- Механізм скорочення м’язового волокна. (Лекція 6)

Содержание
- 2. Електромеханічний зв’язк Потенціал дії Деполяризація мембрани Т-трубочок Деполяризація мембрани СР Вихід йонів кальцію Скорочення м’яза
- 3. Рис.1. Коливання концентрації йонів кальцію під час генерації ПД і активації скорочення м’язу. 1. Генерація потенціалу
- 4. Характеристики деполяризації м’язових волокон 1) при тривалій деполяризації м′язового волокна після початкового скорочення відбувається мимовільне його
- 5. Особливості активації скорочення у тонічних м′язових волокнах Волокна за нормальних умов не здатні до генерації ПД.
- 6. Рис. 2. Будова м′язового волокна (І) і міофібрили (ІІ): А – анізотропні диски, І – ізотропні
- 7. Рис. 3. Схема ковзання ниток актину і міозину в ході скорочення: а) І – розслаблена міофібрила;
- 8. Механізми, що лежать в основі скорочення м′язового волокна можна об’єднати у дві групи: взаємне переміщення (ковзання)
- 9. Рис. 4. Механізм утворення акто-міозинового комплексу. А- зображення актинової і міозинової ниток у продольному перетині; Б
- 10. А – модель механізму руху: міозинова нитка з поперечними містками, що прикріплені до сусідніх актинових ниток;
- 11. Послідовність процесу скорочення м’яза згідно теорії ковзання
- 12. Рис. 6. Передача ПД через нервово-мязовий синапс
- 13. Рис. 7. Відкриття місця зв’язування на актині
- 14. Рис. 8. Вихід кальцію із СР і приєднання до актину
- 15. Рис. 9. Приєднання міозинової головки до актину
- 16. Рис. 10. Поворот міозинової головки
- 17. Рис. 11. Роз’єднання акто-міозинового містка
- 18. Рис. 12. Відновлення початкового положення міозинової головки
- 19. Рис. 13. Повернення кальцію у СР
- 20. Рис. 14. Розміщення актинових та міозинових ниток Теорія вкручування
- 21. Енергетичне забезпечення м'язового скорочення. АТФ – є безпосереднім джерелом для м'язового скорочення. АТФ при цьому виконує
- 22. Ресинтез АТФ в анаеробних умовах відбувається за рахунок енергії: повільного дефосфорилювання креатинфосфату на креатин і фосфорну
- 23. Ресинтез АТФ в аеробних умовах Аеробні процеси відбуваються за участю кисню і супроводжуються окисленням ліпідів і
- 24. Рис. 15. Передача на робочий механізм м′язового волокна енергії, що вивільняється при окисних процесах.
- 26. Скачать презентацию

Електромеханічний зв’язк
Потенціал дії
Деполяризація мембрани Т-трубочок
Деполяризація мембрани СР
Вихід йонів кальцію
Скорочення
Електромеханічний зв’язк
Потенціал дії
Деполяризація мембрани Т-трубочок
Деполяризація мембрани СР
Вихід йонів кальцію
Скорочення
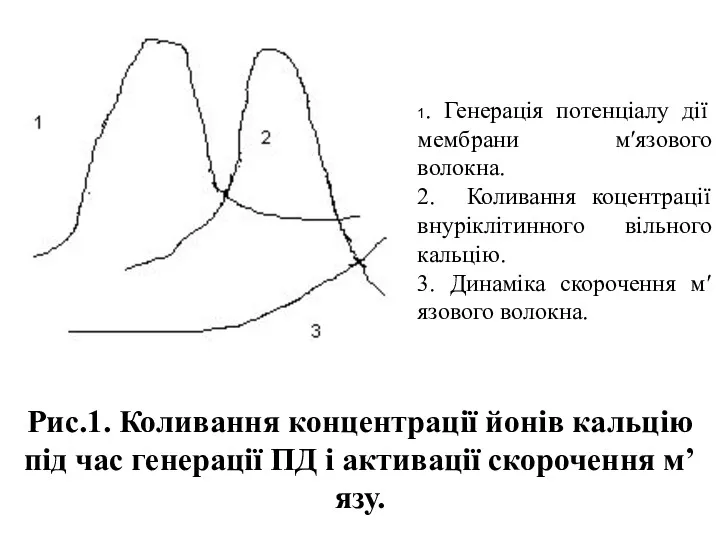
Рис.1. Коливання концентрації йонів кальцію під час генерації ПД і активації
Рис.1. Коливання концентрації йонів кальцію під час генерації ПД і активації

Характеристики деполяризації м’язових волокон
1) при тривалій деполяризації м′язового волокна після
Характеристики деполяризації м’язових волокон
1) при тривалій деполяризації м′язового волокна після

Особливості активації скорочення у тонічних м′язових волокнах
Волокна за нормальних умов не
Особливості активації скорочення у тонічних м′язових волокнах
Волокна за нормальних умов не
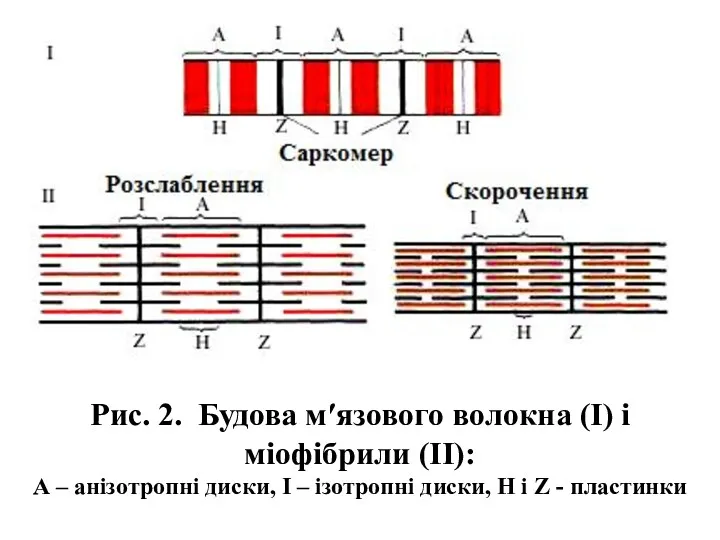
Рис. 2. Будова м′язового волокна (І) і міофібрили (ІІ):
А – анізотропні
Рис. 2. Будова м′язового волокна (І) і міофібрили (ІІ): А – анізотропні
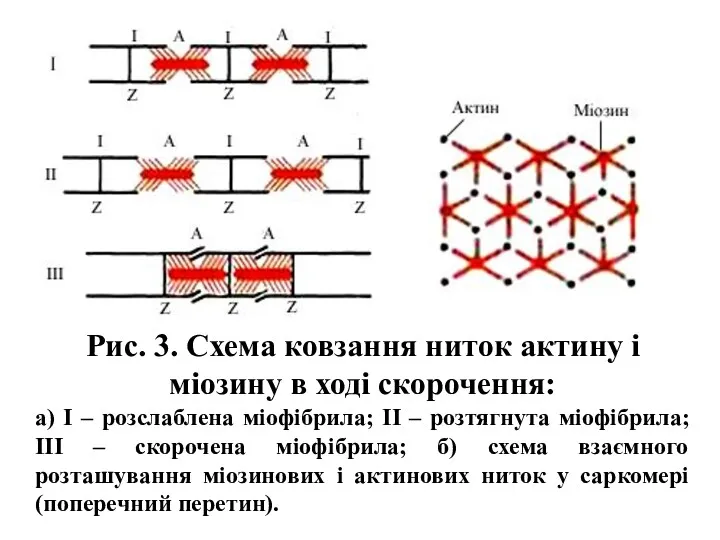
Рис. 3. Схема ковзання ниток актину і міозину в ході скорочення:
Рис. 3. Схема ковзання ниток актину і міозину в ході скорочення:

Механізми, що лежать в основі скорочення м′язового волокна можна об’єднати у
Механізми, що лежать в основі скорочення м′язового волокна можна об’єднати у
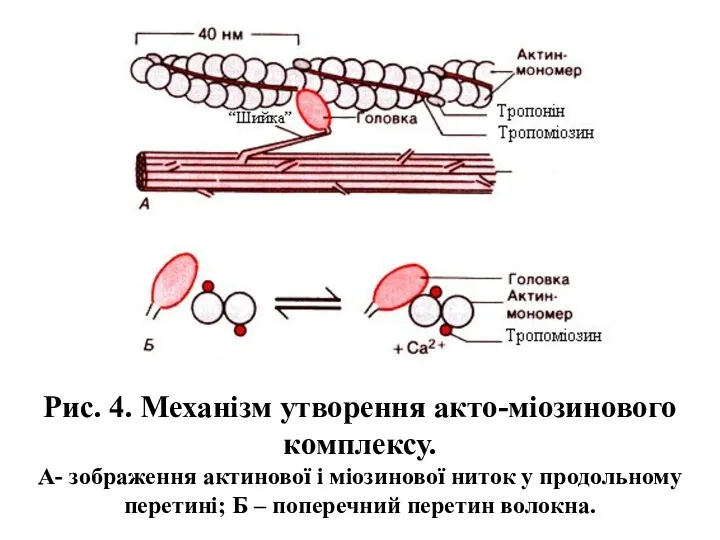
Рис. 4. Механізм утворення акто-міозинового комплексу.
А- зображення актинової і міозинової ниток
Рис. 4. Механізм утворення акто-міозинового комплексу. А- зображення актинової і міозинової ниток
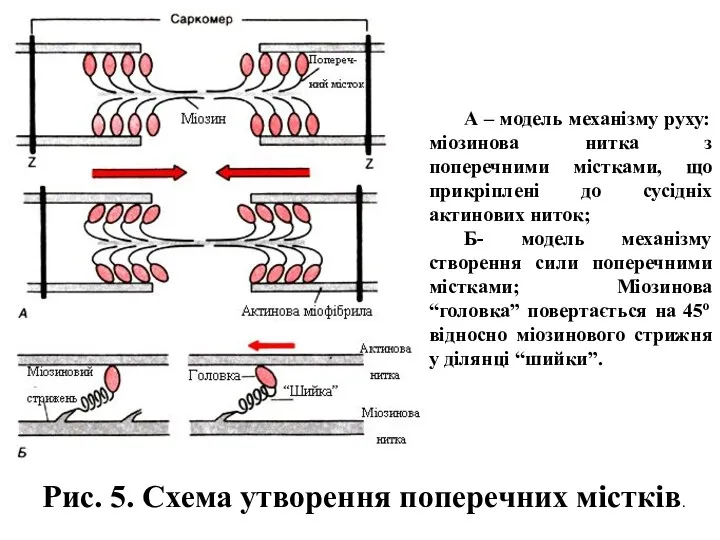
А – модель механізму руху: міозинова нитка з поперечними містками, що
А – модель механізму руху: міозинова нитка з поперечними містками, що

Послідовність процесу скорочення м’яза згідно теорії ковзання
Послідовність процесу скорочення м’яза згідно теорії ковзання
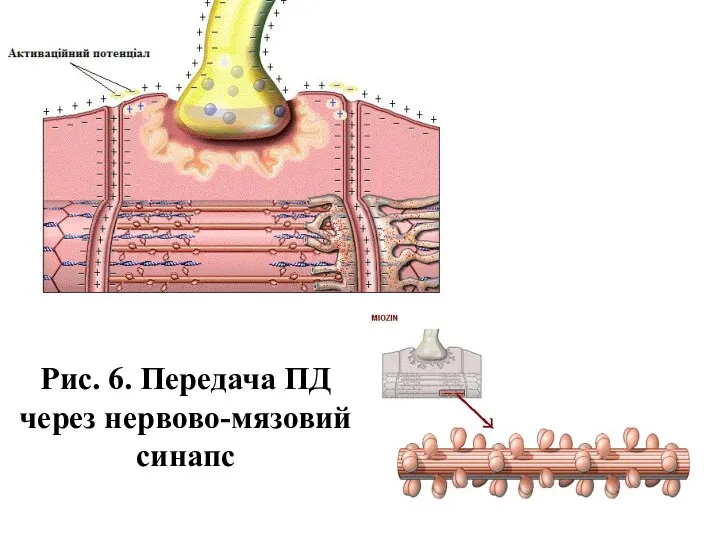
Рис. 6. Передача ПД через нервово-мязовий синапс
Рис. 6. Передача ПД через нервово-мязовий синапс
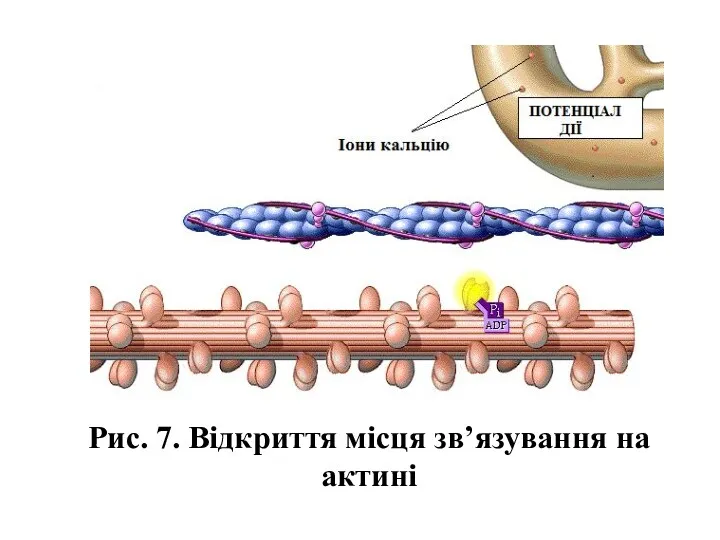
Рис. 7. Відкриття місця зв’язування на актині
Рис. 7. Відкриття місця зв’язування на актині
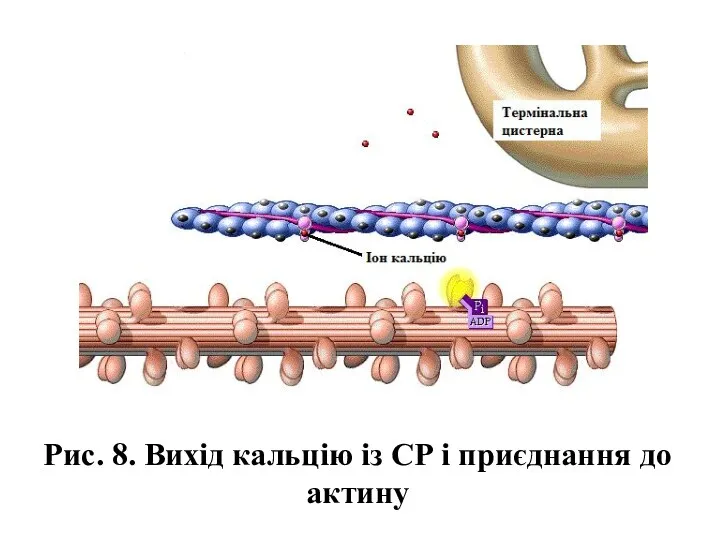
Рис. 8. Вихід кальцію із СР і приєднання до актину
Рис. 8. Вихід кальцію із СР і приєднання до актину
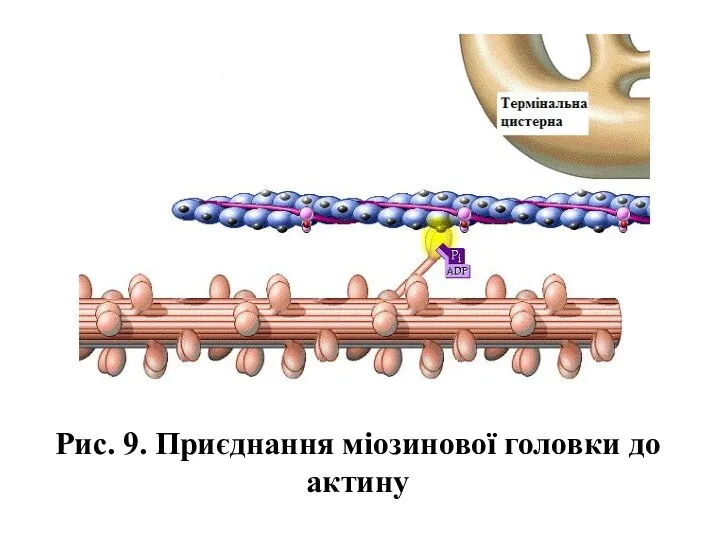
Рис. 9. Приєднання міозинової головки до актину
Рис. 9. Приєднання міозинової головки до актину

Рис. 10. Поворот міозинової головки
Рис. 10. Поворот міозинової головки
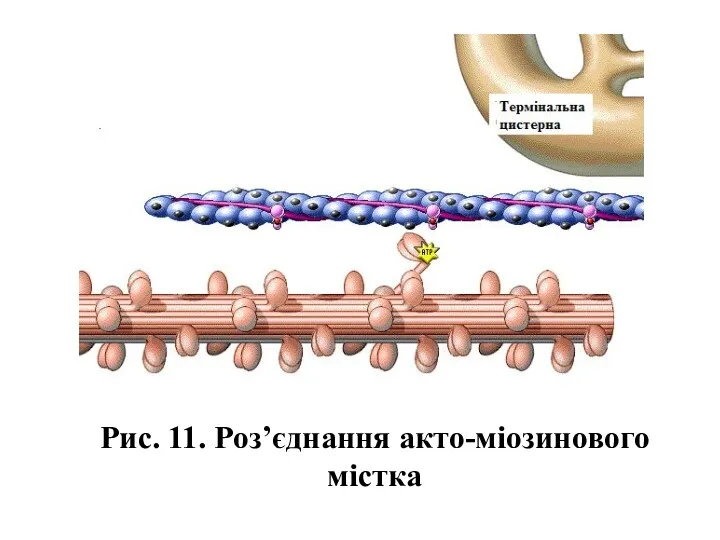
Рис. 11. Роз’єднання акто-міозинового містка
Рис. 11. Роз’єднання акто-міозинового містка
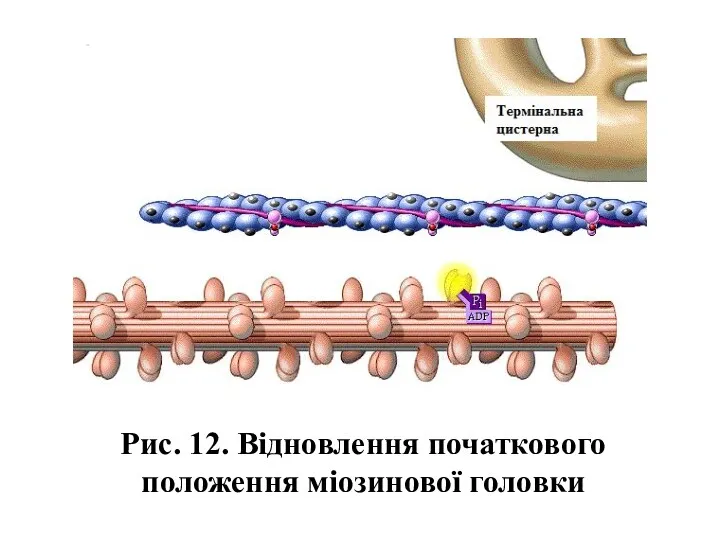
Рис. 12. Відновлення початкового положення міозинової головки
Рис. 12. Відновлення початкового положення міозинової головки
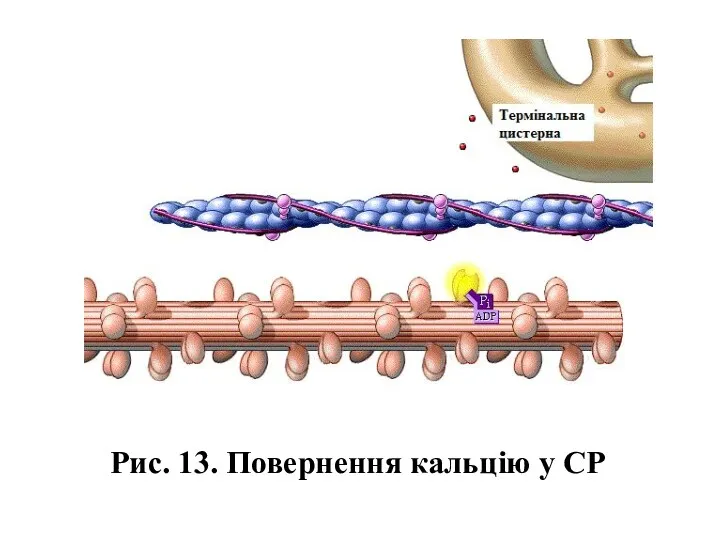
Рис. 13. Повернення кальцію у СР
Рис. 13. Повернення кальцію у СР
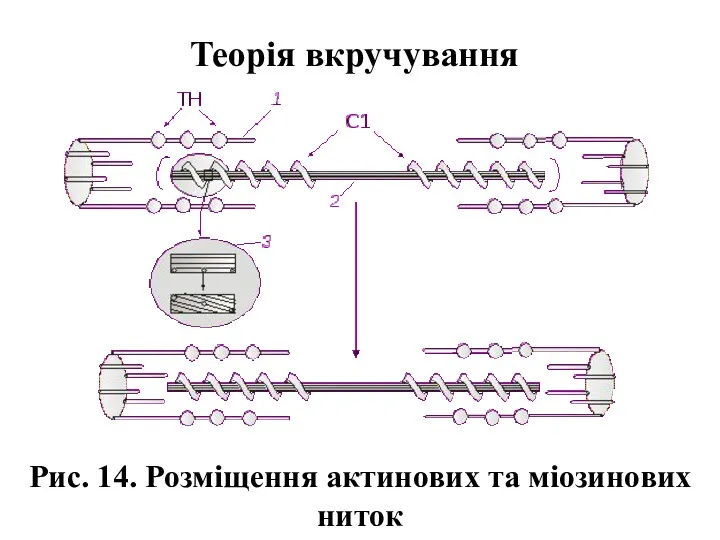
Рис. 14. Розміщення актинових та міозинових ниток
Теорія вкручування
Рис. 14. Розміщення актинових та міозинових ниток
Теорія вкручування

Енергетичне забезпечення м'язового скорочення.
АТФ – є безпосереднім джерелом для м'язового
Енергетичне забезпечення м'язового скорочення.
АТФ – є безпосереднім джерелом для м'язового

Ресинтез АТФ в анаеробних умовах відбувається за рахунок енергії:
повільного дефосфорилювання
Ресинтез АТФ в анаеробних умовах відбувається за рахунок енергії:
повільного дефосфорилювання

Ресинтез АТФ в аеробних умовах
Аеробні процеси відбуваються за участю кисню і
Ресинтез АТФ в аеробних умовах
Аеробні процеси відбуваються за участю кисню і
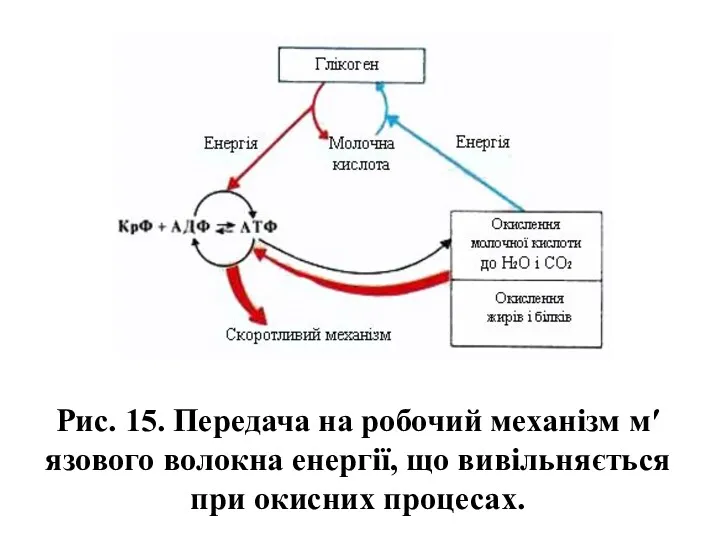
Рис. 15. Передача на робочий механізм м′язового волокна енергії, що вивільняється
Рис. 15. Передача на робочий механізм м′язового волокна енергії, що вивільняється
 Самые необычные животные
Самые необычные животные Путешествие по материкам. Животный мир Евразии
Путешествие по материкам. Животный мир Евразии Многообразие и происхождение пресмыкающихся
Многообразие и происхождение пресмыкающихся Слюнные железы
Слюнные железы Жизнь диких животных зимой
Жизнь диких животных зимой Морфологическая характеристика лекарственных и ядовитых растений
Морфологическая характеристика лекарственных и ядовитых растений Движение. 6 класс
Движение. 6 класс Мозжечок (cerebellum). Сагиттальный разрез головного мозга
Мозжечок (cerebellum). Сагиттальный разрез головного мозга Тихоходки. Что? Как? Почему?
Тихоходки. Что? Как? Почему? Проект Судогда - река, город, жизнь...
Проект Судогда - река, город, жизнь... Интересное о растениях
Интересное о растениях Класс Ракообразные
Класс Ракообразные Строение клетки. Растительная и животная клетки. (9 класс)
Строение клетки. Растительная и животная клетки. (9 класс) презентация Интеллектуальный марафон
презентация Интеллектуальный марафон Виды корней и типы корневых систем (7 класс)
Виды корней и типы корневых систем (7 класс) Химические элементы и неорганические вещества клетки
Химические элементы и неорганические вещества клетки Размножение растений семенами. 5 класс
Размножение растений семенами. 5 класс Антропогенез. Этапы эволюции человека
Антропогенез. Этапы эволюции человека Физиология высшей нервной деятельности. Формирование поведения в онтогенезе
Физиология высшей нервной деятельности. Формирование поведения в онтогенезе Вода, її роль у життєдіяльності організмів
Вода, її роль у життєдіяльності організмів Питание клетки
Питание клетки Мифы и легенды о животных
Мифы и легенды о животных Лекарственные растения Томской области
Лекарственные растения Томской области Животные Красной Книги. Белый медведь
Животные Красной Книги. Белый медведь Класс рыбы: Хрящевые, Костные
Класс рыбы: Хрящевые, Костные Биологические мембраны. Транспорт веществ через мембраны
Биологические мембраны. Транспорт веществ через мембраны Сцепленное наследование генов
Сцепленное наследование генов Отряд Рукокрылые
Отряд Рукокрылые