- Обмен аммиака. Механизмы обезвреживания

Содержание
- 2. Основным источником аммиака является катаболизм аминокислот в тканях. Часть аммиака образуется в клетках при распаде азотсодержащих
- 3. Из организма аммиак выводится почками в виде конечных продуктов азотистого обмена: • мочевины - синтезируется в
- 4. Основной реакцией обезвреживания аммиака почти во всех тканях является синтез глутамина под действием глутаминсинтетазы: глутаминсинтетаза обладает
- 5. Глутамин можно считать транспортной формой аммиака, он является нейтральной аминокислотой и способен легко проникать через клеточные
- 6. Из тканей глутамин транспортируется в почки и кишечник. В клетках кишечника под действием фермента глутаминазы происходит
- 7. В почках глутамин также подвергается действию фермента глутаминазы и расщепляется на глутамат, который реабсорбируется и возвращается
- 8. Глутаминаза почек активируется при ацидозе; образовавшийся аммиак используется для нейтрализации кислых продуктов и образования аммонийных солей
- 9. Использование глутамина в почках для поддержания кислотно-щелочного баланса
- 10. В мозге для обезвреживания аммиака используется р-я восстановительного аминирования α-кетоглутарата под действием глутаматдегидрогеназы. Этот путь в
- 11. Из мышц, клеток кишечника и некоторых других тканей избыток азота выводится в кровь в виде аланина
- 12. Особенно много аланина выделяют мышцы в силу их большой массы. Работающие мышцы часть энергии могут получать
- 13. Аланин поступает в печень, где подвергается дезаминированию. Аммиак обезвреживается в процессе синтеза мочевины, а пируват включается
- 14. В печени аммиак обезвреживается путем связывания с СО2 и образования карбамоилфосфата . Реакцию катализирует карбамоилфосфатсинтетаза I,
- 15. Пути обмена азота, аминокислот и аммиака. А - выведение азота из мышц и кишечника в составе
- 16. Орнитиновый цикл мочевинообразования. (цикл Кребса-Гензелейта). Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины.
- 17. Большая часть свободного аммиака и азота в составе АК (глутамин, аланин) поступают в печень, где из
- 18. Реакции орнитинового цикла Предварительно в митохондриях под действием карбамоилфосфатсинтетазы I с затратой 2 АТФ аммиак связывается
- 19. 1. В митохондриях с участием фермента орнитинкарбамоилтрансферазы образуется цитруллин.
- 20. 2. В цитозоле аргининосукцинатсинтетаза с затратой 1 АТФ связывает цитруллин с аспартатом и образует аргининосукцинат (аргининоянтарная
- 21. 3. В цитозоле аргининсукцинатлиаза расщепляет аргининосукцинат на аргинин и фумарат (аминогруппа аспартата оказывается в аргинине)
- 22. 4. В цитозоле аргиназа гидролизует аргинин на орнитин и мочевину. У аргиназы кофакторы ионы Са2+ или
- 23. Орнитиновый цикл Кребса-Гензелейта.
- 24. Суммарное уравнение . NH3 + CO2 + 3ATФ + аспарагиновая кислота + 2H2O → мочевина +
- 25. Взаимосвязь орнитинового цикла и общего пути катаболизма (ЦТК). Фумарат, образующийся в результате расщепления аргининосукцината, превращается в
- 26. Энергетический баланс орнитинового цикла. На синтез 1 мочевины расходуются 4 связи 3 АТФ. Дополнительные затраты энергии
- 27. Орнитиновый цикл в печени выполняет 2 функции: 1. превращение азота АК в мочевину, которая экскретируется и
- 28. Эффективность работы орнитинового цикла при нормальном питании человека и умеренных физических нагрузках составляет примерно 60% его
- 29. Некоторые патологические состояния, характеризующиеся интенсивным распадом белков тканей (сахарный диабет и др.), также сопровождаются активацией орнитинового
- 30. Выделение азота из организма. Азот выводиться из организма с мочой, калом, потом и с выдыхаемым воздухом
- 31. ГИПЕРАММОНИЕМИЯ. Аммиак превращается в мочевину только в печени, поэтому при заболеваниях печени или наследственных дефектах ферментов
- 32. Все нарушения орнитинового цикла приводят к значительному повышению в крови концентрации: аммиака, глутамина, аланина. Для снижения
- 33. ПУТИ ИСПОЛЬЗОВАНИЯ БЕЗАЗОТИСТЫХ ОСТАТКОВ АМИНОКИСЛОТ
- 34. Катаболизм аминокислот -это образование шести соединений, являющихся метаболитами ОПК: пируват, ацетил-КоА, α-кетоглутарат, сукцинил-КоА, фумарат, оксалоацетат.
- 35. К глюкогенным относятся аминокислоты , при распаде которых образуются пируват и метаболиты ЦТК, например, оксалоацетат или
- 36. Полное окисление безазотистых остатков аминокислот до углекислого газа и воды реального энергетического значения не имеет. Основной
- 37. Можно выделить пять анаплеротических реакций: Первые две реакции являются основными анаплеротическими реакциями.
- 38. БИОСИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ Углеродный скелет восьми заменимых аминокислот (Ала, Асп, Асн, Сер, Гли, Про, Глу, Глн)
- 41. Частично заменимые аминокислоты Арг и Гис синтезируются в небольших количествах, которые не отвечают потребностям организма, что
- 43. Скачать презентацию

Основным источником аммиака является катаболизм аминокислот в тканях.
Часть аммиака образуется в клетках
Основным источником аммиака является катаболизм аминокислот в тканях. Часть аммиака образуется в клетках
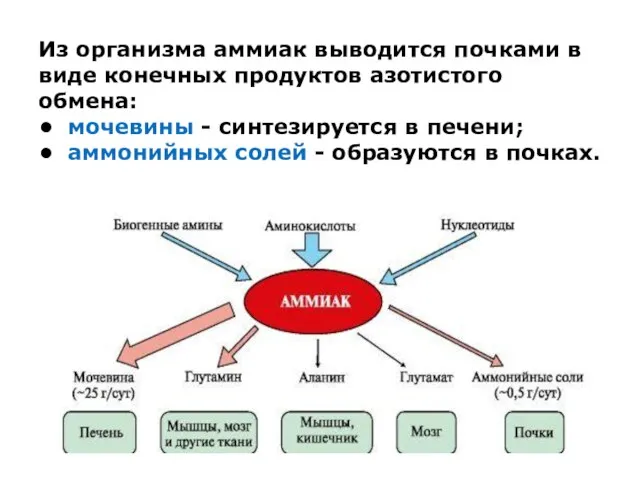
Из организма аммиак выводится почками в виде конечных продуктов азотистого обмена:
• мочевины -
Из организма аммиак выводится почками в виде конечных продуктов азотистого обмена: • мочевины -

Основной реакцией обезвреживания аммиака почти во всех тканях является синтез глутамина под действием глутаминсинтетазы:
глутаминсинтетаза обладает высоким
Основной реакцией обезвреживания аммиака почти во всех тканях является синтез глутамина под действием глутаминсинтетазы: глутаминсинтетаза обладает высоким

Глутамин можно считать транспортной формой аммиака, он является нейтральной аминокислотой и способен легко проникать
Глутамин можно считать транспортной формой аммиака, он является нейтральной аминокислотой и способен легко проникать

Из тканей глутамин транспортируется в почки и кишечник. В клетках кишечника под действием
Из тканей глутамин транспортируется в почки и кишечник. В клетках кишечника под действием

В почках глутамин также подвергается действию фермента глутаминазы и
расщепляется на глутамат, который реабсорбируется
В почках глутамин также подвергается действию фермента глутаминазы и расщепляется на глутамат, который реабсорбируется

Глутаминаза почек активируется при ацидозе; образовавшийся аммиак используется для нейтрализации кислых продуктов
Глутаминаза почек активируется при ацидозе; образовавшийся аммиак используется для нейтрализации кислых продуктов
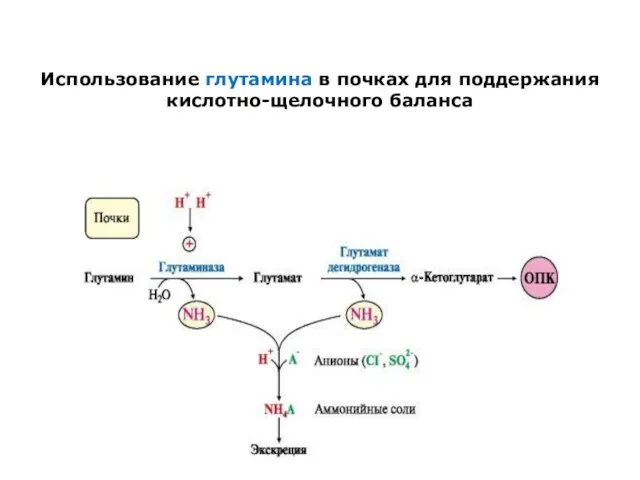
Использование глутамина в почках для поддержания кислотно-щелочного баланса
Использование глутамина в почках для поддержания кислотно-щелочного баланса

В мозге для обезвреживания аммиака используется р-я восстановительного аминирования α-кетоглутарата под действием
В мозге для обезвреживания аммиака используется р-я восстановительного аминирования α-кетоглутарата под действием
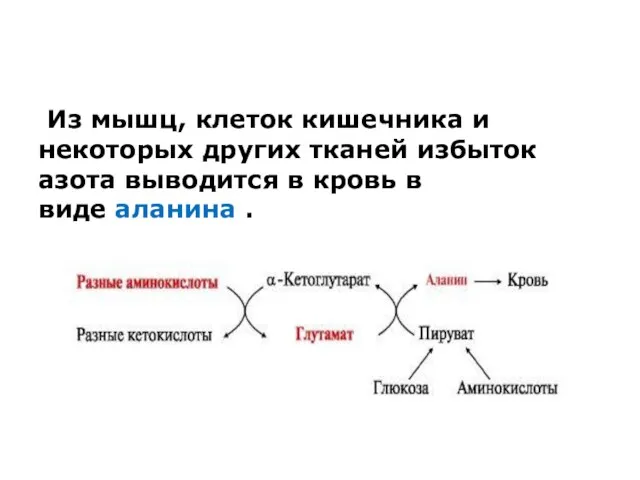
Из мышц, клеток кишечника и некоторых других тканей избыток азота выводится в
Из мышц, клеток кишечника и некоторых других тканей избыток азота выводится в
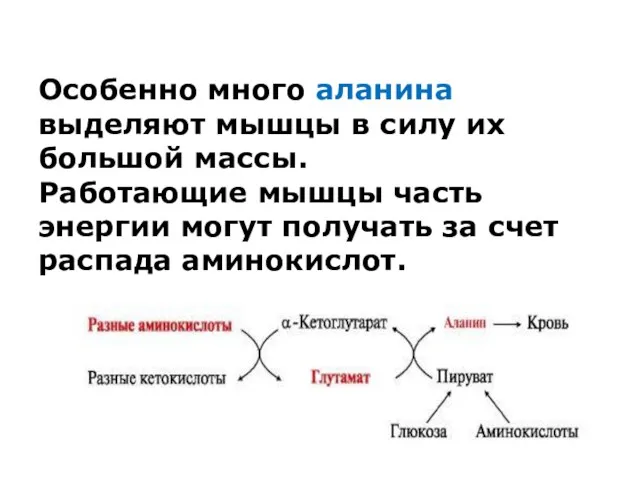
Особенно много аланина выделяют мышцы в силу их большой массы.
Работающие
Особенно много аланина выделяют мышцы в силу их большой массы. Работающие
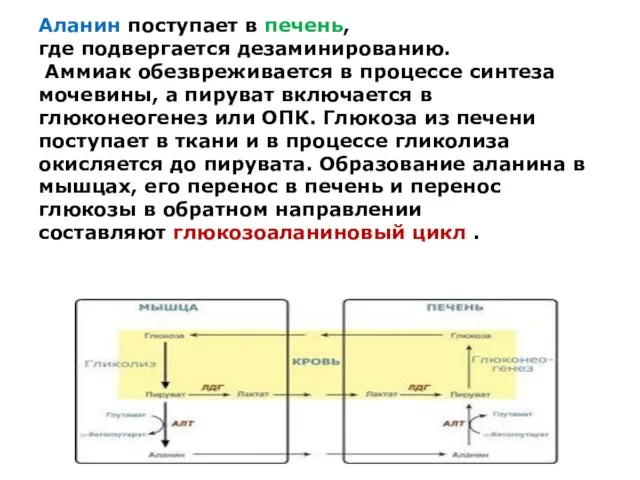
Аланин поступает в печень,
где подвергается дезаминированию.
Аммиак обезвреживается в процессе синтеза
Аланин поступает в печень, где подвергается дезаминированию. Аммиак обезвреживается в процессе синтеза

В печени аммиак обезвреживается путем связывания с СО2 и образования карбамоилфосфата .
Реакцию катализирует
карбамоилфосфатсинтетаза I, которая
В печени аммиак обезвреживается путем связывания с СО2 и образования карбамоилфосфата . Реакцию катализирует карбамоилфосфатсинтетаза I, которая
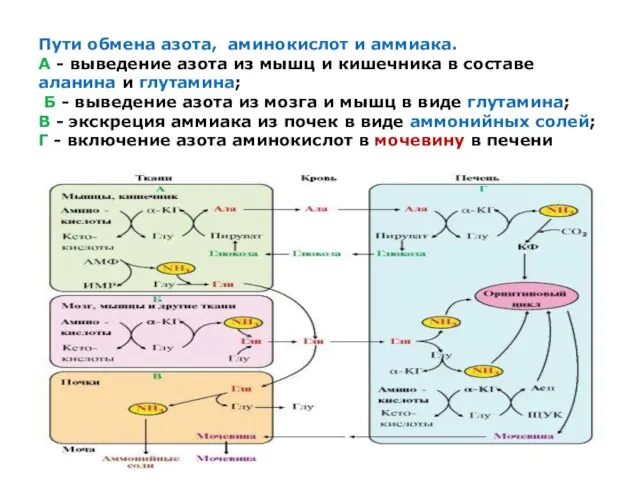
Пути обмена азота, аминокислот и аммиака.
А - выведение азота из мышц и
Пути обмена азота, аминокислот и аммиака. А - выведение азота из мышц и

Орнитиновый цикл мочевинообразования.
(цикл Кребса-Гензелейта).
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины.
Орнитиновый цикл мочевинообразования.
(цикл Кребса-Гензелейта).
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины.
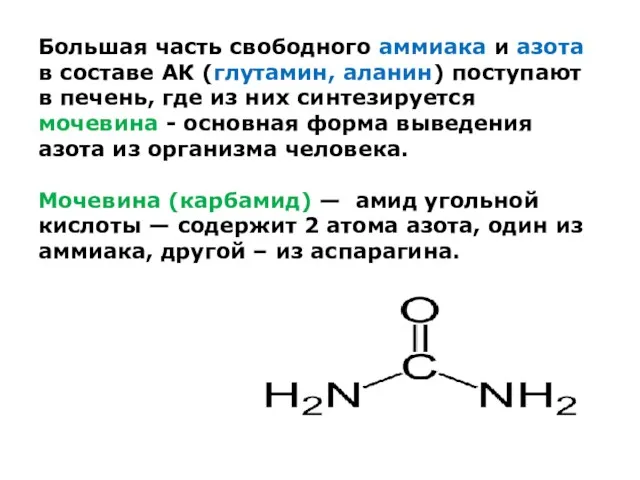
Большая часть свободного аммиака и азота в составе АК (глутамин, аланин)
Большая часть свободного аммиака и азота в составе АК (глутамин, аланин)

Реакции орнитинового цикла
Предварительно в митохондриях под действием карбамоилфосфатсинтетазы I с затратой
2 АТФ
Реакции орнитинового цикла Предварительно в митохондриях под действием карбамоилфосфатсинтетазы I с затратой 2 АТФ

1. В митохондриях с участием фермента орнитинкарбамоилтрансферазы
образуется цитруллин.
1. В митохондриях с участием фермента орнитинкарбамоилтрансферазы
образуется цитруллин.

2. В цитозоле аргининосукцинатсинтетаза с затратой 1 АТФ связывает цитруллин с аспартатом и
2. В цитозоле аргининосукцинатсинтетаза с затратой 1 АТФ связывает цитруллин с аспартатом и

3. В цитозоле аргининсукцинатлиаза расщепляет аргининосукцинат на аргинин и фумарат (аминогруппа аспартата
3. В цитозоле аргининсукцинатлиаза расщепляет аргининосукцинат на аргинин и фумарат (аминогруппа аспартата

4. В цитозоле аргиназа гидролизует аргинин на орнитин и мочевину. У аргиназы кофакторы
4. В цитозоле аргиназа гидролизует аргинин на орнитин и мочевину. У аргиназы кофакторы
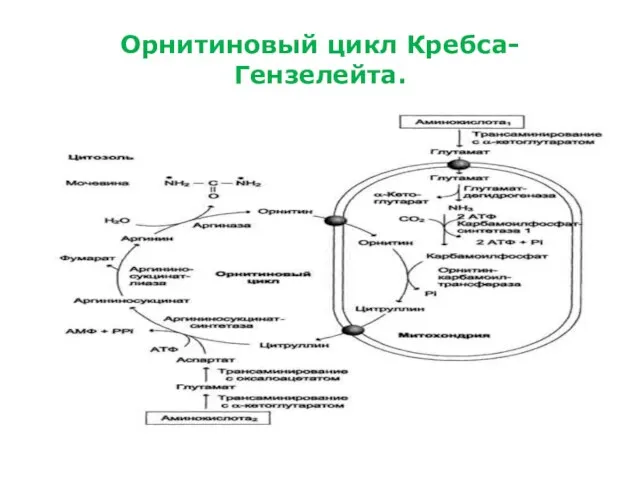
Орнитиновый цикл Кребса-Гензелейта.
Орнитиновый цикл Кребса-Гензелейта.
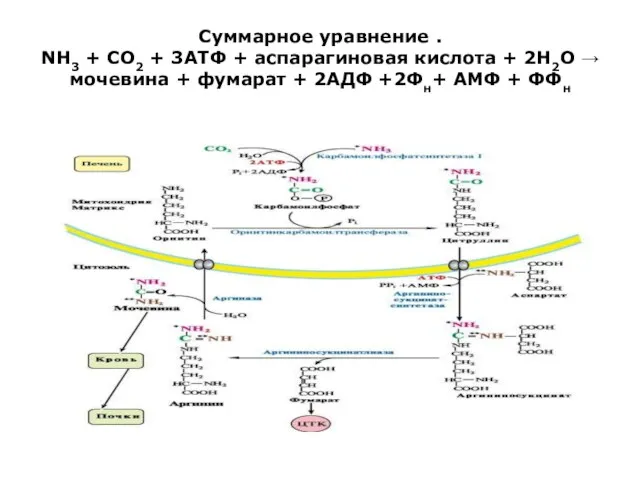
Суммарное уравнение .
NH3 + CO2 + 3ATФ + аспарагиновая кислота + 2H2O →
Суммарное уравнение . NH3 + CO2 + 3ATФ + аспарагиновая кислота + 2H2O →
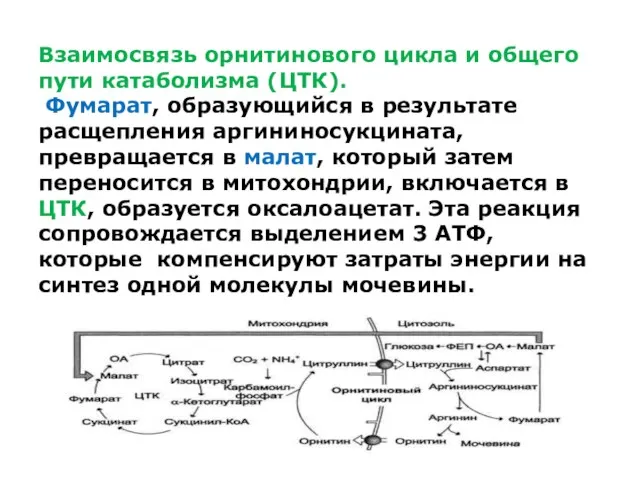
Взаимосвязь орнитинового цикла и общего пути катаболизма (ЦТК).
Фумарат, образующийся в
Взаимосвязь орнитинового цикла и общего пути катаболизма (ЦТК). Фумарат, образующийся в

Энергетический баланс орнитинового цикла.
На синтез 1 мочевины расходуются
4 связи 3
Энергетический баланс орнитинового цикла. На синтез 1 мочевины расходуются 4 связи 3
Орнитиновый цикл в печени выполняет 2 функции:
1. превращение азота АК в
Орнитиновый цикл в печени выполняет 2 функции: 1. превращение азота АК в

Эффективность работы орнитинового цикла при нормальном питании человека и умеренных физических
Эффективность работы орнитинового цикла при нормальном питании человека и умеренных физических

Некоторые патологические состояния, характеризующиеся интенсивным распадом белков тканей (сахарный диабет и
Некоторые патологические состояния, характеризующиеся интенсивным распадом белков тканей (сахарный диабет и

Выделение азота из организма.
Азот выводиться из организма с мочой, калом, потом
Выделение азота из организма. Азот выводиться из организма с мочой, калом, потом

ГИПЕРАММОНИЕМИЯ.
Аммиак превращается в мочевину только в печени, поэтому при заболеваниях печени
ГИПЕРАММОНИЕМИЯ. Аммиак превращается в мочевину только в печени, поэтому при заболеваниях печени

Все нарушения орнитинового цикла приводят к значительному повышению в крови концентрации: аммиака, глутамина, аланина.
Для снижения
Все нарушения орнитинового цикла приводят к значительному повышению в крови концентрации: аммиака, глутамина, аланина. Для снижения

ПУТИ ИСПОЛЬЗОВАНИЯ БЕЗАЗОТИСТЫХ ОСТАТКОВ АМИНОКИСЛОТ
ПУТИ ИСПОЛЬЗОВАНИЯ БЕЗАЗОТИСТЫХ ОСТАТКОВ АМИНОКИСЛОТ

Катаболизм аминокислот -это образование шести соединений, являющихся метаболитами ОПК: пируват, ацетил-КоА,
Катаболизм аминокислот -это образование шести соединений, являющихся метаболитами ОПК: пируват, ацетил-КоА,

К глюкогенным относятся аминокислоты , при распаде которых образуются пируват и метаболиты ЦТК,
К глюкогенным относятся аминокислоты , при распаде которых образуются пируват и метаболиты ЦТК,

Полное окисление безазотистых остатков аминокислот до углекислого газа и воды реального
Полное окисление безазотистых остатков аминокислот до углекислого газа и воды реального

Можно выделить пять анаплеротических реакций:
Первые две реакции являются основными анаплеротическими реакциями.
Можно выделить пять анаплеротических реакций:
Первые две реакции являются основными анаплеротическими реакциями.

БИОСИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ
Углеродный скелет восьми заменимых аминокислот (Ала, Асп, Асн, Сер, Гли,
БИОСИНТЕЗ ЗАМЕНИМЫХ АМИНОКИСЛОТ Углеродный скелет восьми заменимых аминокислот (Ала, Асп, Асн, Сер, Гли,


Частично заменимые аминокислоты Арг и Гис синтезируются в небольших количествах, которые не
Частично заменимые аминокислоты Арг и Гис синтезируются в небольших количествах, которые не
 Викторина Растительная отгадайка. 6 класс
Викторина Растительная отгадайка. 6 класс Лист — боковой орган побега
Лист — боковой орган побега Бұршақ тұқымдастар
Бұршақ тұқымдастар Проект группы дыхания. Как мы дышим
Проект группы дыхания. Как мы дышим Размножение комнатных растений
Размножение комнатных растений Растения Дальневосточного региона
Растения Дальневосточного региона Хемосинтез. Автотрофное питание
Хемосинтез. Автотрофное питание Клас Павукоподібні
Клас Павукоподібні Функции отдельных желез внутренней секреции
Функции отдельных желез внутренней секреции Общая характеристика царства растения
Общая характеристика царства растения Откуда берутся шоколад, изюм и мед
Откуда берутся шоколад, изюм и мед Взаимосвязи компонентов природы. Природный комплекс. (проект Урок в Москве)
Взаимосвязи компонентов природы. Природный комплекс. (проект Урок в Москве) Строение, свойства костей типы их соединения
Строение, свойства костей типы их соединения Мейоз
Мейоз Тип Моллюски
Тип Моллюски Рослинний матеріал у озелененні населених місць
Рослинний матеріал у озелененні населених місць Перелетные птицы
Перелетные птицы презентация Строение почек
презентация Строение почек Класс - двудольные. Семейство - крестоцветные. Род - капуста
Класс - двудольные. Семейство - крестоцветные. Род - капуста Разнообразие цветов. Строение цветка
Разнообразие цветов. Строение цветка Мой домашний зоопарк
Мой домашний зоопарк Изучение биоритмов человека – их влияние на жизнедеятельность
Изучение биоритмов человека – их влияние на жизнедеятельность Методы исследования генетики человека
Методы исследования генетики человека Віроїди, мікоплазми, рикетсії, нематоди, квіткові рослини-паразити як збудники хвороб рослин
Віроїди, мікоплазми, рикетсії, нематоди, квіткові рослини-паразити як збудники хвороб рослин Самые популярные рептилии для террариума
Самые популярные рептилии для террариума Проблемы Антропогенеза (фрагмент курса Эволюция Биосферы)
Проблемы Антропогенеза (фрагмент курса Эволюция Биосферы) Животные семейства кошачьих (фотографии)
Животные семейства кошачьих (фотографии) Кровь и остальные компоненты внутренней среды организма
Кровь и остальные компоненты внутренней среды организма