- Молекулярная генетика онтогенеза

Содержание
- 2. Существует мнение: Все, что сделано ранее и делается сегодня – редукционистская биология. Надо изучать биологические системы
- 3. Системная биология (systems biology) — междисциплинарная наука о жизни, изучающая сложные взаимодействия в живых системах, использующая
- 4. Компьютерная модель полного жизненного цикла патогена Mycoplasma genitalium (2012 г.) Идея: Если на базе огромного количества
- 5. Однако: Без достижений редукционистской биологии невозможно создать «системную биологию».
- 6. 2006 – Э. Файер и К. Мелло «за открытие РНК-интерференции — эффекта гашения активности генов». 2007
- 7. Утром белки-активаторы BMal1 и Clock переходят из цитоплазмы в ядро клетки и там прикрепляются к участку
- 8. Генная инженерия in vitro Векторы (плазмиды, вирусы, фаги и др.) Ферменты (рестриктазы, лигазы, фосфатазы, ревертазы) (1967-1970
- 9. культивирование соматических клеток животных in vitro (конец 50-х годов XX века, в России – Н.И. Шапиро),
- 10. Селективная среда ГAT (НАТ) (гипоксантин, аминоптерин и тимидин) 1) Основной путь синтеза пуриновых и пиримидиновых оснований
- 11. 1) иммунизация 2) выделение В-лимфоцитов 3) клетки миеломы ГФРТ- 4) слияние 5) сегрегация 6) Селекция 7)
- 12. Пластмассовые антитела? Смесь мономеров и антигена
- 13. Трансдукция (transduction) [лат. transductio — перемещение] — перенос ДНК (генов) из одной бактериальной клетки в другую,
- 14. Эксперименты с пневмококками Гриффита (1928 г.) и Эвери, Маклауда и Маккарти (1944 г.) Вирулентная S-форма Невирулентная
- 15. Трансфекция (transfection) [лат. trans — сквозь, через, за и inficere — портить, заражать] — 1) в
- 16. Искусственный перенос генов в целые организмы и клетки (трансфекция) – начальный этап Бенуа (1957 г.) –
- 17. Методы, используемые для транфекции эукариотических клеток Кальций-фосфатный метод (Грехем, Ван дер Еб, 1973) Декстрановый метод ДМСО-метод
- 18. Липофекция
- 19. Электропорация
- 20. Баллистическая трансфекция клеток
- 21. Импалефекция – метод доставки генов с применением наноматериалов (углеродные нанотрубки и нановолокна).
- 22. Микроинъекция
- 23. Перенос ранних зародышей от донора реципиенту 1890 г. – Уолтор Хип – первый успешный перенос ранних
- 24. Культивирование эмбрионов in vitro 1958 – Мак Ларен и Бриггс – культивирование эмбрионов мыши со стадии
- 25. Зигота человека Зигота свиньи
- 26. Перенос эмбрионов в воронку яйцевода свиньи
- 27. 1-й день – выделение яйцеклеток из фолликулов яичников шприцом с иглой (в каждом яичнике от 10
- 28. Химера (chimera, chimaera) [греч. Chimaira — мифическое чудовище, имеющее голову льва, туловище козы и хвост дракона]
- 29. Виды химеризма Первичный химеризм: - искусственная комбинация клеток зародышей предимплатационных стадий, - спонтанное оплодотворение сперматозоидами яйцеклетки
- 30. Агрегационные химеры А. Тарковский, 1961, Б. Минц, 1962 1 2 3 4 5 6 Ограничение: от
- 31. Вид бластоцисты под электронным микроскопом
- 32. Инъекционные химеры (Гарднер, 1968) Преимущество: возможно формирование химер из разных видов организмов
- 34. Овцекоза (1984 г.) Fehilly, C. B. et al.
- 35. Химерные макаки как результат агрегации бластомеров (Ш. Миталипов и др., 2012 г.) Потомки 12-ти родителей
- 36. ВЫВОДЫ: ЭСК и ВКМ у приматов не могут образовывать химеры; это способны делать 3-6 делящихся эмбрионов
- 37. Главный вывод: Культивированные in vitro эмбриональные стволовые клетки не обладают полностью свойствами стволовых клеток, существующих в
- 38. Новый вариант получения химер - оптотрансфекция
- 39. Распределение тканей в различных типах химер
- 40. Что можно выяснять с помощью метода создания химер? Используя две клеточные популяции, можно проследить в онтогенезе
- 41. Примеры использования химер для решения проблем онтогенеза Тарковский: наличие в зачатке гонады клеток мужского пола предотвращают
- 42. Стволовые клетки
- 43. Тотипотентность (totipotency) [лат. totus — весь, целый и potentia — сила] — способность клетки дифференцироваться в
- 44. Плюри- или тотипотентны эмбриональные стволовые клетки?
- 46. Скачать презентацию

Существует мнение:
Все, что сделано ранее и делается сегодня – редукционистская биология.
Надо
Существует мнение:
Все, что сделано ранее и делается сегодня – редукционистская биология.
Надо

Системная биология (systems biology) — междисциплинарная наука о жизни, изучающая сложные
Системная биология (systems biology) — междисциплинарная наука о жизни, изучающая сложные

Компьютерная модель полного жизненного цикла патогена Mycoplasma genitalium (2012 г.)
Идея:
Компьютерная модель полного жизненного цикла патогена Mycoplasma genitalium (2012 г.)
Идея:

Однако:
Без достижений редукционистской биологии невозможно создать «системную биологию».
Однако:
Без достижений редукционистской биологии невозможно создать «системную биологию».

2006 – Э. Файер и К. Мелло «за открытие РНК-интерференции — эффекта
2006 – Э. Файер и К. Мелло «за открытие РНК-интерференции — эффекта

Утром белки-активаторы BMal1 и Clock переходят из цитоплазмы в ядро клетки и там
Утром белки-активаторы BMal1 и Clock переходят из цитоплазмы в ядро клетки и там

Генная инженерия in vitro
Векторы (плазмиды, вирусы, фаги и др.)
Ферменты (рестриктазы, лигазы,
Генная инженерия in vitro
Векторы (плазмиды, вирусы, фаги и др.)
Ферменты (рестриктазы, лигазы,

культивирование соматических клеток животных in vitro
(конец 50-х годов XX века,
культивирование соматических клеток животных in vitro
(конец 50-х годов XX века,

Селективная среда ГAT (НАТ)
(гипоксантин, аминоптерин и тимидин)
1) Основной путь синтеза
Селективная среда ГAT (НАТ)
(гипоксантин, аминоптерин и тимидин)
1) Основной путь синтеза
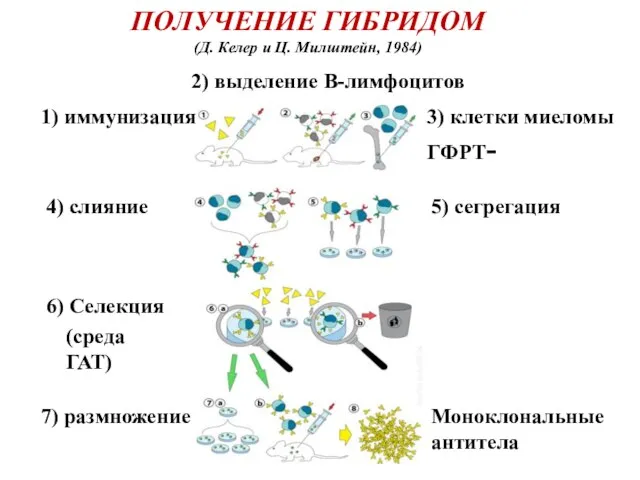
1) иммунизация
2) выделение В-лимфоцитов
3) клетки миеломы ГФРТ-
4) слияние
5) сегрегация
6) Селекция
7) размножение
1) иммунизация
2) выделение В-лимфоцитов
3) клетки миеломы ГФРТ-
4) слияние
5) сегрегация
6) Селекция
7) размножение
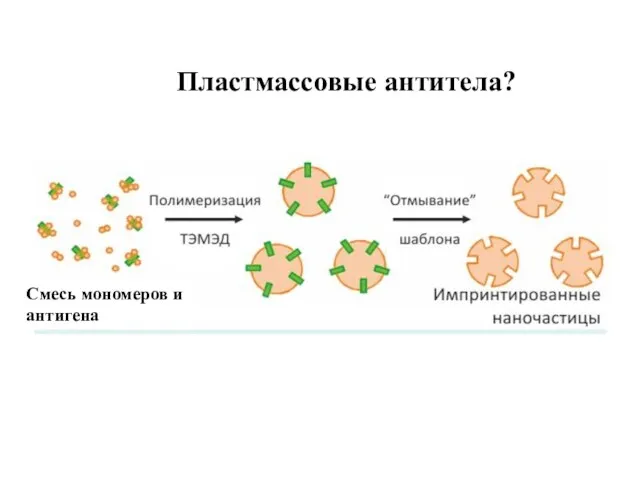
Пластмассовые антитела?
Смесь мономеров и антигена
Пластмассовые антитела?
Смесь мономеров и антигена
![Трансдукция (transduction) [лат. transductio — перемещение] — перенос ДНК (генов)](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/433665/slide-12.jpg)
Трансдукция (transduction) [лат. transductio — перемещение] — перенос ДНК (генов) из
Трансдукция (transduction) [лат. transductio — перемещение] — перенос ДНК (генов) из
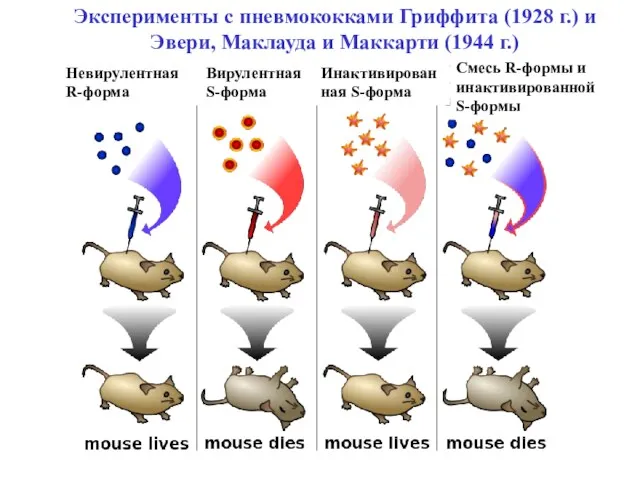
Эксперименты с пневмококками Гриффита (1928 г.) и Эвери, Маклауда и Маккарти
Эксперименты с пневмококками Гриффита (1928 г.) и Эвери, Маклауда и Маккарти

Трансфекция (transfection) [лат. trans — сквозь, через, за и inficere —
Трансфекция (transfection) [лат. trans — сквозь, через, за и inficere —

Искусственный перенос генов в целые организмы и клетки (трансфекция) – начальный
Искусственный перенос генов в целые организмы и клетки (трансфекция) – начальный

Методы, используемые для транфекции эукариотических клеток
Кальций-фосфатный метод
(Грехем, Ван дер
Методы, используемые для транфекции эукариотических клеток
Кальций-фосфатный метод
(Грехем, Ван дер
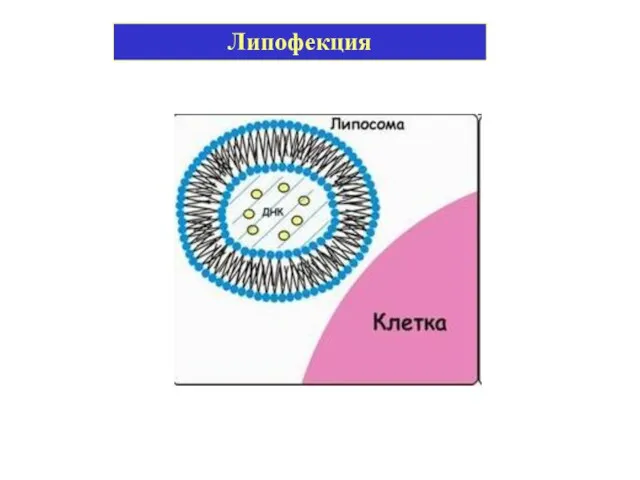
Липофекция
Липофекция
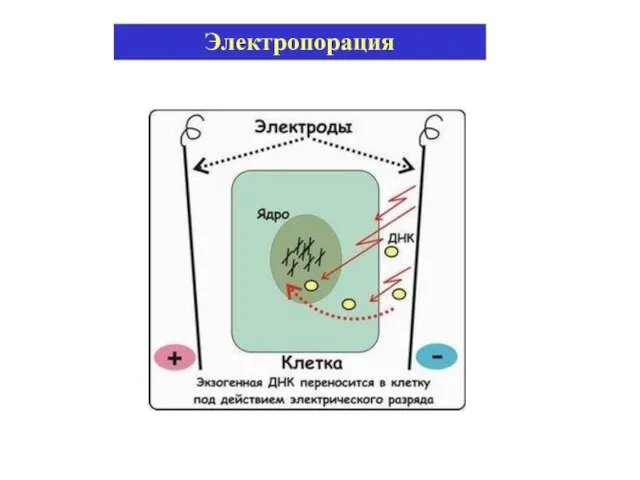
Электропорация
Электропорация
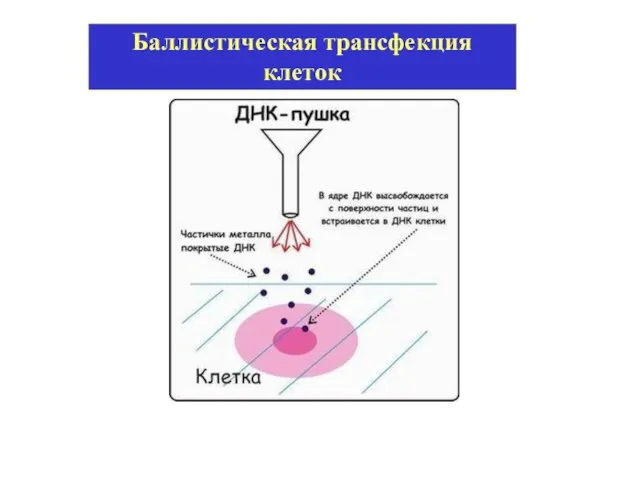
Баллистическая трансфекция клеток
Баллистическая трансфекция клеток

Импалефекция – метод доставки генов с применением наноматериалов (углеродные нанотрубки и
Импалефекция – метод доставки генов с применением наноматериалов (углеродные нанотрубки и
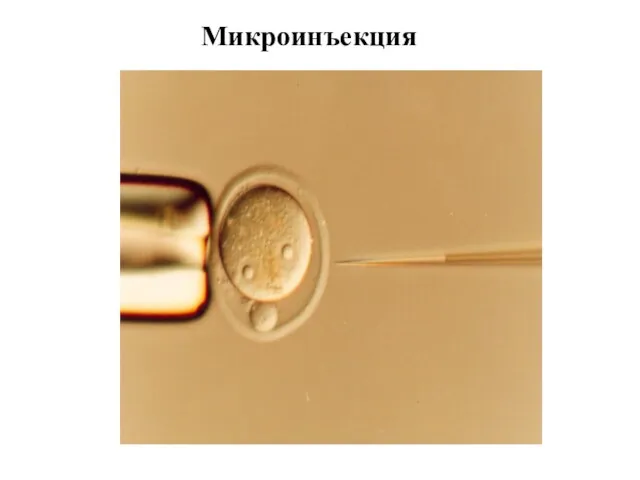
Микроинъекция
Микроинъекция

Перенос ранних зародышей от донора реципиенту
1890 г. – Уолтор Хип –
Перенос ранних зародышей от донора реципиенту
1890 г. – Уолтор Хип –

Культивирование эмбрионов in vitro
1958 – Мак Ларен и Бриггс – культивирование
Культивирование эмбрионов in vitro
1958 – Мак Ларен и Бриггс – культивирование
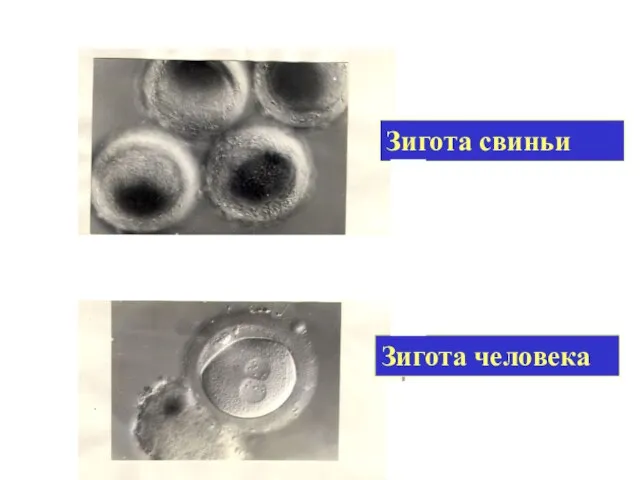
Зигота человека
Зигота свиньи
Зигота человека
Зигота свиньи
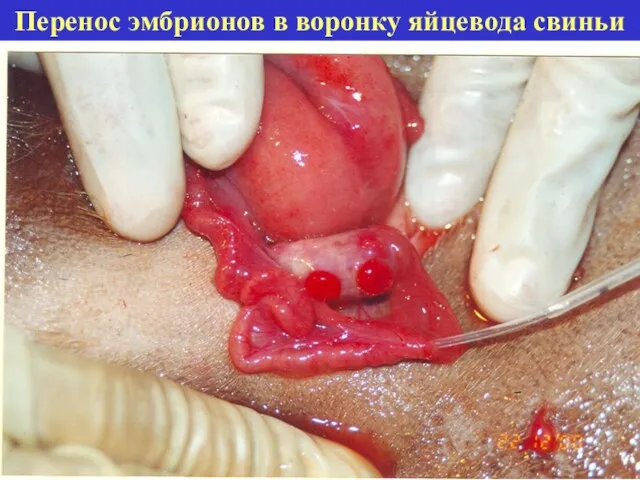
Перенос эмбрионов в воронку яйцевода свиньи
Перенос эмбрионов в воронку яйцевода свиньи

1-й день – выделение яйцеклеток из фолликулов яичников шприцом с иглой
1-й день – выделение яйцеклеток из фолликулов яичников шприцом с иглой

Химера (chimera, chimaera) [греч. Chimaira — мифическое чудовище, имеющее голову льва,
Химера (chimera, chimaera) [греч. Chimaira — мифическое чудовище, имеющее голову льва,

Виды химеризма
Первичный химеризм: - искусственная комбинация клеток зародышей предимплатационных стадий,
Виды химеризма
Первичный химеризм: - искусственная комбинация клеток зародышей предимплатационных стадий,
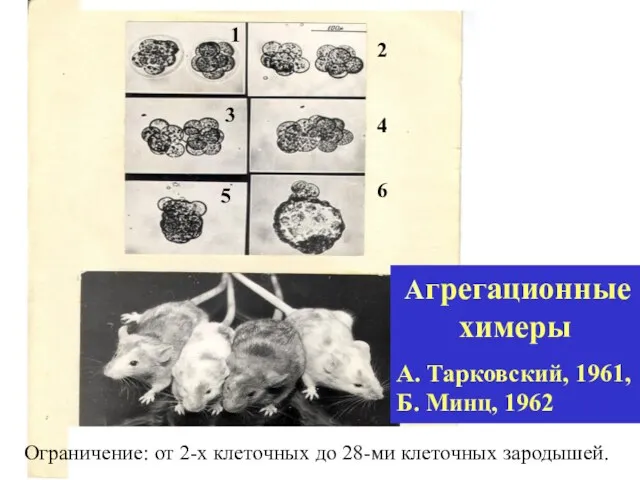
Агрегационные химеры
А. Тарковский, 1961, Б. Минц, 1962
1
2
3
4
5
6
Ограничение: от 2-х клеточных
Агрегационные химеры
А. Тарковский, 1961, Б. Минц, 1962
1
2
3
4
5
6
Ограничение: от 2-х клеточных
Вид бластоцисты под электронным микроскопом
Вид бластоцисты под электронным микроскопом
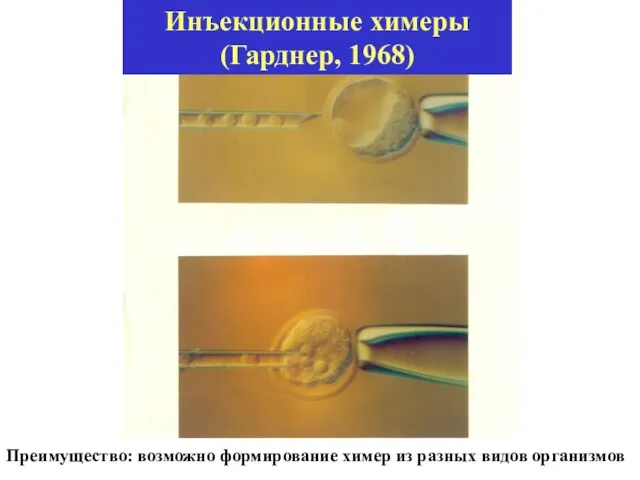
Инъекционные химеры (Гарднер, 1968)
Преимущество: возможно формирование химер из разных видов организмов
Инъекционные химеры (Гарднер, 1968)
Преимущество: возможно формирование химер из разных видов организмов

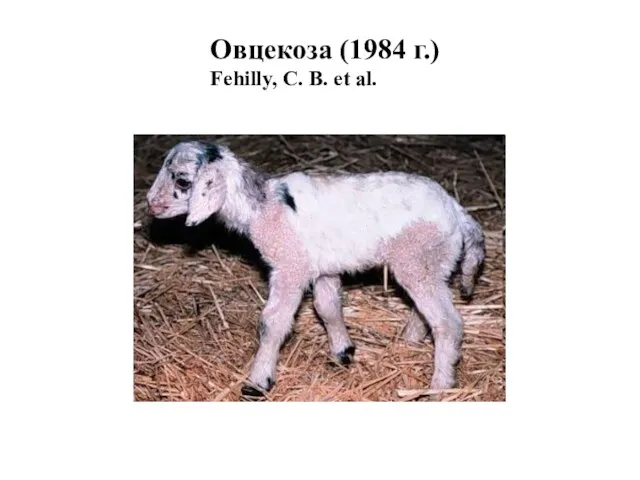
Овцекоза (1984 г.)
Fehilly, C. B. et al.
Овцекоза (1984 г.)
Fehilly, C. B. et al.

Химерные макаки как результат агрегации бластомеров
(Ш. Миталипов и др., 2012 г.)
Потомки
Химерные макаки как результат агрегации бластомеров
(Ш. Миталипов и др., 2012 г.)
Потомки
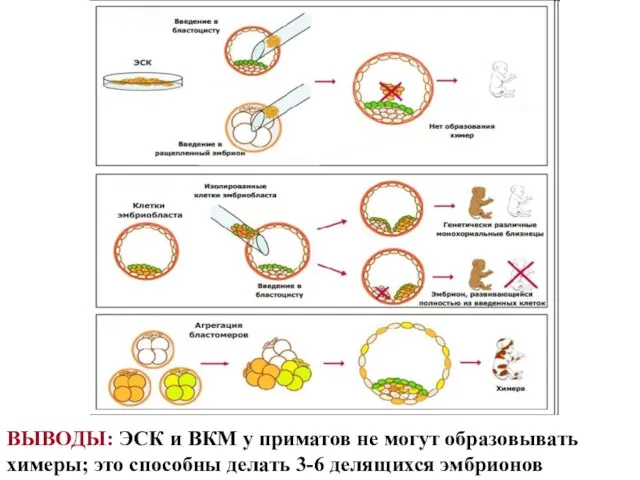
ВЫВОДЫ: ЭСК и ВКМ у приматов не могут образовывать химеры; это
ВЫВОДЫ: ЭСК и ВКМ у приматов не могут образовывать химеры; это

Главный вывод:
Культивированные in vitro эмбриональные стволовые клетки не обладают полностью свойствами
Главный вывод:
Культивированные in vitro эмбриональные стволовые клетки не обладают полностью свойствами
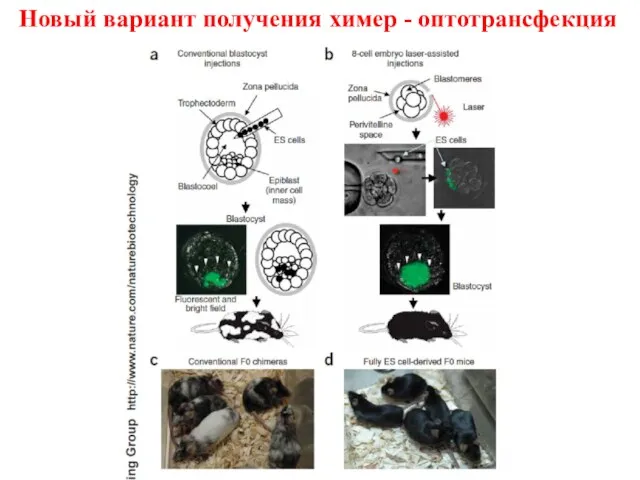
Новый вариант получения химер - оптотрансфекция
Новый вариант получения химер - оптотрансфекция
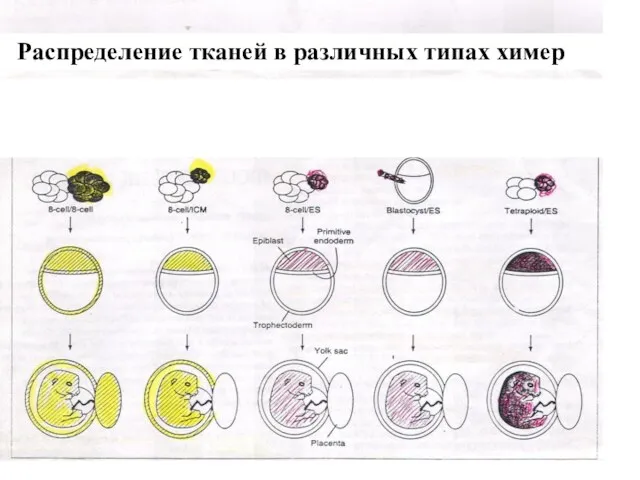
Распределение тканей в различных типах химер
Распределение тканей в различных типах химер

Что можно выяснять с помощью метода создания химер?
Используя две клеточные
Что можно выяснять с помощью метода создания химер?
Используя две клеточные

Примеры использования химер для решения проблем онтогенеза
Тарковский: наличие в зачатке гонады
Примеры использования химер для решения проблем онтогенеза
Тарковский: наличие в зачатке гонады

Стволовые клетки
Стволовые клетки

Тотипотентность (totipotency) [лат. totus — весь, целый и potentia — сила]
Тотипотентность (totipotency) [лат. totus — весь, целый и potentia — сила]

Плюри- или тотипотентны эмбриональные стволовые клетки?
Плюри- или тотипотентны эмбриональные стволовые клетки?
 Тип хордовые
Тип хордовые Клетка.10 класс
Клетка.10 класс Презентация к уроку биологии 7 класс Круглые черви
Презентация к уроку биологии 7 класс Круглые черви Презентация к уроку на тему Видоизменение побегов в 6 классе
Презентация к уроку на тему Видоизменение побегов в 6 классе Многолетние бобовые травы. Клевер луговой, гибридный, ползучий
Многолетние бобовые травы. Клевер луговой, гибридный, ползучий Зарождение человека. Процесс оплодотворения. Эмбриональное развитие плода
Зарождение человека. Процесс оплодотворения. Эмбриональное развитие плода Царство Бактерии
Царство Бактерии Лист – вегетативный орган растения
Лист – вегетативный орган растения Алоэ өсімдігінің шипалық қасиеті
Алоэ өсімдігінің шипалық қасиеті Раздел анатомии спланхнология. (Лекция 1)
Раздел анатомии спланхнология. (Лекция 1) Фрукти і овочі: характеристика, вимоги до якості, умови зберігання
Фрукти і овочі: характеристика, вимоги до якості, умови зберігання Sistemul nervos
Sistemul nervos Презентация Гигиена органов пищеварения
Презентация Гигиена органов пищеварения Ғарыш және биологиялық ырғақ
Ғарыш және биологиялық ырғақ Строение, свойства и функции углеводов и липидов в клетке
Строение, свойства и функции углеводов и липидов в клетке Генна інженерія та її досягнення
Генна інженерія та її досягнення Ядро клетки. Хромосомный набор клетки
Ядро клетки. Хромосомный набор клетки туберкулез
туберкулез Испарение воды растениями
Испарение воды растениями Тренажёр Вода
Тренажёр Вода Методическая копилка
Методическая копилка Три среды обитания организмов
Три среды обитания организмов Азотное питание растений
Азотное питание растений Культивирование бактерий
Культивирование бактерий Семейства кошачих. Домашние кошки, тигры
Семейства кошачих. Домашние кошки, тигры Класс Двудольные. Семейство Крестоцветные
Класс Двудольные. Семейство Крестоцветные Life cycle assessment
Life cycle assessment Биологические ритмы человека. Сон и его значение
Биологические ритмы человека. Сон и его значение