- Механизмы биоэлектрогенеза

Содержание
- 2. ВОПРОСЫ 1. Ионные насосы и ионные каналы плазмолеммы. 2. Потенциал покоя. 3. Потенциал действия
- 3. Интеграция организма и вся жизнедеятельность, включая мышление, мышечное сокращение, есть процесс электрический
- 4. Три типа потенциалов
- 5. Луиджи Гальвани 1737-1798 (1791 книга) и Алессандро Вольта
- 6. Алан Ходжкин и Эндрью Хаксли
- 7. Стивен Каффлер, Джон Экклс и Бернард Катц
- 8. В.Маунткастл и Анри Лабори
- 9. Рефлекторные симпатические реакции от рецепторов и афферентных волокон кишки
- 10. Кальмар
- 11. Электровозбудимые клетки: нейроны, миоциты, гландоциты: имеют потенциал покоя ПП и потенциал действия ПД
- 12. Нервная ткань Нейроны Глиоциты
- 13. Электрофизиологические свойства нейронов 1.Потенциал покоя нейронов близок к 60-80 мВ, амплитуда ПД составляет 80-110 мВ.
- 14. Насосы
- 15. Ионы асимметрично распределены во внутренней среде
- 16. ИОННЫЕ ГРАДИЕНТЫ НУЖНЫ ДЛЯ: ЗАПАСАНИЯ ЭНЕРГИИ ГЕНЕРАЦИИ ВОЗБУЖДЕНИЯ ТРАНСПОРТА ГЛЮКОЗЫ И АМИНОКИСЛОТ
- 17. Ионные градиенты
- 18. Для создания потенциалов нужны ионы Na+, K+, Ca++, Cl- Na+ и K+ существенно отличаются по физико-химическим
- 19. физико-химические отличия ионов 1.Для создания электрохимического потенциала ионов K+ нужно на порядок меньше, чем Na+ 2.У
- 20. Ионная шуба натрия (он любит воду = гидрофил)
- 21. физико-химические отличия ионов Из-за высокой гидрофобности K+ проникает через мембрану в 5 раз легче, чем гидрофильный
- 22. физико-химические отличия ионов
- 24. Транспортеры и обменники
- 25. Цикл натриевой помпы
- 26. Структура натриевой помпы
- 27. Стехиометрия Для встречного перемещения (антипорта) натрия и калия Na-K-АТФазой характерна стехиометрия, для аксона кальмара равная 3/2.
- 28. 30% метаболизма Калий-натриевая помпа локализована в мембранах практически всех клеток организма, но относительно много молекул –
- 29. Са-АТФазы Кальциевый насос поддерживает содержание ионов Са++ в цитозоле на низком уровне. В качестве депо кальция
- 30. Са-АТФазы В саркоплазматической сети на долю Са-АТФазы приходится 60% общего мембранного белка. По-видимому, в мембране саркоплазматической
- 31. Кальциевые насосы
- 32. Кроме ионных помп есть каналы: отличия в том, что по каналам ионы идут по градиенту концентрации
- 33. Ионные каналы Некоторые из них являются высокоизбирательными для определенных ионов, их относят к классу селективных (например,
- 34. потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно осуществляется конформацией белковой молекулы при
- 35. Типы ионных каналов и их структура
- 36. Управление ионными каналами
- 37. Структура ионного канала
- 38. Schematic drawing of the voltage-gated Na channel α-subunit. Drawing (a) adapted from Goldin A(2002) The evolution
- 39. Voltage-gated K channel and its associated cytoplasmic β-subunit
- 40. Потенциал - управляемые каналы – воротный механизм Сенсор напряжения домен S4 Закрывает «Шар на веревочке»
- 41. Холинорецептор – лиганд-управляемый канал
- 42. Токи в одном канале: «все или ничего»
- 43. Формирование ПП
- 44. Формирование ПП
- 45. ПП При потенциале покоя внутренняя сторона клеточной мембраны имеет заряд, знак которого (отрицательность) определяется наличием в
- 46. Мембранная проницаемость ионных каналов создает ПП
- 47. ПП Из-за этого в клетке наблюдается избыток отрицательных ионов, и, следовательно, отрицательных зарядов, а в интерстиции
- 48. Уравнение Нернста
- 49. Имеет значение сума ионных концентраций
- 50. Механизм электрогенеза Уравнение Гольдмана Здесь R – газовая постоянная, T –температура, z – заряд иона, а
- 51. РП Равновесный потенциал – такой потенциал плазмолеммы клетки, при котором суммарный ток конкретного иона через мембрану
- 52. Равновесный потенциал Ек = –75 мВ; ЕNa = +55 мВ; ЕCa = +150 мВ; ЕCl =
- 53. Закон Ома I=U/R iNa = gNa (Vm – ENa), где gNa = 1/R – проводимость, величина,
- 54. Направление ионного тока iК = gК(Vm–EК) При Vm = EК ток равен 0 При Vm если
- 55. Функции ПП 1. Поляризация мембраны является условием для возбуждения и торможения. 2.Поляризация определяет объем выделения медиатора
- 57. Скачать презентацию

ВОПРОСЫ
1. Ионные насосы и ионные каналы плазмолеммы.
2. Потенциал покоя.
3. Потенциал
ВОПРОСЫ
1. Ионные насосы и ионные каналы плазмолеммы.
2. Потенциал покоя.
3. Потенциал

Интеграция организма и вся жизнедеятельность, включая мышление, мышечное сокращение, есть процесс
Интеграция организма и вся жизнедеятельность, включая мышление, мышечное сокращение, есть процесс

Три типа потенциалов
Три типа потенциалов

Луиджи Гальвани 1737-1798
(1791 книга)
и Алессандро Вольта
Луиджи Гальвани 1737-1798
(1791 книга)
и Алессандро Вольта

Алан Ходжкин и Эндрью Хаксли
Алан Ходжкин и Эндрью Хаксли

Стивен Каффлер, Джон Экклс и Бернард Катц
Стивен Каффлер, Джон Экклс и Бернард Катц

В.Маунткастл и Анри Лабори
В.Маунткастл и Анри Лабори
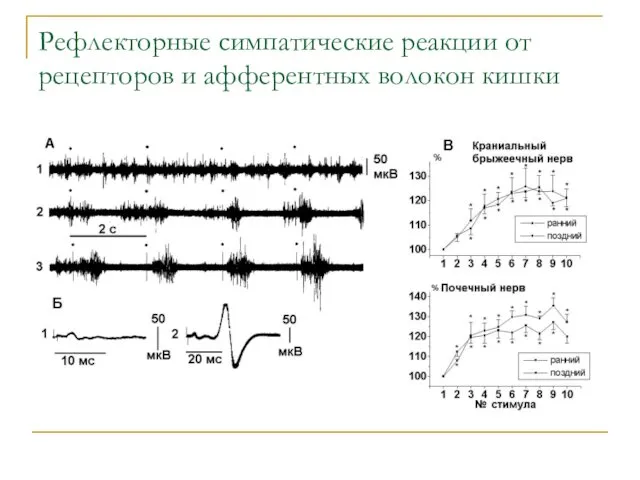
Рефлекторные симпатические реакции от рецепторов и афферентных волокон кишки
Рефлекторные симпатические реакции от рецепторов и афферентных волокон кишки

Кальмар
Кальмар
Электровозбудимые клетки: нейроны, миоциты, гландоциты: имеют потенциал покоя ПП и потенциал
Электровозбудимые клетки: нейроны, миоциты, гландоциты: имеют потенциал покоя ПП и потенциал

Нервная ткань
Нейроны
Глиоциты
Нервная ткань
Нейроны
Глиоциты

Электрофизиологические свойства нейронов
1.Потенциал покоя нейронов близок к 60-80 мВ, амплитуда ПД
Электрофизиологические свойства нейронов
1.Потенциал покоя нейронов близок к 60-80 мВ, амплитуда ПД

Насосы
Насосы
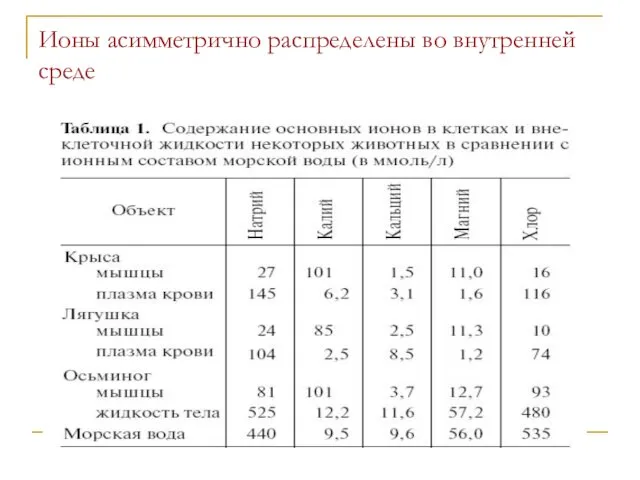
Ионы асимметрично распределены во внутренней среде
Ионы асимметрично распределены во внутренней среде

ИОННЫЕ ГРАДИЕНТЫ НУЖНЫ ДЛЯ:
ЗАПАСАНИЯ ЭНЕРГИИ
ГЕНЕРАЦИИ ВОЗБУЖДЕНИЯ
ТРАНСПОРТА ГЛЮКОЗЫ И АМИНОКИСЛОТ
ИОННЫЕ ГРАДИЕНТЫ НУЖНЫ ДЛЯ:
ЗАПАСАНИЯ ЭНЕРГИИ
ГЕНЕРАЦИИ ВОЗБУЖДЕНИЯ
ТРАНСПОРТА ГЛЮКОЗЫ И АМИНОКИСЛОТ

Ионные градиенты
Ионные градиенты

Для создания потенциалов нужны ионы Na+, K+, Ca++, Cl-
Na+ и K+
Для создания потенциалов нужны ионы Na+, K+, Ca++, Cl-
Na+ и K+

физико-химические отличия ионов
1.Для создания электрохимического потенциала ионов K+ нужно на порядок
физико-химические отличия ионов
1.Для создания электрохимического потенциала ионов K+ нужно на порядок

Ионная шуба натрия (он любит воду = гидрофил)
Ионная шуба натрия (он любит воду = гидрофил)

физико-химические отличия ионов
Из-за высокой гидрофобности K+ проникает через мембрану в 5
физико-химические отличия ионов
Из-за высокой гидрофобности K+ проникает через мембрану в 5
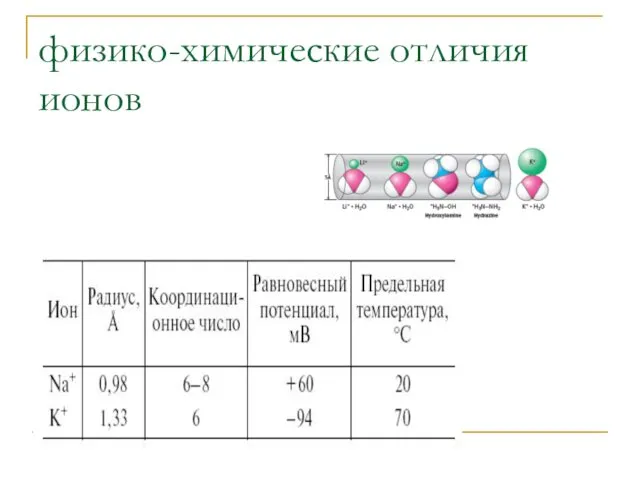
физико-химические отличия ионов
физико-химические отличия ионов
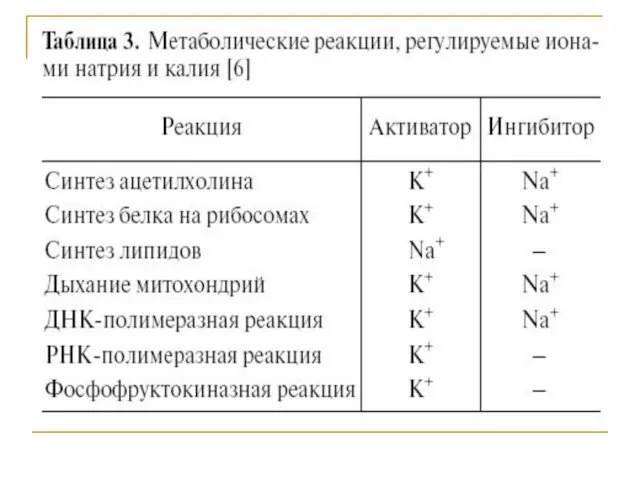

Транспортеры и обменники
Транспортеры и обменники

Цикл натриевой помпы
Цикл натриевой помпы

Структура натриевой помпы
Структура натриевой помпы

Стехиометрия
Для встречного перемещения (антипорта) натрия и калия Na-K-АТФазой характерна стехиометрия, для
Стехиометрия
Для встречного перемещения (антипорта) натрия и калия Na-K-АТФазой характерна стехиометрия, для

30% метаболизма
Калий-натриевая помпа локализована в мембранах практически всех клеток организма, но
30% метаболизма
Калий-натриевая помпа локализована в мембранах практически всех клеток организма, но

Са-АТФазы
Кальциевый насос поддерживает содержание ионов Са++ в цитозоле на низком уровне.
Са-АТФазы
Кальциевый насос поддерживает содержание ионов Са++ в цитозоле на низком уровне.

Са-АТФазы
В саркоплазматической сети на долю Са-АТФазы приходится 60% общего мембранного белка.
Са-АТФазы
В саркоплазматической сети на долю Са-АТФазы приходится 60% общего мембранного белка.

Кальциевые насосы
Кальциевые насосы

Кроме ионных помп есть каналы: отличия в том, что по каналам
Кроме ионных помп есть каналы: отличия в том, что по каналам

Ионные каналы
Некоторые из них являются высокоизбирательными для определенных ионов, их относят
Ионные каналы
Некоторые из них являются высокоизбирательными для определенных ионов, их относят

потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно
потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно

Типы ионных каналов и их структура
Типы ионных каналов и их структура

Управление ионными каналами
Управление ионными каналами

Структура ионного канала
Структура ионного канала

Schematic drawing of the voltage-gated Na channel α-subunit.
Drawing (a) adapted from
Schematic drawing of the voltage-gated Na channel α-subunit. Drawing (a) adapted from

Voltage-gated K channel and
its associated cytoplasmic β-subunit
Voltage-gated K channel and
its associated cytoplasmic β-subunit

Потенциал - управляемые каналы – воротный механизм
Сенсор напряжения домен S4
Закрывает «Шар
Потенциал - управляемые каналы – воротный механизм
Сенсор напряжения домен S4
Закрывает «Шар

Холинорецептор –
лиганд-управляемый канал
Холинорецептор –
лиганд-управляемый канал

Токи в одном канале: «все или ничего»
Токи в одном канале: «все или ничего»

Формирование ПП
Формирование ПП

Формирование ПП
Формирование ПП

ПП
При потенциале покоя внутренняя сторона клеточной мембраны имеет заряд, знак которого
ПП
При потенциале покоя внутренняя сторона клеточной мембраны имеет заряд, знак которого

Мембранная проницаемость ионных каналов создает ПП
Мембранная проницаемость ионных каналов создает ПП

ПП
Из-за этого в клетке наблюдается избыток отрицательных ионов, и, следовательно, отрицательных
ПП
Из-за этого в клетке наблюдается избыток отрицательных ионов, и, следовательно, отрицательных

Уравнение Нернста
Уравнение Нернста

Имеет значение сума ионных концентраций
Имеет значение сума ионных концентраций

Механизм электрогенеза
Уравнение Гольдмана
Здесь R – газовая постоянная, T –температура, z –
Механизм электрогенеза
Уравнение Гольдмана
Здесь R – газовая постоянная, T –температура, z –

РП
Равновесный потенциал – такой потенциал плазмолеммы клетки, при котором суммарный ток
РП
Равновесный потенциал – такой потенциал плазмолеммы клетки, при котором суммарный ток

Равновесный потенциал
Ек = –75 мВ;
ЕNa = +55 мВ;
ЕCa = +150 мВ;
ЕCl
Равновесный потенциал
Ек = –75 мВ;
ЕNa = +55 мВ;
ЕCa = +150 мВ;
ЕCl

Закон Ома I=U/R
iNa = gNa (Vm – ENa),
где gNa = 1/R
Закон Ома I=U/R
iNa = gNa (Vm – ENa),
где gNa = 1/R

Направление ионного тока
iК = gК(Vm–EК)
При Vm = EК ток равен 0
При
Направление ионного тока
iК = gК(Vm–EК)
При Vm = EК ток равен 0
При

Функции ПП
1. Поляризация мембраны является условием для возбуждения и торможения.
2.Поляризация определяет
Функции ПП
1. Поляризация мембраны является условием для возбуждения и торможения.
2.Поляризация определяет
 Биогеоценоз смешанного леса
Биогеоценоз смешанного леса Естественный отбор и борьба за существование
Естественный отбор и борьба за существование Чем живое отличается от неживго
Чем живое отличается от неживго Широколиственный лес
Широколиственный лес Мир бабочек. Бабочка из бумаги
Мир бабочек. Бабочка из бумаги Красноухая черепаха
Красноухая черепаха Видоизменения побегов растения
Видоизменения побегов растения Мир и человек. Живые обитатели планеты
Мир и человек. Живые обитатели планеты Тема урока: Кровообращение (9-й класс)
Тема урока: Кровообращение (9-й класс) Розпізнавання антигену специфічною імунною системою. Антитіла: структура і функції
Розпізнавання антигену специфічною імунною системою. Антитіла: структура і функції Грибы. Конкурс пословиц
Грибы. Конкурс пословиц презентация по биологии 6классАстрология комнатных растений
презентация по биологии 6классАстрология комнатных растений Бактериофаги
Бактериофаги Пищеварительная система
Пищеварительная система Влияние пищевых добавок на организм человека 1 часть
Влияние пищевых добавок на организм человека 1 часть Тип Хордовые. Ланцетник 7 класс
Тип Хордовые. Ланцетник 7 класс Пивной фронт: за кем победа?
Пивной фронт: за кем победа? Презентация к уроку биологии в 8 классе по теме История развития знаний о строении и функциях организма человека
Презентация к уроку биологии в 8 классе по теме История развития знаний о строении и функциях организма человека Животные. Интересные факты о животных
Животные. Интересные факты о животных Дыхательная система. Особенности строения, работа и гигиена органов дыхания
Дыхательная система. Особенности строения, работа и гигиена органов дыхания Ядовитые растения Чеченской Республики
Ядовитые растения Чеченской Республики портфолио учителя биологии Канунниковой Л.Н.
портфолио учителя биологии Канунниковой Л.Н. Язык животных
Язык животных Презентация к игре Биологическая викторина
Презентация к игре Биологическая викторина Отряд хищные
Отряд хищные Evolution 3 – Genetic Variation
Evolution 3 – Genetic Variation Тайга. Флора и фауна
Тайга. Флора и фауна Ягодные и декоративные кустарники
Ягодные и декоративные кустарники