- Механизмы передачи сигнала: фермент-связывающие и фермент-содержащие рецепторы

Содержание
- 2. План занятия 1) Общие представления 2) Механизм действия рецепторов на примере а) липофильных гормонов (внутриклеточные рецепторы)
- 3. Медиатор, нейромедиатор, мессенджер (от англ. – messenger – посредник) – молекула, адресно передающая информацию от клетки
- 4. Сайт, место связывания, площадка узнавания – участок рецептор-ной молекулы либо фермента, специфично связывающийся с лигандом (субстратом)
- 5. Лигандом может быть нейромедиатор гормон ксенобиотик естественное вещество, биологического происхождения, не являющееся специфичным для данного вида
- 6. Определение понятия «клеточный рецептор» Клеточный рецептор — молекула (белок или гетерополисахарид) или функциональная группа молекул находящаяся
- 7. обладать высоким сродством (специфичностью) к лиганду ткани что определяет наличие или отсутствие данных функций в клетках
- 8. В плазматических мембранах про- и эукариотических клеток локализованы различные специализированные рецепторные системы, процесс функционирования которых включают
- 9. Полная передача межклеточного сигнала предполагает примерно следующую схему: 1) внешний агент (медиатор, стимул) взаимодействует с клеточным
- 10. Механизм действия дистантных липофильных гормонов Местом действия этих биорегуляторов являются ядра клеток-мишеней. В крови липофильные гормоны
- 11. Рецепторы липофильных гормонов принадлежат к группе редких белков. присутствуют в клетках-мишенях в количестве 10−3 – 10−4
- 12. 6) комплекс связывается с регуляторными участками генов, которые именуются гормон-респонсивные элементы (ГРЭ (HRE)). 7) ГРЭ –
- 13. Секреция гормона Связывание гормона с транспортными белками Транспорт гормона сквозь цитоплазматическую мембрану Образование комплекса гормон-рецептор Экспрессия
- 15. Рецепторы липофильных гормонов 1) во многом сходны, поскольку принадлежат к одному семей-ству белков; 2) образованы несколькими
- 16. Рецепторы липофильных гормонов
- 17. Рецептор стероидных гормонов Biophysical and Structural Characterization of Nuclear Hormone Receptors Mark Remec Pavlin College Scholars:
- 18. Механизм действия альдостерона
- 19. Механизм действия эстрадиола Белок Hsp90 – шаперон, помогает другим белкам правильно сворачиваться в трехмерные глобулы. Он
- 20. Взаимодействие липофильного гормона с внутриклеточным рецептором
- 21. Различия механизмов реализации эффектов липо- и гидрофильных гормонов
- 22. Механизм стимуляции функции клеток предстательной железы андрогенами
- 23. В семейство цинксодержащих белков входят рецепторы: стероидных гормонов, тироксина (и других тиреоидных гормонов), ретиноевой кислоты, вирусный
- 24. С помощью химического синтеза получают вещества, не идентичные гормонам, но обладающие свойством связываться с рецепторами. Например,
- 25. Механизм действия гидрофильных гормонов Большинство гидрофильных сигнальных веществ не спо-собны проходить через липофильную клеточную мембрану. Поэтому
- 26. Различают три типа рецепторов Рецепторы первого типа Ионные каналы Рецепторы третьего типа, сопряженные с ГТФ-связывающими белками
- 27. Рецепторы первого типа являются белками, имеющими одну трансмембранную полипептидную цепь это аллостерические ферменты, активный центр которых
- 28. 1) связывание сигнального вещества ведет к димеризации рецептора первого типа 2) это активирует фермент и фосфорилирование
- 29. Ионные каналы (рецепторы второго типа ) рецепторы второго типа являются олигомерными мембранными белками, образующими лиганд-активируемый ионный
- 30. Ацетилхолиновый рецептор в электровозбудимой мембране Структура рецептора Ацетилхолин Сайт связывания α-субъединицы Ворота Закрыты Открыты Функция рецептора
- 31. Рецепторы третьего типа, сопряженные с ГТФ-связывающими белками полипептидная цепь этих белков включает семь трансмембранных тяжей, проникающих
- 32. Связывание сигнального вещества с мембранным рецептором влечет за собой один из трех вариантов внутриклеточного ответа: рецепторные
- 33. Все три системы передачи сигнала взаимосвязаны Например, образование вторичного мессенджера цАМФ (сАМР) активирует протеинкиназу А [ПК-А
- 34. 5-HT серотониновый рецептор
- 35. Липофильный лиганд Гидрофильный лиганд Ионный канал Лиганд связанный с матриксом Рецепторы, сопряжённые с G-белком Рецептор клеточной
- 36. Преобразование сигнала G-белками G-белки переносят сигнал с рецептора третьего типа на белки-эффекторы. построены из трех субъединиц:
- 37. Рецептор, связанный с G-белком Рецептор Лиганд G-белок субъединицы
- 38. Передача сигнала с помощью рецептора третьего типа осуществляется поэтапно: 1) связывание сигнального вещества с рецептором третьего
- 39. Рецептор, связанный с G-белком Лиганд ГТФ ГТФ ГДФ ГДФ G-белок Экспрессия гена ФДЭ Активация протеинкиназы Инактивация
- 40. 5) медленный гидролиз связанного ГТФ до ГДФ переводит α-субъединицу в неактивное состояние и она вновь связывается
- 41. Вторичные мессенджеры Вторичные мессенджеры (посредники) – внутриклеточные вещества, передающие информацию от мембранного рецептора на исполняющие внутриклеточные
- 42. Циклический АМФ Биосинтез: нуклеотид цАМФ (3',5'-циклоаденозинмонофосфат, сАМP) синтезируется мембранными аденилатциклазами – семей-ством ферментов, катализирующих реакцию циклизации
- 43. Синтез циклического АМФ
- 44. Механизм действия цАМФ является аллостерическим эффектором протеинкиназ А (ПК-Α) и ионных каналов. В неактивном состоянии ПК-Α
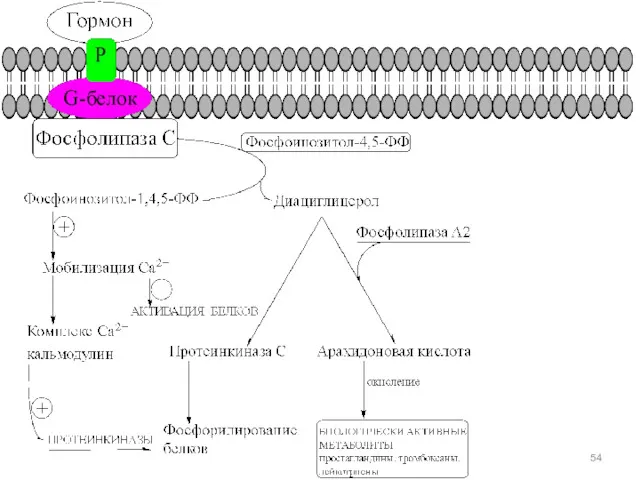
- 45. Кроме цАМФ функции вторичного мессенджера могутт выполнять цГМФ (cGMP), либо фосфоинозитолдифосфат, инозитолтрифосфат, диацилглицерол (ДАК), все соединения
- 46. Цепь реакций с участником внутриклеточного посредника цАМФ Стимулирующий внешний сигнал Ингибирующий внешний сигнал Форскалин Включение Колюшный
- 47. Цепь реакций с участником внутриклеточного посредника цАМФ Rs — возбуждающий рецептор, Ri — тормозный рецептор, Gs
- 48. Роль ионов кальция Концентрация Са2+ в цитоплазме нестимулированной клетки очень низка (10-100 нМ). Низкий уровень поддерживается
- 49. Канал может быть открыт деполяризацией мембран или действием сигнальных веществ, нейромедиаторов (глутамат и АТФ), вторичных мессенджеров
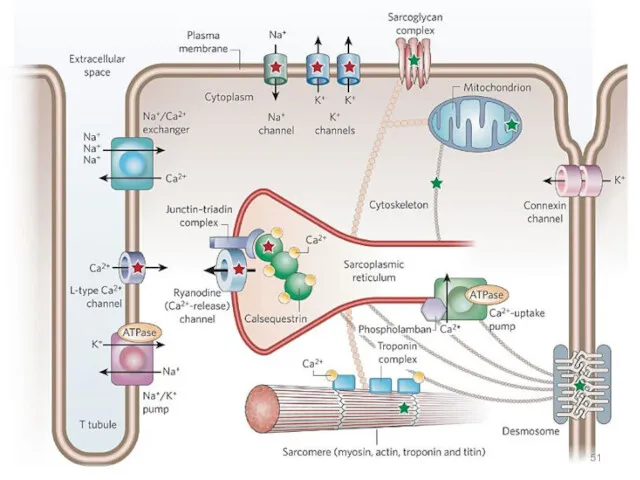
- 50. Действие кальция Опосредовано специальными Са2+-связывающими белками («кальциевыми сенсорами»): аннексин, кальмодулин и тропонин. Кальмодулин — сравнительно небольшой
- 52. Инозит-1,4,5-трифосфат и диацилглицерин Фосфолипаза С гидролизует фосфатидилинозит-4,5-дифосфат [ФИФ2 (PlnsP2)], что приводит к образованию двух вторичных мессенджеров:
- 53. Гормон Рецептор Фосфолипаза С Протеинкиназа С Цитоплазма ДАГ PTI Ca2+-канал Ca2+-канал IP3 открытие Плазматическая мембрана Гладкий
- 55. Рецепторы с собственной ферментативной активностью имеют внеклеточный лиганд-связывающий домен, трансмембранный участок и внутриклеточную часть, внутриклеточная часть
- 56. Рецепторы с собственной ферментативной активностью бывают: с тирозинкиназной активностью; с серин-треонин киназной активностью; связанные с тирозинкиназами;
- 57. 1. Рецепторы с тирозинкиназной активностью при связывании с сигнальной молекулой димеризуются, что включает их активность и
- 58. 3. Рецепторы, связанные с тирозинкиназами не являются ферментами, но тесно связаны с тирозинкиназами, находящимися в цитозоле,
- 59. Рецепторы цитокинов Цитокины — гидрофильные сигнальные вещества, действие которых опосредовано тирозинкиназными рецепторами на внешней стороне плазматической
- 60. Семейство цитокиновых рецепторов Семейство рецепторов интерлейкинов тип I Семейство рецепторов интерлейкинов тип II Рецептор фактора некроза
- 61. Цитокиновые рецепторы могут образовывать гетеродимеры за счет связывания со вспомогательными белками-переносчиками сигнала [БПС (STP)] или стимулировать
- 62. ПСАТ белки относятся к семейству БПС, имеющих в структуре SH2-домен, узнающий остатки фосфотирозина. Поэтому они обладают
- 63. Работа янус киназы Двуликий Янус EPO – рецептор эритропоэтина STAT белки – семейство факторов транскрипции эукариот,
- 64. Дополнительная информация В совокупности цитокины образуют регуляторную сеть (каскад цитокинов) с многофункциональным действием. Взаимоперекрывание между цитокинами
- 65. Сигнальные пути Иногда активация рецептора внешним стимулом сразу приводит к ответу клетки. Например, когда нейротрансмиттер ГАМК
- 66. Сигнальный путь Notch относительно короток. Активация этого рецептора позволяет протеазе расщепить его, часть белка может затем
- 67. Например, эпидермальный фактор роста, связывая свой рецептор, активирует его аутокаталитическое фосфорилирование. Фосфорилированный рецептор может связывать адапторный
- 68. Сеть цитокиновых взаимодействий Внимание! Это демонстрация. Не для попыток заучивания
- 70. Скачать презентацию

План занятия
1) Общие представления
2) Механизм действия рецепторов на примере
а)
План занятия
1) Общие представления
2) Механизм действия рецепторов на примере
а)

Медиатор, нейромедиатор, мессенджер (от англ. – messenger – посредник) – молекула,
Медиатор, нейромедиатор, мессенджер (от англ. – messenger – посредник) – молекула,

Сайт, место связывания, площадка узнавания – участок рецептор-ной молекулы либо фермента,
Сайт, место связывания, площадка узнавания – участок рецептор-ной молекулы либо фермента,
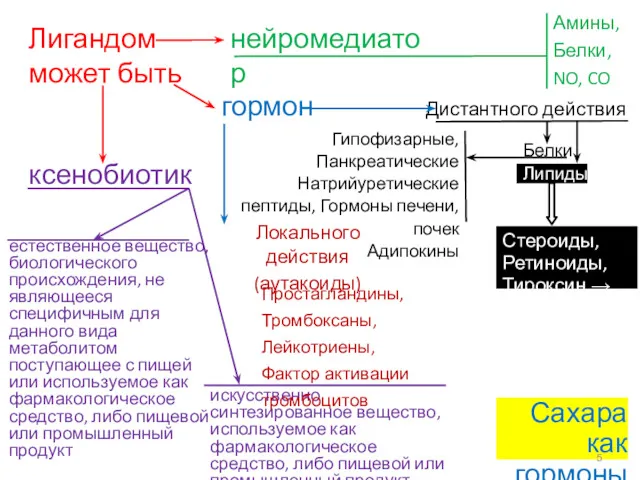
Лигандом может быть
нейромедиатор
гормон
ксенобиотик
естественное вещество, биологического происхождения, не являющееся специфичным для данного
Лигандом может быть
нейромедиатор
гормон
ксенобиотик
естественное вещество, биологического происхождения, не являющееся специфичным для данного

Определение понятия «клеточный рецептор»
Клеточный рецептор — молекула (белок или гетерополисахарид) или
Определение понятия «клеточный рецептор»
Клеточный рецептор — молекула (белок или гетерополисахарид) или

обладать высоким сродством (специфичностью) к
лиганду
ткани
что определяет наличие или отсутствие данных функций
обладать высоким сродством (специфичностью) к
лиганду
ткани
что определяет наличие или отсутствие данных функций

В плазматических мембранах про- и эукариотических клеток локализованы различные специализированные рецепторные
В плазматических мембранах про- и эукариотических клеток локализованы различные специализированные рецепторные

Полная передача межклеточного сигнала предполагает примерно следующую схему:
1) внешний агент (медиатор,
Полная передача межклеточного сигнала предполагает примерно следующую схему:
1) внешний агент (медиатор,
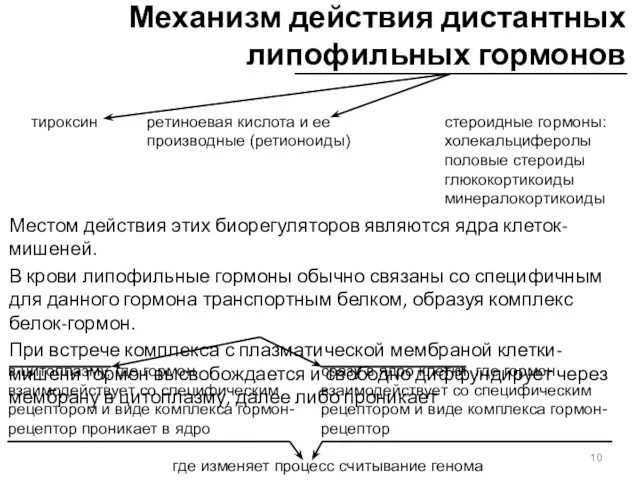
Механизм действия дистантных липофильных гормонов
Местом действия этих биорегуляторов являются ядра клеток-мишеней.
В
Механизм действия дистантных липофильных гормонов
Местом действия этих биорегуляторов являются ядра клеток-мишеней.
В

Рецепторы липофильных гормонов
принадлежат к группе редких белков.
присутствуют в клетках-мишенях
Рецепторы липофильных гормонов
принадлежат к группе редких белков.
присутствуют в клетках-мишенях

6) комплекс связывается с регуляторными участками генов, которые именуются гормон-респонсивные элементы
6) комплекс связывается с регуляторными участками генов, которые именуются гормон-респонсивные элементы
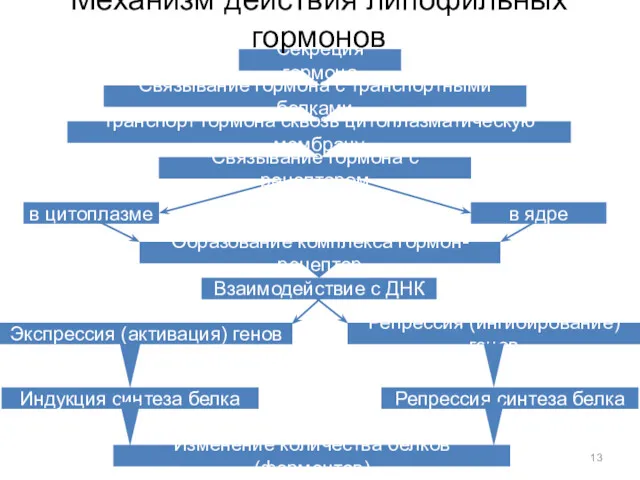
Секреция гормона
Связывание гормона с транспортными белками
Транспорт гормона сквозь цитоплазматическую мембрану
Образование комплекса
Секреция гормона
Связывание гормона с транспортными белками
Транспорт гормона сквозь цитоплазматическую мембрану
Образование комплекса
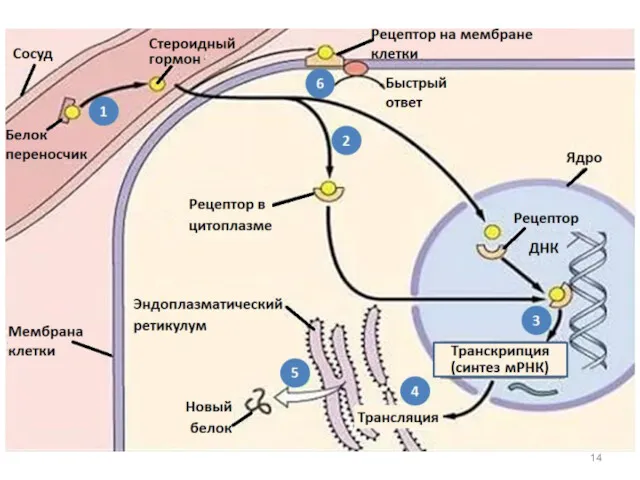

Рецепторы липофильных гормонов
1) во многом сходны, поскольку принадлежат к одному семей-ству
Рецепторы липофильных гормонов
1) во многом сходны, поскольку принадлежат к одному семей-ству
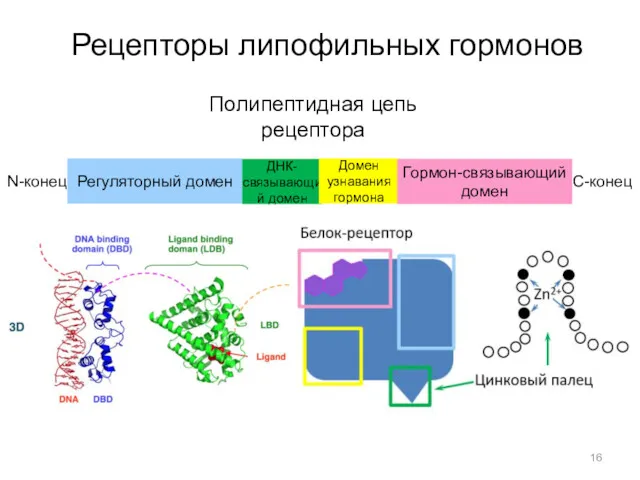
Рецепторы липофильных гормонов
Рецепторы липофильных гормонов
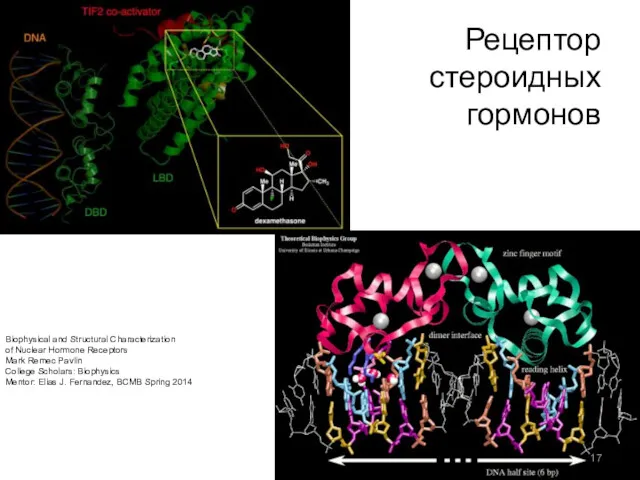
Рецептор стероидных гормонов
Biophysical and Structural Characterization
of Nuclear Hormone Receptors
Mark Remec
Рецептор стероидных гормонов
Biophysical and Structural Characterization
of Nuclear Hormone Receptors
Mark Remec
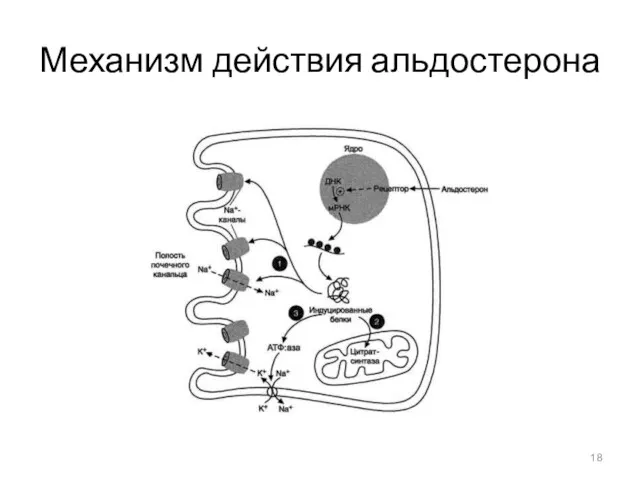
Механизм действия альдостерона
Механизм действия альдостерона
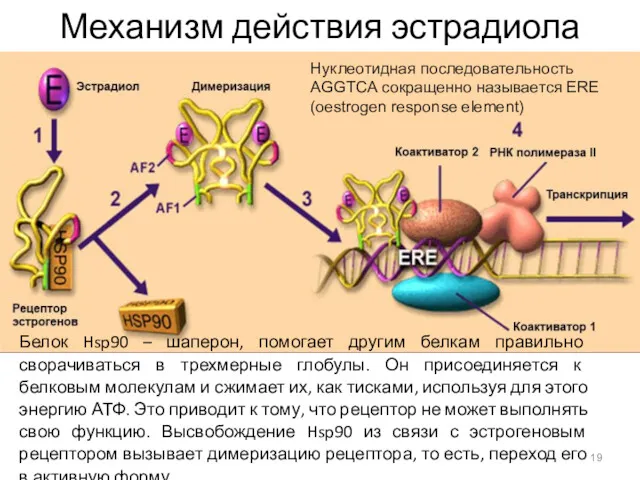
Механизм действия эстрадиола
Белок Hsp90 – шаперон, помогает другим белкам правильно сворачиваться
Механизм действия эстрадиола
Белок Hsp90 – шаперон, помогает другим белкам правильно сворачиваться
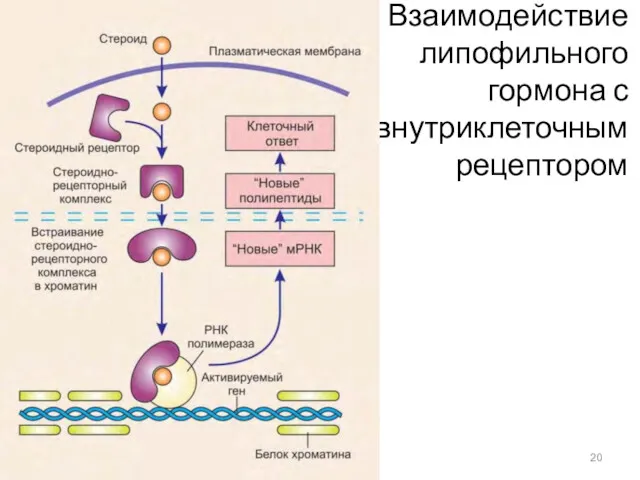
Взаимодействие липофильного гормона с внутриклеточным рецептором
Взаимодействие липофильного гормона с внутриклеточным рецептором
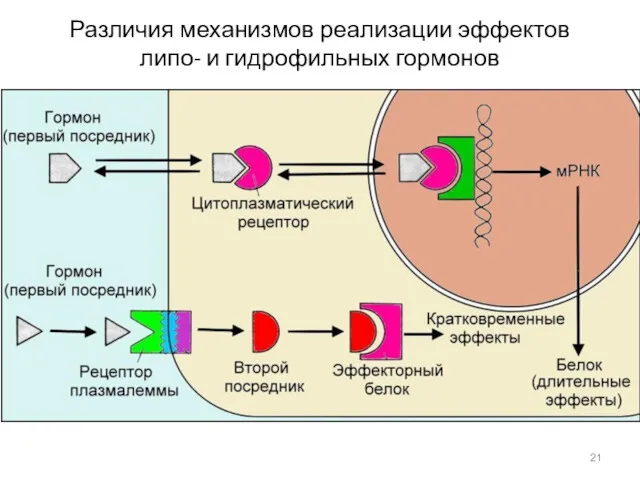
Различия механизмов реализации эффектов липо- и гидрофильных гормонов
Различия механизмов реализации эффектов липо- и гидрофильных гормонов
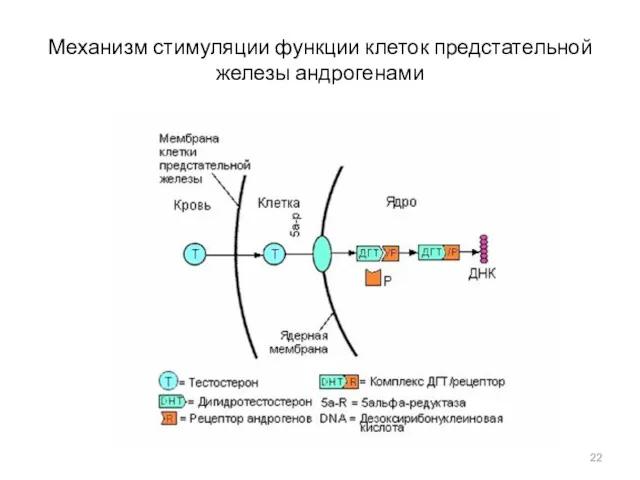
Механизм стимуляции функции клеток предстательной железы андрогенами
Механизм стимуляции функции клеток предстательной железы андрогенами

В семейство цинксодержащих белков входят рецепторы:
стероидных гормонов,
тироксина (и других
В семейство цинксодержащих белков входят рецепторы:
стероидных гормонов,
тироксина (и других

С помощью химического синтеза получают вещества, не идентичные гормонам, но обладающие
С помощью химического синтеза получают вещества, не идентичные гормонам, но обладающие

Механизм действия гидрофильных гормонов
Большинство гидрофильных сигнальных веществ не спо-собны проходить
Механизм действия гидрофильных гормонов
Большинство гидрофильных сигнальных веществ не спо-собны проходить

Различают три типа рецепторов
Рецепторы первого типа
Ионные каналы
Рецепторы третьего
Различают три типа рецепторов
Рецепторы первого типа
Ионные каналы
Рецепторы третьего

Рецепторы первого типа
являются белками, имеющими одну трансмембранную полипептидную цепь
это аллостерические
Рецепторы первого типа
являются белками, имеющими одну трансмембранную полипептидную цепь
это аллостерические

1) связывание сигнального вещества ведет к димеризации рецептора первого типа
2) это
1) связывание сигнального вещества ведет к димеризации рецептора первого типа
2) это

Ионные каналы
(рецепторы второго типа )
рецепторы второго типа являются олигомерными мембранными
Ионные каналы
(рецепторы второго типа )
рецепторы второго типа являются олигомерными мембранными
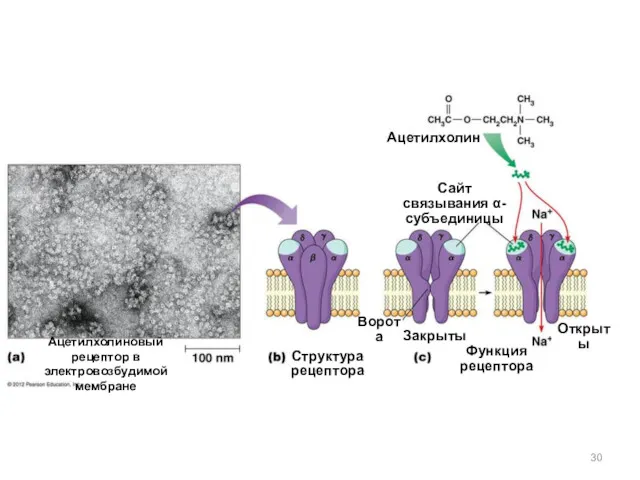
Ацетилхолиновый рецептор в электровозбудимой мембране
Структура
рецептора
Ацетилхолин
Сайт связывания α-субъединицы
Ворота
Закрыты
Открыты
Функция рецептора
Ацетилхолиновый рецептор в электровозбудимой мембране
Структура
рецептора
Ацетилхолин
Сайт связывания α-субъединицы
Ворота
Закрыты
Открыты
Функция рецептора

Рецепторы третьего типа, сопряженные с ГТФ-связывающими белками
полипептидная цепь этих белков
Рецепторы третьего типа, сопряженные с ГТФ-связывающими белками
полипептидная цепь этих белков

Связывание сигнального вещества с мембранным рецептором влечет за собой один из
Связывание сигнального вещества с мембранным рецептором влечет за собой один из

Все три системы передачи сигнала взаимосвязаны
Например, образование вторичного мессенджера цАМФ (сАМР)
Все три системы передачи сигнала взаимосвязаны
Например, образование вторичного мессенджера цАМФ (сАМР)
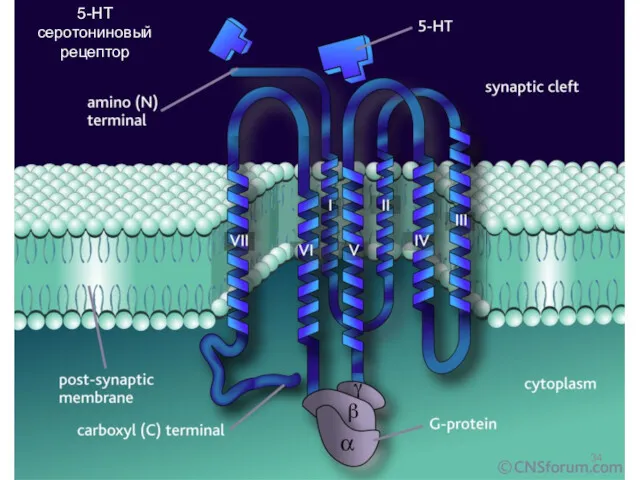
5-HT серотониновый рецептор
5-HT серотониновый рецептор
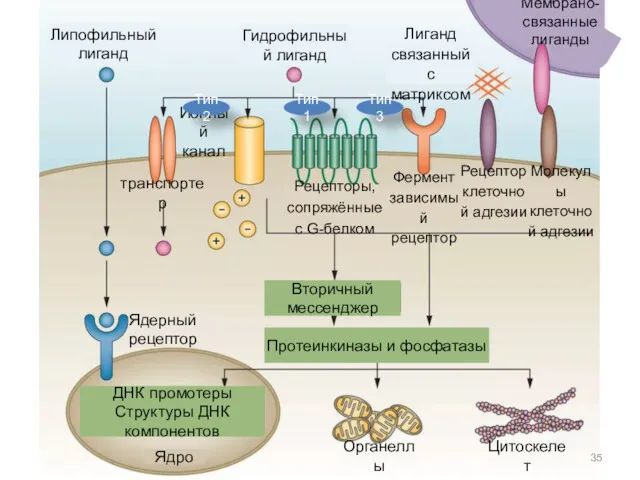
Липофильный лиганд
Гидрофильный лиганд
Ионный канал
Лиганд
связанный с матриксом
Рецепторы, сопряжённые с G-белком
Рецептор клеточной
Липофильный лиганд
Гидрофильный лиганд
Ионный канал
Лиганд
связанный с матриксом
Рецепторы, сопряжённые с G-белком
Рецептор клеточной

Преобразование сигнала G-белками
G-белки переносят сигнал с рецептора третьего типа на
Преобразование сигнала G-белками
G-белки переносят сигнал с рецептора третьего типа на
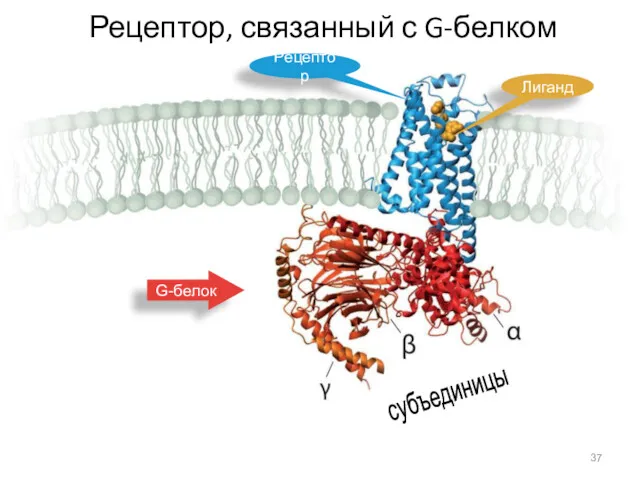
Рецептор, связанный с G-белком
Рецептор
Лиганд
G-белок
субъединицы
Рецептор, связанный с G-белком
Рецептор
Лиганд
G-белок
субъединицы

Передача сигнала с помощью рецептора третьего типа осуществляется поэтапно:
1) связывание сигнального
Передача сигнала с помощью рецептора третьего типа осуществляется поэтапно:
1) связывание сигнального
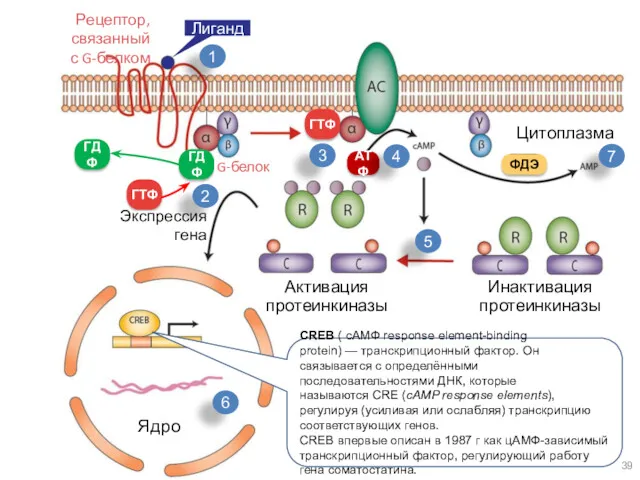
Рецептор, связанный
с G-белком
Лиганд
ГТФ
ГТФ
ГДФ
ГДФ
G-белок
Экспрессия гена
ФДЭ
Активация протеинкиназы
Инактивация протеинкиназы
Цитоплазма
Ядро
CREB ( сAMФ response element-binding
Рецептор, связанный
с G-белком
Лиганд
ГТФ
ГТФ
ГДФ
ГДФ
G-белок
Экспрессия гена
ФДЭ
Активация протеинкиназы
Инактивация протеинкиназы
Цитоплазма
Ядро
CREB ( сAMФ response element-binding

5) медленный гидролиз связанного ГТФ до ГДФ переводит α-субъединицу в неактивное
5) медленный гидролиз связанного ГТФ до ГДФ переводит α-субъединицу в неактивное

Вторичные мессенджеры
Вторичные мессенджеры (посредники) – внутриклеточные вещества, передающие информацию от мембранного
Вторичные мессенджеры
Вторичные мессенджеры (посредники) – внутриклеточные вещества, передающие информацию от мембранного

Циклический АМФ
Биосинтез: нуклеотид цАМФ (3',5'-циклоаденозинмонофосфат, сАМP) синтезируется мембранными аденилатциклазами –
Циклический АМФ
Биосинтез: нуклеотид цАМФ (3',5'-циклоаденозинмонофосфат, сАМP) синтезируется мембранными аденилатциклазами –
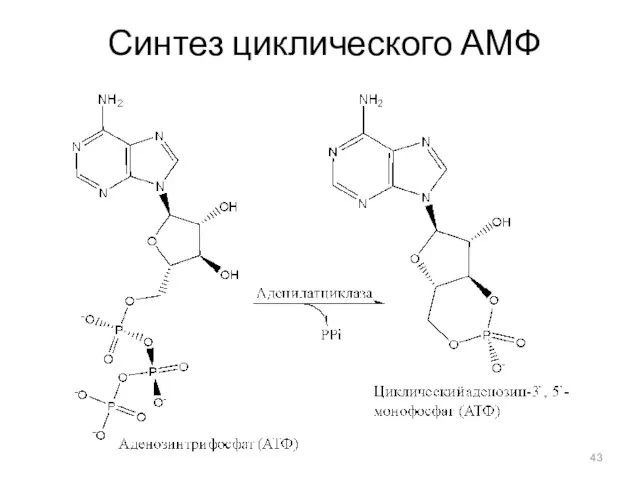
Синтез циклического АМФ
Синтез циклического АМФ

Механизм действия
цАМФ является аллостерическим эффектором протеинкиназ А (ПК-Α) и ионных каналов.
В
Механизм действия
цАМФ является аллостерическим эффектором протеинкиназ А (ПК-Α) и ионных каналов.
В
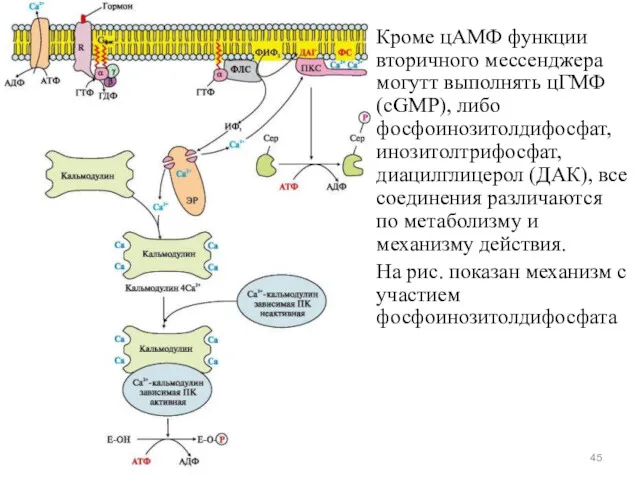
Кроме цАМФ функции вторичного мессенджера могутт выполнять цГМФ (cGMP), либо фосфоинозитолдифосфат,
Кроме цАМФ функции вторичного мессенджера могутт выполнять цГМФ (cGMP), либо фосфоинозитолдифосфат,
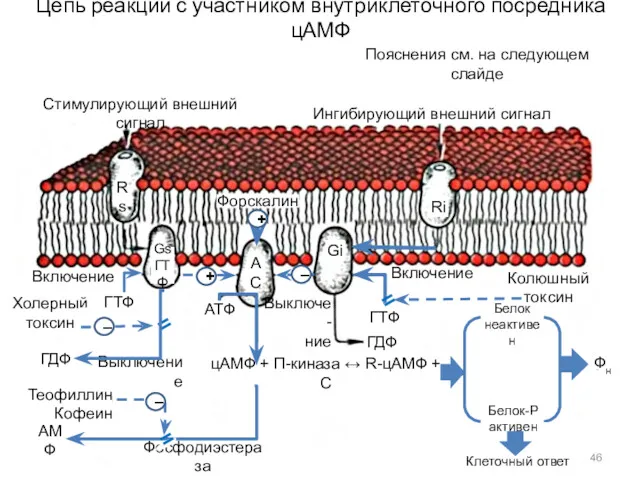
Цепь реакций с участником внутриклеточного посредника цАМФ
Стимулирующий
Цепь реакций с участником внутриклеточного посредника цАМФ
Стимулирующий

Цепь реакций с участником внутриклеточного посредника цАМФ
Rs — возбуждающий рецептор,
Ri —
Цепь реакций с участником внутриклеточного посредника цАМФ
Rs — возбуждающий рецептор,
Ri —

Роль ионов кальция
Концентрация Са2+ в цитоплазме нестимулированной клетки очень низка
Роль ионов кальция
Концентрация Са2+ в цитоплазме нестимулированной клетки очень низка
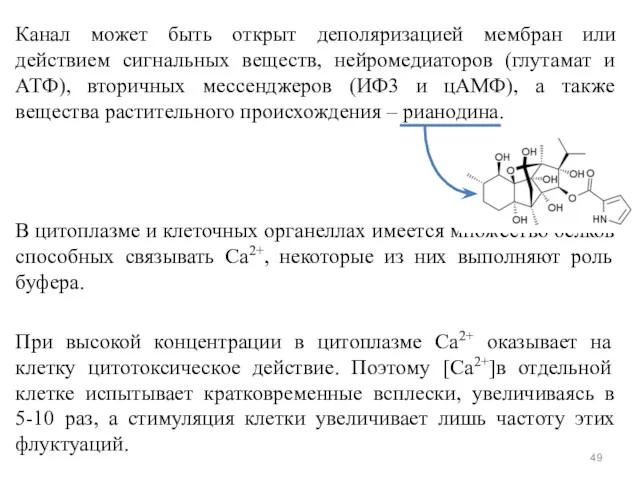
Канал может быть открыт деполяризацией мембран или действием сигнальных веществ, нейромедиаторов
Канал может быть открыт деполяризацией мембран или действием сигнальных веществ, нейромедиаторов

Действие кальция
Опосредовано специальными Са2+-связывающими белками («кальциевыми сенсорами»): аннексин, кальмодулин и тропонин.
Кальмодулин
Действие кальция
Опосредовано специальными Са2+-связывающими белками («кальциевыми сенсорами»): аннексин, кальмодулин и тропонин.
Кальмодулин
![Инозит-1,4,5-трифосфат и диацилглицерин Фосфолипаза С гидролизует фосфатидилинозит-4,5-дифосфат [ФИФ2 (PlnsP2)], что](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/157733/slide-51.jpg)
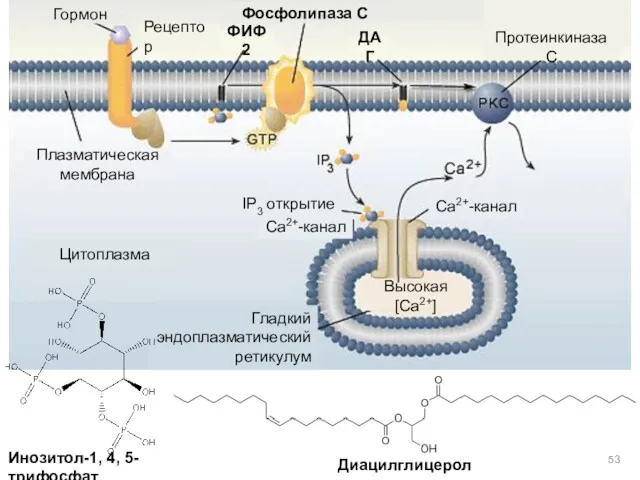
Инозит-1,4,5-трифосфат и диацилглицерин
Фосфолипаза С гидролизует фосфатидилинозит-4,5-дифосфат [ФИФ2 (PlnsP2)], что приводит
Инозит-1,4,5-трифосфат и диацилглицерин
Фосфолипаза С гидролизует фосфатидилинозит-4,5-дифосфат [ФИФ2 (PlnsP2)], что приводит
Гормон
Рецептор
Фосфолипаза С
Протеинкиназа С
Цитоплазма
ДАГ
PTI
Ca2+-канал
Ca2+-канал
IP3 открытие
Плазматическая мембрана
Гладкий
эндоплазматический
ретикулум
Высокая [Ca2+]
ФИФ2
Диацилглицерол
Инозитол-1, 4, 5-трифосфат
Гормон
Рецептор
Фосфолипаза С
Протеинкиназа С
Цитоплазма
ДАГ
PTI
Ca2+-канал
Ca2+-канал
IP3 открытие
Плазматическая мембрана
Гладкий
эндоплазматический
ретикулум
Высокая [Ca2+]
ФИФ2
Диацилглицерол
Инозитол-1, 4, 5-трифосфат

Рецепторы с собственной ферментативной активностью
имеют
внеклеточный лиганд-связывающий домен,
трансмембранный участок и
внутриклеточную
Рецепторы с собственной ферментативной активностью
имеют
внеклеточный лиганд-связывающий домен,
трансмембранный участок и
внутриклеточную

Рецепторы с собственной ферментативной активностью бывают:
с тирозинкиназной активностью;
с серин-треонин киназной активностью;
Рецепторы с собственной ферментативной активностью бывают:
с тирозинкиназной активностью;
с серин-треонин киназной активностью;

1. Рецепторы с тирозинкиназной активностью при связывании с сигнальной молекулой димеризуются, что
1. Рецепторы с тирозинкиназной активностью при связывании с сигнальной молекулой димеризуются, что

3. Рецепторы, связанные с тирозинкиназами не являются ферментами, но тесно связаны с
3. Рецепторы, связанные с тирозинкиназами не являются ферментами, но тесно связаны с

Рецепторы цитокинов
Цитокины — гидрофильные сигнальные вещества, действие которых опосредовано тирозинкиназными рецепторами
Рецепторы цитокинов
Цитокины — гидрофильные сигнальные вещества, действие которых опосредовано тирозинкиназными рецепторами
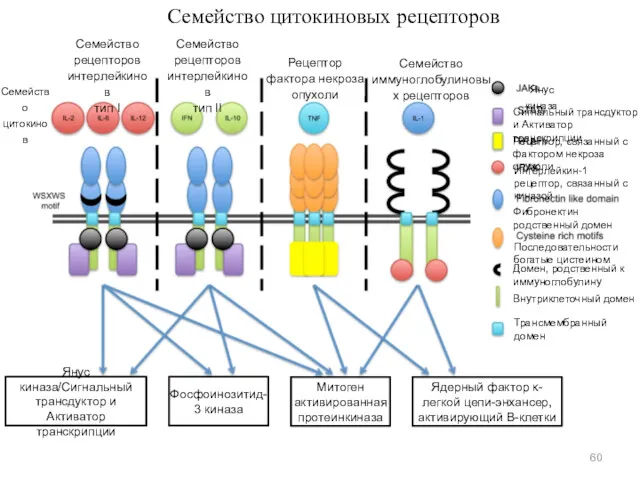
Семейство цитокиновых рецепторов
Семейство
рецепторов
интерлейкинов
тип I
Семейство
рецепторов
интерлейкинов
тип II
Рецептор фактора некроза
Семейство цитокиновых рецепторов
Семейство
рецепторов
интерлейкинов
тип I
Семейство
рецепторов
интерлейкинов
тип II
Рецептор фактора некроза

Цитокиновые рецепторы могут образовывать гетеродимеры за счет связывания со вспомогательными белками-переносчиками
Цитокиновые рецепторы могут образовывать гетеродимеры за счет связывания со вспомогательными белками-переносчиками

ПСАТ белки относятся к семейству БПС, имеющих в структуре SH2-домен, узнающий
ПСАТ белки относятся к семейству БПС, имеющих в структуре SH2-домен, узнающий
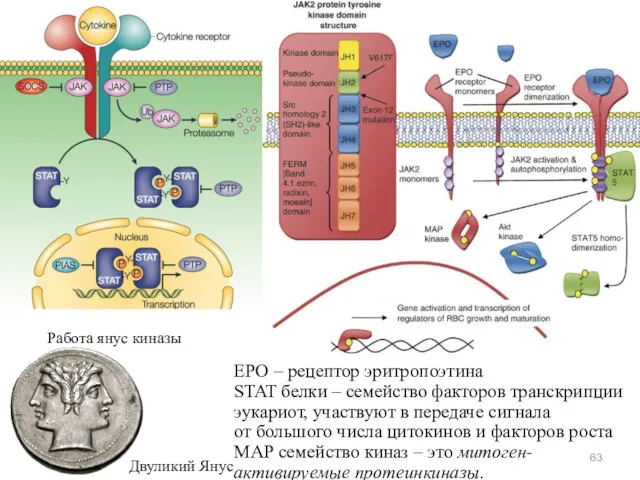
Работа янус киназы
Двуликий Янус
EPO – рецептор эритропоэтина
STAT белки – семейство факторов
Работа янус киназы
Двуликий Янус
EPO – рецептор эритропоэтина
STAT белки – семейство факторов
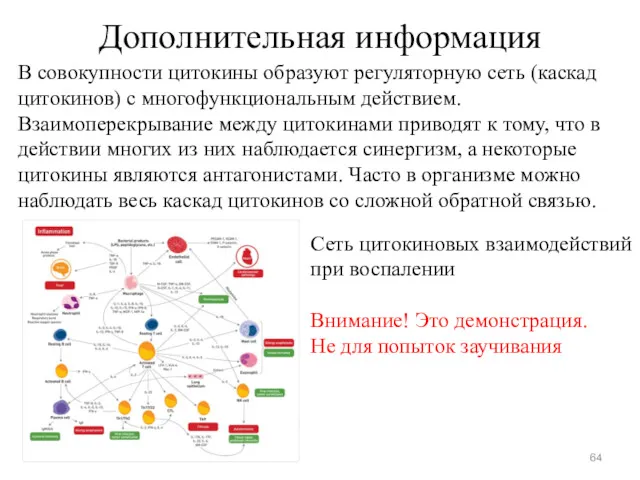
Дополнительная информация
В совокупности цитокины образуют регуляторную сеть (каскад цитокинов) с многофункциональным
Дополнительная информация
В совокупности цитокины образуют регуляторную сеть (каскад цитокинов) с многофункциональным

Сигнальные пути
Иногда активация рецептора внешним стимулом сразу приводит к ответу клетки.
Сигнальные пути
Иногда активация рецептора внешним стимулом сразу приводит к ответу клетки.

Сигнальный путь Notch относительно короток. Активация этого рецептора позволяет протеазе расщепить
Сигнальный путь Notch относительно короток. Активация этого рецептора позволяет протеазе расщепить

Например, эпидермальный фактор роста, связывая свой рецептор, активирует его аутокаталитическое фосфорилирование.
Фосфорилированный
Например, эпидермальный фактор роста, связывая свой рецептор, активирует его аутокаталитическое фосфорилирование.
Фосфорилированный
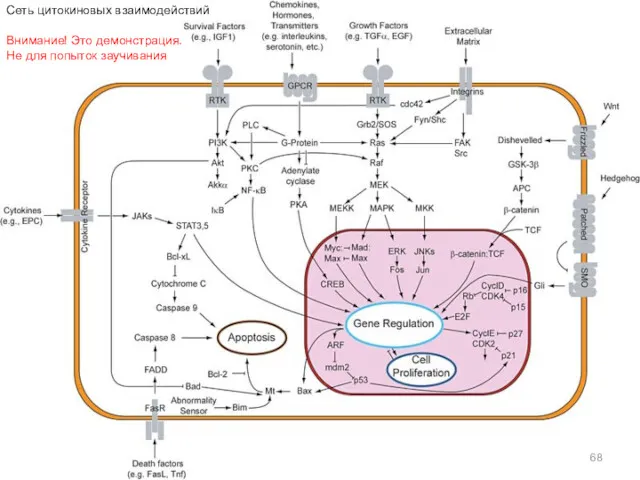
Сеть цитокиновых взаимодействий
Внимание! Это демонстрация.
Не для попыток заучивания
Сеть цитокиновых взаимодействий
Внимание! Это демонстрация.
Не для попыток заучивания
 Система опоры и движения. Скелет
Система опоры и движения. Скелет Строение цветка. Соцветие. Значение оплодотворения
Строение цветка. Соцветие. Значение оплодотворения Флуор. Його роль в організмі людини
Флуор. Його роль в організмі людини Животные лесов умеренной зоны
Животные лесов умеренной зоны Наследственность и изменчивость организмов
Наследственность и изменчивость организмов Бұғылар, бітеумүйізділер (лат. Cervidae)
Бұғылар, бітеумүйізділер (лат. Cervidae) Ферменты. Гормоны
Ферменты. Гормоны Корень. Урок биологии 6 класс
Корень. Урок биологии 6 класс Городские птицы
Городские птицы презентация к уроку Ветер
презентация к уроку Ветер методическая разработка урока на тему: клеточное строение листа для 6 класса
методическая разработка урока на тему: клеточное строение листа для 6 класса Презентация Первая помощь при кровотечениях.
Презентация Первая помощь при кровотечениях. Биологические ритмы и сон
Биологические ритмы и сон Основы здорового питания собак и кошек
Основы здорового питания собак и кошек Интерактивное пособие по теме Мышцы
Интерактивное пособие по теме Мышцы Сезонные изменения в жизни организмов
Сезонные изменения в жизни организмов Строение волос
Строение волос Клеточная теория
Клеточная теория Ядовитые растения и животные
Ядовитые растения и животные Ферменты. Функции белков
Ферменты. Функции белков Введення в анатомію та фізіологію
Введення в анатомію та фізіологію Система живой природы
Система живой природы Спинномозговые нервы
Спинномозговые нервы Красная книга. Коала
Красная книга. Коала Тип Кольчатые черви
Тип Кольчатые черви Органические вещества в составе почвы
Органические вещества в составе почвы Класс Пресмыкающиеся, или Рептилии
Класс Пресмыкающиеся, или Рептилии Лекарственные растения желчегонного действия
Лекарственные растения желчегонного действия