Механизмы окислительного повреждения аминокислот, пептидов, белков, нуклеотидов и нуклеиновых кислот. Лекция 5 презентация
- Механизмы окислительного повреждения аминокислот, пептидов, белков, нуклеотидов и нуклеиновых кислот. Лекция 5

Содержание
- 2. Механизмы окислительной модификации белков (ОМБ) В качестве основных индукторов ОМБ, в первую очередь, рассматриваются активные формы
- 3. Окисление полипептидной цепи, приводящее к поперечному связыванию или разрыву белков
- 4. Основной агент, вызывающий окислительное повреждение белков, — гидроксил-радикал. Окислительная атака на белок начинается с удаления электрона
- 5. Все боковые остатки аминокислот подвержены окислению, но в разной степени. Наиболее уязвимы цистеин и метионин, однако
- 6. Присутствующие на поверхности многих белков остатки метионина можно рассматривать как встроенный в белок предохранитель от АФК,
- 7. Обратимое окисление остатков серусодержащих аминокислот метионина и цистеина в составе белков
- 8. Нерепарируемому окислению чаще других подвергаются лизин и аргинин. В результате их окисления, а также в результате
- 9. Необратимое окисление остатков лизина, аргинина, глутаминовой кислоты и пролина в составе белков
- 10. Окисление глутаминовой кислоты и пролина часто приводит к разрыву полипептидной цепи. Разрыв цепи, обусловленный окислением пролина,
- 11. В настоящее время предложены следующие механизмы ОМБ. Первый механизм ОМБ – коньюгация липидных пероксидов с аминокислотными
- 12. Наиболее важным следствием ОМБ белков является инактивация ферментов. Например, альдегиды вызывают инактивацию мембранных транспортеров, таких, как
- 13. Гликирование белков Гликирование, или неферментативное гликозилирование, — реакция между восстанавливающими углеводами (глюкоза, фруктоза и др.) и
- 14. Участие углеводов и белков в оксидативном стрессе Продукты Амадори – модифицированные белки (при образовании с углеводами
- 15. Взаимодействие белков, липидов и углеводов при оксидативном стрессе
- 16. Окислительные повреждения нуклеиновых кислот Очень тяжелые последствия имеет окисление нуклеиновых кислот, особенно ДНК. В этом случае
- 18. Наиболее распространенные окислительные модификации азотистых оснований, входящих в состав нуклеиновых кислот с образованием производных пиримидина. Кроме
- 19. Идентифицировано приблизительно 20 окислительных повреждений азотистых оснований. У пуринов присоединение гидроксил-радикала происходит к С4, С5 и
- 20. В клетках всех организмов существует многокомпонентная система репарации повреждений ДНК. Окисленные основания ДНК удаляются двумя способами.
- 22. Липофусцин (lipofuscinum; от греч. lipo – ≪жир≫ и лат. fuscus – ≪темный≫;), также известный как ≪пигмент
- 23. Липофусцин – гликопротеид, в состав которого входят: - Жиры - Аминокислоты - Ферменты - Флавиновые соединения
- 24. Ультраструктура липофусцина: Представляет из себя электронно-плотные гранулы, окруженные двойной мембраной, содержащей миелиноподобные структуры. Эти гранулы называют
- 26. Скачать презентацию

Механизмы окислительной модификации белков (ОМБ)
В качестве основных индукторов ОМБ, в первую
Механизмы окислительной модификации белков (ОМБ)
В качестве основных индукторов ОМБ, в первую
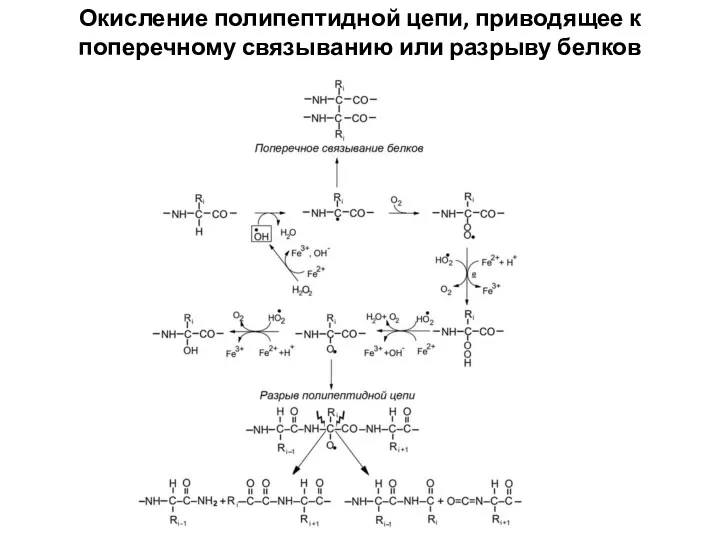
Окисление полипептидной цепи, приводящее к поперечному связыванию или разрыву белков
Окисление полипептидной цепи, приводящее к поперечному связыванию или разрыву белков

Основной агент, вызывающий окислительное повреждение белков, — гидроксил-радикал. Окислительная атака на
Основной агент, вызывающий окислительное повреждение белков, — гидроксил-радикал. Окислительная атака на

Все боковые остатки аминокислот подвержены окислению, но в разной степени. Наиболее
Все боковые остатки аминокислот подвержены окислению, но в разной степени. Наиболее

Присутствующие на поверхности многих белков остатки метионина можно рассматривать как
Присутствующие на поверхности многих белков остатки метионина можно рассматривать как
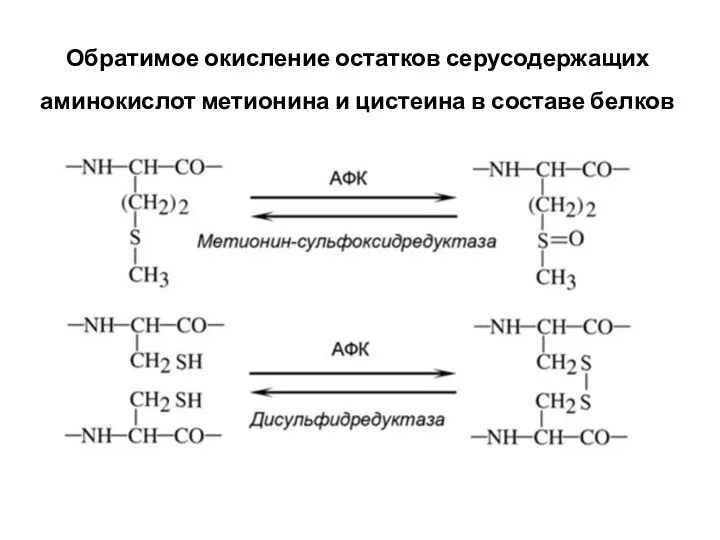
Обратимое окисление остатков серусодержащих аминокислот метионина и цистеина в составе белков
Обратимое окисление остатков серусодержащих аминокислот метионина и цистеина в составе белков

Нерепарируемому окислению чаще других подвергаются лизин и аргинин. В результате их
Нерепарируемому окислению чаще других подвергаются лизин и аргинин. В результате их
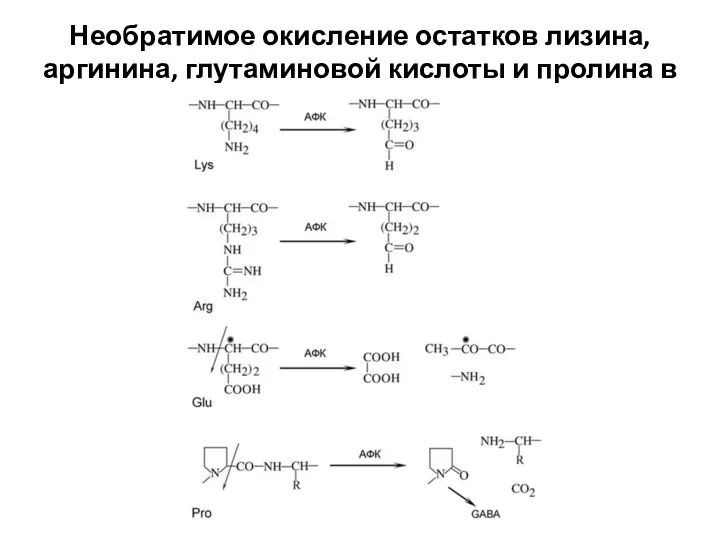
Необратимое окисление остатков лизина, аргинина, глутаминовой кислоты и пролина в составе
Необратимое окисление остатков лизина, аргинина, глутаминовой кислоты и пролина в составе

Окисление глутаминовой кислоты и пролина часто приводит к разрыву полипептидной цепи.
Окисление глутаминовой кислоты и пролина часто приводит к разрыву полипептидной цепи.

В настоящее время предложены следующие механизмы ОМБ.
Первый механизм ОМБ –
В настоящее время предложены следующие механизмы ОМБ.
Первый механизм ОМБ –

Наиболее важным следствием ОМБ белков является инактивация ферментов. Например, альдегиды вызывают
Наиболее важным следствием ОМБ белков является инактивация ферментов. Например, альдегиды вызывают

Гликирование белков
Гликирование, или неферментативное гликозилирование, — реакция между восстанавливающими углеводами (глюкоза, фруктоза и др.) и свободными аминогруппами белков, липидов и нуклеиновых кислот живого организма,
Гликирование белков
Гликирование, или неферментативное гликозилирование, — реакция между восстанавливающими углеводами (глюкоза, фруктоза и др.) и свободными аминогруппами белков, липидов и нуклеиновых кислот живого организма,
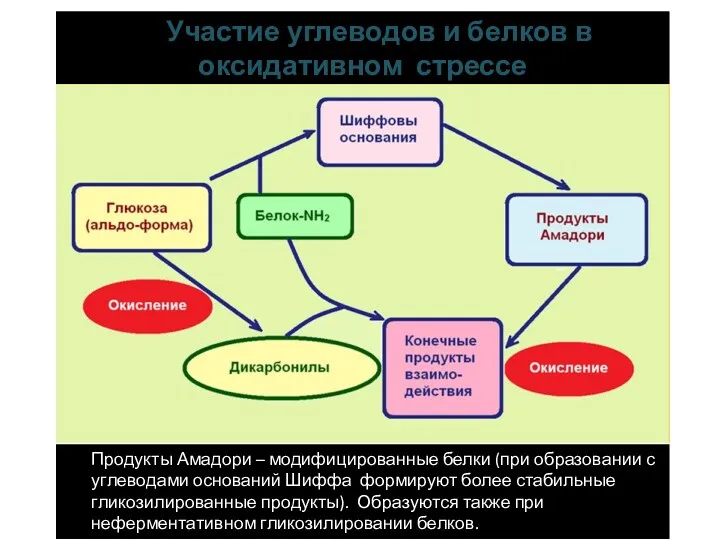
Участие углеводов и белков в оксидативном стрессе
Продукты Амадори – модифицированные
Участие углеводов и белков в оксидативном стрессе
Продукты Амадори – модифицированные
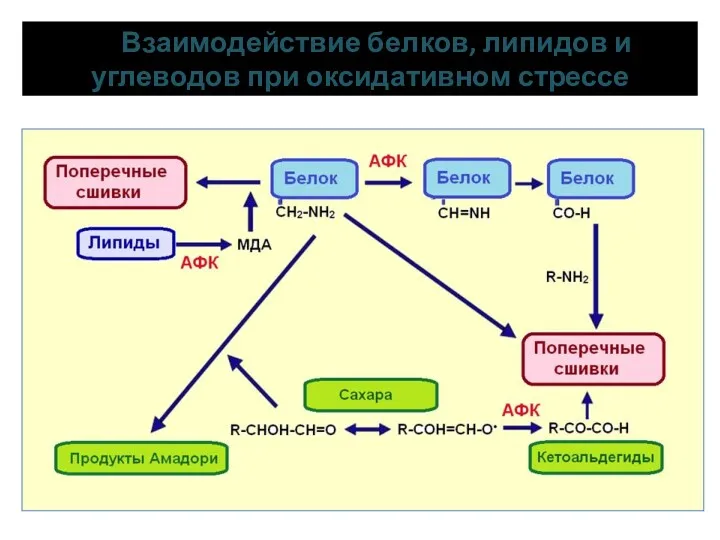
Взаимодействие белков, липидов и углеводов при оксидативном стрессе
Взаимодействие белков, липидов и углеводов при оксидативном стрессе

Окислительные повреждения нуклеиновых кислот
Очень тяжелые последствия имеет окисление нуклеиновых кислот, особенно
Окислительные повреждения нуклеиновых кислот
Очень тяжелые последствия имеет окисление нуклеиновых кислот, особенно
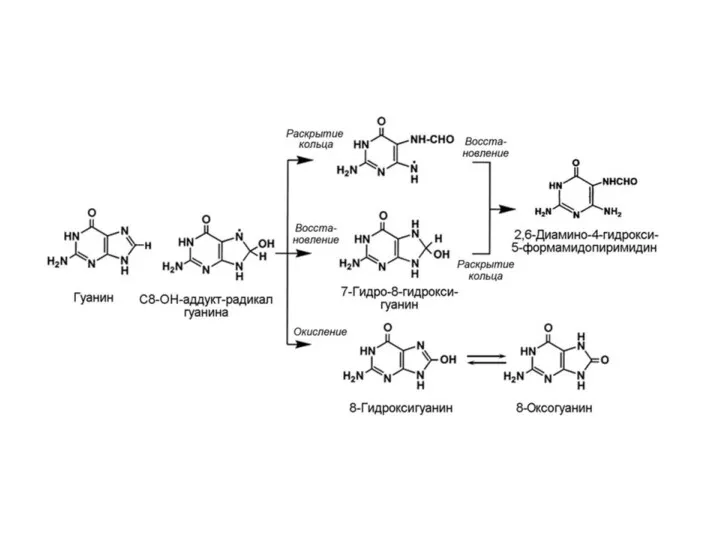
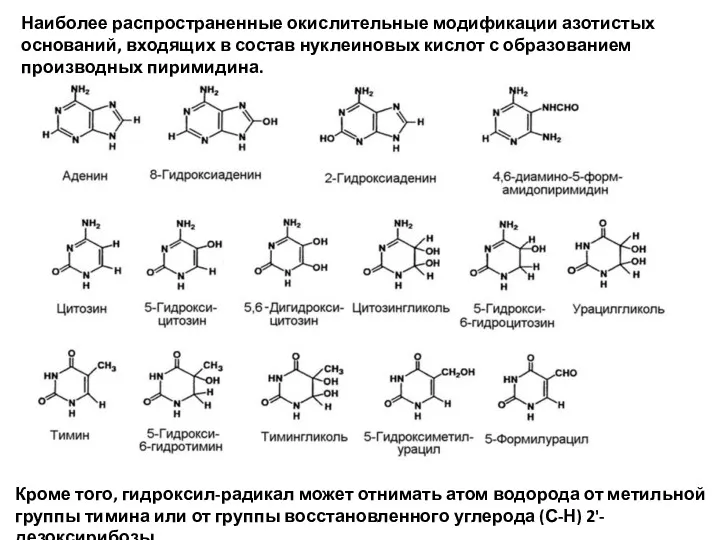
Наиболее распространенные окислительные модификации азотистых оснований, входящих в состав нуклеиновых кислот
Наиболее распространенные окислительные модификации азотистых оснований, входящих в состав нуклеиновых кислот

Идентифицировано приблизительно 20 окислительных повреждений азотистых оснований. У пуринов присоединение гидроксил-радикала
Идентифицировано приблизительно 20 окислительных повреждений азотистых оснований. У пуринов присоединение гидроксил-радикала

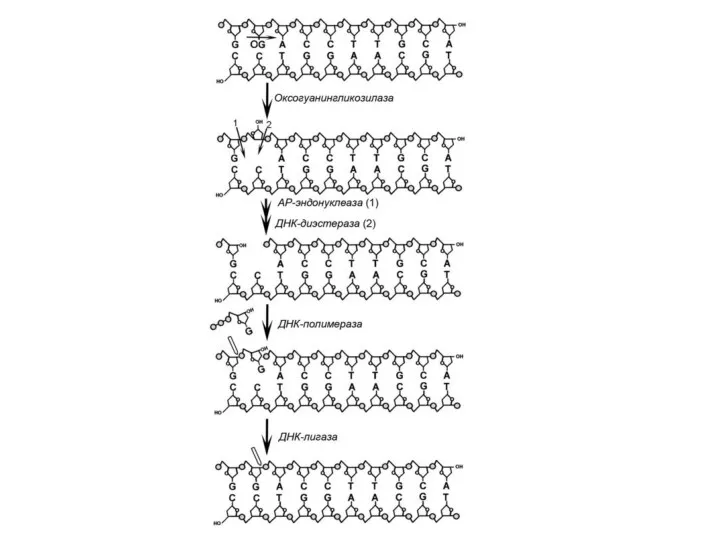
В клетках всех организмов существует многокомпонентная система репарации повреждений ДНК. Окисленные
В клетках всех организмов существует многокомпонентная система репарации повреждений ДНК. Окисленные

Липофусцин (lipofuscinum; от греч. lipo – ≪жир≫ и лат. fuscus –
Липофусцин (lipofuscinum; от греч. lipo – ≪жир≫ и лат. fuscus –

Липофусцин – гликопротеид, в состав которого входят:
- Жиры
- Аминокислоты
- Ферменты
- Флавиновые
Липофусцин – гликопротеид, в состав которого входят:
- Жиры
- Аминокислоты
- Ферменты
- Флавиновые
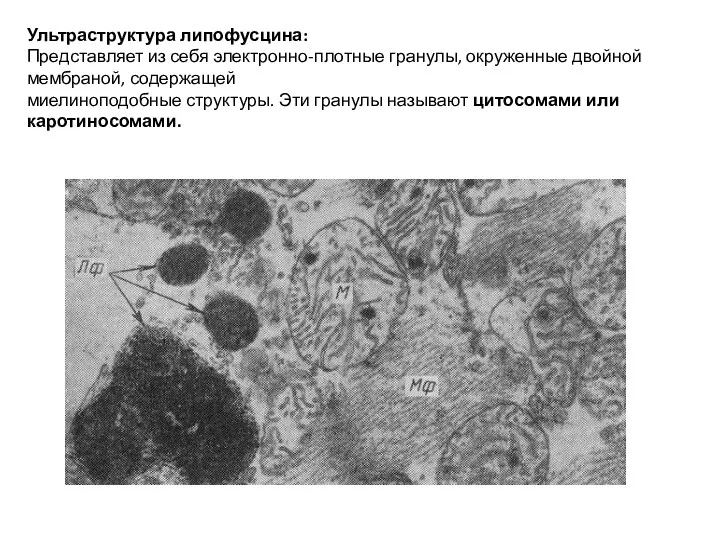
Ультраструктура липофусцина:
Представляет из себя электронно-плотные гранулы, окруженные двойной мембраной, содержащей
миелиноподобные структуры.
Ультраструктура липофусцина:
Представляет из себя электронно-плотные гранулы, окруженные двойной мембраной, содержащей
миелиноподобные структуры.
 Гнилостные бактерии
Гнилостные бактерии Необычные приветствия. Сквернословие
Необычные приветствия. Сквернословие Ракообразные. Классификация, строение, функции, размножение
Ракообразные. Классификация, строение, функции, размножение Покрытосеменные, или Цветковые
Покрытосеменные, или Цветковые Игра. Угадай животное
Игра. Угадай животное Cattle breeds
Cattle breeds Биологические науки
Биологические науки Большой (конечный) мозг
Большой (конечный) мозг Створення зелених насаджень
Створення зелених насаджень Репликация и транскрипция
Репликация и транскрипция Надцарство доядерные
Надцарство доядерные Эндокринная система
Эндокринная система Видоизменения побегов
Видоизменения побегов Биология – наука о живом мире. Общие свойства живых организмов.
Биология – наука о живом мире. Общие свойства живых организмов. Цитоплазматическая наследственность
Цитоплазматическая наследственность Ботанический сад во время блокады Ленинграда
Ботанический сад во время блокады Ленинграда Презентация по теме Постэмбриональное развитие
Презентация по теме Постэмбриональное развитие Класс насекомые. Стрекозы
Класс насекомые. Стрекозы Пришкольный участок. Уход за растениями
Пришкольный участок. Уход за растениями Квезаль - священная птица индейцев майя
Квезаль - священная птица индейцев майя Основы здорового питания собак и кошек
Основы здорового питания собак и кошек Биогенетический закон
Биогенетический закон Отряд приматы
Отряд приматы Развитие жизни в протерозойской и палеозойской эрах
Развитие жизни в протерозойской и палеозойской эрах Разведение кроликов
Разведение кроликов Органы и системы органов животных
Органы и системы органов животных Наследственность и изменчивость организмов
Наследственность и изменчивость организмов Кабинет биологии, фото
Кабинет биологии, фото