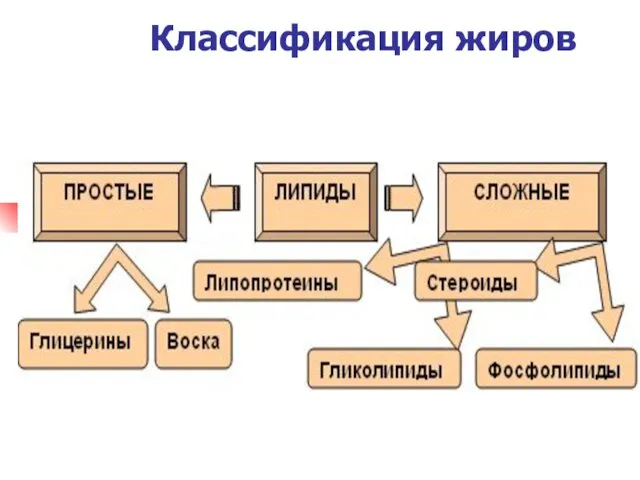
Классификация жиров
Классификация жиров

Биологическая роль липидов
Энергетическая: 1 г жира = 39кДж.
Биологическая роль липидов
Энергетическая: 1 г жира = 39кДж.

Жирные кислоты -
длинноцепочечные органические кислоты, содержат одну полярную карбоксильную
Жирные кислоты -
длинноцепочечные органические кислоты, содержат одну полярную карбоксильную
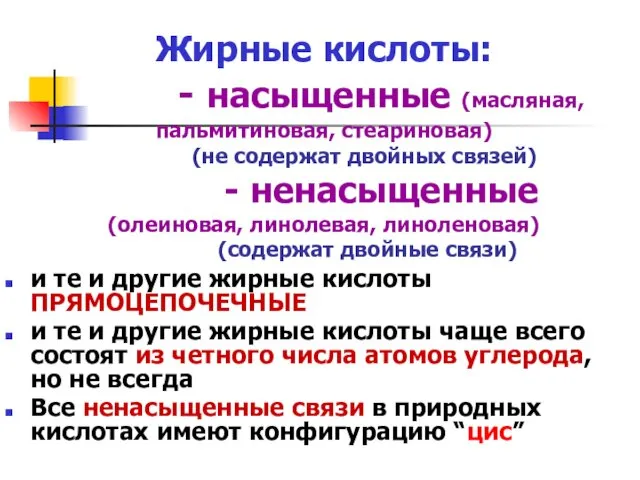
Жирные кислоты: - насыщенные (масляная, пальмитиновая, стеариновая) (не содержат двойных
Жирные кислоты: - насыщенные (масляная, пальмитиновая, стеариновая) (не содержат двойных

Функции ЖК
1. Жирные кислоты являются строительными блоками для фосфолипидов и гликолипидов.
Функции ЖК
1. Жирные кислоты являются строительными блоками для фосфолипидов и гликолипидов.

3. Жирные кислоты являются топливными молекулами. Они запасаются в виде триацилглицеролов.
3. Жирные кислоты являются топливными молекулами. Они запасаются в виде триацилглицеролов.

Функции незаменимых ЖК:
1) из них образуются биорегуляторы - эйкозаноиды;
2) необходимы
Функции незаменимых ЖК:
1) из них образуются биорегуляторы - эйкозаноиды;
2) необходимы

Эйкозаноиды
Эйкозаноиды - это производные эйкозаполиеновых жирных кислот, т.е. С20-жирных кислот (арахидоновой
Эйкозаноиды
Эйкозаноиды - это производные эйкозаполиеновых жирных кислот, т.е. С20-жирных кислот (арахидоновой
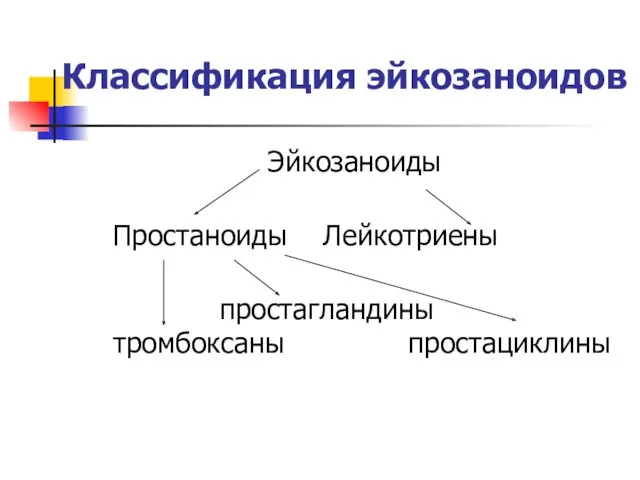
Классификация эйкозаноидов
Эйкозаноиды
Простаноиды Лейкотриены
простагландины тромбоксаны простациклины
Классификация эйкозаноидов
Эйкозаноиды
Простаноиды Лейкотриены
простагландины тромбоксаны простациклины
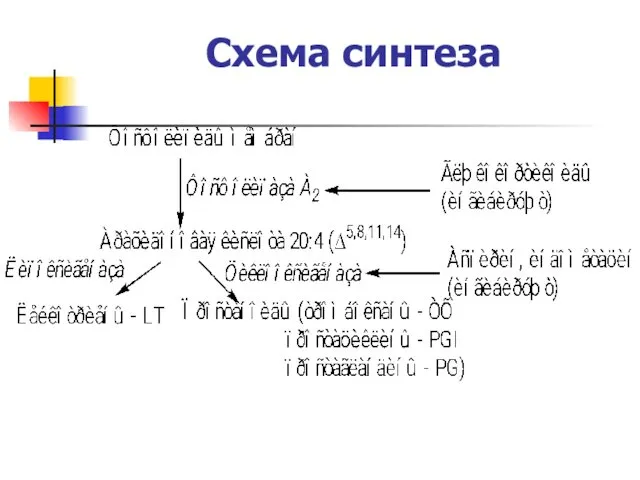
Схема синтеза
Схема синтеза
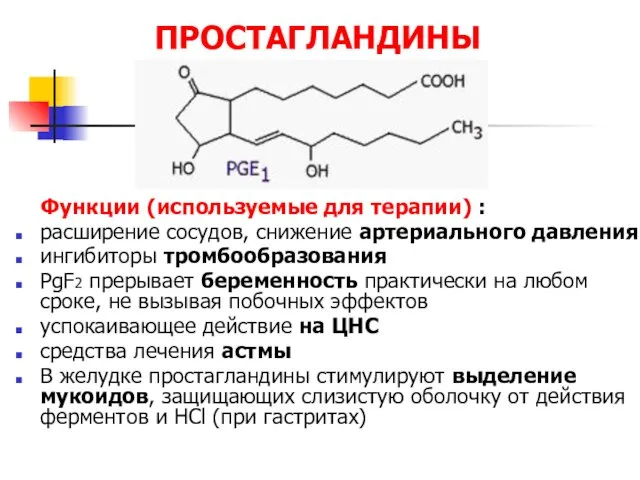
ПРОСТАГЛАНДИНЫ
Функции (используемые для терапии) :
расширение сосудов, снижение артериального давления
ингибиторы тромбообразования
PgF2
ПРОСТАГЛАНДИНЫ
Функции (используемые для терапии) :
расширение сосудов, снижение артериального давления
ингибиторы тромбообразования
PgF2

Биологическая активность эйкозаноидов
Эйкозаноиды – локальные биорегуляторы, действуют
путем связывания
Биологическая активность эйкозаноидов
Эйкозаноиды – локальные биорегуляторы, действуют путем связывания

Переваривание глицерофосфолипидов
Распад глицерофосфолипидов происходит в кишечнике при участии фосфолипаз, секретируемых
Переваривание глицерофосфолипидов
Распад глицерофосфолипидов происходит в кишечнике при участии фосфолипаз, секретируемых

Фосфолипаза А2 катализирует гидролитическое отщепление жирной кислоты во втором положении глицерофосфолипида.
Фосфолипаза А2 катализирует гидролитическое отщепление жирной кислоты во втором положении глицерофосфолипида.
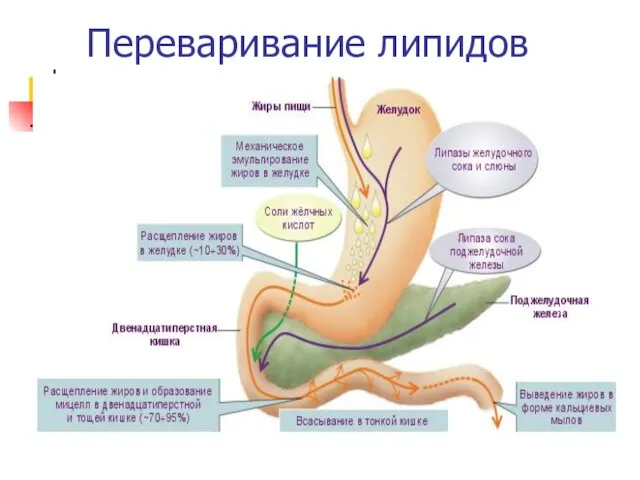
Переваривание липидов
Переваривание липидов

Таким образом, в результате действия фосфолипаз глицерофосфолипиды расщепляются до глицерола, высших
Таким образом, в результате действия фосфолипаз глицерофосфолипиды расщепляются до глицерола, высших

Транспорт липидов
Жиры гидрофобны, поэтому существуют специальные механизмы их транспорта
Транспорт липидов
Жиры гидрофобны, поэтому существуют специальные механизмы их транспорта

Строение липопротеинов
Липопротеины являются молекулярными комплексами, состоящими из липидов и белков.
Существует
Строение липопротеинов
Липопротеины являются молекулярными комплексами, состоящими из липидов и белков.
Существует
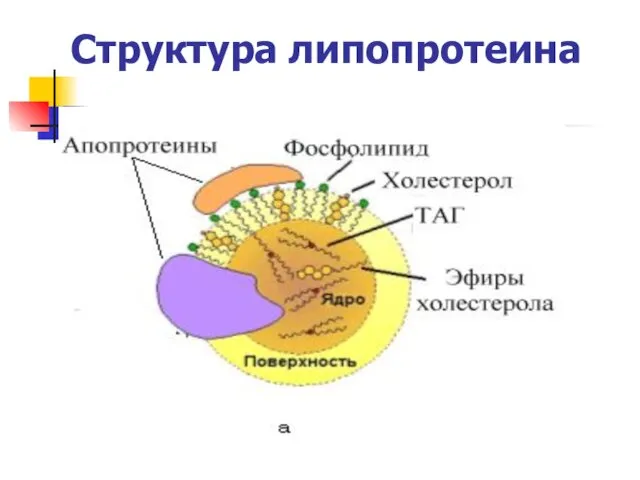
Структура липопротеина
Структура липопротеина

Функции аполипопротеинов
1) являются структурными компонентами липопротеинов;
2) участвуют в узнавании и
Функции аполипопротеинов
1) являются структурными компонентами липопротеинов;
2) участвуют в узнавании и

Липопротеины подразделяются на 4 основные класса в зависимости от плотности (определяемой
Липопротеины подразделяются на 4 основные класса в зависимости от плотности (определяемой

Сложные липиды и стероиды
Сложные липиды и стероиды

Ацилглицеролы
Ацилглицеролы (ацилглицерины, нейтральные жиры) – это сложные эфиры трехатомного спирта
Ацилглицеролы
Ацилглицеролы (ацилглицерины, нейтральные жиры) – это сложные эфиры трехатомного спирта
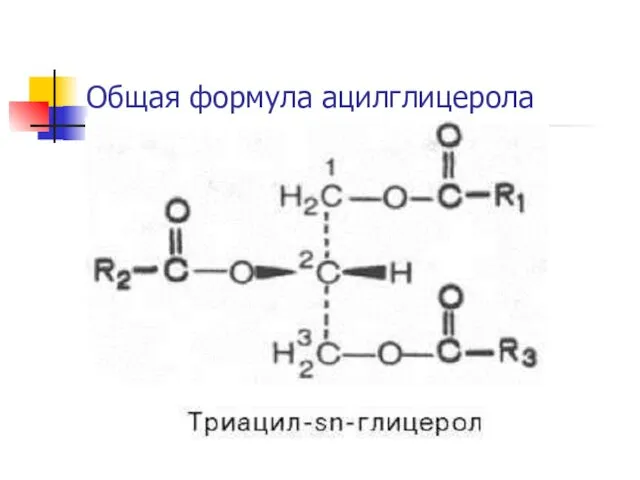
Общая формула ацилглицерола
Общая формула ацилглицерола
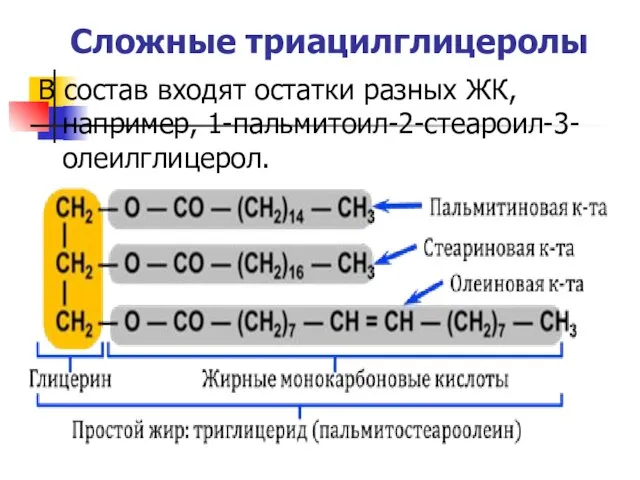
Сложные триацилглицеролы
В состав входят остатки разных ЖК, например, 1-пальмитоил-2-стеароил-3-олеилглицерол.
Сложные триацилглицеролы
В состав входят остатки разных ЖК, например, 1-пальмитоил-2-стеароил-3-олеилглицерол.

Значение
Животные жиры и растительные масла являются важнейшими составляющими пищи человека. ЖК
Значение
Животные жиры и растительные масла являются важнейшими составляющими пищи человека. ЖК

Воски
сложные эфиры высших многоатомных
спиртов и высших жирных кислот
Воски
сложные эфиры высших многоатомных
спиртов и высших жирных кислот

Фосфолипиды
- это сложные эфиры различных многоатомных и аминоспиртов с жирными
Фосфолипиды
- это сложные эфиры различных многоатомных и аминоспиртов с жирными
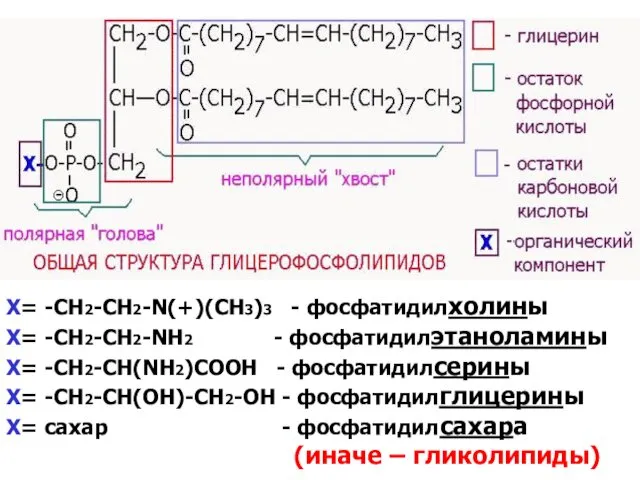
Х= -СH2-CH2-N(+)(СН3)3 - фосфатидилхолины
Х= -СН2-СН2-NH2 - фосфатидилэтаноламины
Х= -СН2-СН(NH2)COOH - фосфатидилсерины
Х=
Х= -СH2-CH2-N(+)(СН3)3 - фосфатидилхолины
Х= -СН2-СН2-NH2 - фосфатидилэтаноламины
Х= -СН2-СН(NH2)COOH - фосфатидилсерины
Х=
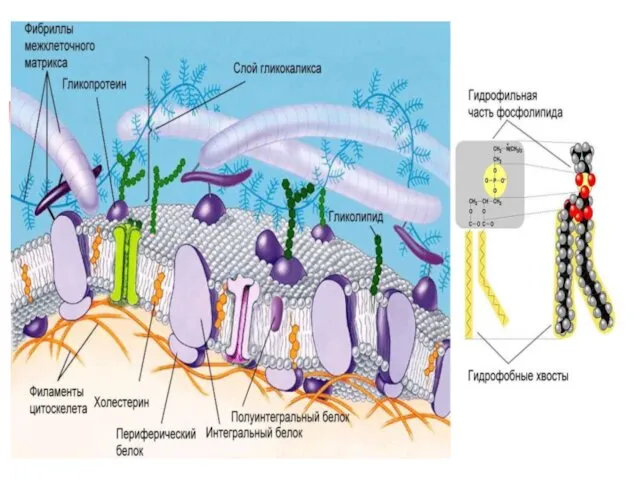

Гликолипиды
ГЛИКОСФИНГОЛИПИДЫ отличаются от фосфолипидов: - нет остатка фосфорной кислоты -
Гликолипиды
ГЛИКОСФИНГОЛИПИДЫ отличаются от фосфолипидов: - нет остатка фосфорной кислоты -
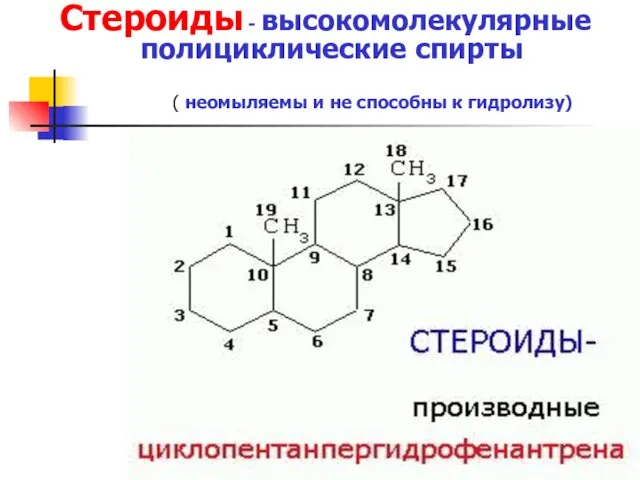
Стероиды - высокомолекулярные полициклические спирты
( неомыляемы и не способны
Стероиды - высокомолекулярные полициклические спирты
( неомыляемы и не способны
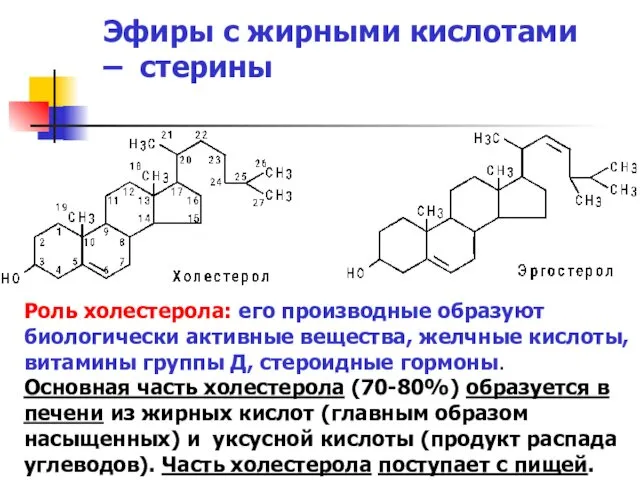
Эфиры с жирными кислотами – стерины
Роль холестерола: его производные образуют биологически
Эфиры с жирными кислотами – стерины
Роль холестерола: его производные образуют биологически

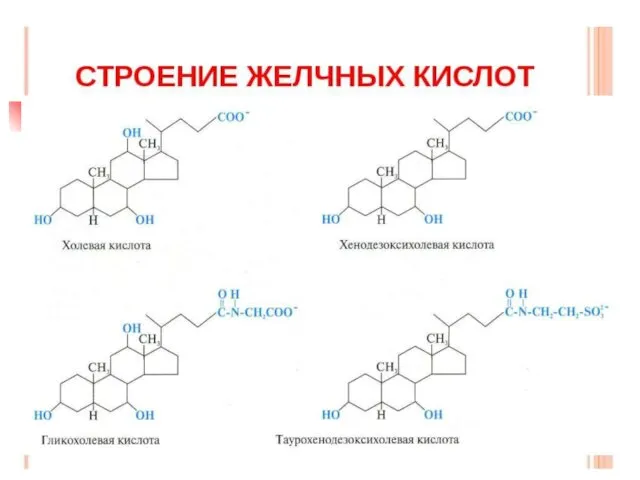
Желчные кислоты
Это производные холановой кислоты или С24-стероиды. Они являются основными продуктами
Желчные кислоты
Это производные холановой кислоты или С24-стероиды. Они являются основными продуктами

ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЛИПИДОВ
1) гидролиз ТАГ – липолиз
2) окисление ЖК
ВНУТРИКЛЕТОЧНЫЙ ОБМЕН ЛИПИДОВ
1) гидролиз ТАГ – липолиз
2) окисление ЖК

1) Липолиз
Мобилизация ТАГ из депо – гидролитическое расщепление липидов до СЖК
1) Липолиз
Мобилизация ТАГ из депо – гидролитическое расщепление липидов до СЖК

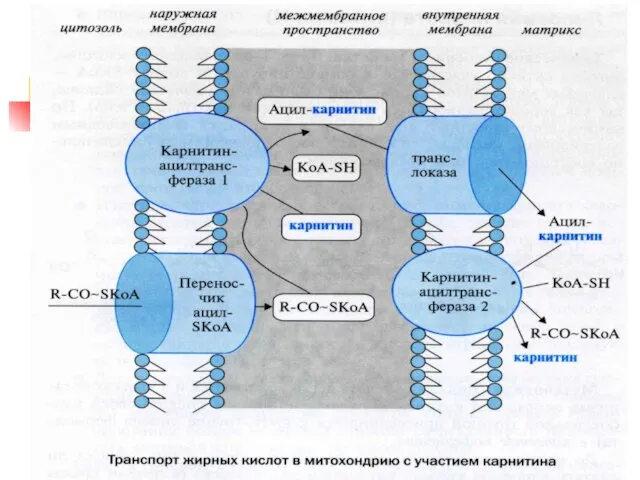
2) β-окисление жирных кислот
Впервые механизм изучен в 1904 г.
2) β-окисление жирных кислот
Впервые механизм изучен в 1904 г.

2 этап – непосредственно β-окисление
R-CH2-CH2-CO~SКоА
R-CH=CH-CO~SКоА
R-CHОН-CH2-CO~SКоА
R-CО-CH2-CO~SКоА
R-CО~SКоА
2 этап – непосредственно β-окисление
R-CH2-CH2-CO~SКоА
R-CH=CH-CO~SКоА
R-CHОН-CH2-CO~SКоА
R-CО-CH2-CO~SКоА
R-CО~SКоА

Окисление ненасыщенных жирных кислот
При окислении ненасыщенных жирных кислот возникает потребность
Окисление ненасыщенных жирных кислот
При окислении ненасыщенных жирных кислот возникает потребность

Окисление жирных кислот с нечетным числом атомов углерода
Жирные кислоты с
Окисление жирных кислот с нечетным числом атомов углерода
Жирные кислоты с
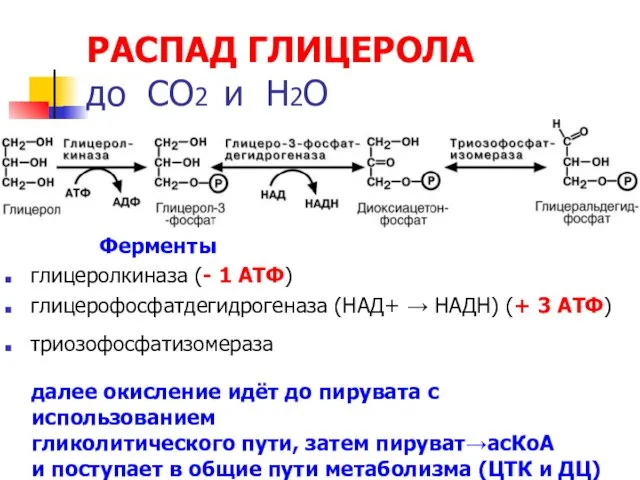
РАСПАД ГЛИЦЕРОЛА
до СО2 и Н2О
Ферменты
глицеролкиназа (- 1 АТФ)
глицерофосфатдегидрогеназа (НАД+ →
РАСПАД ГЛИЦЕРОЛА
до СО2 и Н2О
Ферменты
глицеролкиназа (- 1 АТФ)
глицерофосфатдегидрогеназа (НАД+ →

Предшественники
для синтеза липидов
ДОАФ (промежуточный продукт гликолиза) – для глицерола, если
Предшественники
для синтеза липидов
ДОАФ (промежуточный продукт гликолиза) – для глицерола, если

3) Синтез жирных кислот
При синтезе наращиваются 2С-фрагменты
и затем восстанавливаются
3) Синтез жирных кислот
При синтезе наращиваются 2С-фрагменты и затем восстанавливаются

1 этап – образование малонил-КоА
СН3СО~КоА + СО2 → СООН-СН2СО~КоА
Фермент карбоксилаза (кофермент
1 этап – образование малонил-КоА
СН3СО~КоА + СО2 → СООН-СН2СО~КоА
Фермент карбоксилаза (кофермент

Синтез триацилглицеролов
2 пути синтеза ТАГ:
Печень использует оба пути 1)
Синтез триацилглицеролов
2 пути синтеза ТАГ: Печень использует оба пути 1)

Транспорт жиров из печени
В печени синтезируется большое количество различных видов эндогенного
Транспорт жиров из печени
В печени синтезируется большое количество различных видов эндогенного

5) Синтез фосфолипидов
Фосфатидная кислота – ключевой продукт синтеза
До стадии
5) Синтез фосфолипидов
Фосфатидная кислота – ключевой продукт синтеза До стадии

СИНТЕЗ ХОЛЕСТЕРОЛА
Холестерол – стероид, характерный только для животных организмов.
50%
СИНТЕЗ ХОЛЕСТЕРОЛА
Холестерол – стероид, характерный только для животных организмов. 50%

II этап – мевалоновая кислота конденсируется в сквален
Сначала мевалоновая
II этап – мевалоновая кислота конденсируется в сквален
Сначала мевалоновая
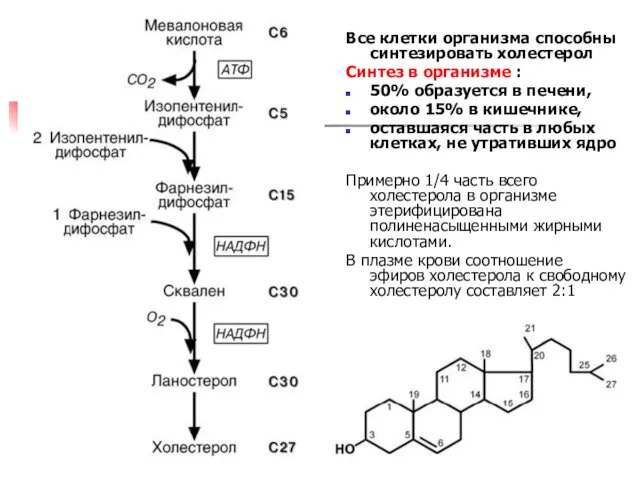
Все клетки организма способны синтезировать холестерол
Синтез в организме :
50% образуется
Все клетки организма способны синтезировать холестерол
Синтез в организме :
50% образуется

ГМГ-редуктаза ключевой фермент синтеза
холестерола, его активность регулируется:
высокие концентрации ХС
ГМГ-редуктаза ключевой фермент синтеза
холестерола, его активность регулируется:
высокие концентрации ХС

Основные виды патологии липидного обмена
Желчекаменная болезнь
Холестерол является компонентом желчных кислот
Основные виды патологии липидного обмена
Желчекаменная болезнь Холестерол является компонентом желчных кислот

Атеросклероз
При повреждении эндотелия сосудов ЛП проникают в сосудистую стенку и поглощаются
Атеросклероз
При повреждении эндотелия сосудов ЛП проникают в сосудистую стенку и поглощаются

• холестериновый коэффициент атерогенности
k = ХС общ – ХС
• холестериновый коэффициент атерогенности
k = ХС общ – ХС
 Бионика
Бионика Перспективы развития биологии
Перспективы развития биологии Положение человека в системе животного мира
Положение человека в системе животного мира Регуляция матричных синтезов. Ингибиторы матричных синтезов
Регуляция матричных синтезов. Ингибиторы матричных синтезов Скелет туловища
Скелет туловища Роль гормонов в обмене веществ, росте и развитии организма
Роль гормонов в обмене веществ, росте и развитии организма Кровеносная система
Кровеносная система Особенности адаптации респираторной системы к длительным физическим нагрузкам
Особенности адаптации респираторной системы к длительным физическим нагрузкам Ящерицы на территории эко-сквера
Ящерицы на территории эко-сквера Клетка как биологическая система. Подготовка к ЕГЭ
Клетка как биологическая система. Подготовка к ЕГЭ Живая и неживая природа (начальная школа)
Живая и неживая природа (начальная школа) Екотоксикологічна характеристика мікотоксинів
Екотоксикологічна характеристика мікотоксинів Птицы. Забота о потомстве
Птицы. Забота о потомстве Артерии большого и малого круга кровообращения
Артерии большого и малого круга кровообращения Животные живого уголка
Животные живого уголка Тип Arthropoda – Членистоногие
Тип Arthropoda – Членистоногие Парад лета
Парад лета Биология наука о жизни
Биология наука о жизни Топографическая анатомия наружного, среднего и внутреннего уха
Топографическая анатомия наружного, среднего и внутреннего уха Полевые культуры как экологическая система. Понятие биологического потенциала продуктивности растений
Полевые культуры как экологическая система. Понятие биологического потенциала продуктивности растений Презентация по теме Кайнозойская эра, 11 класс
Презентация по теме Кайнозойская эра, 11 класс Елементи біофізики при вивченні електрики
Елементи біофізики при вивченні електрики Самые опасные животные в мире
Самые опасные животные в мире Физиология кровообращения
Физиология кровообращения Презентация Скелет и мускулатура птиц
Презентация Скелет и мускулатура птиц Адам миының құрылысы және қызметі
Адам миының құрылысы және қызметі Голоса птиц
Голоса птиц Статистические закономерности модификационной изменчивости
Статистические закономерности модификационной изменчивости