- Многообразие внутриклеточных каскадов в клетке

Содержание
- 2. Передача сигнала липофильными гормонами
- 4. Передача сигнала посредством внутриклеточных рецепторов
- 5. Механизм действия стероидных гормонов (СГ) 1.СГ→СГ+белок-переносчик→по кровотоку к клетке-мишени → диссоциация комплекса → диффузия СГ внутрь
- 7. Механизм регуляции экспрессии генов посредством стероидных и тиреоидных гормонов, ретиноидной кислоты и витамина Д
- 9. Вторичные посредники Пути образования и проведение сигнала (цАМФ, цГМФ, NO, липидные мессенджеры)
- 10. Вторичные посредники – это низкомолекулярные вещества, небелковой природы, образуются и действуют внутри клеток, и обеспечивают передачу
- 11. Вторичные мессенджеры: циклические нуклеотиды (цАМФ, цГМФ), мембранные липиды и их производные (ИФ3, ДАГ), ионы (Са2+), оксид
- 13. Ключевые этапы передачи сигнала посредством ВП являются общими для регуляторных систем: агонист→рецептор→эффекторный белок→вторичный посредник→модулируемый белок→физиологический эффект
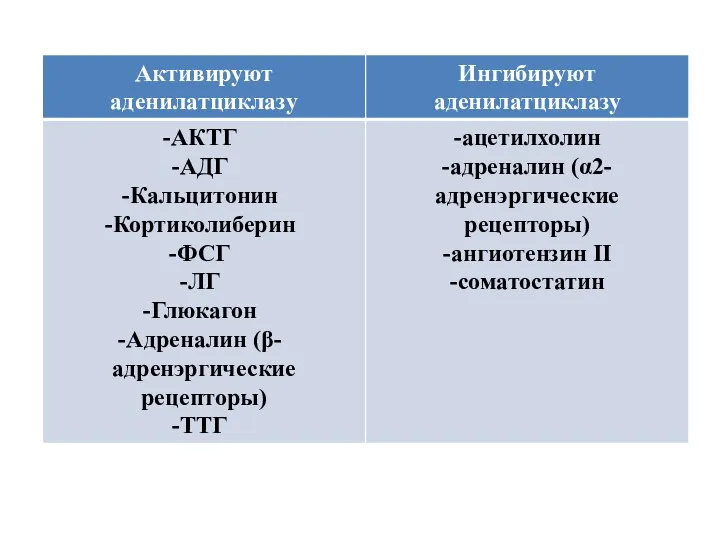
- 14. Пути проведения метаболического и пролиферативного сигнала рецепторами, сопряженными с G-белками G-белки Gs-белки Gi-белки Gq-белки Gs, Gi,
- 15. Существует 2 пути, с помощью которых GPCRs запускают образование вторичных посредников: сАМР – путь: активация GPCRs
- 16. Циклический АМФ (цАМФ) - важнейший ВП, это вещество «влияет на все – от памяти до кончиков
- 17. Образование цАМФ
- 18. Характеристика аденилатциклазы (АЦ) Интегральный белок плазматической мембраны Гликопротеин, М – 110-180 кДа Полипептидная цепь содержит 12
- 21. Упрощенная схема активации АЦ вследствие связывания гормона (адреналин, глюкагон) с рецепторм, сопряженным с G-белком
- 25. Передача сигнала через обонятельные рецепторы
- 28. Образование вторичного посредника цГМФ катализируется гуанилатциклазами
- 29. Образование цГМФ
- 30. Гуанилатциклазы (ГЦ) 1. ГЦ катализирует образование вторичного посредника цГМФ из ГТФ. 2. В клетке имеется 2
- 31. Механизм действия цГМФ
- 32. Регуляция активности мембранной и цитозольной ГЦ
- 34. Структура мембранной гуанилатциклазы
- 35. Структура растворимой ГЦ
- 36. Схема активации растворимой гуанилатциклазы оксидом азота
- 38. 1. В неактивированном состоянии гем гуанилатциклазы связан координационно с четырьмя атомами азота центрального кольца гема, а
- 39. NO и цГМФ-зависимые сигнальные пути
- 40. Схема активации зрительного каскада и регуляция цГМФ-активируемых натриевых каналов
- 41. цГМФ и гуанилатциклазы (Нельсон, Кокс, 2011)
- 42. NO как вторичный посредник. Биосинтез оксида азота и структура нейрональной NOS FMN, ВH4, гем NOS:NADFH, FAD
- 43. Строение NO-синтазы
- 44. Активность NOS регулируется : Фосфорилированием - ПКА - ПКС - Са/кальмодулин-зависимой киназой Дефосфорилированием - фосфатазой 1
- 45. NO запускает различные downstream пути и регулирует: вазодилятацию нейротрансмиссию макрофагальную цитотоксичность релаксацию гладкомышечных клеток ЖКТ бронходилятацию
- 46. Роль NO в работе возбуждающего нейрона
- 47. Липидные вторичные посредники образуются путем расщепления эфирных связей фосфолипидов фосфолипазами
- 48. Характеристика фосфолипаз (ФЛ) ФЛ - гидролазы, катализирующие катаболизм глицеро-фосфолипидов. Различают: 1) секреторные ФЛ (панкреа-тический сок); 2)
- 51. Роль фосфолипазы А2 в синтезе эйкозаноидов
- 52. Мембранные липиды и их производные: инозитол-1,4,5-трифосфат и 1,2-диацилглицерол
- 54. Синтез производных фосфатидилинозитола в плазматической мембране, гидролиз ФИФ2 фосфолипазой С ФИ-4-киназа → ФИ → ФИ-4-фосфат (ФИФ)
- 55. Характеристика фосфолипазы С 1.Известно 3 класса ФЛС: ФЛСβ, ФЛСγ, ФЛСδ, которые включают 16 изоформ. 2.ФЛСβ активируется
- 58. Механизм связывания с мембраной и активация ФЛС 1.Начальное связывание ФЛС с мембраной происходит через домен РН
- 63. Са2+ -единственный вторичный посредник, функционирующий во всех типах живых клеток
- 64. Са 2+ - ключевой вторичный посредник Различают 3 состояния Са2+ в клетках нескелетообра-зующих тканей: 1.Са2+, локализованный
- 65. Особенности Са 2+ как вторичного посредника 1. Са 2+ - неметаболизируемый (стабильный) катион и его внутриклеточной
- 66. Внутриклеточные Са2+-связывающие белки определяют Са2-опосредованные сигналы Выделяют 3 группы Са2+-связывающих белков: 1.ЕF-hand-белки (цитоплазма, ядро)- содержат EFH-фраг-мент
- 67. Са2+ - связывающие белки
- 76. Источники перекиси водорода в клетке: НАДФН-оксидаза Электрон-транспортная цепь митохондрий Электрон-транспортная цепь микросом Ксантиноксидоредуктаза (КОР) Супероксиддисмутаза Пути
- 77. Образование перекиси водорода НАДФН-оксидазой
- 78. Строение флавоцитохрома b558 НАДФН → ФАД → ФАДН• → гем(внутр) → гем(внеш) → О2 → О2‾•
- 79. Механизм активации NOX1-3 на плазматической мембране: лиганд→РТП→ФЛСγ→ПКС→фосфорилирование цитозольных белков-регуляторов→сборка НАДФН-оксидазного комплекса→продукция О2‾• и Н2О2 лиганд→РТП→Ras-белок→ПК→фосфорилирование цитозольных
- 80. Электрон-транспортная цепь митохондрий (5-6% АФК)
- 81. Продукция супероксида и перекиси водорода в митохондриях
- 82. Образование О2‾• и Н2О2 в системе микросомального окисления (75% АФК) Главной функцией монооксигеназ является детоксикация ксенобиотиков
- 83. Ксантиноксидоредуктаза – источник перекиси водорода в клетке Ксантиноксидоредуктаза представлена двумя изоформами: ксантиндегидрогеназой (КД) и ксантиноксидазой (КО).
- 84. Схема катаболизма пуринов, катализируемого ксантиноксидоредуктазой. Ксантиноксидаза – источник перекиси водорода
- 85. Схема функционирования ксантиноксидоредуктазы
- 86. Супероксиддисмутазы (СОД) – суперсемейство ферментов, относящихся к классу оксидоредуктаз и катализирующих реакцию дисмутации супероксидного анион-радикала с
- 87. Структура различных изоферментов СОД
- 88. Mn-СОД и АФК- сигналинг
- 89. Элиминация перекиси водорода в клетке осуществляется ферментативным путем: 1.Каталаза – гемсодержащий внутриклеточный фермент (тетрамер): 2Н2О2 →
- 90. Предполагаемый механизм действия перекиси водорода и ее мишени в клетке: тирозиновые фосфатазы (ингибирование), тирозиновые киназы (активация)
- 91. Дополнительные слайды
- 100. Скачать презентацию

Передача сигнала липофильными гормонами
Передача сигнала липофильными гормонами
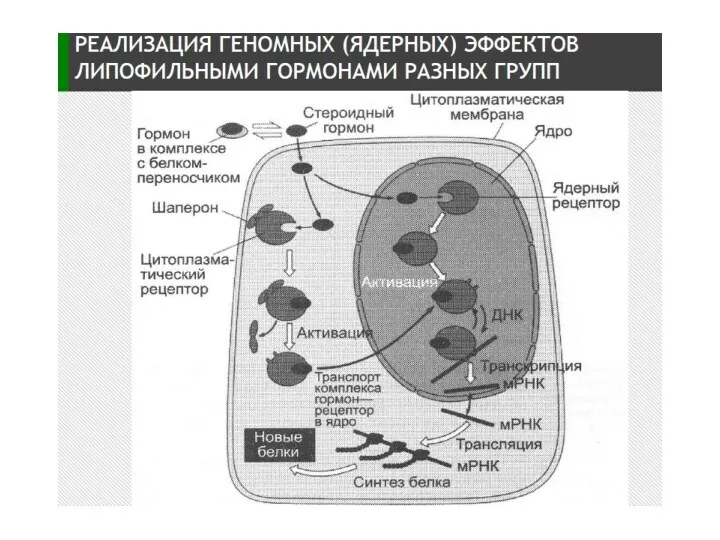
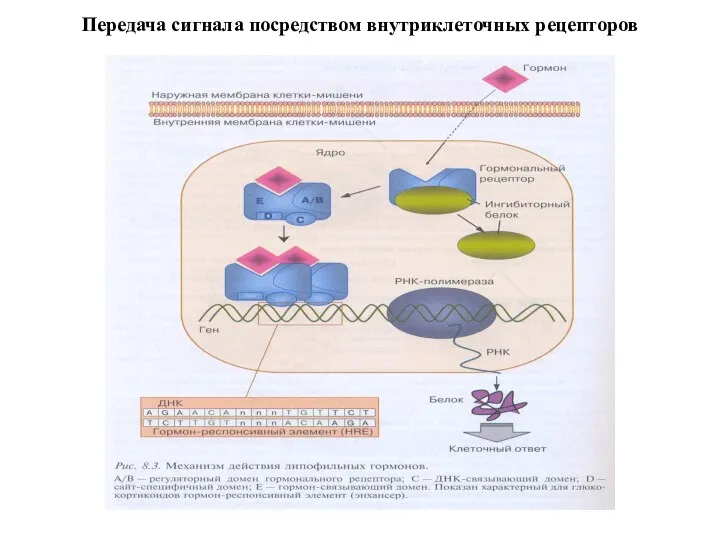
Передача сигнала посредством внутриклеточных рецепторов
Передача сигнала посредством внутриклеточных рецепторов

Механизм действия стероидных гормонов (СГ)
1.СГ→СГ+белок-переносчик→по кровотоку к клетке-мишени → диссоциация комплекса
Механизм действия стероидных гормонов (СГ)
1.СГ→СГ+белок-переносчик→по кровотоку к клетке-мишени → диссоциация комплекса
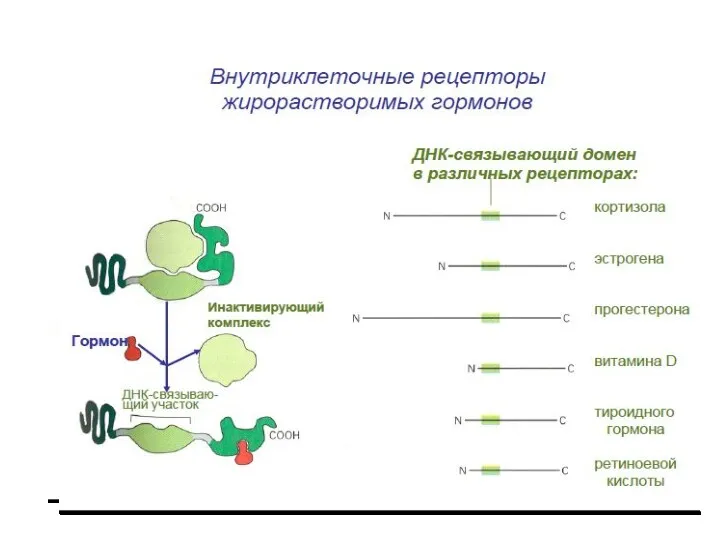
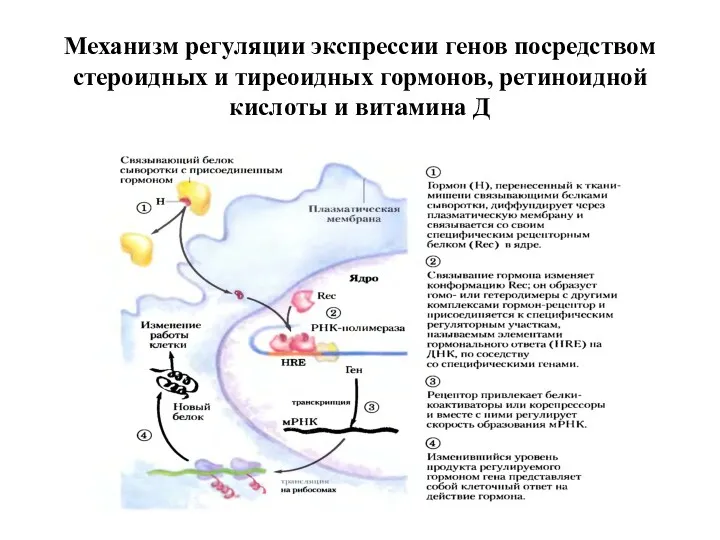
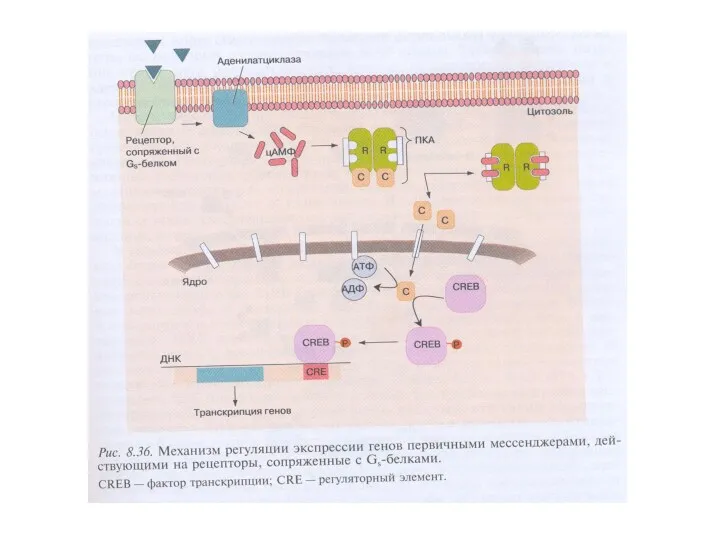
Механизм регуляции экспрессии генов посредством стероидных и тиреоидных гормонов, ретиноидной кислоты
Механизм регуляции экспрессии генов посредством стероидных и тиреоидных гормонов, ретиноидной кислоты


Вторичные посредники
Пути образования и проведение сигнала (цАМФ, цГМФ, NO, липидные мессенджеры)
Вторичные посредники
Пути образования и проведение сигнала (цАМФ, цГМФ, NO, липидные мессенджеры)

Вторичные посредники – это низкомолекулярные вещества, небелковой природы, образуются и
Вторичные посредники – это низкомолекулярные вещества, небелковой природы, образуются и
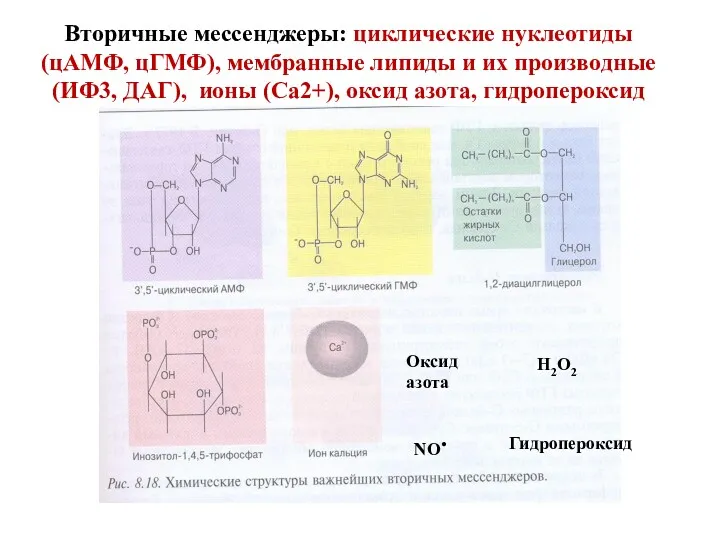
Вторичные мессенджеры: циклические нуклеотиды (цАМФ, цГМФ), мембранные липиды и их производные
Вторичные мессенджеры: циклические нуклеотиды (цАМФ, цГМФ), мембранные липиды и их производные
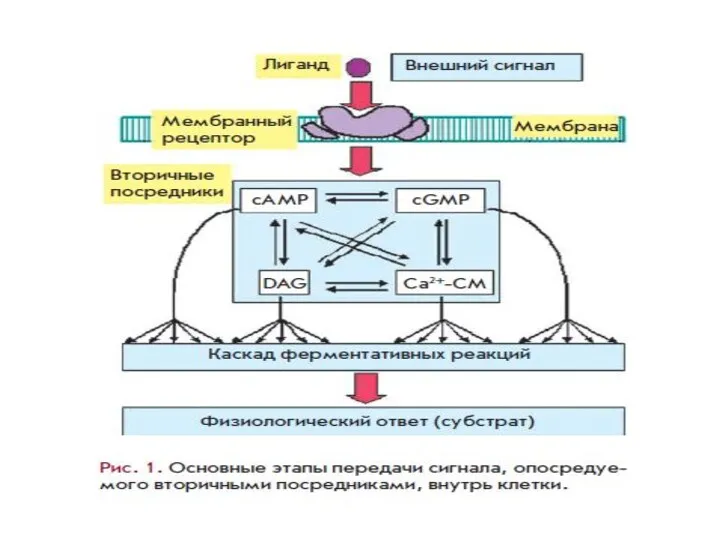

Ключевые этапы передачи сигнала посредством ВП являются общими для регуляторных систем:
агонист→рецептор→эффекторный
Ключевые этапы передачи сигнала посредством ВП являются общими для регуляторных систем:
агонист→рецептор→эффекторный
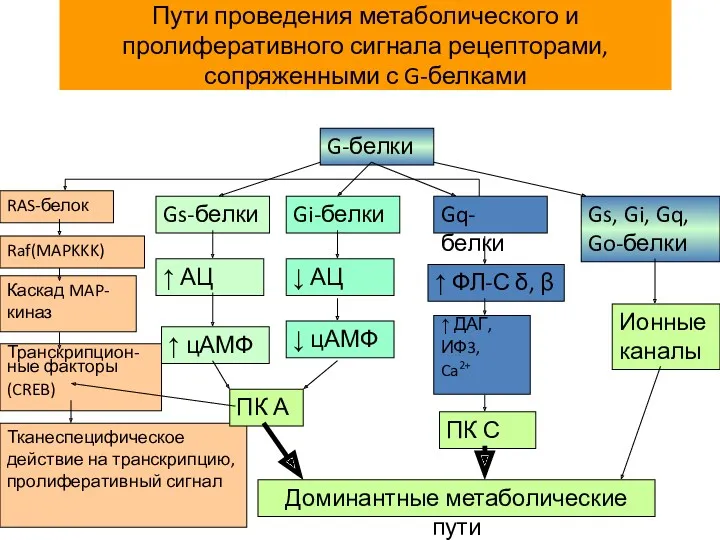
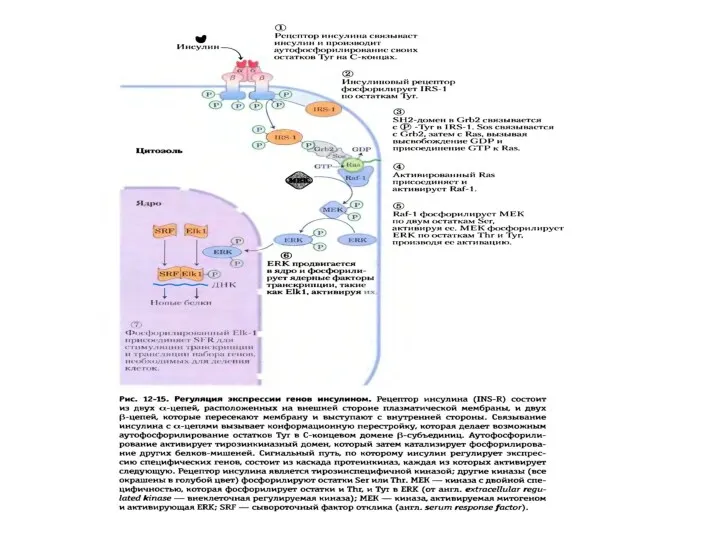
Пути проведения метаболического и пролиферативного сигнала рецепторами, сопряженными с G-белками
G-белки
Gs-белки
Gi-белки
Gq-белки
Gs, Gi,
Пути проведения метаболического и пролиферативного сигнала рецепторами, сопряженными с G-белками
G-белки
Gs-белки
Gi-белки
Gq-белки
Gs, Gi,

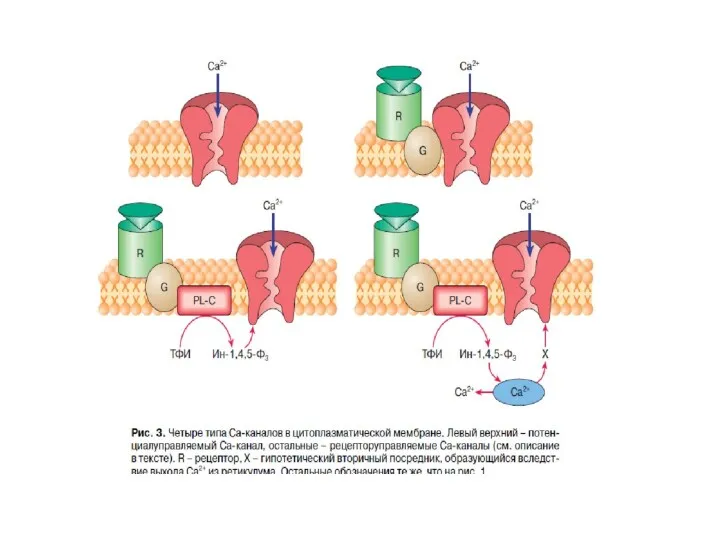
Существует 2 пути, с помощью которых GPCRs запускают образование вторичных посредников:
Существует 2 пути, с помощью которых GPCRs запускают образование вторичных посредников:
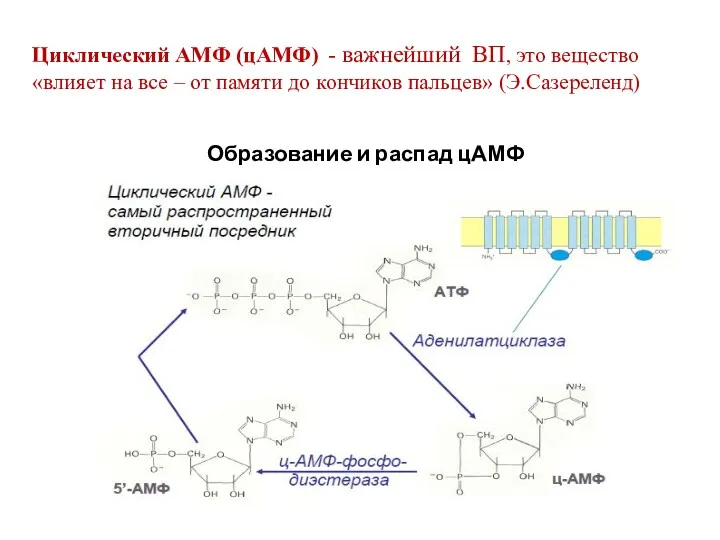
Циклический АМФ (цАМФ) - важнейший ВП, это вещество «влияет на все
Циклический АМФ (цАМФ) - важнейший ВП, это вещество «влияет на все
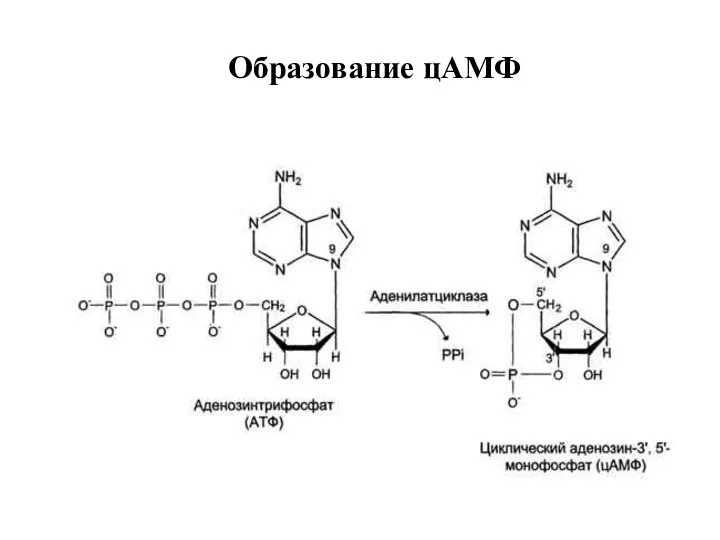
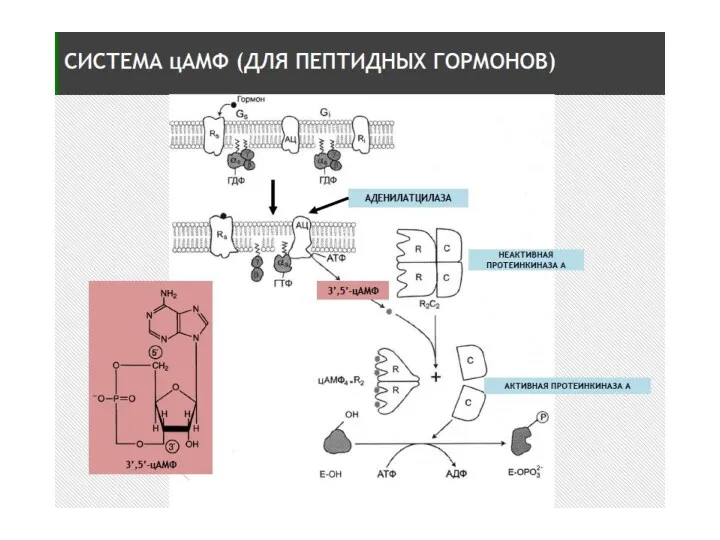
Образование цАМФ
Образование цАМФ

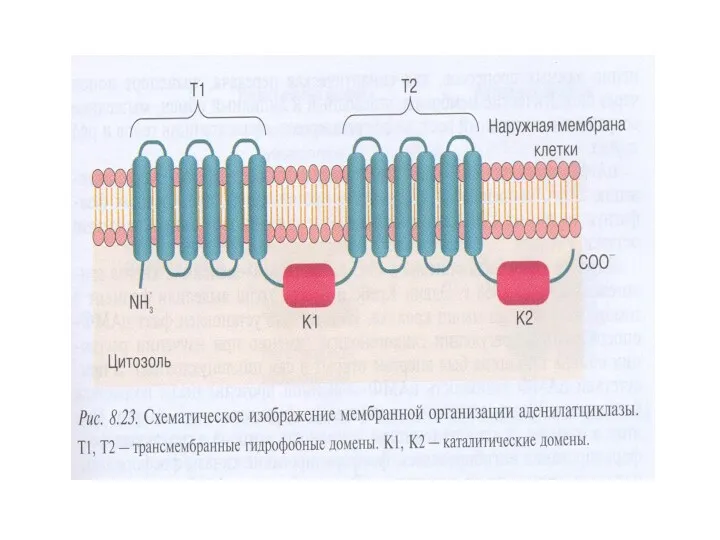
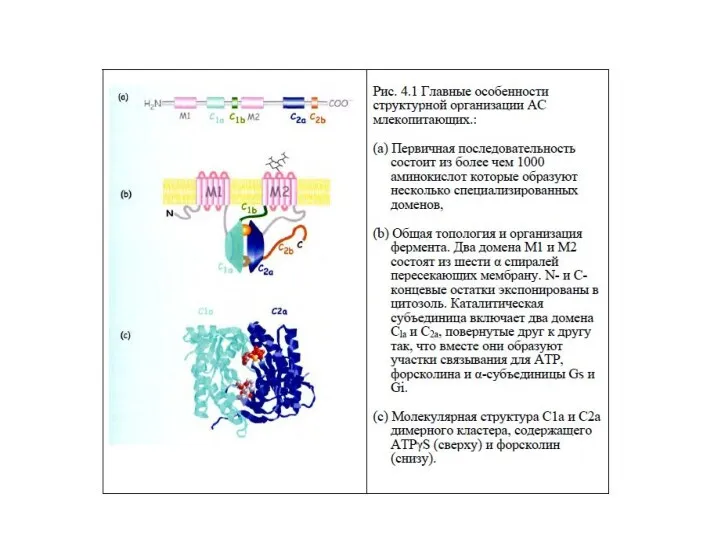
Характеристика аденилатциклазы (АЦ)
Интегральный белок плазматической мембраны
Гликопротеин, М – 110-180 кДа
Полипептидная цепь
Характеристика аденилатциклазы (АЦ)
Интегральный белок плазматической мембраны
Гликопротеин, М – 110-180 кДа
Полипептидная цепь
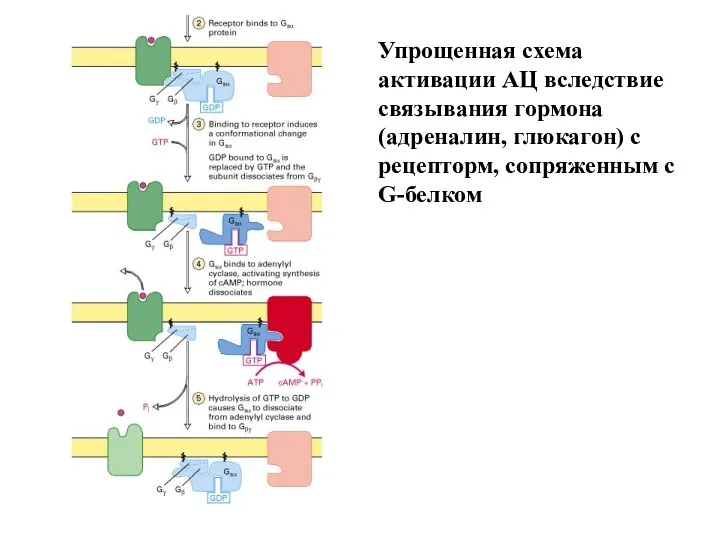
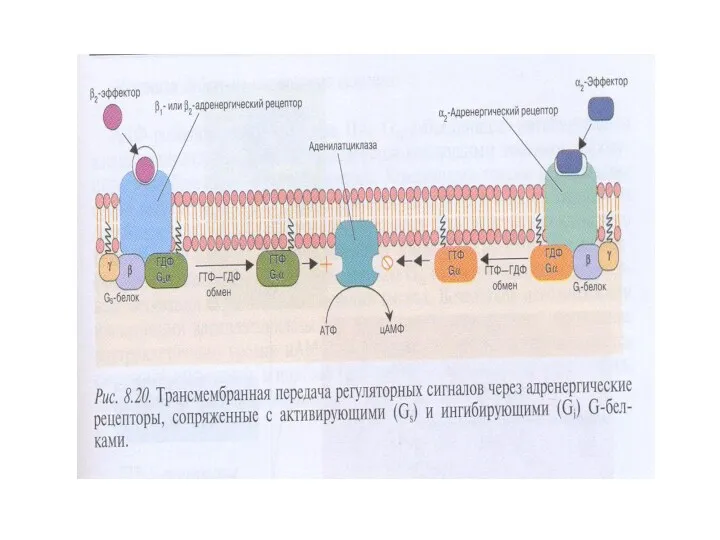
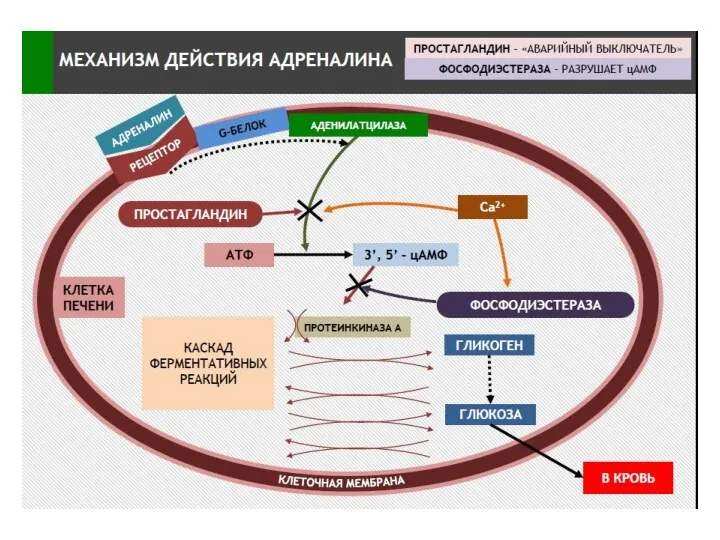
Упрощенная схема активации АЦ вследствие связывания гормона
(адреналин, глюкагон) с рецепторм, сопряженным
Упрощенная схема активации АЦ вследствие связывания гормона
(адреналин, глюкагон) с рецепторм, сопряженным
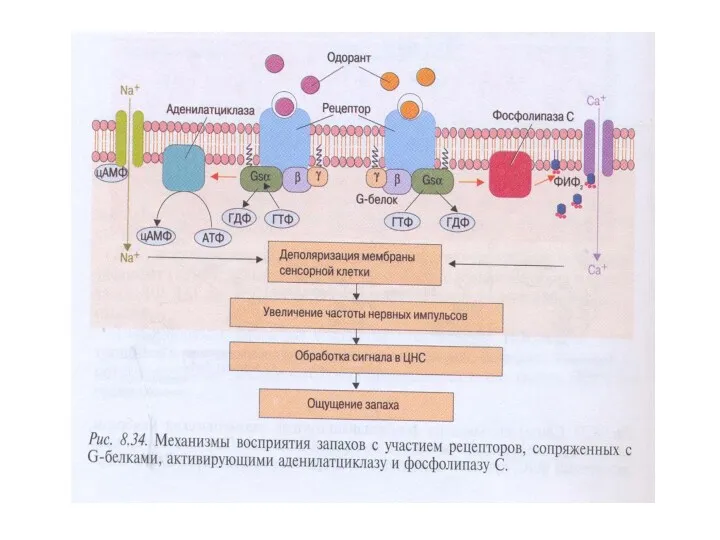
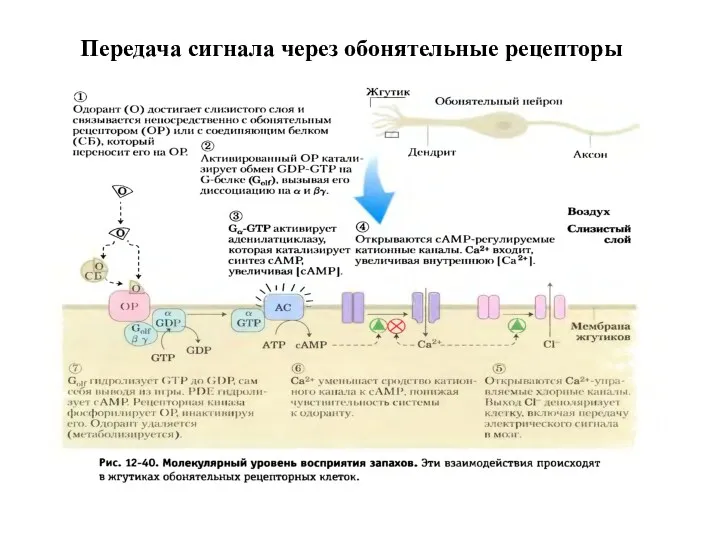
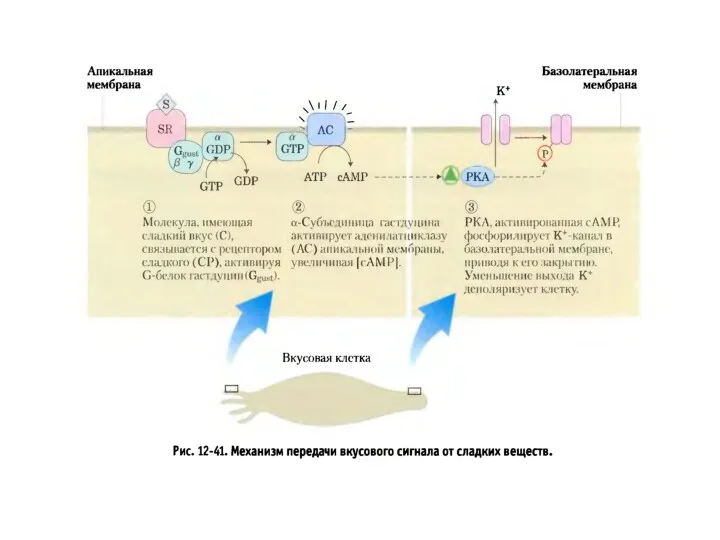
Передача сигнала через обонятельные рецепторы
Передача сигнала через обонятельные рецепторы
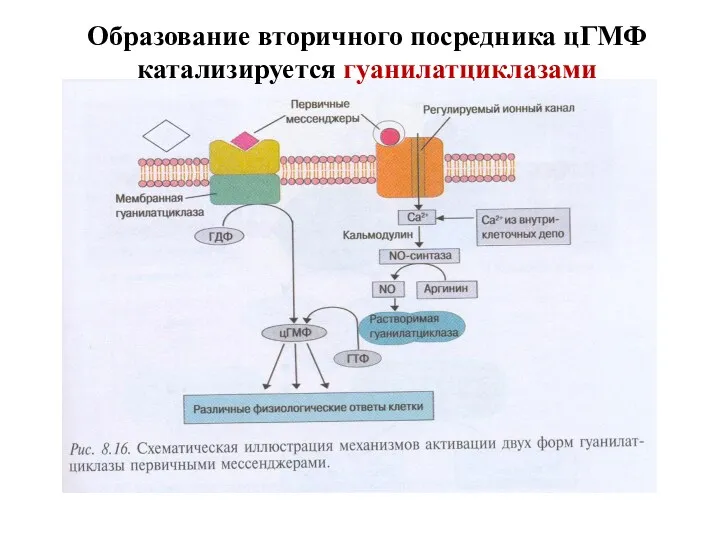
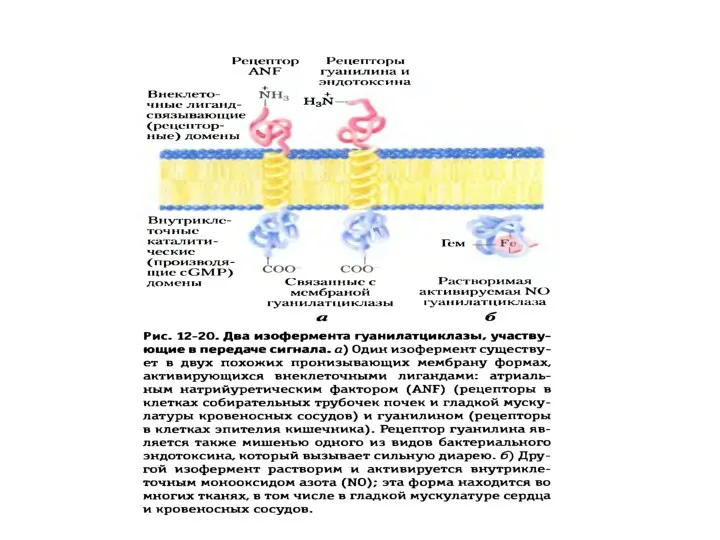
Образование вторичного посредника цГМФ катализируется гуанилатциклазами
Образование вторичного посредника цГМФ катализируется гуанилатциклазами
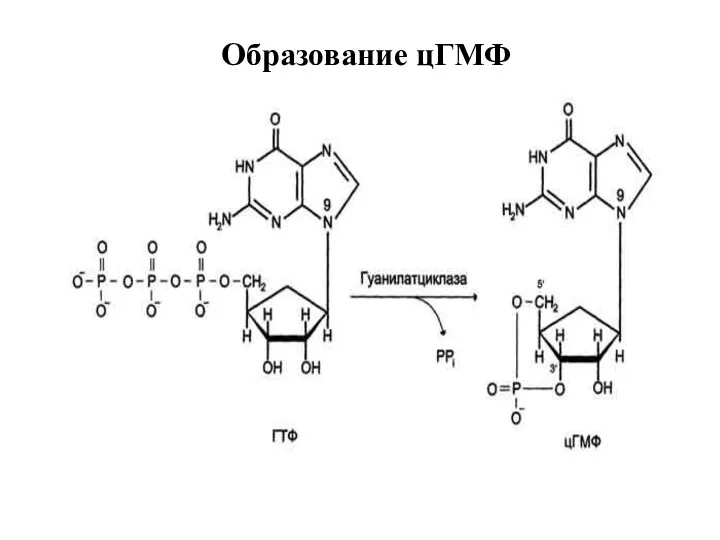
Образование цГМФ
Образование цГМФ

Гуанилатциклазы (ГЦ)
1. ГЦ катализирует образование вторичного посредника цГМФ из ГТФ.
2. В
Гуанилатциклазы (ГЦ)
1. ГЦ катализирует образование вторичного посредника цГМФ из ГТФ.
2. В
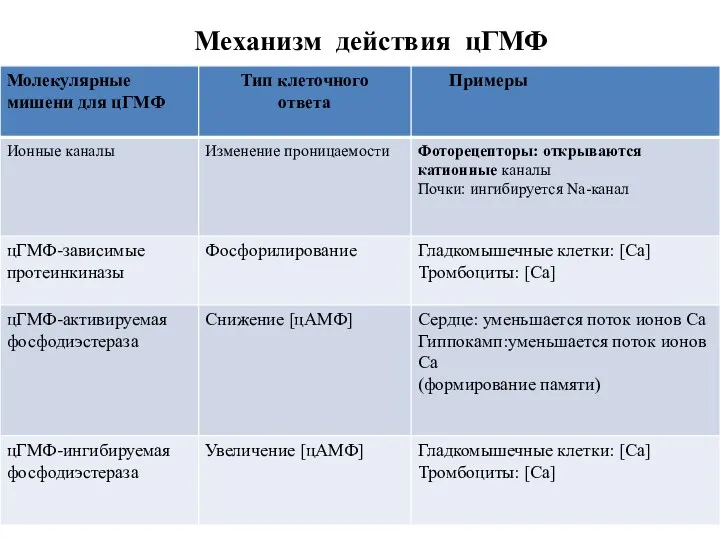
Механизм действия цГМФ
Механизм действия цГМФ
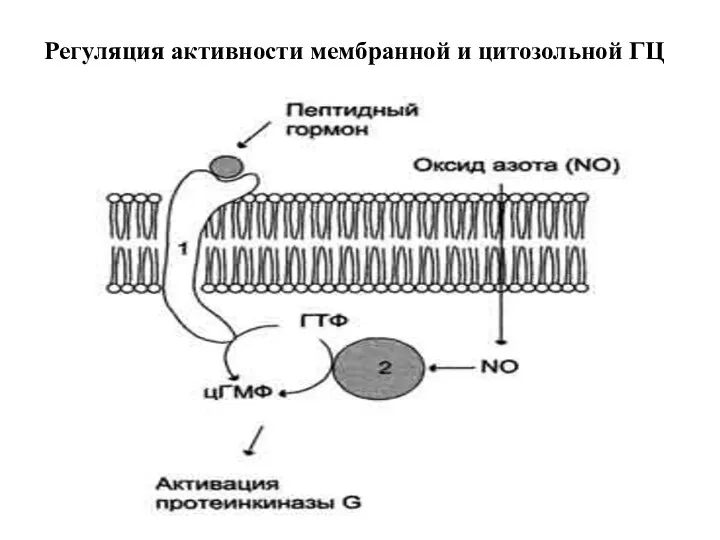
Регуляция активности мембранной и цитозольной ГЦ
Регуляция активности мембранной и цитозольной ГЦ
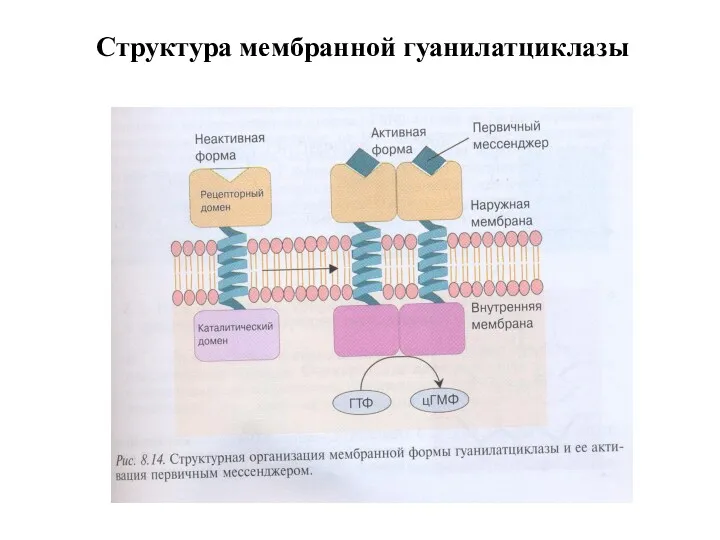
Структура мембранной гуанилатциклазы
Структура мембранной гуанилатциклазы
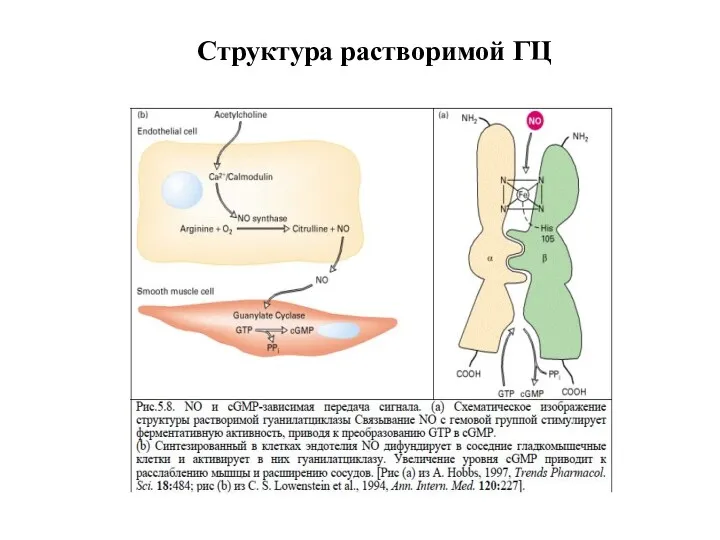
Структура растворимой ГЦ
Структура растворимой ГЦ
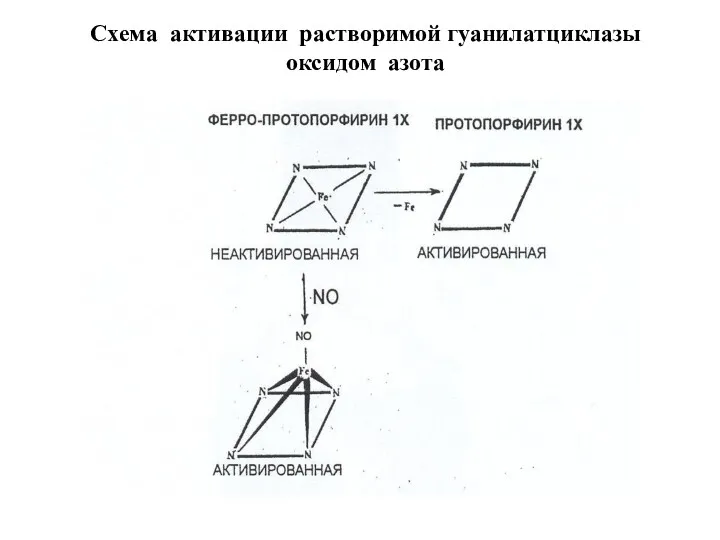
Схема активации растворимой гуанилатциклазы оксидом азота
Схема активации растворимой гуанилатциклазы оксидом азота
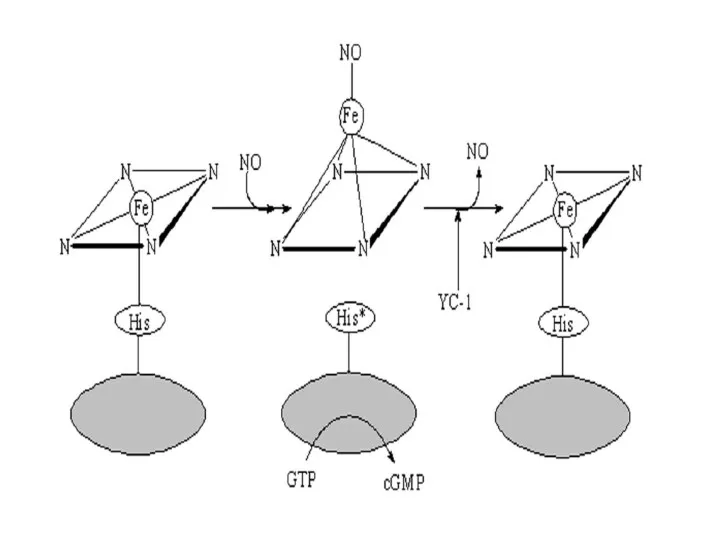

1. В неактивированном состоянии гем гуанилатциклазы связан координационно с четырьмя атомами
1. В неактивированном состоянии гем гуанилатциклазы связан координационно с четырьмя атомами
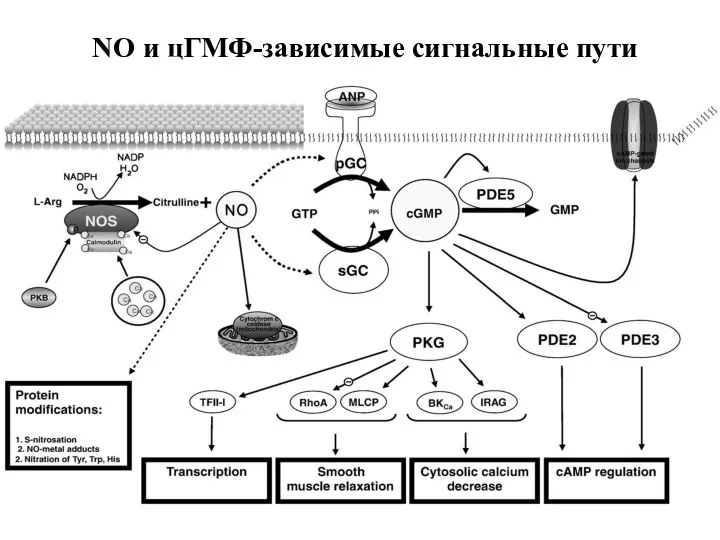
NO и цГМФ-зависимые сигнальные пути
NO и цГМФ-зависимые сигнальные пути
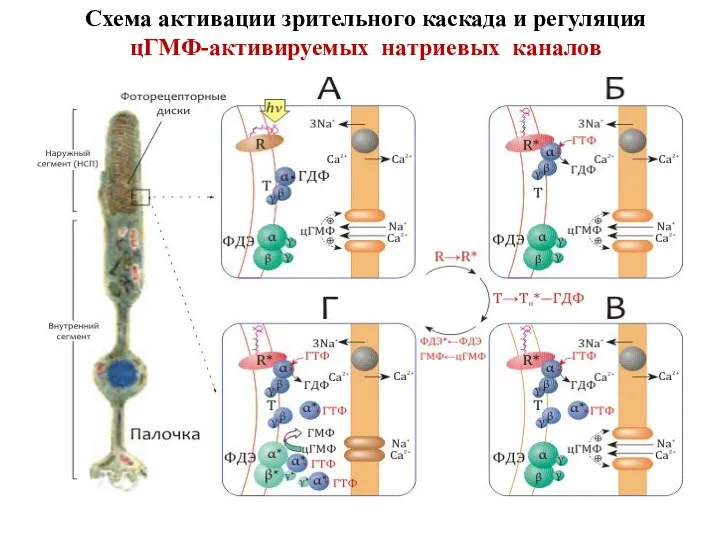
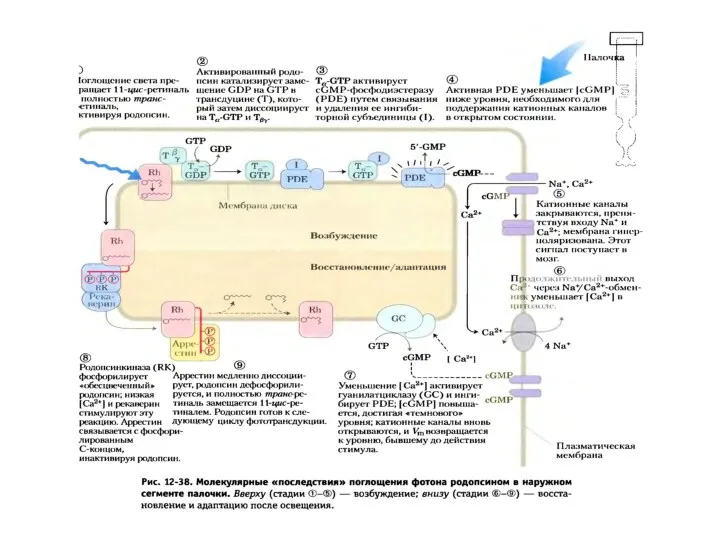
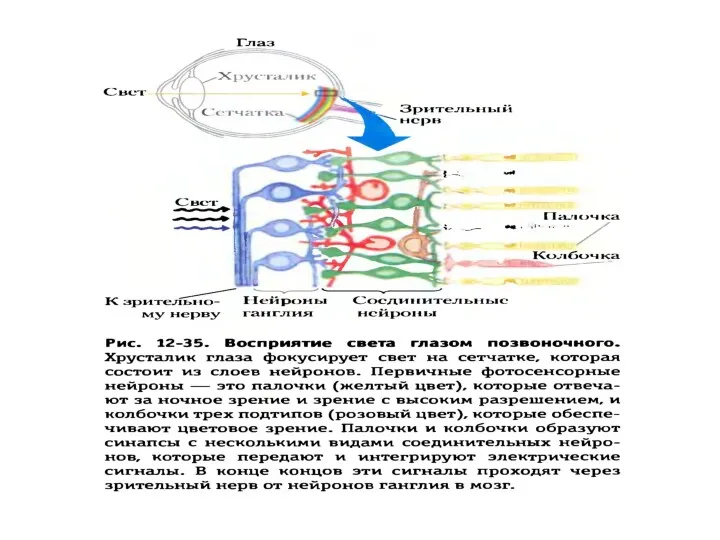
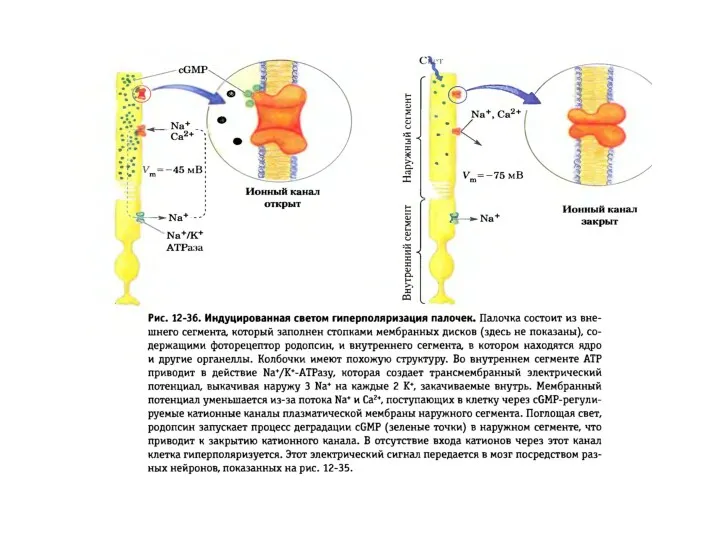
Схема активации зрительного каскада и регуляция цГМФ-активируемых натриевых каналов
Схема активации зрительного каскада и регуляция цГМФ-активируемых натриевых каналов

цГМФ и гуанилатциклазы (Нельсон, Кокс, 2011)
цГМФ и гуанилатциклазы (Нельсон, Кокс, 2011)
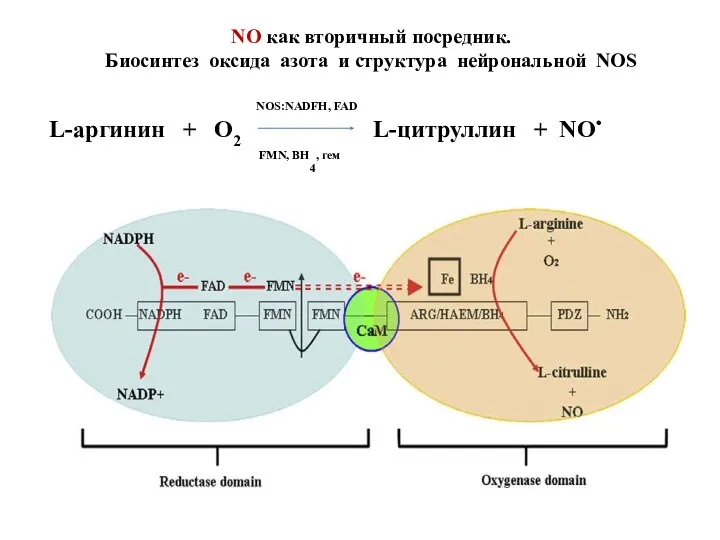
NO как вторичный посредник.
Биосинтез оксида азота и структура нейрональной NOS
FMN, ВH4,
NO как вторичный посредник.
Биосинтез оксида азота и структура нейрональной NOS
FMN, ВH4,
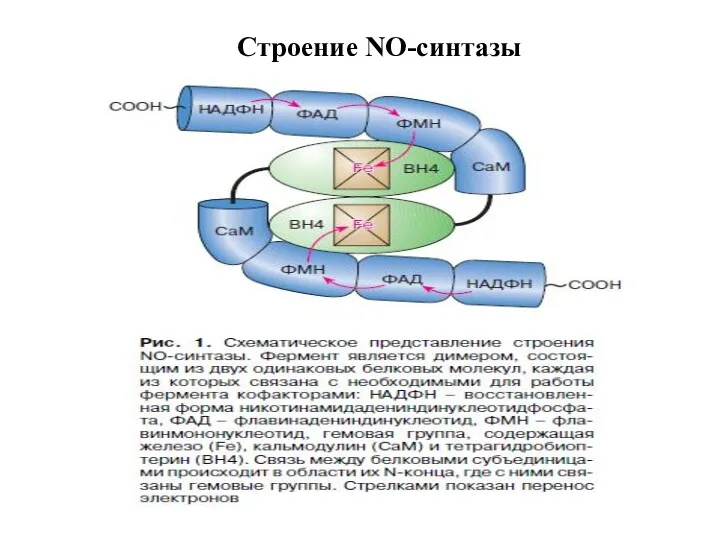
Строение NO-синтазы
Строение NO-синтазы

Активность NOS регулируется :
Фосфорилированием
- ПКА
- ПКС
- Са/кальмодулин-зависимой
Активность NOS регулируется :
Фосфорилированием
- ПКА
- ПКС
- Са/кальмодулин-зависимой

NO запускает различные downstream пути и регулирует:
вазодилятацию
нейротрансмиссию
макрофагальную цитотоксичность
релаксацию гладкомышечных клеток ЖКТ
бронходилятацию
модуляцию
NO запускает различные downstream пути и регулирует:
вазодилятацию
нейротрансмиссию
макрофагальную цитотоксичность
релаксацию гладкомышечных клеток ЖКТ
бронходилятацию
модуляцию
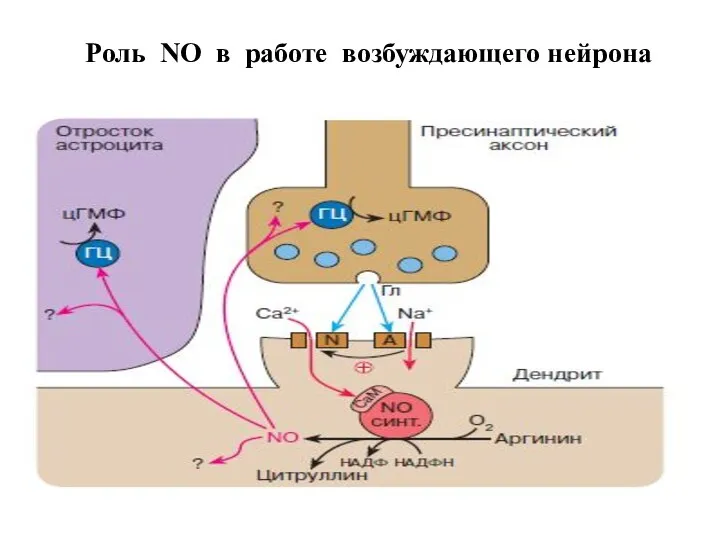
Роль NO в работе возбуждающего нейрона
Роль NO в работе возбуждающего нейрона
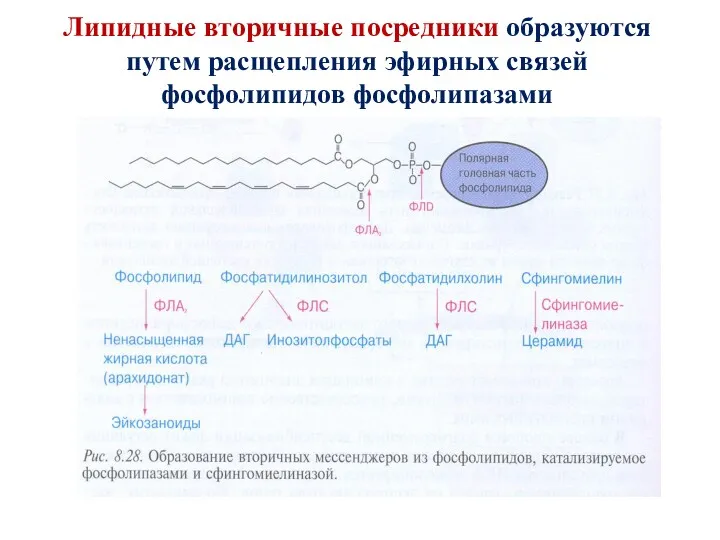
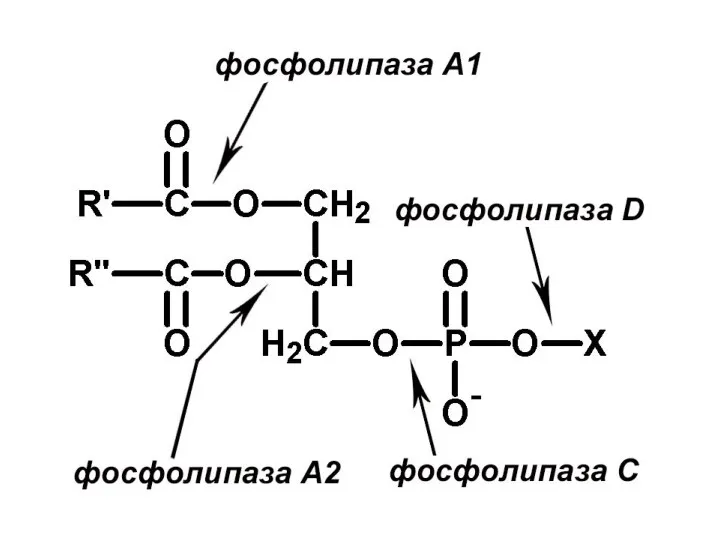
Липидные вторичные посредники образуются путем расщепления эфирных связей фосфолипидов фосфолипазами
Липидные вторичные посредники образуются путем расщепления эфирных связей фосфолипидов фосфолипазами

Характеристика фосфолипаз (ФЛ)
ФЛ - гидролазы, катализирующие катаболизм глицеро-фосфолипидов. Различают: 1) секреторные
Характеристика фосфолипаз (ФЛ)
ФЛ - гидролазы, катализирующие катаболизм глицеро-фосфолипидов. Различают: 1) секреторные
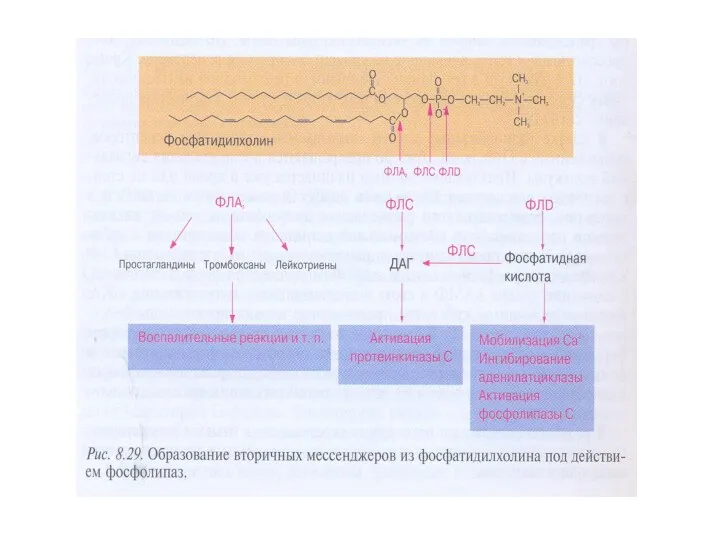
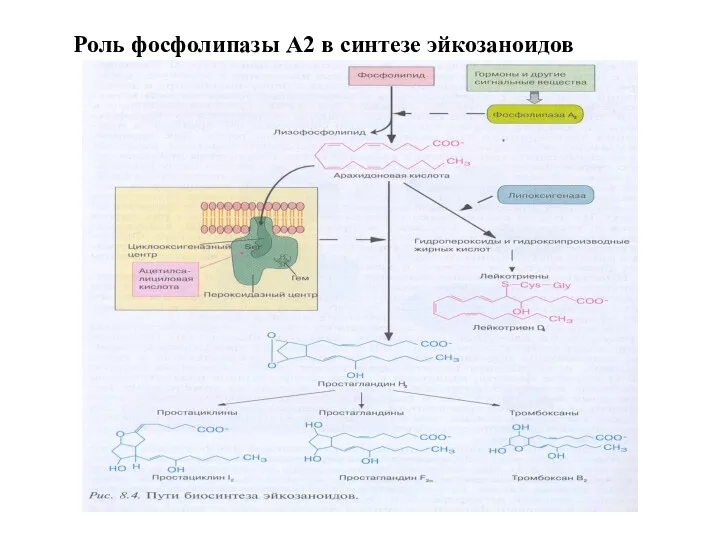
Роль фосфолипазы А2 в синтезе эйкозаноидов
Роль фосфолипазы А2 в синтезе эйкозаноидов
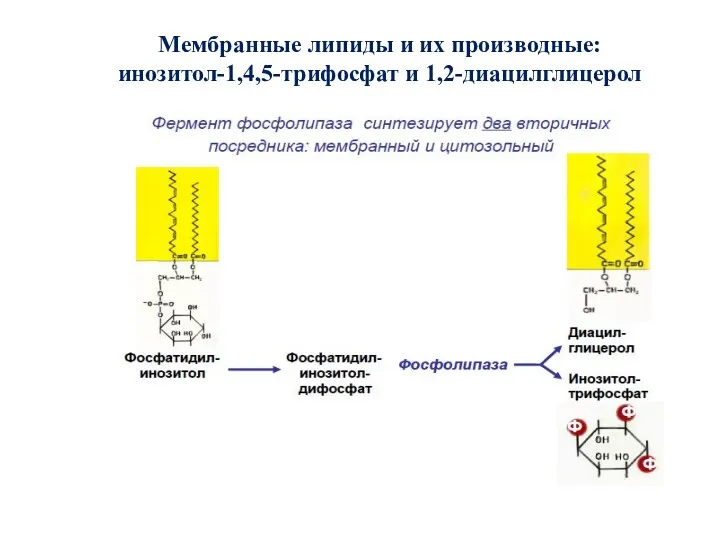
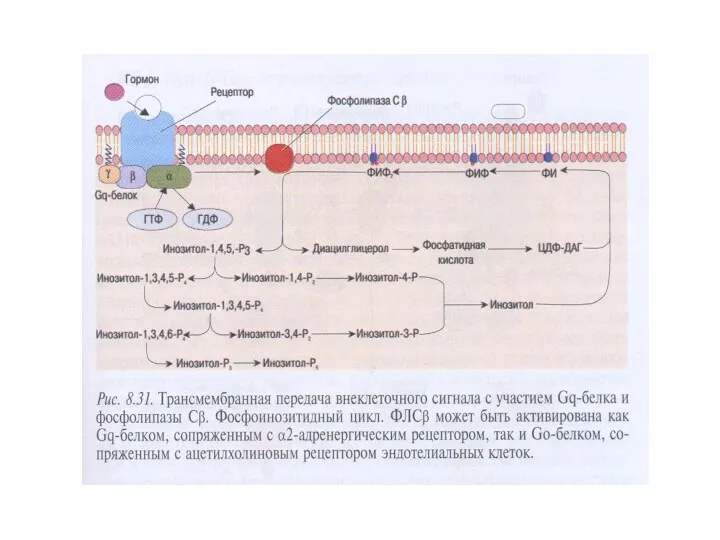
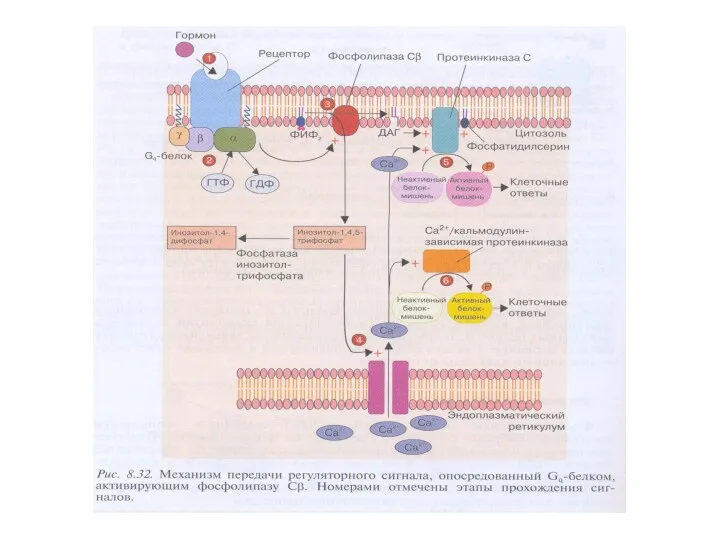
Мембранные липиды и их производные: инозитол-1,4,5-трифосфат и 1,2-диацилглицерол
Мембранные липиды и их производные: инозитол-1,4,5-трифосфат и 1,2-диацилглицерол
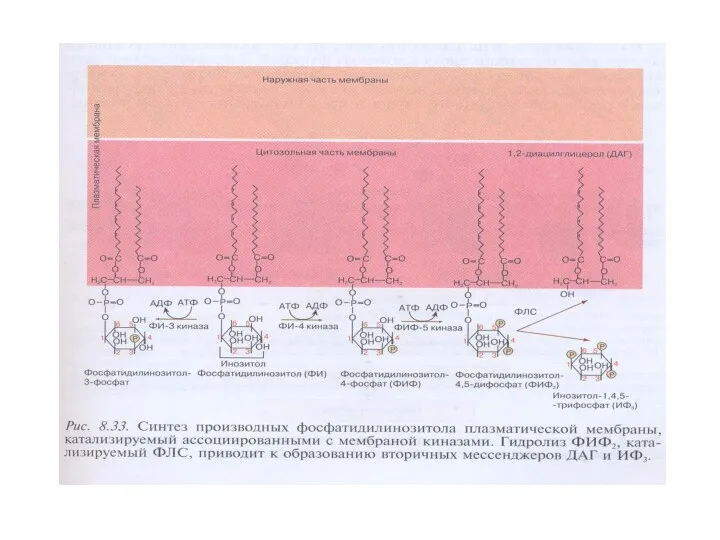

Синтез производных фосфатидилинозитола в плазматической мембране, гидролиз ФИФ2
фосфолипазой С
ФИ-4-киназа →
Синтез производных фосфатидилинозитола в плазматической мембране, гидролиз ФИФ2
фосфолипазой С
ФИ-4-киназа →

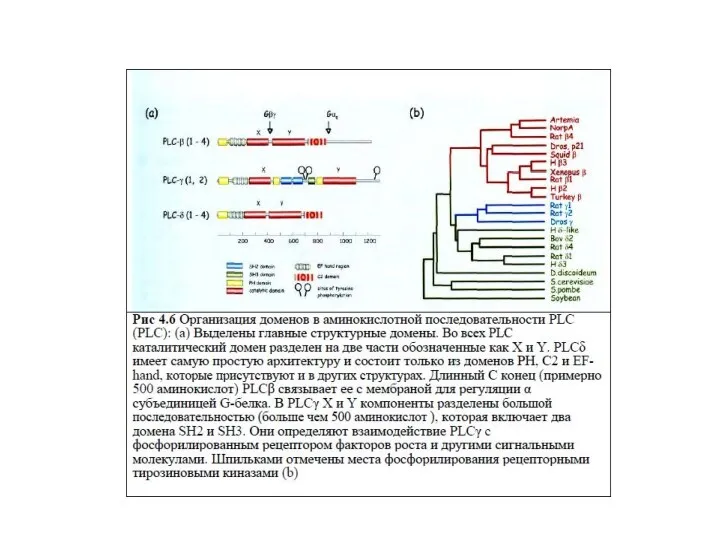
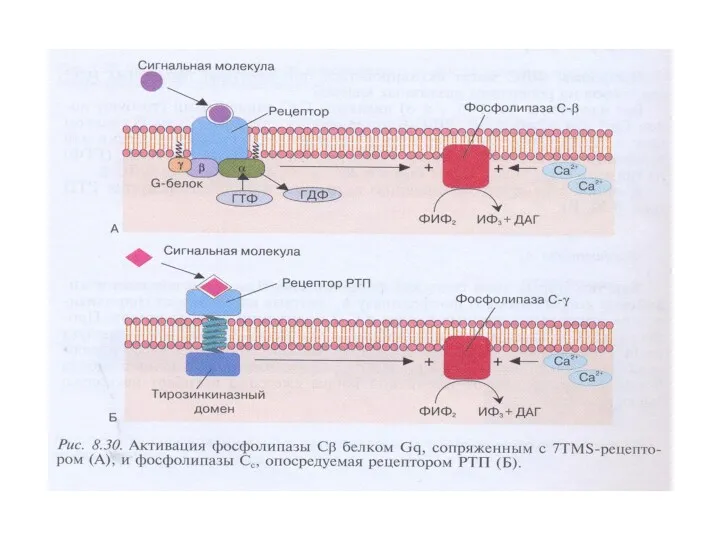
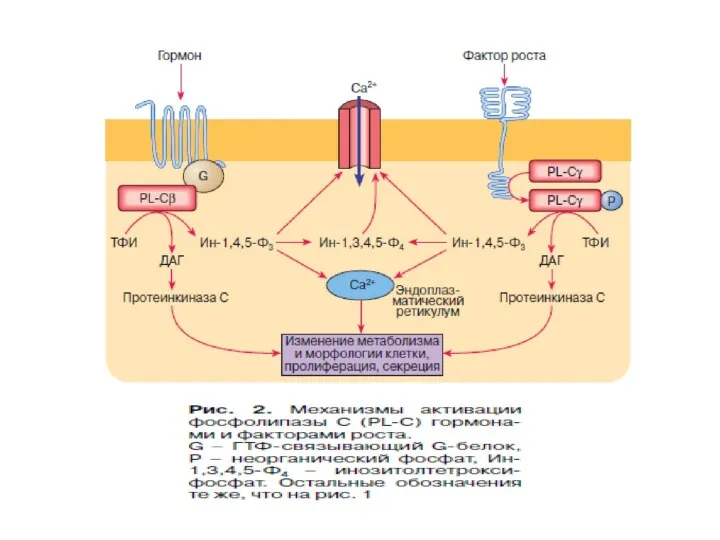
Характеристика фосфолипазы С
1.Известно 3 класса ФЛС: ФЛСβ, ФЛСγ, ФЛСδ, которые включают
Характеристика фосфолипазы С
1.Известно 3 класса ФЛС: ФЛСβ, ФЛСγ, ФЛСδ, которые включают
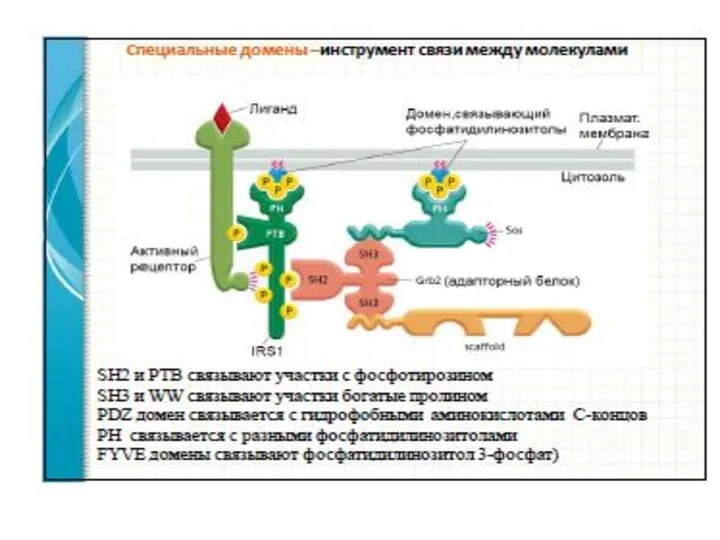

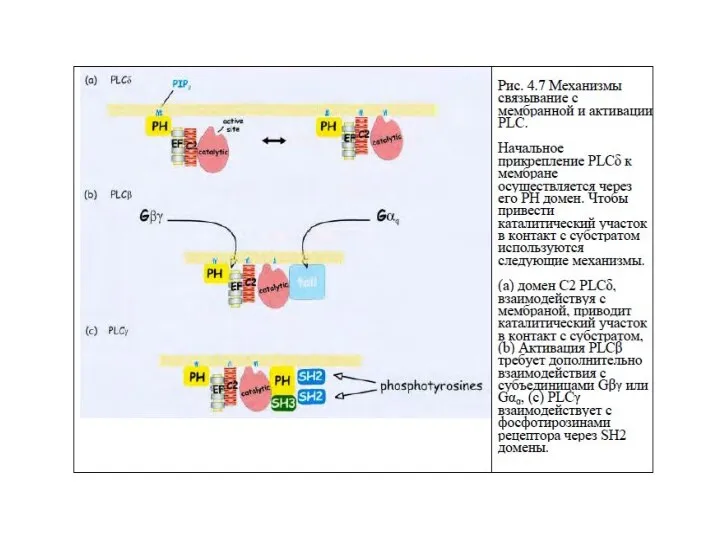
Механизм связывания с мембраной и активация ФЛС
1.Начальное связывание ФЛС с мембраной
Механизм связывания с мембраной и активация ФЛС
1.Начальное связывание ФЛС с мембраной
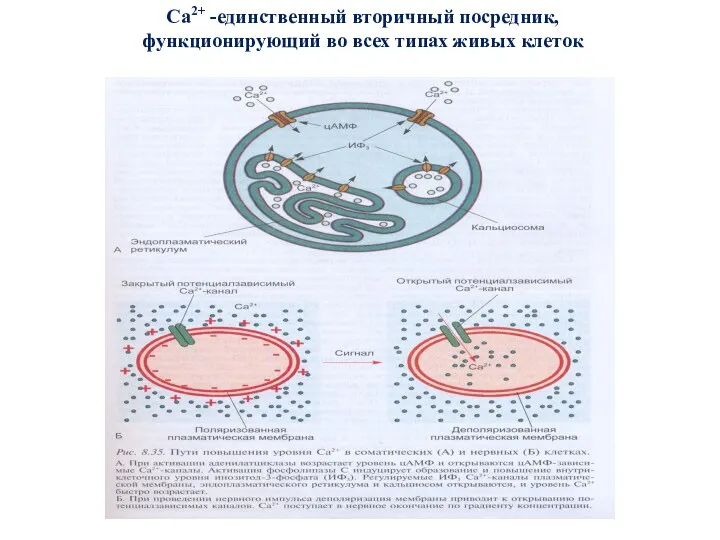
Са2+ -единственный вторичный посредник, функционирующий во всех типах живых клеток
Са2+ -единственный вторичный посредник, функционирующий во всех типах живых клеток

Са 2+ - ключевой вторичный посредник
Различают 3 состояния Са2+ в
Са 2+ - ключевой вторичный посредник
Различают 3 состояния Са2+ в

Особенности Са 2+ как вторичного посредника
1. Са 2+ - неметаболизируемый
Особенности Са 2+ как вторичного посредника
1. Са 2+ - неметаболизируемый

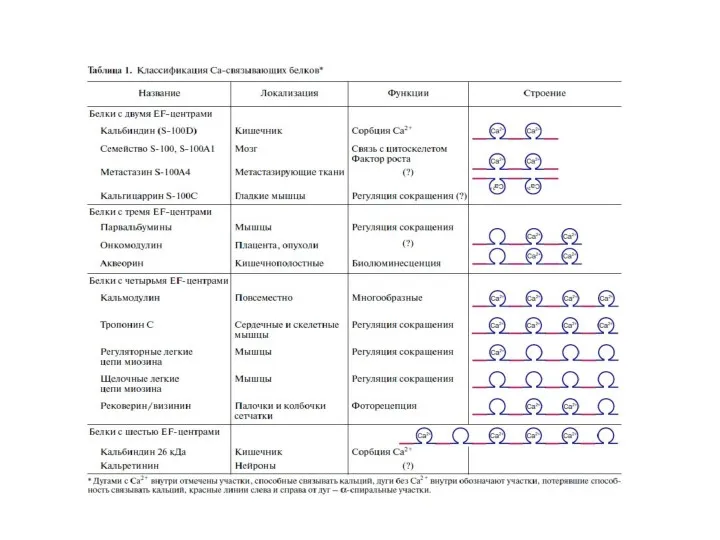
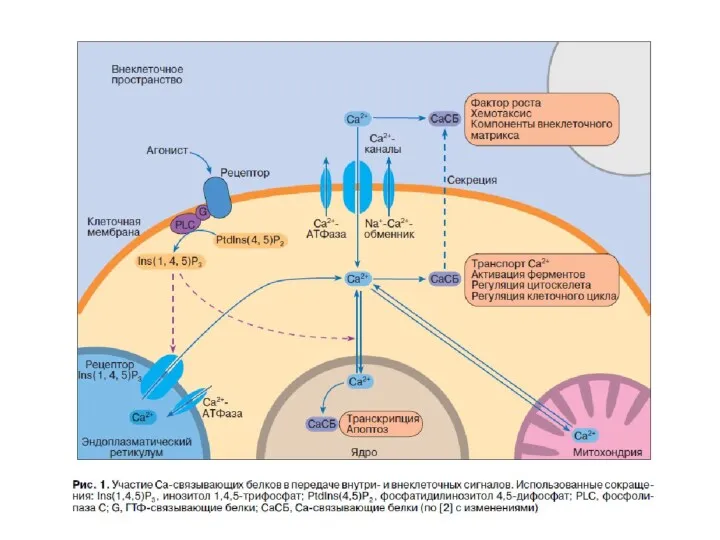
Внутриклеточные Са2+-связывающие белки определяют Са2-опосредованные сигналы
Выделяют 3 группы Са2+-связывающих белков:
1.ЕF-hand-белки (цитоплазма,
Внутриклеточные Са2+-связывающие белки определяют Са2-опосредованные сигналы
Выделяют 3 группы Са2+-связывающих белков:
1.ЕF-hand-белки (цитоплазма,
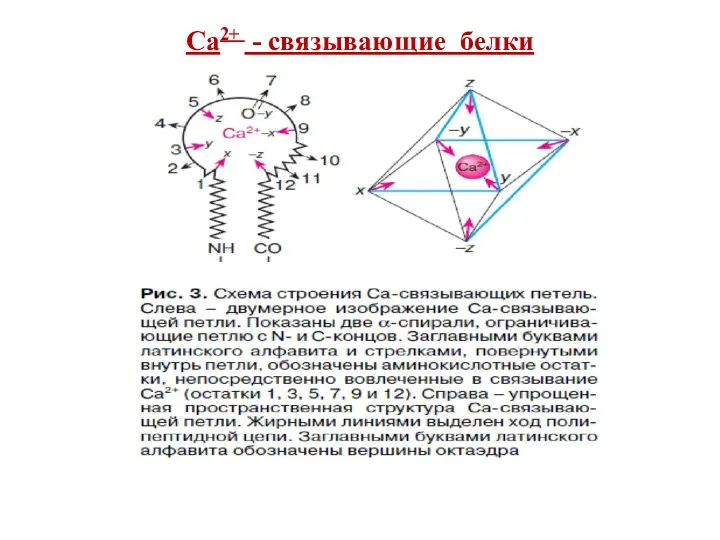
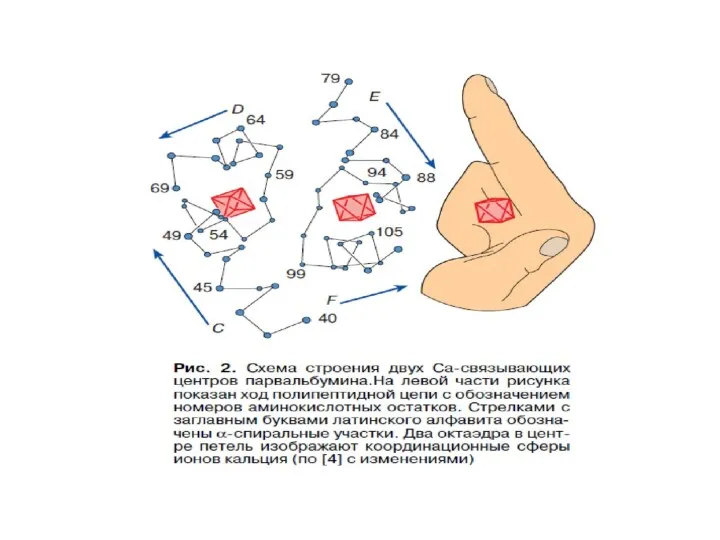
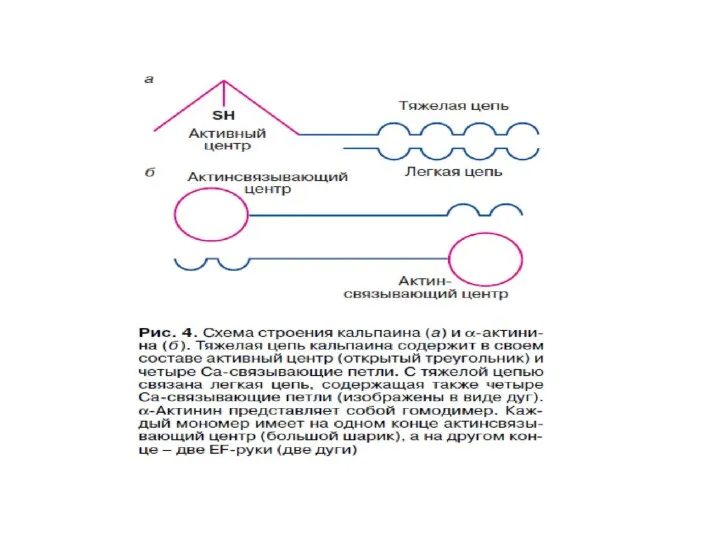
Са2+ - связывающие белки
Са2+ - связывающие белки

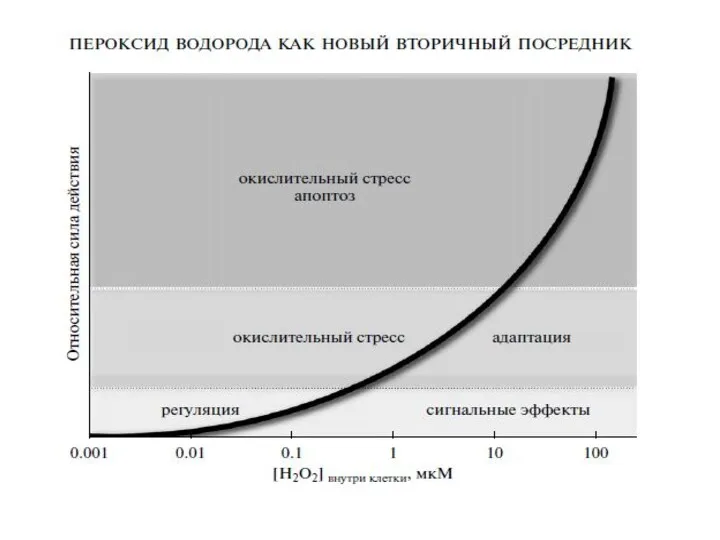

Источники перекиси водорода в клетке:
НАДФН-оксидаза
Электрон-транспортная цепь митохондрий
Электрон-транспортная цепь микросом
Ксантиноксидоредуктаза (КОР)
Супероксиддисмутаза
Источники перекиси водорода в клетке:
НАДФН-оксидаза
Электрон-транспортная цепь митохондрий
Электрон-транспортная цепь микросом
Ксантиноксидоредуктаза (КОР)
Супероксиддисмутаза
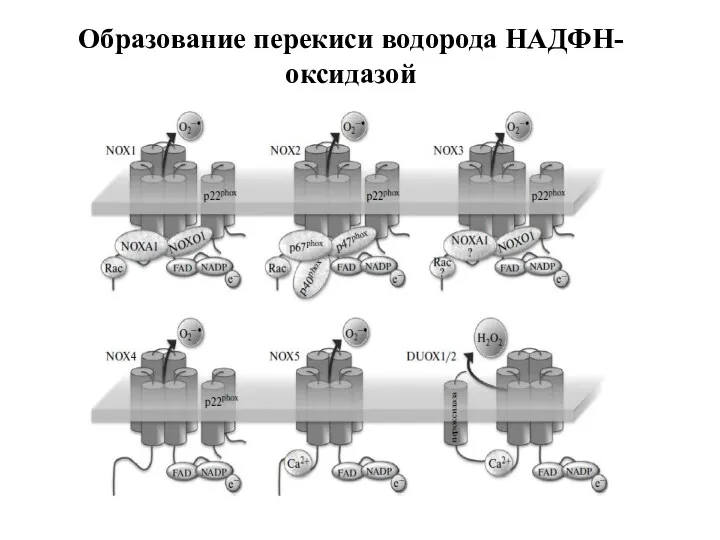
Образование перекиси водорода НАДФН-оксидазой
Образование перекиси водорода НАДФН-оксидазой
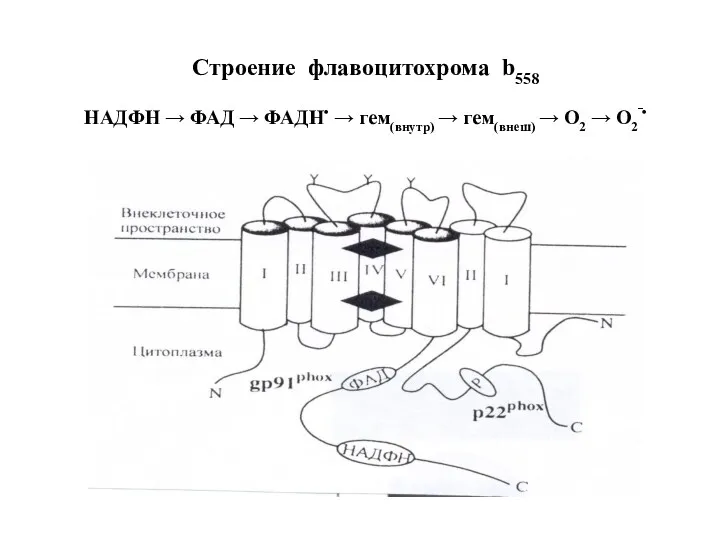
Строение флавоцитохрома b558
НАДФН → ФАД → ФАДН• → гем(внутр) → гем(внеш)
Строение флавоцитохрома b558 НАДФН → ФАД → ФАДН• → гем(внутр) → гем(внеш)
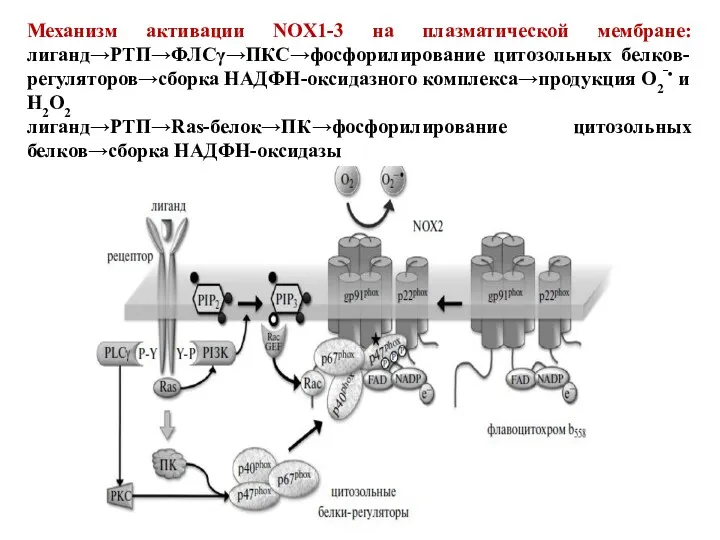
Механизм активации NOX1-3 на плазматической мембране: лиганд→РТП→ФЛСγ→ПКС→фосфорилирование цитозольных белков-регуляторов→сборка НАДФН-оксидазного комплекса→продукция
Механизм активации NOX1-3 на плазматической мембране: лиганд→РТП→ФЛСγ→ПКС→фосфорилирование цитозольных белков-регуляторов→сборка НАДФН-оксидазного комплекса→продукция
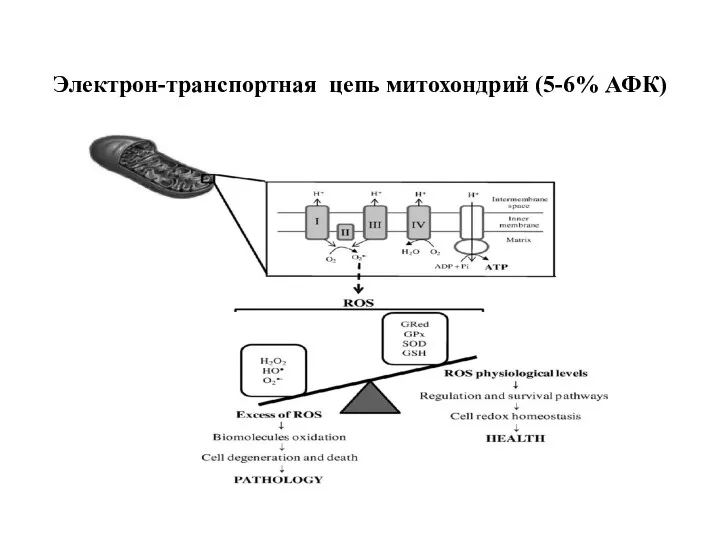
Электрон-транспортная цепь митохондрий (5-6% АФК)
Электрон-транспортная цепь митохондрий (5-6% АФК)
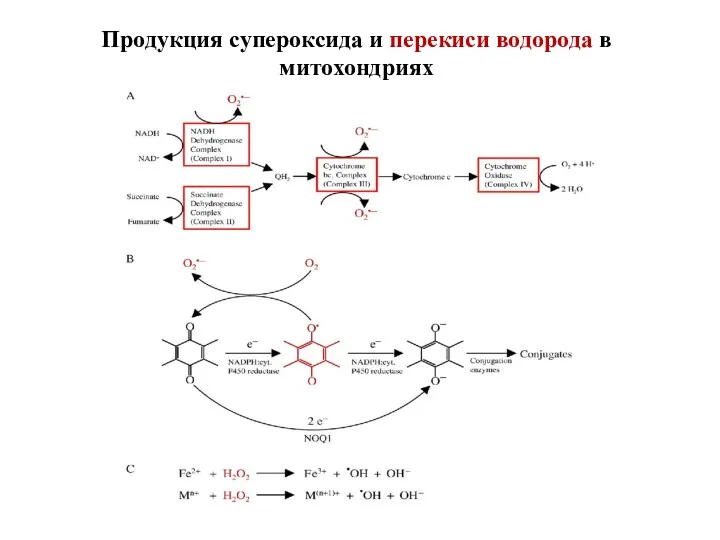
Продукция супероксида и перекиси водорода в митохондриях
Продукция супероксида и перекиси водорода в митохондриях
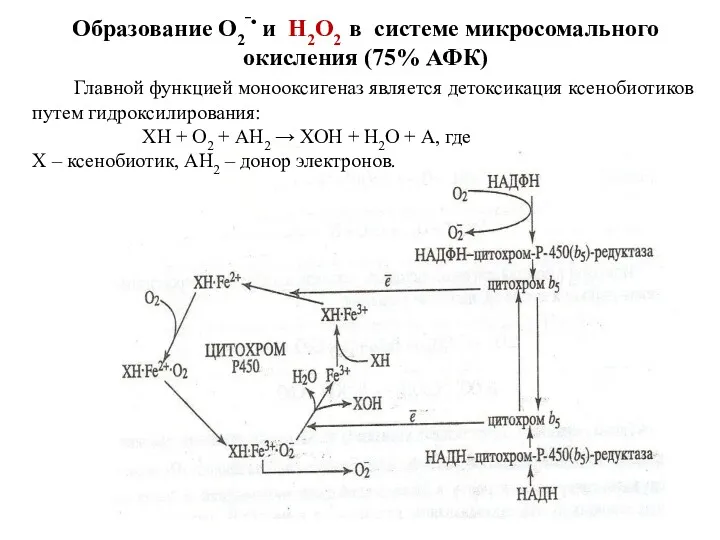
Образование О2‾• и Н2О2 в системе микросомального окисления (75% АФК)
Главной
Образование О2‾• и Н2О2 в системе микросомального окисления (75% АФК)
Главной

Ксантиноксидоредуктаза – источник перекиси водорода в клетке
Ксантиноксидоредуктаза представлена двумя изоформами:
Ксантиноксидоредуктаза – источник перекиси водорода в клетке
Ксантиноксидоредуктаза представлена двумя изоформами:
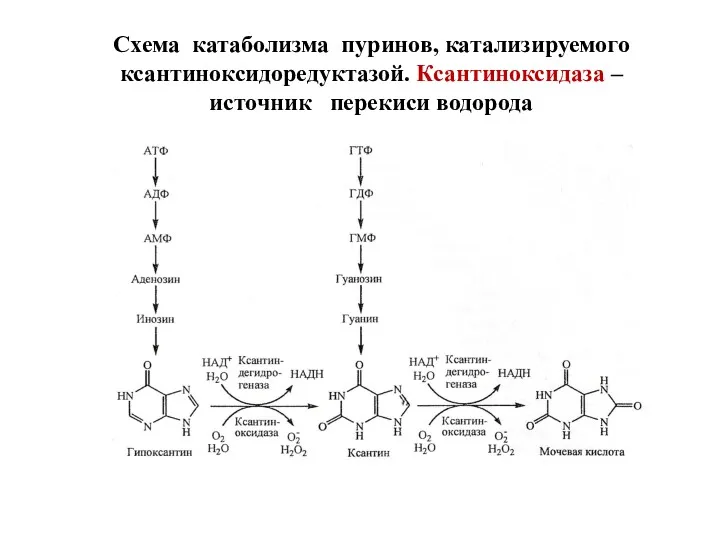
Схема катаболизма пуринов, катализируемого ксантиноксидоредуктазой. Ксантиноксидаза – источник перекиси водорода
Схема катаболизма пуринов, катализируемого ксантиноксидоредуктазой. Ксантиноксидаза – источник перекиси водорода
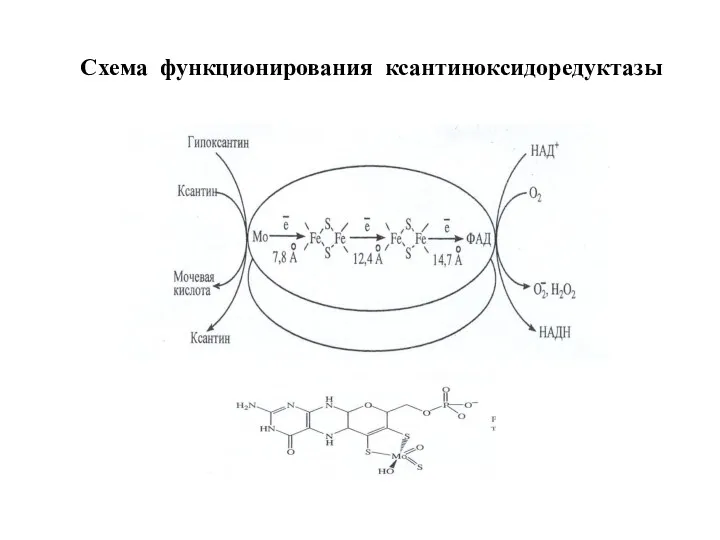
Схема функционирования ксантиноксидоредуктазы
Схема функционирования ксантиноксидоредуктазы

Супероксиддисмутазы (СОД) – суперсемейство ферментов, относящихся к классу оксидоредуктаз и катализирующих
Супероксиддисмутазы (СОД) – суперсемейство ферментов, относящихся к классу оксидоредуктаз и катализирующих
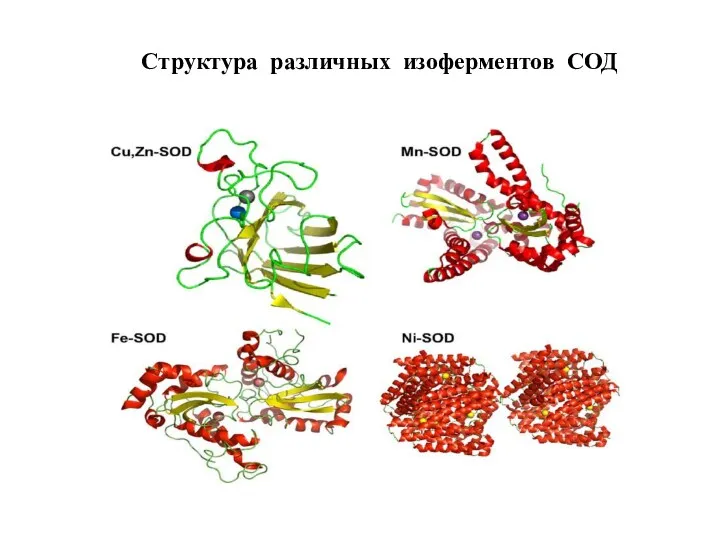
Структура различных изоферментов СОД
Структура различных изоферментов СОД
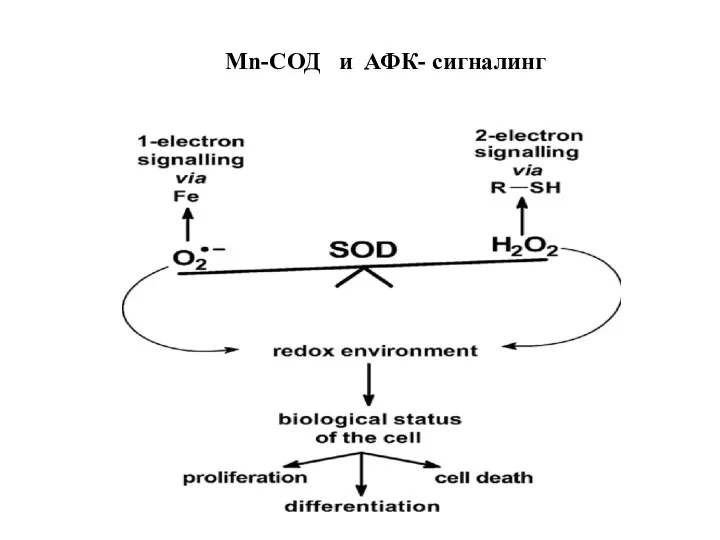
Mn-СОД и АФК- сигналинг
Mn-СОД и АФК- сигналинг

Элиминация перекиси водорода в клетке осуществляется ферментативным путем:
1.Каталаза – гемсодержащий внутриклеточный
Элиминация перекиси водорода в клетке осуществляется ферментативным путем:
1.Каталаза – гемсодержащий внутриклеточный
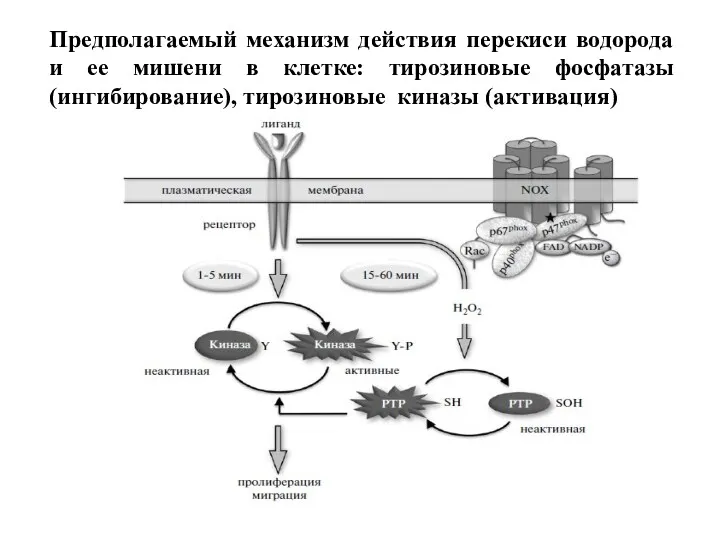
Предполагаемый механизм действия перекиси водорода и ее мишени в клетке: тирозиновые
Предполагаемый механизм действия перекиси водорода и ее мишени в клетке: тирозиновые

Дополнительные слайды
Дополнительные слайды

 Растительный и животный мир пустыни
Растительный и животный мир пустыни Промысловые рыбы
Промысловые рыбы Энергообеспечение мышечной деятельности
Энергообеспечение мышечной деятельности Урок-презентация. Плоды, их строение, значение, распространение в природе
Урок-презентация. Плоды, их строение, значение, распространение в природе Жизнь в Мировом океане
Жизнь в Мировом океане Что такое онтогенез? Представления об онтогенезе: эпигенез, преформизм, современное
Что такое онтогенез? Представления об онтогенезе: эпигенез, преформизм, современное Биотехнология в селекции растений. Часть 1. Селекция на важнейшие хозяйственные свойства
Биотехнология в селекции растений. Часть 1. Селекция на важнейшие хозяйственные свойства Пищеварение в кишечнике. 8 класс
Пищеварение в кишечнике. 8 класс Уход за комнатными растениями в уголке природы
Уход за комнатными растениями в уголке природы Семя, его строение и значение
Семя, его строение и значение Влияние экологических факторов на организмы
Влияние экологических факторов на организмы КЛЕТКА
КЛЕТКА Генетическая инженерия. Тема №1
Генетическая инженерия. Тема №1 Вид. Критерии вида
Вид. Критерии вида Популяционно-статистический метод
Популяционно-статистический метод Сүйектің байланысу түрлері
Сүйектің байланысу түрлері Тип круглые черви
Тип круглые черви Проявления генов в развитии. Плейотропное действие гена
Проявления генов в развитии. Плейотропное действие гена Энергетический обмен углеводов. 10 класс
Энергетический обмен углеводов. 10 класс среда обитания 9 класс
среда обитания 9 класс Повреждение ядра клетки
Повреждение ядра клетки Достижения в селекции животных
Достижения в селекции животных головоногия моллюски
головоногия моллюски 7 самых ядовитых растений России
7 самых ядовитых растений России Адаптация организма человека к факторам окружающей среды
Адаптация организма человека к факторам окружающей среды презентация Интеллектуальный марафон
презентация Интеллектуальный марафон Корень. Типы корневых систем. Урок биологии, 6 класс
Корень. Типы корневых систем. Урок биологии, 6 класс Физиология вегетативной нервной системы (ВНС)
Физиология вегетативной нервной системы (ВНС)