- Направленность и пути макроэволюции. Лекция 3

Содержание
- 2. Проблема направленности эволюции Направленность - это проявления в ходе макроэволюции общих тенденций в организации эволюционных процессов.
- 3. Существуют 2 крайние точки зрения: 1. эволюционный оппортунизм (селекционизм, классический дарвинизм, СТЭ) - эволюция лишена устойчивости
- 5. 3. явление Инадаптации (от лат. in — приставка, означающая отрицание, и адаптация) совокупность несовершенных приспособлений, возникающая
- 6. В эволюции копытных млекопитающих — несовершенные изменения в скелете и суставах конечностей у большинства групп при
- 7. Инадаптивные формы появлялись раньше адаптивных, но затем вытеснялись последними. Так, плацентарные млекопитающие вытеснили сумчатых, сохранившихся в
- 8. 2. Номогенез - эволюция представляет запрограммированную изменчивость, а роль естественного отбора - выбраковка. Работы Л.С.Берга, А.А.Любищева
- 9. Аргументы номогенеза: 1. Повышение организации - магистральная линия эволюции, (запрограммированное направление)
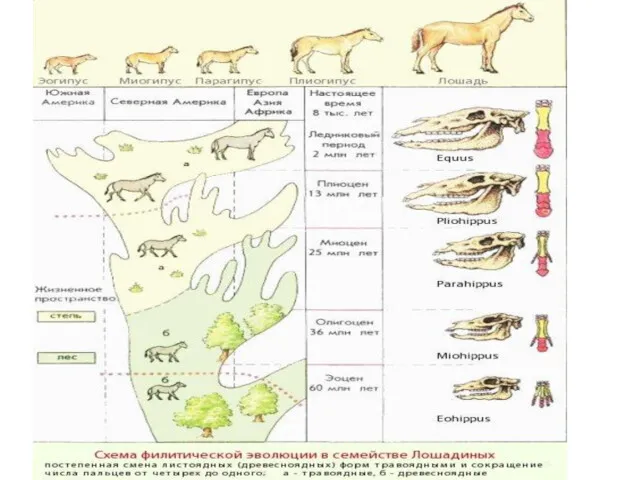
- 10. 2. Параллельная эволюция и филетические линии (лошадиные непарнокопытные - 55 млн. лет развитие в сходном направлении:
- 11. 3. Появление гипертрофированных органов, то есть сохранение эволюционной инерции, вопреки действию естественного отбора.
- 12. 1.Эволюционный оппортунизм (случайная изменчивость) 2. Номогенез (запрограммированная изменчивость) Абсолютизируют разные факторы направляющую роль отбора роль организационной
- 13. Два разных подхода к изучению эволюции: а).«элементаристский» - разложение целого на элементы (СТЭ, борьба за существование,
- 14. 3-я точка зрения - системный взгляд на направленность эволюции за счет взаимодействия живого как системы со
- 15. отсюда, существуют две категории направляющих факторов эволюции: 1.внеорганизменные - силы отбора соответствующие изменениям внешней среды 2.организменные
- 16. 2. Организменные факторы А) генетические корреляции некоторые мутации запрещены из-за отсутствия биохимических предпосылок (синие глаза у
- 17. Корреляции действуют через ключевой эволюционный запрет Он определяет характер адаптации в зависимых системах органов. Затем возникает
- 18. Комплекс преобразований челюстного аппарата у териодонтов был обусловлен ключевой адаптацией - совершенствованием переработки пищи челюстями.
- 19. Корреляции в исторически сложившихся системах Трахейная система насекомых как специализированное эффективное приспособление, наложила отпечаток на всю
- 20. Каскадные взаимодействия Редукция кровеносной системы невозможность снабжения массивных органов ограничение размеров тела; Особенности поведения и образа
- 21. Корреляции в исторически сложившихся системах . Способ вентиляции легких – кожное дыхание и строение легких земноводных
- 22. Каскадные взаимодействия При этом ороговение не имеет б/х и морфологических запретов, (могут появляться мутантные особи с
- 23. Если естественный отбор длительное время благоприятствовал развитию каких-либо адаптаций (лошадиные - бег по уплотненной почве, питание
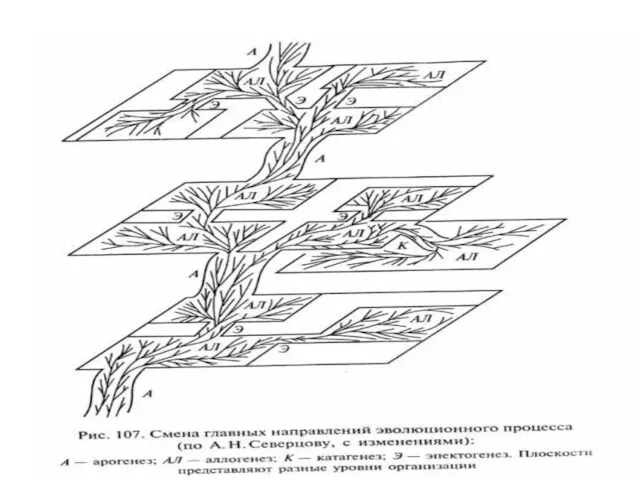
- 24. Путь направленной эволюции – арогенез Отбор благоприятствует ароморфозам, если они обеспечивают важные преимущества в борьбе за
- 25. Избыточность биосистем - прогнозирование будущего при цикличности повторяемости событий. Филогенез - прогнозная модель, т.к. разнообразные группы.
- 26. Канализирующая роль строения организма. Адаптивный компромисс. Это означает ограничение эволюционных возможностей какими-то факторами и, как следствие,
- 27. Примеры метода «оптимизированного случайного тыка» 1. Работа иммунной системы. Антитела к новым возбудителям, с которыми организм
- 28. Законы композиции и роста V. Законы самоорганизации III. Взаимосвязанность онто-филогенеза VII. коэволюционность процессов в сообществе, экосистеме
- 29. Механизмы направленности эволюции I. Закон композиции и законы роста, связанные с системной организацией канализированность структурных преобразований.
- 30. Числа Фибоначчи на главном вокзале Цюриха Плотная пища жён Фибоначчи Только на пользу им шла, не
- 31. Одинаковые наборы типов листорасположения Формулы листорасположения - дроби, где цифры в числителе и знаменателе, соответствуют взятым
- 32. Геометрические классы спиралей = у разных моллюсков (хорошие пловцы) и фораминифер Сходство цветков соцветий
- 34. II. Законы формы (морфологии) Производные генетики, физиологии и экологии (генетические механизмы + функция + внешнее приспособление).
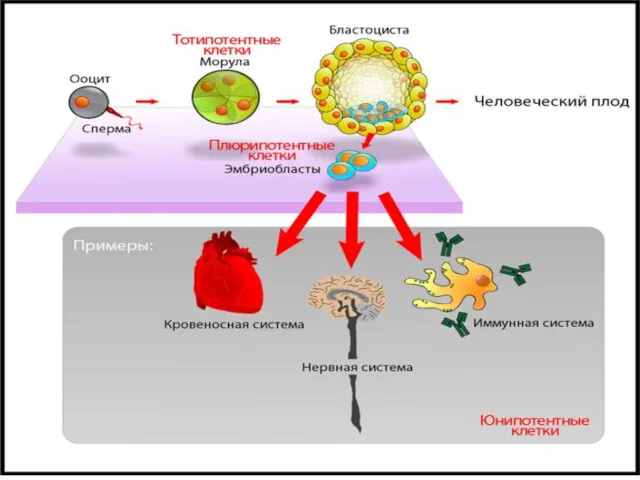
- 35. III. Взаимосвязанность онто-филогенеза А.) Новую линию в эволюции могут дать мутации большой амплитуды, проявляющимися на ранних
- 36. Б). Стабилизация онтогенеза (несмотря на присущую ему стохастику), за счет специальных адаптаций - системы повышения помехоустойчивости.
- 37. центральный фрагмент генной сети биосинтеза холестерина в клетке (регуляция по механизму отрицательной обратной связи) Принципиальная схема
- 40. Пути передачи сигналов: МАР-киназный путь передачи сигнала в ядро клетки, активируемый ростовыми факторами, контролирующий процесс клеточного
- 41. Важную роль в стабилизации онтогенеза может играть «избыточность» генно-регуляторных сетей, управляющих развитием. Благодаря избыточности ГРК выход
- 43. г). Эпигенетическая изменчивость (М. А. Шишкин). Сильное внешнее воздействие (например, резкое изменение среды) приводит к дестабилизации
- 45. Эпигенетическая теория эволюции рассматривает один из возможных механизмов наследования приобретенных свойств – генетическую ассимиляцию морфозов. Т.е.
- 46. Морфозы в широком смысле, т. е. изменение фенотипа без изменения генома, может вести к смене направленности
- 47. Когда развитие куриного эмбриона достигает определенного этапа, клетки кожи, соединяющей зачатки пальцев, гибнут. Это не происходит
- 48. Основополагающее влияние на наследственность оказывает не геном, а эпигенетическая система (ЭС) – совокупность факторов, воздействующих на
- 49. Эволюция по ЭТЭ (эпигенетическая теория эволюции) заключается в преобразовании одного креода в другой при возмущающем воздействии
- 50. Участок эпигенетического ландшафта, показывающий перепады высоты стенок (порогов устойчивости) креода в местах ответвления аберративных долин. Онтогенез
- 51. Дестабилизация развития может существенно облегчать формирование новых адаптаций. Дестабилизация «перекидывает мостик» от одного стабильного состояния к
- 52. VI. Термодинамическая направленность эволюции определяемая законом минимизации диссипации энергии при функционировании открытых систем Эволюция направлена на
- 53. фенотипическая устойчивость реализуется в потенциальной яме пространства возможных состояний + приспособленность к экологической нише
- 54. IV. Законы самоорганизации Эволюция сложных природных неравновесных систем рассматривается как процесс самоорганизации. Самоорганизация означает образование в
- 55. Точки бифуркации
- 56. В процессе эволюции система переходит в новое состояние. Целое задает правила поведения своим частям. Системы стремятся
- 57. Поведение системы в состоянии неустойчивости, может зависеть от поведения очень немногих переменных. Эти факторы называются параметрами
- 59. Параметры порядка играют доминирующую роль в концепции синергетики. Они «подчиняют» отдельные части, т. е. определяют поведение
- 61. В процессе развития природных систем также можно выделить несколько главных переменных, т.е параметров порядка, к которым
- 62. При комплексном исследовании почв территории параметр порядка - рельеф, При исследовании поверхностных водных источников – геометрия
- 63. Для микро-, мезо-(или макро-) и мега- уровня параметры порядка - это долгоживущие коллективные переменные, задающие язык
- 64. Управляющий параметр может быть один и несколько. Их количество фиксировано и налагается на систему извне. Управляющие
- 68. Управляющий параметр выполняет роль триггера, запускающего сложные сетевые процессы. Он включает каскады взаимодействий (параметров порядка). Взаимосвязь
- 69. Ю.Л. Климонтович, изучая процессы самоорганизации, в 1983 году доказал S-теорему, которая гласит, что по мере удаления
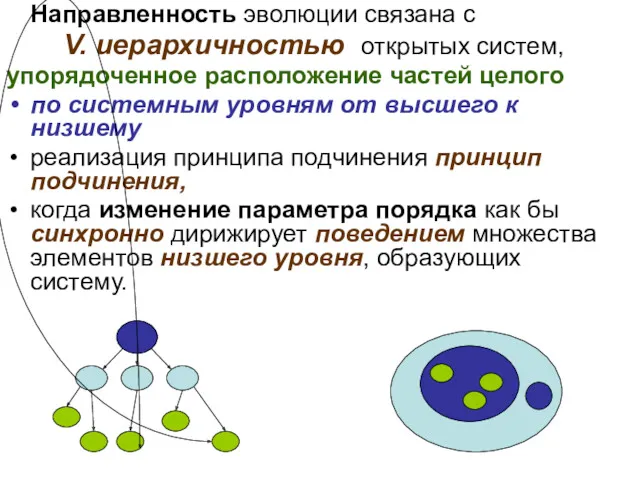
- 70. Направленность эволюции связана с V. иерархичностью открытых систем, упорядоченное расположение частей целого по системным уровням от
- 71. Особую роль в иерархии систем играет время, и синергетический принцип подчинения Хакена формулируется именно для временной
- 72. Развитие выступает как направленный процесс количественных и качественных изменений в структуре объектов, определяемых их последовательным включением
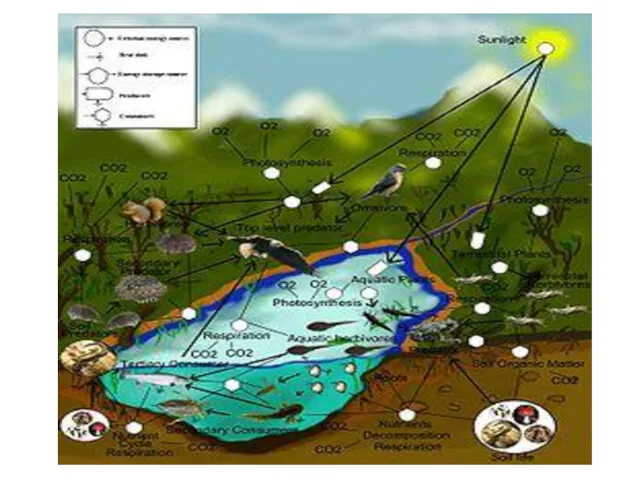
- 73. Направленность эволюции связана с VII. коэволюционностью процессов от сообщества до биосферы (Коэволюция - это эволюционное взаимодействие
- 74. А). Взаимодействие видов через экологические ниши. - С одной стороны, усиливает эволюционный процесс. Молодые э/с с
- 75. В развитом сообществе все возможные ниши обычно заняты, и новым видам просто некуда втиснуться. Если же
- 76. Вхождение вида в сообщество может быть в роли доминанта, а в других случаях - второстепенного вида.
- 77. Каждая экосистема предоставляет организмам, живущим в ней или вселяющимся экологическую лицензию, то есть место в потоках
- 78. Поразительные примеры параллельной эволюции наблюдались в гибнущем Аральском море, который разделился на два изолированных, быстро высыхающих
- 79. Массовое вымирание освободило множество ниш. Например, вымерли все моллюски-фильтраторы (тогда как многие зарывающиеся грунтоеды уцелели). Пустующие
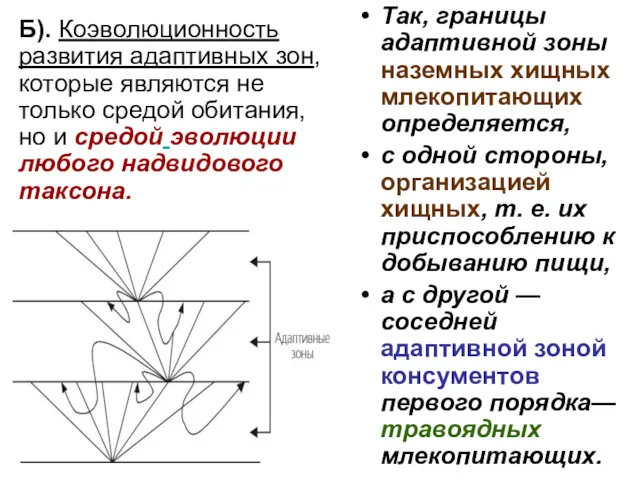
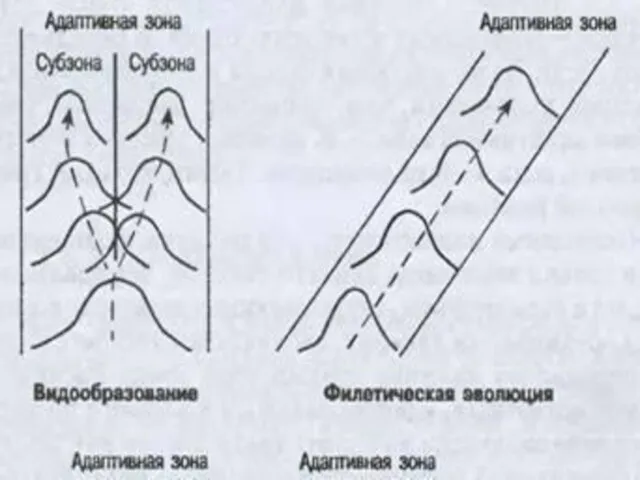
- 80. Б). Коэволюционность развития адаптивных зон, которые являются не только средой обитания, но и средой эволюции любого
- 81. Т.о. понятие адаптивной зоны подразумевает не только эволюцию данного таксона, определяемую средой его обитания, но и
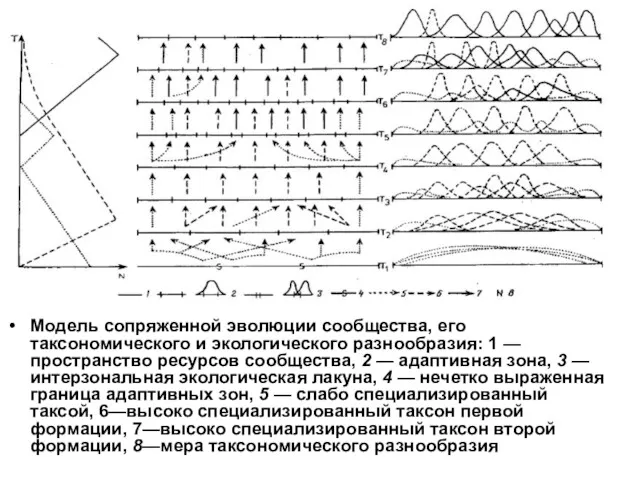
- 83. Модель сопряженной эволюции сообщества, его таксономического и экологического разнообразия: 1 — пространство ресурсов сообщества, 2 —
- 84. В). В экосистеме сохранение вида, его ниши происходит за счет непрерывной адаптивной эволюция (колеблющееся равновесие между
- 85. 2). Направленность эволюции групп связана с эволюцией самих биогеоценозов как систем. Это согласованная "когерентная" эволюция (Красилов)
- 86. Кибернетическая схема зависимости эволюционного процесса (Шмальгаузен И.И.)
- 87. Пьер Тейя́р де Шарде́н (1881 -1955) — французский теолог, философ, священник-иезуит, один из создателей теории ноосферы.
- 88. Эволюция - это возрастание функциональной сложности параллельно росту специфичности, индивидуализации, дискретности. Эволюция сочетает: радиальную энергию (повышение
- 89. Будущее открыто и непредсказуемо, но оно не произвольно – т.к. существуют спектры возможных будущих состояний в
- 90. Эволюция, по всей видимости, – процесс в целом и в самом главном детерминированный, но в деталях
- 91. Основные пути эволюции Прогресс и регресс Прогресс выражается в подъеме общей организации и энергии жизнедеятельности, но
- 92. Формы прогрессивного развития: 1. Неограниченный, 2. . Морфофизиологический, 3. Биологический, 4. Групповой, 5. Биотехнический.
- 93. Неограниченный прогресс: от прокариот – до человека, от индивидуума – к социальности
- 94. 2. Морфофизиологический прогресс Усложнение организации группы Проблема выделения критериев: 1. наличие ароморфозов, крупные изменения в плане
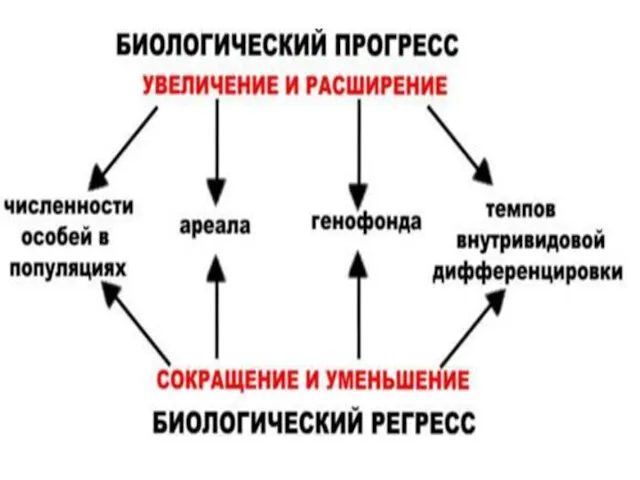
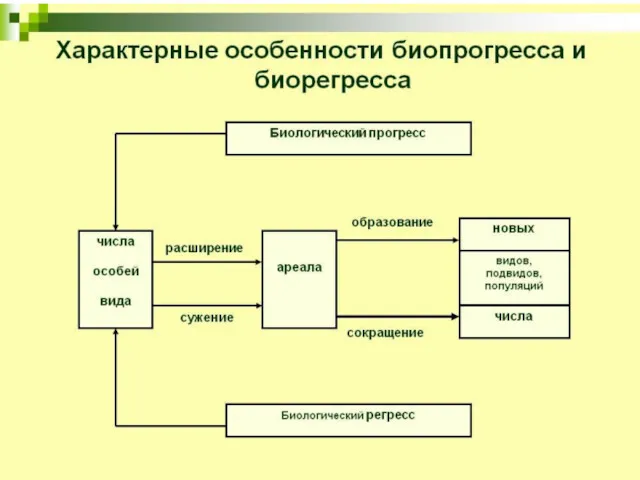
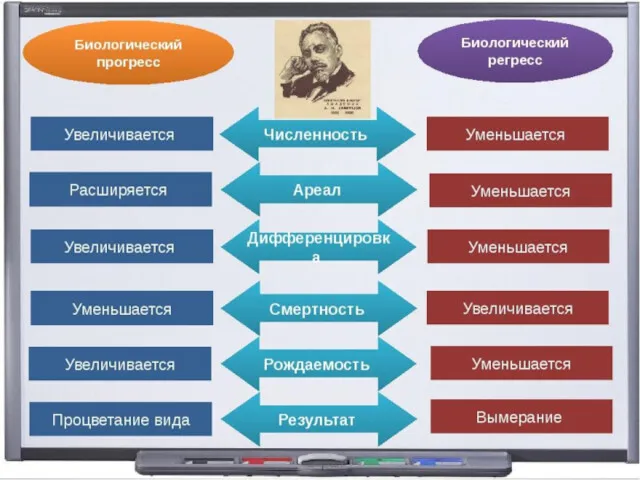
- 95. 3. Биологический прогресс Процветание группы : увеличение численности, расширение ареала, высокая скорость видообразования. Может достигаться разными
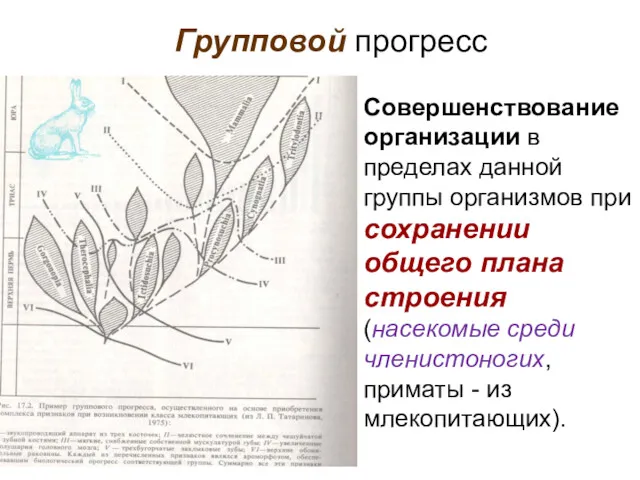
- 99. Групповой прогресс Совершенствование организации в пределах данной группы организмов при сохранении общего плана строения (насекомые среди
- 100. Биотехнический прогресс - большая эффективность выполнения функций у некоторых групп. Основной критерий - энергетические показатели, КПД
- 101. Критерии прогресса Повышение общей Е жизнедеятельности (Северцев). Совершенствование системы регуляции (Грант). Регуляция управления, основанного на термодинамических
- 102. Биологический регресс Признаки обратные прогрессу: уменьшение численности, сужение ареала, снижение скорости видообразования. Может привести к вымиранию.
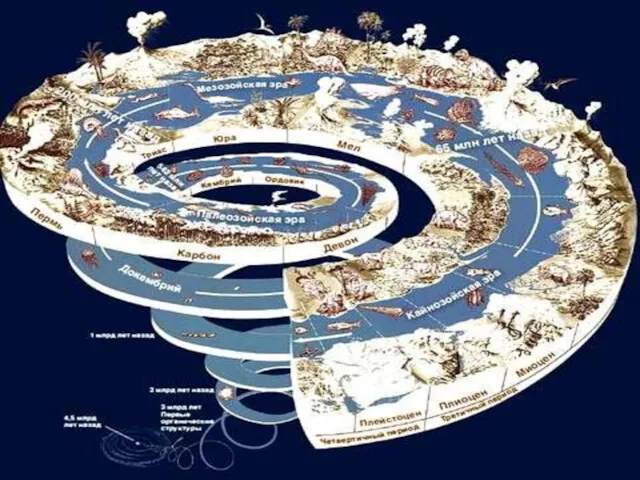
- 107. На границе перми и триаса, то есть палеозойской и мезозойской эр, произошло массовое вымирание во многих
- 110. 440 млн лет назад — ордовикско-силурийское вымирание — исчезло более 60 % видов морских беспозвоночных; 364
- 114. Достигнутые эволюционной биологией на настоящий момент рубежи отличаются от представленных ранее постулатов СТЭ: 1.Постулат о популяции
- 116. Скачать презентацию

Проблема направленности эволюции
Направленность
- это проявления в ходе макроэволюции общих тенденций
Проблема направленности эволюции
Направленность
- это проявления в ходе макроэволюции общих тенденций

Существуют 2 крайние точки зрения:
1. эволюционный оппортунизм (селекционизм, классический дарвинизм,
Существуют 2 крайние точки зрения:
1. эволюционный оппортунизм (селекционизм, классический дарвинизм,

3. явление Инадаптации
(от лат. in — приставка, означающая отрицание, и адаптация)
3. явление Инадаптации
(от лат. in — приставка, означающая отрицание, и адаптация)

В эволюции копытных млекопитающих — несовершенные изменения
в скелете и суставах
В эволюции копытных млекопитающих — несовершенные изменения
в скелете и суставах

Инадаптивные формы появлялись раньше адаптивных,
но затем вытеснялись последними.
Так, плацентарные
Инадаптивные формы появлялись раньше адаптивных,
но затем вытеснялись последними.
Так, плацентарные

2. Номогенез - эволюция представляет запрограммированную изменчивость,
а роль естественного отбора
2. Номогенез - эволюция представляет запрограммированную изменчивость,
а роль естественного отбора

Аргументы номогенеза:
1. Повышение организации - магистральная линия эволюции,
(запрограммированное направление)
Аргументы номогенеза:
1. Повышение организации - магистральная линия эволюции,
(запрограммированное направление)

2. Параллельная эволюция и филетические линии (лошадиные непарнокопытные - 55 млн.
2. Параллельная эволюция и филетические линии (лошадиные непарнокопытные - 55 млн.

3. Появление гипертрофированных органов, то есть сохранение эволюционной инерции,
вопреки действию
3. Появление гипертрофированных органов, то есть сохранение эволюционной инерции,
вопреки действию
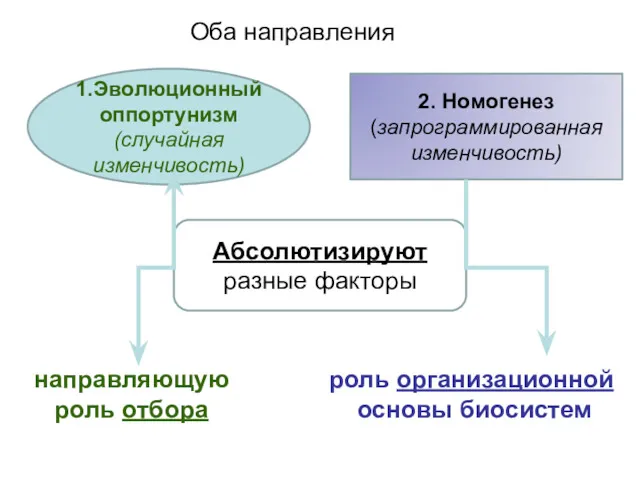
1.Эволюционный оппортунизм
(случайная изменчивость)
2. Номогенез (запрограммированная изменчивость)
Абсолютизируют разные факторы
направляющую
роль отбора
роль организационной
1.Эволюционный оппортунизм
(случайная изменчивость)
2. Номогенез (запрограммированная изменчивость)
Абсолютизируют разные факторы
направляющую
роль отбора
роль организационной

Два разных подхода к изучению эволюции: а).«элементаристский» - разложение целого
Два разных подхода к изучению эволюции: а).«элементаристский» - разложение целого
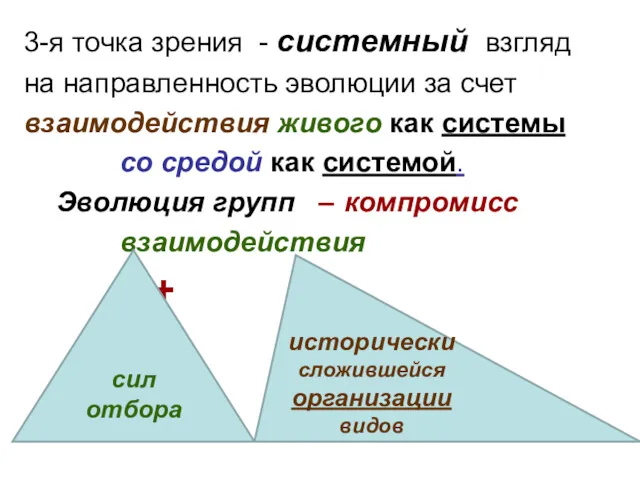
3-я точка зрения - системный взгляд
на направленность эволюции за счет
взаимодействия
3-я точка зрения - системный взгляд
на направленность эволюции за счет
взаимодействия

отсюда, существуют две категории направляющих факторов эволюции:
1.внеорганизменные
- силы отбора соответствующие
отсюда, существуют две категории направляющих факторов эволюции:
1.внеорганизменные
- силы отбора соответствующие

2. Организменные факторы
А) генетические корреляции
некоторые мутации запрещены из-за отсутствия
2. Организменные факторы
А) генетические корреляции
некоторые мутации запрещены из-за отсутствия

Корреляции действуют через ключевой эволюционный запрет
Он определяет характер адаптации в
Корреляции действуют через ключевой эволюционный запрет
Он определяет характер адаптации в
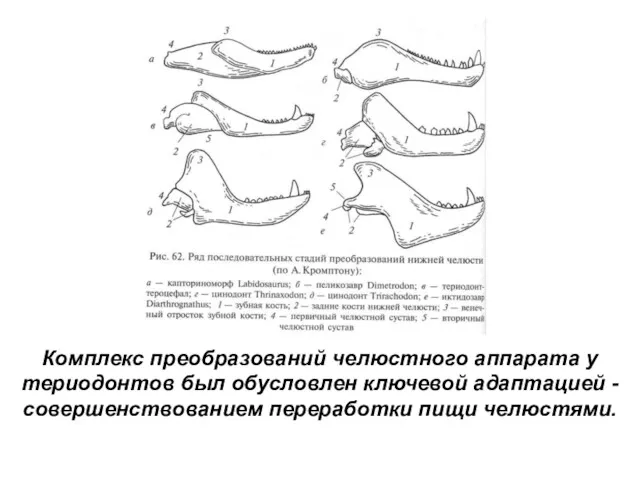
Комплекс преобразований челюстного аппарата у териодонтов был обусловлен ключевой адаптацией -
Комплекс преобразований челюстного аппарата у териодонтов был обусловлен ключевой адаптацией -
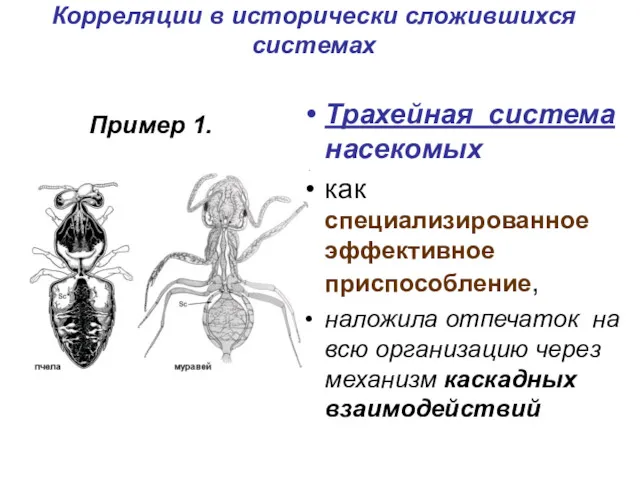
Корреляции в исторически сложившихся системах
Трахейная система насекомых
как специализированное эффективное
Корреляции в исторически сложившихся системах
Трахейная система насекомых
как специализированное эффективное

Каскадные взаимодействия
Редукция кровеносной системы невозможность снабжения массивных органов
Каскадные взаимодействия
Редукция кровеносной системы невозможность снабжения массивных органов

Корреляции в исторически сложившихся системах
.
Способ вентиляции легких –
кожное дыхание и
Корреляции в исторически сложившихся системах
.
Способ вентиляции легких –
кожное дыхание и

Каскадные взаимодействия
При этом ороговение не имеет б/х и морфологических запретов,
Каскадные взаимодействия
При этом ороговение не имеет б/х и морфологических запретов,

Если естественный отбор длительное время благоприятствовал развитию каких-либо адаптаций (лошадиные
Если естественный отбор длительное время благоприятствовал развитию каких-либо адаптаций (лошадиные

Путь направленной эволюции – арогенез
Отбор благоприятствует ароморфозам, если они
Путь направленной эволюции – арогенез
Отбор благоприятствует ароморфозам, если они

Избыточность биосистем - прогнозирование будущего при цикличности повторяемости событий.
Филогенез -
Избыточность биосистем - прогнозирование будущего при цикличности повторяемости событий.
Филогенез -

Канализирующая роль строения организма.
Адаптивный компромисс. Это означает ограничение эволюционных
Канализирующая роль строения организма.
Адаптивный компромисс. Это означает ограничение эволюционных

Примеры метода «оптимизированного случайного тыка»
1. Работа иммунной системы.
Антитела к новым возбудителям,
Примеры метода «оптимизированного случайного тыка»
1. Работа иммунной системы.
Антитела к новым возбудителям,
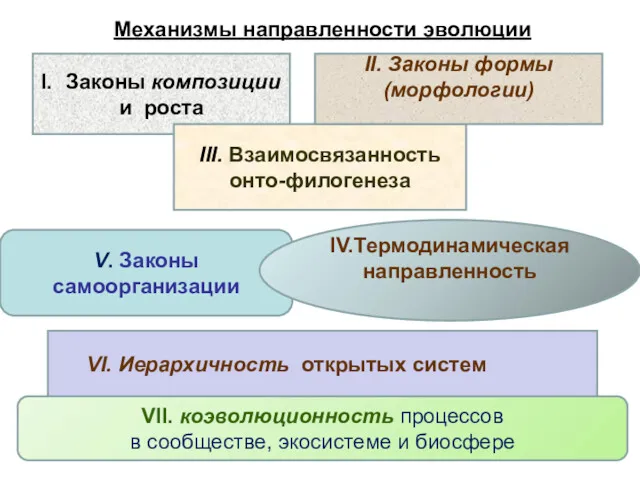
Законы композиции
и роста
V. Законы самоорганизации
III. Взаимосвязанность
онто-филогенеза
VII. коэволюционность процессов
в сообществе,
Законы композиции
и роста
V. Законы самоорганизации
III. Взаимосвязанность
онто-филогенеза
VII. коэволюционность процессов
в сообществе,

Механизмы направленности эволюции
I. Закон композиции и законы роста, связанные
Механизмы направленности эволюции
I. Закон композиции и законы роста, связанные

Числа Фибоначчи
на главном вокзале Цюриха
Плотная пища жён Фибоначчи
Только на
Числа Фибоначчи
на главном вокзале Цюриха
Плотная пища жён Фибоначчи Только на
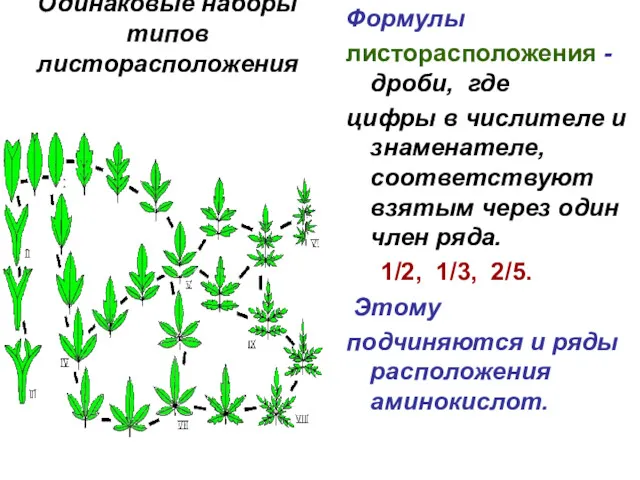
Одинаковые наборы
типов листорасположения
Формулы
листорасположения - дроби, где
цифры в числителе и
Одинаковые наборы
типов листорасположения
Формулы
листорасположения - дроби, где
цифры в числителе и

Геометрические классы спиралей
= у разных моллюсков (хорошие пловцы) и фораминифер
Геометрические классы спиралей
= у разных моллюсков (хорошие пловцы) и фораминифер


II. Законы формы (морфологии)
Производные генетики, физиологии и экологии
(генетические механизмы
II. Законы формы (морфологии)
Производные генетики, физиологии и экологии
(генетические механизмы

III. Взаимосвязанность онто-филогенеза
А.) Новую линию в эволюции могут дать мутации большой
III. Взаимосвязанность онто-филогенеза
А.) Новую линию в эволюции могут дать мутации большой

Б). Стабилизация онтогенеза (несмотря на присущую ему стохастику), за счет специальных
Б). Стабилизация онтогенеза (несмотря на присущую ему стохастику), за счет специальных
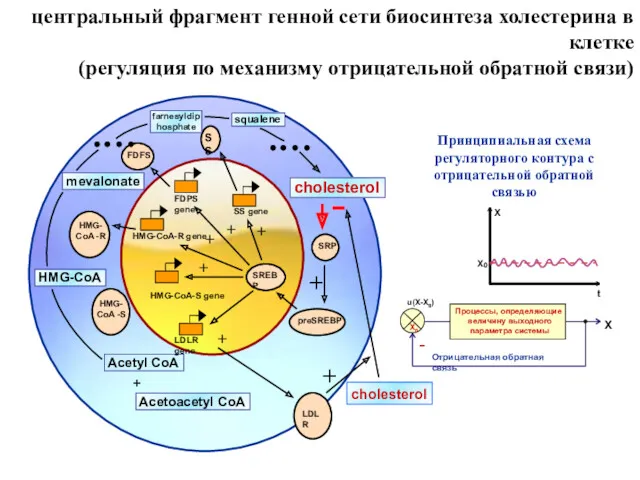
центральный фрагмент генной сети биосинтеза холестерина в клетке
центральный фрагмент генной сети биосинтеза холестерина в клетке
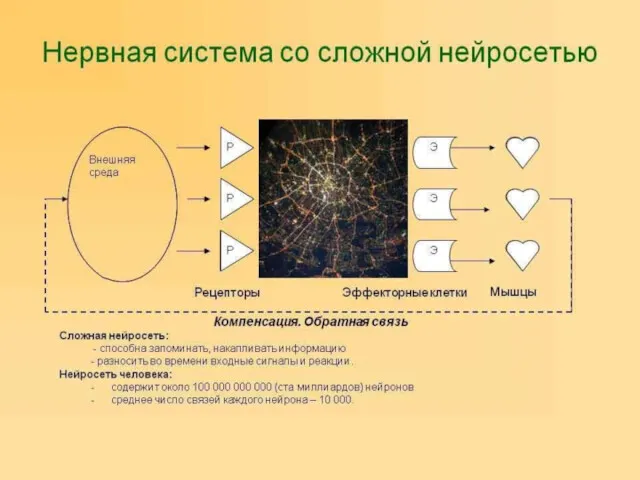
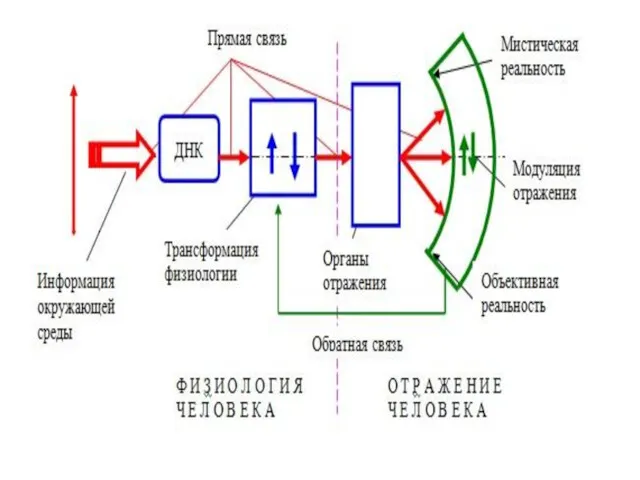
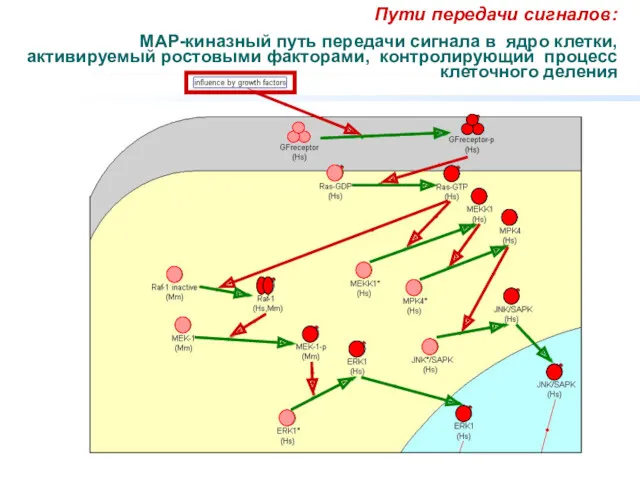
Пути передачи сигналов:
МАР-киназный путь передачи сигнала в ядро клетки, активируемый
Пути передачи сигналов:
МАР-киназный путь передачи сигнала в ядро клетки, активируемый

Важную роль в стабилизации онтогенеза может играть «избыточность» генно-регуляторных сетей, управляющих
Важную роль в стабилизации онтогенеза может играть «избыточность» генно-регуляторных сетей, управляющих
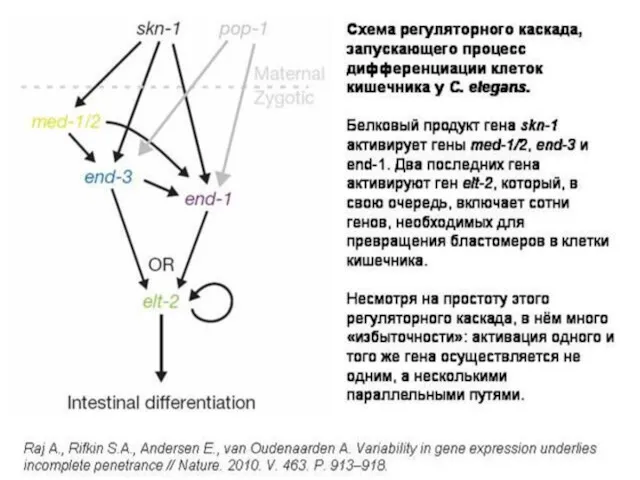

г). Эпигенетическая изменчивость
(М. А. Шишкин).
Сильное внешнее воздействие (например,
г). Эпигенетическая изменчивость
(М. А. Шишкин).
Сильное внешнее воздействие (например,
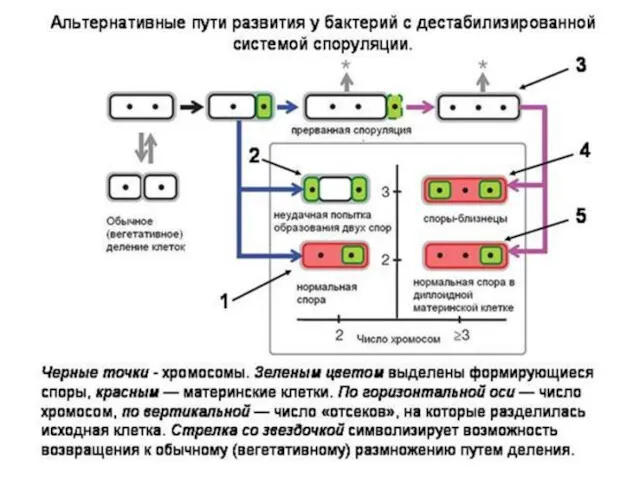

Эпигенетическая теория эволюции рассматривает один из возможных механизмов наследования приобретенных свойств
Эпигенетическая теория эволюции рассматривает один из возможных механизмов наследования приобретенных свойств

Морфозы в широком смысле, т. е. изменение фенотипа без изменения генома,
Морфозы в широком смысле, т. е. изменение фенотипа без изменения генома,

Когда развитие куриного эмбриона достигает определенного этапа, клетки кожи, соединяющей зачатки
Когда развитие куриного эмбриона достигает определенного этапа, клетки кожи, соединяющей зачатки

Основополагающее влияние на наследственность оказывает не геном, а эпигенетическая система (ЭС)
Основополагающее влияние на наследственность оказывает не геном, а эпигенетическая система (ЭС)

Эволюция по ЭТЭ (эпигенетическая теория эволюции) заключается в преобразовании одного креода
Эволюция по ЭТЭ (эпигенетическая теория эволюции) заключается в преобразовании одного креода
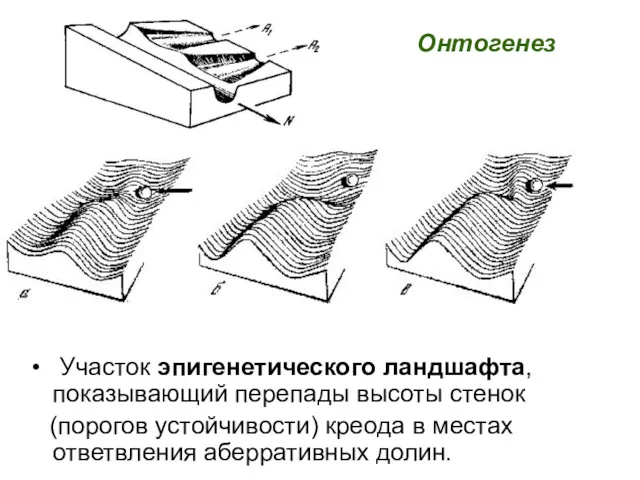
Участок эпигенетического ландшафта, показывающий перепады высоты стенок
(порогов устойчивости)
Участок эпигенетического ландшафта, показывающий перепады высоты стенок
(порогов устойчивости)

Дестабилизация развития может существенно облегчать формирование новых адаптаций.
Дестабилизация «перекидывает мостик»
Дестабилизация развития может существенно облегчать формирование новых адаптаций.
Дестабилизация «перекидывает мостик»

VI. Термодинамическая направленность эволюции
определяемая законом минимизации диссипации энергии при функционировании
VI. Термодинамическая направленность эволюции
определяемая законом минимизации диссипации энергии при функционировании
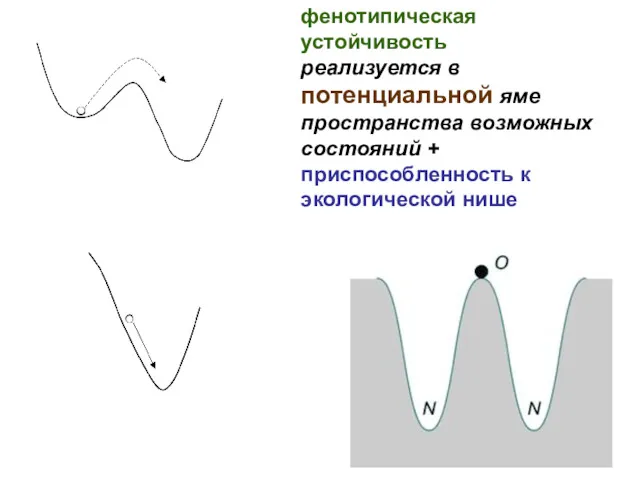
фенотипическая устойчивость
реализуется в
потенциальной яме пространства возможных состояний + приспособленность
фенотипическая устойчивость
реализуется в
потенциальной яме пространства возможных состояний + приспособленность

IV. Законы самоорганизации
Эволюция сложных природных неравновесных систем рассматривается как
IV. Законы самоорганизации
Эволюция сложных природных неравновесных систем рассматривается как
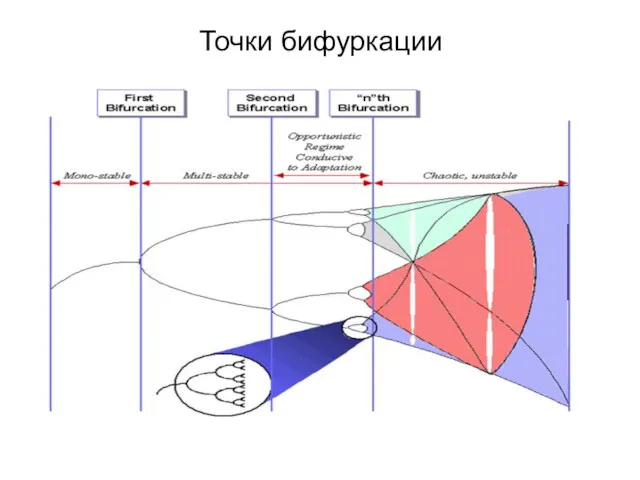
Точки бифуркации
Точки бифуркации

В процессе эволюции система переходит в новое состояние.
Целое задает правила
В процессе эволюции система переходит в новое состояние.
Целое задает правила

Поведение системы в состоянии неустойчивости, может зависеть от поведения очень немногих
Поведение системы в состоянии неустойчивости, может зависеть от поведения очень немногих
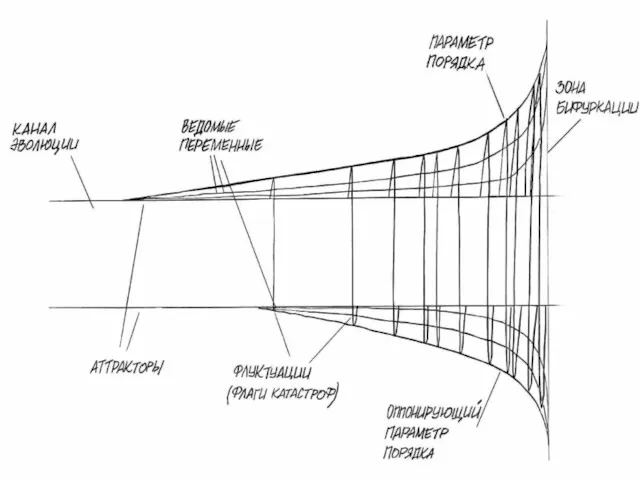
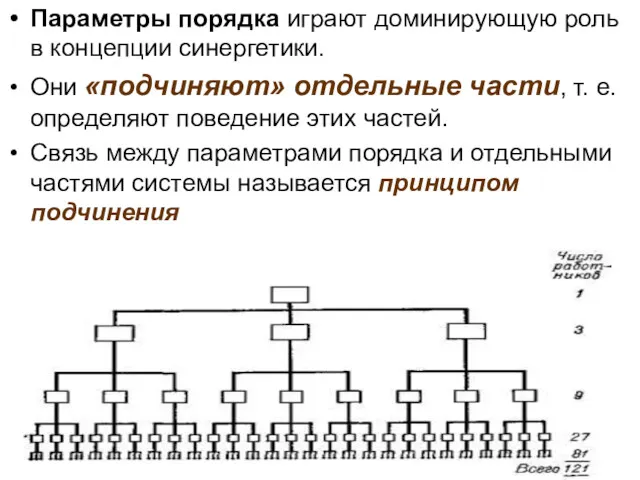
Параметры порядка играют доминирующую роль в концепции синергетики.
Они «подчиняют» отдельные
Параметры порядка играют доминирующую роль в концепции синергетики.
Они «подчиняют» отдельные
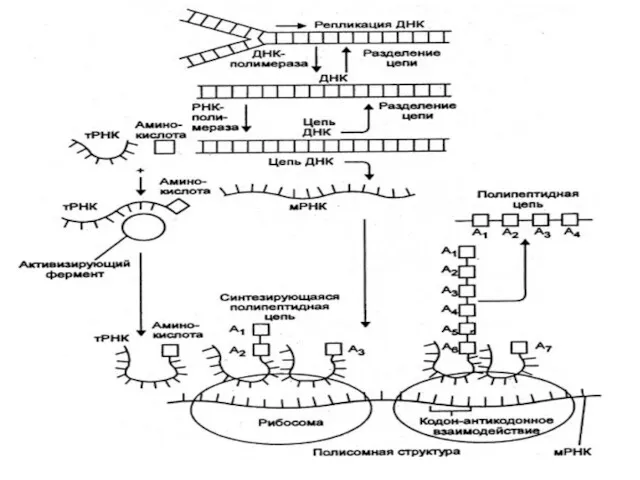

В процессе развития природных систем также можно выделить несколько главных
В процессе развития природных систем также можно выделить несколько главных
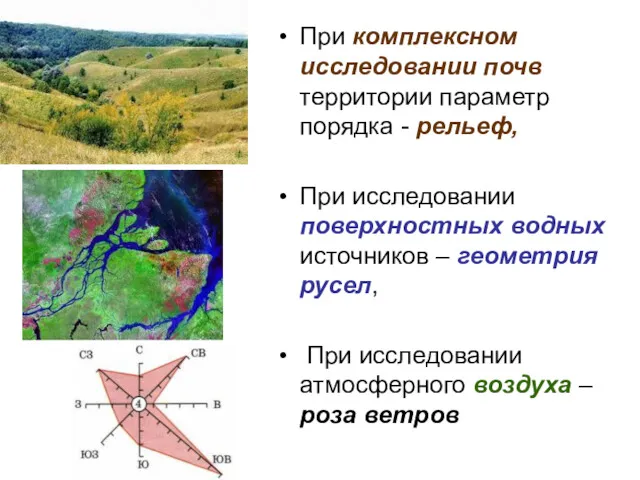
При комплексном исследовании почв территории параметр порядка - рельеф,
При исследовании
При комплексном исследовании почв территории параметр порядка - рельеф,
При исследовании

Для микро-, мезо-(или макро-) и мега- уровня параметры порядка - это
Для микро-, мезо-(или макро-) и мега- уровня параметры порядка - это

Управляющий параметр может быть один и несколько.
Их количество фиксировано и налагается
Управляющий параметр может быть один и несколько.
Их количество фиксировано и налагается


Управляющий параметр выполняет роль триггера, запускающего сложные сетевые процессы.
Он включает
Управляющий параметр выполняет роль триггера, запускающего сложные сетевые процессы.
Он включает

Ю.Л. Климонтович, изучая процессы самоорганизации, в 1983 году доказал S-теорему, которая
Ю.Л. Климонтович, изучая процессы самоорганизации, в 1983 году доказал S-теорему, которая
Направленность эволюции связана с
V. иерархичностью открытых систем,
упорядоченное расположение частей целого
Направленность эволюции связана с
V. иерархичностью открытых систем,
упорядоченное расположение частей целого

Особую роль в иерархии систем играет время,
и синергетический принцип
Особую роль в иерархии систем играет время,
и синергетический принцип

Развитие выступает как направленный процесс количественных и качественных изменений в структуре
Развитие выступает как направленный процесс количественных и качественных изменений в структуре

Направленность эволюции связана с
VII. коэволюционностью процессов
от сообщества до биосферы
Направленность эволюции связана с
VII. коэволюционностью процессов
от сообщества до биосферы

А). Взаимодействие видов через экологические ниши.
- С одной стороны, усиливает
А). Взаимодействие видов через экологические ниши.
- С одной стороны, усиливает

В развитом сообществе все возможные ниши обычно заняты, и новым видам
В развитом сообществе все возможные ниши обычно заняты, и новым видам

Вхождение вида в сообщество может быть в роли доминанта, а в
Вхождение вида в сообщество может быть в роли доминанта, а в

Каждая экосистема предоставляет организмам, живущим в ней или вселяющимся экологическую лицензию,
Каждая экосистема предоставляет организмам, живущим в ней или вселяющимся экологическую лицензию,

Поразительные примеры параллельной эволюции наблюдались в гибнущем Аральском море, который разделился
Поразительные примеры параллельной эволюции наблюдались в гибнущем Аральском море, который разделился

Массовое вымирание освободило множество ниш. Например, вымерли все моллюски-фильтраторы (тогда как
Массовое вымирание освободило множество ниш. Например, вымерли все моллюски-фильтраторы (тогда как
Б). Коэволюционность развития адаптивных зон, которые являются не только средой обитания,
Б). Коэволюционность развития адаптивных зон, которые являются не только средой обитания,

Т.о. понятие адаптивной зоны подразумевает не только эволюцию данного таксона, определяемую
Т.о. понятие адаптивной зоны подразумевает не только эволюцию данного таксона, определяемую
Модель сопряженной эволюции сообщества, его таксономического и экологического разнообразия: 1 —
Модель сопряженной эволюции сообщества, его таксономического и экологического разнообразия: 1 —

В). В экосистеме сохранение вида, его ниши происходит за счет
В). В экосистеме сохранение вида, его ниши происходит за счет

2). Направленность эволюции групп связана с эволюцией самих биогеоценозов как
2). Направленность эволюции групп связана с эволюцией самих биогеоценозов как
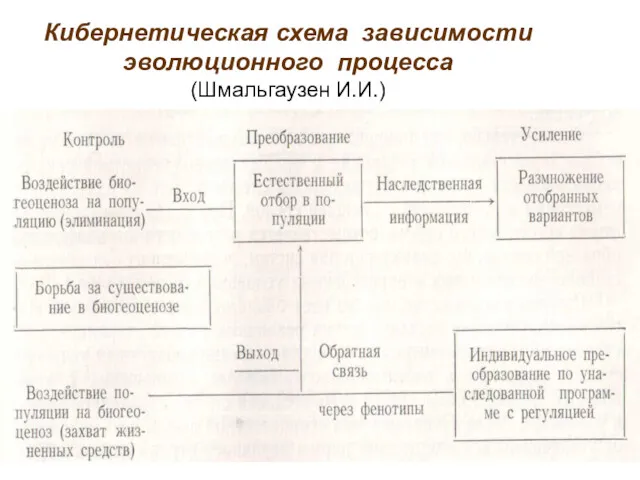
Кибернетическая схема зависимости эволюционного процесса
(Шмальгаузен И.И.)
Кибернетическая схема зависимости эволюционного процесса
(Шмальгаузен И.И.)

Пьер Тейя́р де Шарде́н
(1881 -1955) — французский теолог, философ, священник-иезуит, один
Пьер Тейя́р де Шарде́н
(1881 -1955) — французский теолог, философ, священник-иезуит, один

Эволюция - это возрастание функциональной сложности параллельно росту специфичности, индивидуализации, дискретности.
Эволюция - это возрастание функциональной сложности параллельно росту специфичности, индивидуализации, дискретности.

Будущее открыто и непредсказуемо, но оно не произвольно – т.к.
Будущее открыто и непредсказуемо, но оно не произвольно – т.к.

Эволюция, по всей видимости, – процесс в целом и в самом
Эволюция, по всей видимости, – процесс в целом и в самом

Основные пути эволюции
Прогресс и регресс
Прогресс выражается в подъеме общей
Основные пути эволюции
Прогресс и регресс
Прогресс выражается в подъеме общей

Формы прогрессивного развития:
1. Неограниченный,
2. . Морфофизиологический,
3. Биологический,
4.
Формы прогрессивного развития:
1. Неограниченный,
2. . Морфофизиологический,
3. Биологический,
4.
Неограниченный прогресс:
от прокариот –
до человека,
от индивидуума –
к
Неограниченный прогресс:
от прокариот –
до человека,
от индивидуума –
к

2. Морфофизиологический прогресс
Усложнение организации группы
Проблема выделения критериев:
1. наличие ароморфозов, крупные изменения
2. Морфофизиологический прогресс
Усложнение организации группы
Проблема выделения критериев:
1. наличие ароморфозов, крупные изменения

3. Биологический прогресс
Процветание группы :
увеличение численности,
расширение ареала,
высокая скорость
3. Биологический прогресс
Процветание группы :
увеличение численности,
расширение ареала,
высокая скорость


Групповой прогресс
Совершенствование организации в пределах данной группы организмов при сохранении общего
Групповой прогресс
Совершенствование организации в пределах данной группы организмов при сохранении общего

Биотехнический прогресс - большая эффективность выполнения функций у некоторых групп. Основной
Биотехнический прогресс - большая эффективность выполнения функций у некоторых групп. Основной

Критерии прогресса
Повышение общей Е жизнедеятельности (Северцев).
Совершенствование системы регуляции (Грант).
Критерии прогресса
Повышение общей Е жизнедеятельности (Северцев).
Совершенствование системы регуляции (Грант).

Биологический регресс
Признаки обратные прогрессу:
уменьшение численности,
сужение ареала,
снижение скорости видообразования.
Может привести
Биологический регресс
Признаки обратные прогрессу:
уменьшение численности,
сужение ареала,
снижение скорости видообразования.
Может привести

На границе перми и триаса, то есть палеозойской и мезозойской эр,
На границе перми и триаса, то есть палеозойской и мезозойской эр,
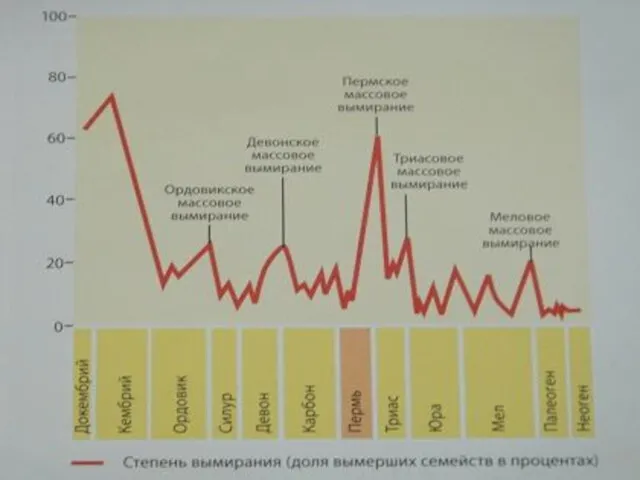
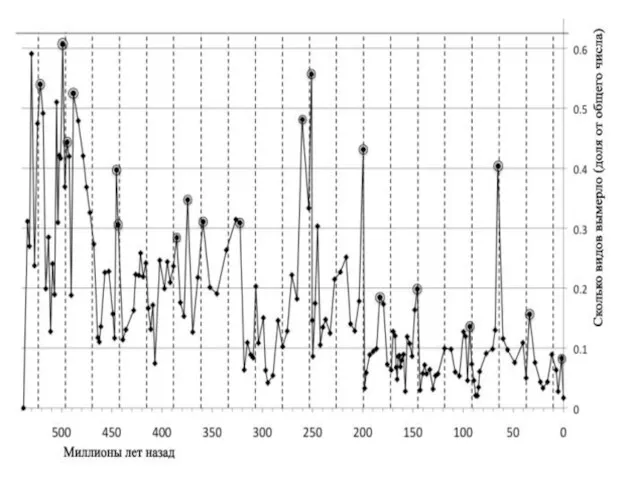

440 млн лет назад — ордовикско-силурийское вымирание — исчезло более 60 % видов морских беспозвоночных;
440 млн лет назад — ордовикско-силурийское вымирание — исчезло более 60 % видов морских беспозвоночных;


Достигнутые эволюционной биологией на настоящий момент рубежи отличаются от представленных
Достигнутые эволюционной биологией на настоящий момент рубежи отличаются от представленных
 Онтогенез как процесс реализации наследственной информации с эпигенетическими факторами. Принципы клеточной регуляции онтогенеза
Онтогенез как процесс реализации наследственной информации с эпигенетическими факторами. Принципы клеточной регуляции онтогенеза Функциональная межполушарная асимметрия мозга
Функциональная межполушарная асимметрия мозга Характеристика водорослей
Характеристика водорослей Основы разведения животных
Основы разведения животных Еңбектенудің физиологиялық негізі
Еңбектенудің физиологиялық негізі Систематические группы птиц
Систематические группы птиц Зачем животным нужен хвост. 4 класс
Зачем животным нужен хвост. 4 класс Абай ауданы, Шығыс Қазақстан облысы
Абай ауданы, Шығыс Қазақстан облысы Введение в эпизоотологию
Введение в эпизоотологию Простые и сложные соцветия
Простые и сложные соцветия Материалы конкурса Уроки качества
Материалы конкурса Уроки качества Микроорганизмы: эукариоты и прокариоты
Микроорганизмы: эукариоты и прокариоты Строение вирусов. Жизненные циклы вирусов,
Строение вирусов. Жизненные циклы вирусов, Методы изучения природы
Методы изучения природы Будова клітин прокаріот та еукаріот
Будова клітин прокаріот та еукаріот Структура и содержание КИМ - 2023
Структура и содержание КИМ - 2023 Цікаві факти з життя комах
Цікаві факти з життя комах ВПР по биологии 6 класс
ВПР по биологии 6 класс Как видят животные
Как видят животные Общие свойства живых организмов
Общие свойства живых организмов Анамнії та амніоти
Анамнії та амніоти Мероприятия экологической акции Скворушка, проводимые в МКОУ БСОШ №7
Мероприятия экологической акции Скворушка, проводимые в МКОУ БСОШ №7 Ферменты. Биологически активные вещества
Ферменты. Биологически активные вещества Механизмы репродукции клеток
Механизмы репродукции клеток Цветок, его строение и значение
Цветок, его строение и значение Иллюзии зрения
Иллюзии зрения Классификация покрытосеменных растений. Отдел Покрытосеменные. Класс Двудольные. Класс Однодольные
Классификация покрытосеменных растений. Отдел Покрытосеменные. Класс Двудольные. Класс Однодольные Гости из далёких стран
Гости из далёких стран