- Морфология и структура вирусов

Содержание
- 2. Вирусы – микроорганизмы, составляющие царство Vira. Отличительные признаки: 1) содержат лишь один тип нуклеиновой кислоты (РНК
- 3. Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса). По форме вирионы могут быть:
- 4. В центре вириона – вирусная нуклеиновая кислота, покрытая белковой оболочкой – капсидом, который имеет строго упорядоченную
- 5. ДНК может быть: 1) двухцепочечной; 2) одноцепочечной; 3) кольцевой; 4) двухцепочечной, но с одной более короткой
- 6. Вирусные белки подразделяют на: 1) геномные – нуклеопротеиды. Обеспечивают репликацию вирусных нуклеиновых кислот и процессы репродукции
- 7. 3) белки суперкапсидной оболочки – это сложные белки, разнообразные по функции. За счет них происходит взаимодействие
- 8. 2. Взаимодействие вирусов с клеткой хозяина Взаимодействие идет в единой биологической системе на генетическом уровне. Существует
- 9. После адсорбции вирионы проникают внутрь путем эндоцитоза (виропексиса) или в результате слияния вирусной и клеточной мембран.
- 10. Нуклеиновые кислоты вирусов реализуют генетическую программу по созданию вирусного потомства и определяют наследственные свойства вирусов. С
- 11. После того как в зараженной клетке накопится достаточное количество компонентов вируса, начинается сборка вирионов потомства. Процесс
- 12. Последним этапом взаимодействия вирусов с клетками является выход или освобождение из клетки дочерних вирусных частиц. Простые
- 13. 3. Культивирование вирусов Основные методы культивирования вирусов: 1) биологический – заражение лабораторных животных. При заражении вирусом
- 14. В результате заражения могут происходить и появляться: 1) гибель эмбриона; 2) дефекты развития: на поверхности оболочек
- 15. Для поддержания клеток культуры ткани используют специальные среды. Это жидкие питательные среды сложного состава, содержащие аминокислоты,
- 16. Основные проявления цитопатического действия вирусов: 1) размножение вируса может сопровождаться гибелью клеток или морфологическими изменениями в
- 17. 4. Особенности противовирусного иммунитета Противовирусный иммунитет начинается со стадии презентации вирусного антигена Т-хелперами. Сильными антигенпрезентирующими свойствами
- 18. 1) нейтрализацию вируса антителами; это препятствует рецепции вируса клеткой и проникновению его внутрь. Опсонизация вируса с
- 19. Недостаточная концентрация антител может усиливать репродукцию вируса. Иногда антитела могут защищать вирус от действия протеолитических ферментов
- 20. Когда вирусы переходят из клетки в клетку по цитоплазматическим мостикам, не контактируя с циркулирующими антителами, то
- 21. Механизм цитотоксического эффекта связан с активацией мембранных ферментных систем в зоне прилипания клеток, образованием цитоплазматических мостиков
- 23. Скачать презентацию

Вирусы – микроорганизмы, составляющие царство Vira.
Отличительные признаки:
1) содержат лишь один тип нуклеиновой
Вирусы – микроорганизмы, составляющие царство Vira.
Отличительные признаки:
1) содержат лишь один тип нуклеиновой

Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса).
По
Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса).
По

В центре вириона – вирусная нуклеиновая кислота, покрытая белковой оболочкой –
В центре вириона – вирусная нуклеиновая кислота, покрытая белковой оболочкой –

ДНК может быть:
1) двухцепочечной;
2) одноцепочечной;
3) кольцевой;
4) двухцепочечной, но с одной более короткой цепью;
5) двухцепочечной, но с
ДНК может быть:
1) двухцепочечной;
2) одноцепочечной;
3) кольцевой;
4) двухцепочечной, но с одной более короткой цепью;
5) двухцепочечной, но с

Вирусные белки подразделяют на:
1) геномные – нуклеопротеиды. Обеспечивают репликацию вирусных нуклеиновых кислот
Вирусные белки подразделяют на:
1) геномные – нуклеопротеиды. Обеспечивают репликацию вирусных нуклеиновых кислот

3) белки суперкапсидной оболочки – это сложные белки, разнообразные по функции. За
3) белки суперкапсидной оболочки – это сложные белки, разнообразные по функции. За

2. Взаимодействие вирусов с клеткой хозяина
Взаимодействие идет в единой биологической системе на
2. Взаимодействие вирусов с клеткой хозяина
Взаимодействие идет в единой биологической системе на

После адсорбции вирионы проникают внутрь путем эндоцитоза (виропексиса) или в результате
После адсорбции вирионы проникают внутрь путем эндоцитоза (виропексиса) или в результате

Нуклеиновые кислоты вирусов реализуют генетическую программу по созданию вирусного потомства и
Нуклеиновые кислоты вирусов реализуют генетическую программу по созданию вирусного потомства и

После того как в зараженной клетке накопится достаточное количество компонентов вируса,
После того как в зараженной клетке накопится достаточное количество компонентов вируса,

Последним этапом взаимодействия вирусов с клетками является выход или освобождение из
Последним этапом взаимодействия вирусов с клетками является выход или освобождение из

3. Культивирование вирусов
Основные методы культивирования вирусов:
1) биологический – заражение лабораторных животных. При заражении
3. Культивирование вирусов
Основные методы культивирования вирусов:
1) биологический – заражение лабораторных животных. При заражении

В результате заражения могут происходить и появляться:
1) гибель эмбриона;
2) дефекты развития: на поверхности
В результате заражения могут происходить и появляться:
1) гибель эмбриона;
2) дефекты развития: на поверхности

Для поддержания клеток культуры ткани используют специальные среды. Это жидкие питательные
Для поддержания клеток культуры ткани используют специальные среды. Это жидкие питательные

Основные проявления цитопатического действия вирусов:
1) размножение вируса может сопровождаться гибелью клеток или
Основные проявления цитопатического действия вирусов:
1) размножение вируса может сопровождаться гибелью клеток или

4. Особенности противовирусного иммунитета
Противовирусный иммунитет начинается со стадии презентации вирусного антигена Т-хелперами.
Сильными
4. Особенности противовирусного иммунитета
Противовирусный иммунитет начинается со стадии презентации вирусного антигена Т-хелперами.
Сильными

1) нейтрализацию вируса антителами; это препятствует рецепции вируса клеткой и проникновению его
1) нейтрализацию вируса антителами; это препятствует рецепции вируса клеткой и проникновению его

Недостаточная концентрация антител может усиливать репродукцию вируса. Иногда антитела могут защищать
Недостаточная концентрация антител может усиливать репродукцию вируса. Иногда антитела могут защищать

Когда вирусы переходят из клетки в клетку по цитоплазматическим мостикам, не
Когда вирусы переходят из клетки в клетку по цитоплазматическим мостикам, не

Механизм цитотоксического эффекта связан с активацией мембранных ферментных систем в зоне
Механизм цитотоксического эффекта связан с активацией мембранных ферментных систем в зоне
 Хвороби бджіл
Хвороби бджіл презентация Клетка
презентация Клетка Генетические и средовые детерминанты вызванных потенциалов
Генетические и средовые детерминанты вызванных потенциалов Элективный курс Биология и будущая профессия 9 класс
Элективный курс Биология и будущая профессия 9 класс Презентация по теме Жизнь, основные свойства живого Диск
Презентация по теме Жизнь, основные свойства живого Диск Выполнение открытки Рыжая красавица – лиса
Выполнение открытки Рыжая красавица – лиса Лекарственные растения
Лекарственные растения презентация Строение и работа сердца. Кровеносные сосуды
презентация Строение и работа сердца. Кровеносные сосуды газообмен в легких
газообмен в легких Ісіктер мен ағзаның иммунды биологиялық әрекеттесу ерекшеліктері
Ісіктер мен ағзаның иммунды биологиялық әрекеттесу ерекшеліктері Влияние спиртов на организм человека
Влияние спиртов на организм человека Амурский тигр - хищник из Красной книги
Амурский тигр - хищник из Красной книги Органы цветковых растений
Органы цветковых растений Мейоз. Значение мейоза
Мейоз. Значение мейоза История органического мира земли
История органического мира земли Физиология пищеварения
Физиология пищеварения Введение в биологию
Введение в биологию Зология беспозвоночных животных
Зология беспозвоночных животных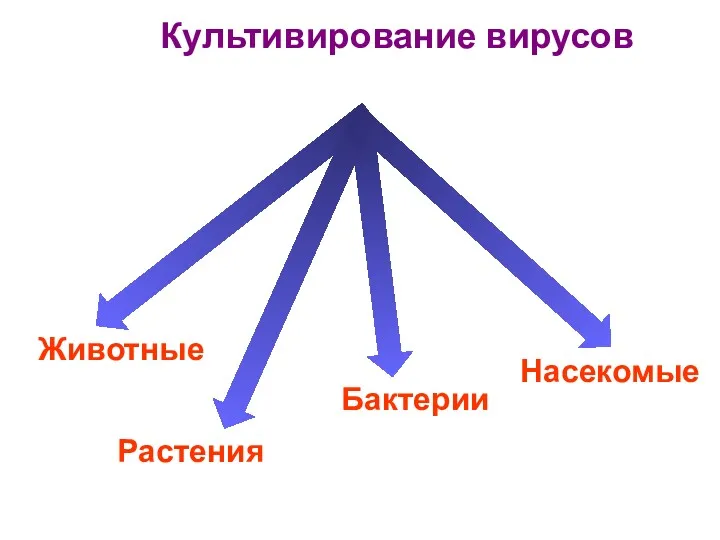 Культивирование вирусов
Культивирование вирусов Органические вещества клетки. Углеводы и белки
Органические вещества клетки. Углеводы и белки Кто в каком домике живёт. (Окружающий мир, 1 класс)
Кто в каком домике живёт. (Окружающий мир, 1 класс) Мир насекомых
Мир насекомых Строение семян однодольных и двудольных растений. Морфология проростка. Прорастание семян
Строение семян однодольных и двудольных растений. Морфология проростка. Прорастание семян Геоботаническое и культуртехническое обследование кормовых угодий
Геоботаническое и культуртехническое обследование кормовых угодий Морфология микроорганизмов. Лекция 2. Морфология бактерий
Морфология микроорганизмов. Лекция 2. Морфология бактерий Кістки лицевого і мозкового відділу черепа
Кістки лицевого і мозкового відділу черепа Органы чувств: глаза
Органы чувств: глаза Анализаторы. Органы чувств
Анализаторы. Органы чувств