- Обмен белков. Источники и пути обезвреживания аммиака. (Лекция 15)

Содержание
- 2. В процессе эволюции сформировались различные типы азотистого обмена, которые классифицируются по основному конечному продукту: 1) аммониотелический
- 3. Обмен аммиака. Основной источник аммиака - АК. Большая часть образовавшегося аммиака обезвреживается в орнитиновом цикле в
- 4. Количество азотсодержащих веществ в моче (%) при нормальном белковом питании.
- 5. поступление аммиака из кишечника в портальную вену
- 7. Функции аммиака Аммиак вовлекается (непосредственно или через глутамин) в синтез многих компонентов организма: заменимые аминокислоты, пурины,
- 8. Аммиак - токсичное соединение. Даже небольшое повышение его концентрации оказывает неблагоприятное действие на организм, и прежде
- 9. Недостаточность α-кетоглутарата приводит к снижению концентрации метаболитов ЦТК, что вызывает ускорение реакции синтеза оксалоацетата из пирувата,
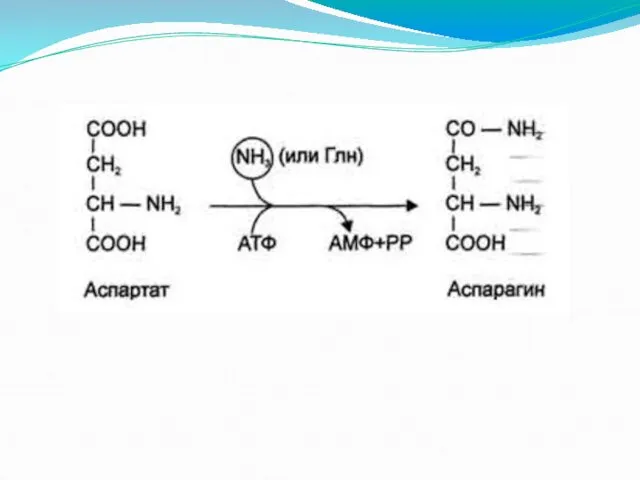
- 10. Обезвреживание аммиака: Основной путь — это связывание аммиака с глутаминовой и реже аспарагиновой кислотами с образованием
- 11. Пути использования глутамина в организме.
- 12. Амидирование остатков глутаминовой и аспарагиновой кислот в составе белков. Восстановительное аминирование 2-оксоглутарата в глутамат. Глутамат в
- 16. Метаболизм амидного азота глутамина в почках Общее обезвреживание аммиака
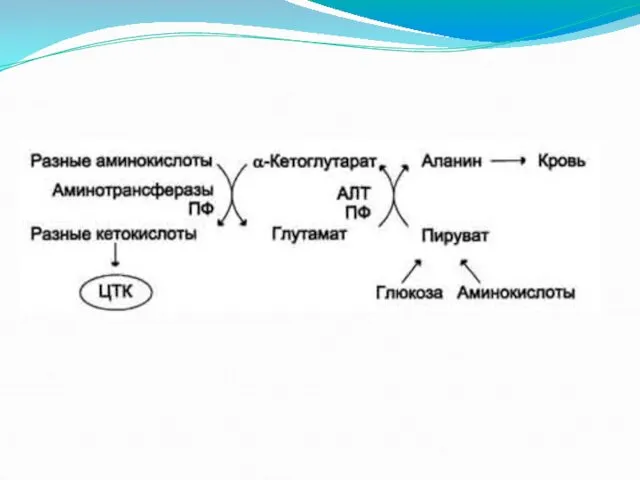
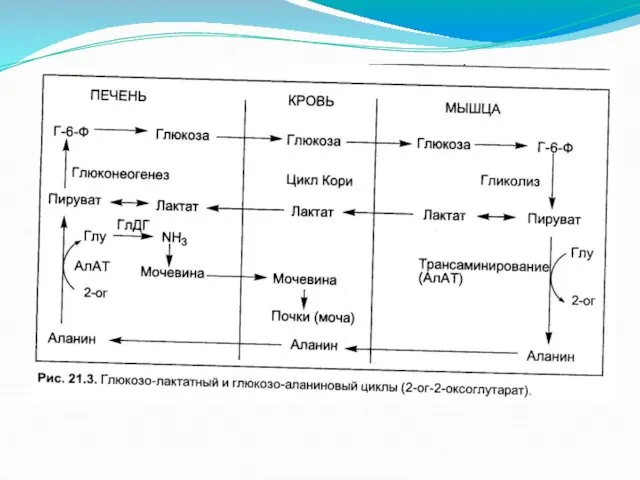
- 17. Глюкозо-аланиновый цикл.
- 18. Орнитиновый цикл Кребса-Гензелейта. Окислительное дезаминирование глутамата происходит в митохондриях. Ферменты орнитинового цикла распределены между митохондриями и
- 19. Синтез мочевины
- 20. Цикл регенерации аспартата, сопряжённый с орнитиновым циклом.
- 21. Орнитиновый цикл в печени выполняет 2 функции: превращение азота аминокислот в мочевину, которая экскретируется и предотвращает
- 22. Взаимосвязь орнитинового цикла и общего пути катаболизма. Фумарат, образующийся в результате расщепления аргининосукцината, превращается в малат,
- 23. Пути выведения аммиака при включении в диету глутамата и фенилацетата (А), бензоата (Б), цитруллина и аргинина
- 24. Нарушения синтеза мочевины
- 25. Нарушение синтеза и выведения мочевины
- 26. Количество азотсодержащих веществ в моче (%) при нормальном белковом питании
- 27. Синтез и распад пуриновых и пиримидиновых оснований
- 28. Строение мононуклеотида
- 29. Пуриновые основания нуклеиновых кислот
- 30. Пиримидиновые основания нуклеиновых кислот
- 31. Углеводы нуклеотидов
- 32. Гидролиз нуклеопротеинов
- 33. Катаболизм пуриновых оснований
- 34. Ингибитор ксантиноксидазы
- 35. Распад пиримидиновых оснований
- 36. Синтез инозиновой кислоты
- 37. Происхождение атомов пуринового кольца
- 38. Синтез пуриновых нуклеотидов
- 39. Биосинтез пиримидинов
- 40. Синтез пиримидиновых нуклеотидов
- 41. Синтез дезоксирибонуклеотидов
- 42. Пуриновые основания нуклеиновых кислот
- 43. Пиримидиновые основания нуклеиновых кислот
- 44. Углеводы нуклеотидов
- 45. Строение мононуклеотида
- 47. Скачать презентацию

В процессе эволюции сформировались различные типы азотистого обмена, которые классифицируются по
В процессе эволюции сформировались различные типы азотистого обмена, которые классифицируются по
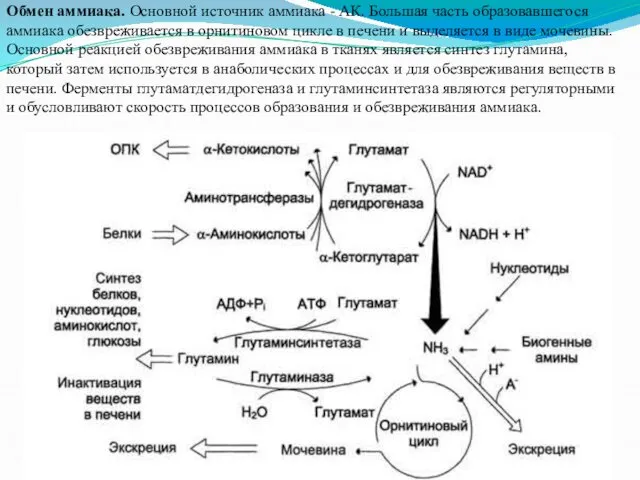
Обмен аммиака. Основной источник аммиака - АК. Большая часть образовавшегося аммиака
Обмен аммиака. Основной источник аммиака - АК. Большая часть образовавшегося аммиака
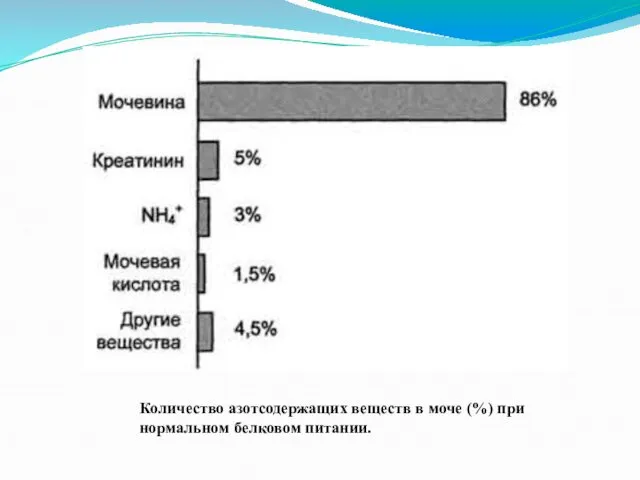
Количество азотсодержащих веществ в моче (%) при нормальном белковом питании.
Количество азотсодержащих веществ в моче (%) при нормальном белковом питании.

поступление аммиака из кишечника в портальную вену
поступление аммиака из кишечника в портальную вену

Функции аммиака
Аммиак вовлекается (непосредственно или через глутамин) в синтез многих компонентов
Функции аммиака
Аммиак вовлекается (непосредственно или через глутамин) в синтез многих компонентов

Аммиак - токсичное соединение. Даже небольшое повышение его концентрации оказывает неблагоприятное
Аммиак - токсичное соединение. Даже небольшое повышение его концентрации оказывает неблагоприятное

Недостаточность α-кетоглутарата приводит к снижению концентрации метаболитов ЦТК, что вызывает ускорение
Недостаточность α-кетоглутарата приводит к снижению концентрации метаболитов ЦТК, что вызывает ускорение
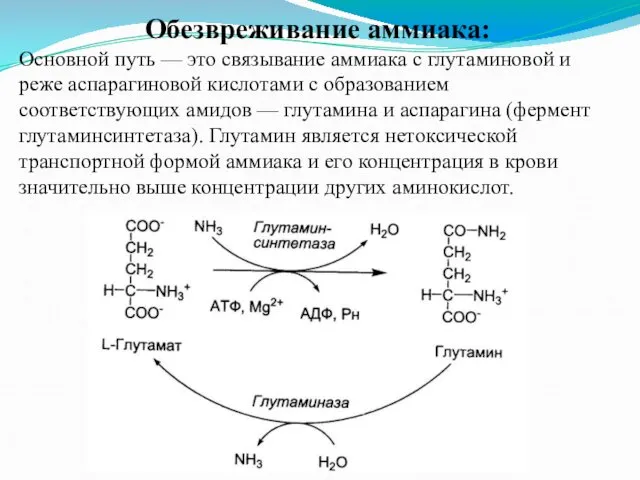
Обезвреживание аммиака:
Основной путь — это связывание аммиака с глутаминовой и реже
Обезвреживание аммиака:
Основной путь — это связывание аммиака с глутаминовой и реже
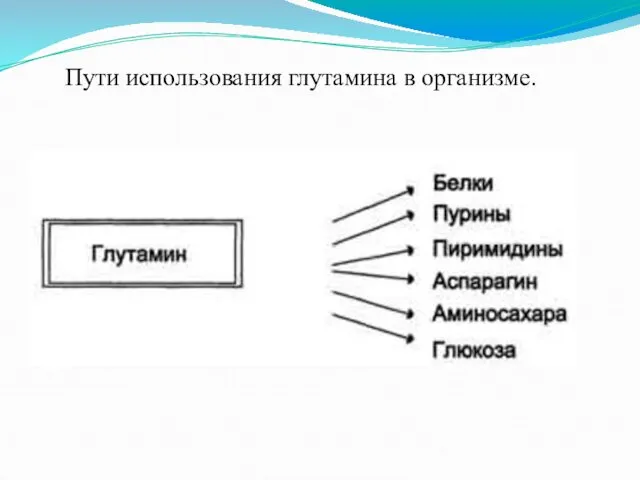
Пути использования глутамина в организме.
Пути использования глутамина в организме.
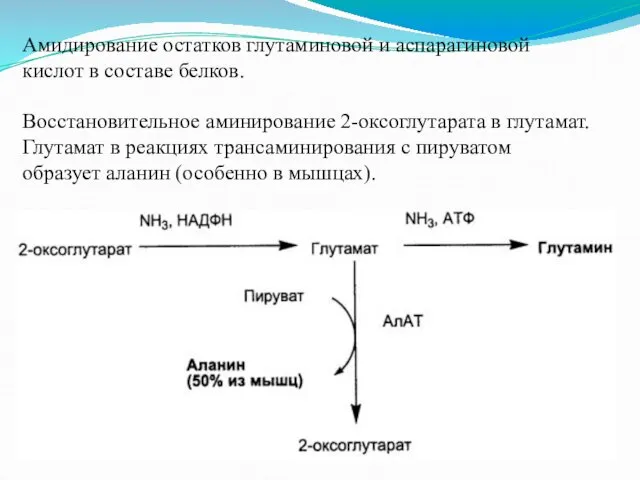
Амидирование остатков глутаминовой и аспарагиновой кислот в составе белков.
Восстановительное аминирование 2-оксоглутарата
Амидирование остатков глутаминовой и аспарагиновой кислот в составе белков.
Восстановительное аминирование 2-оксоглутарата
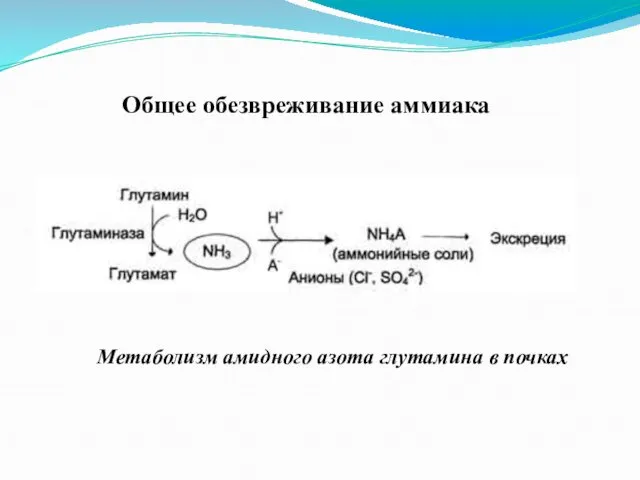
Метаболизм амидного азота глутамина в почках
Общее обезвреживание аммиака
Метаболизм амидного азота глутамина в почках
Общее обезвреживание аммиака
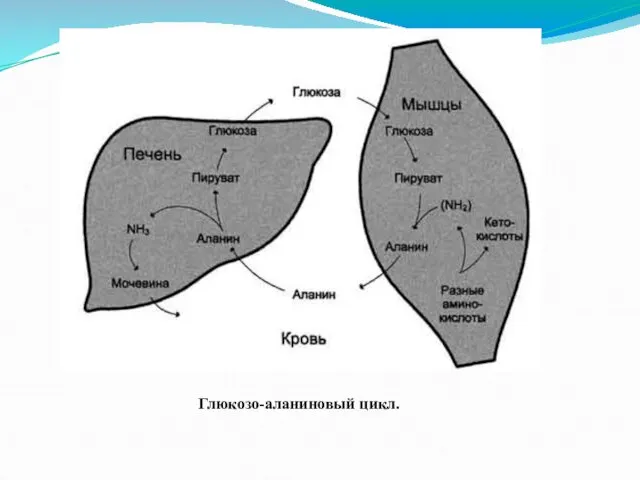
Глюкозо-аланиновый цикл.
Глюкозо-аланиновый цикл.
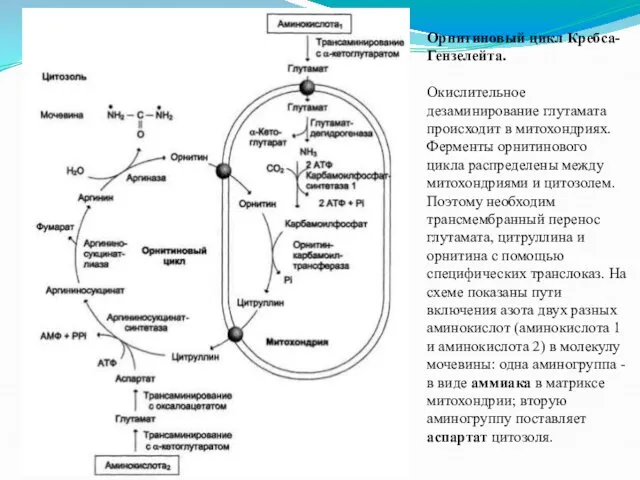
Орнитиновый цикл Кребса-Гензелейта.
Окислительное дезаминирование глутамата происходит в митохондриях. Ферменты орнитинового
Орнитиновый цикл Кребса-Гензелейта.
Окислительное дезаминирование глутамата происходит в митохондриях. Ферменты орнитинового
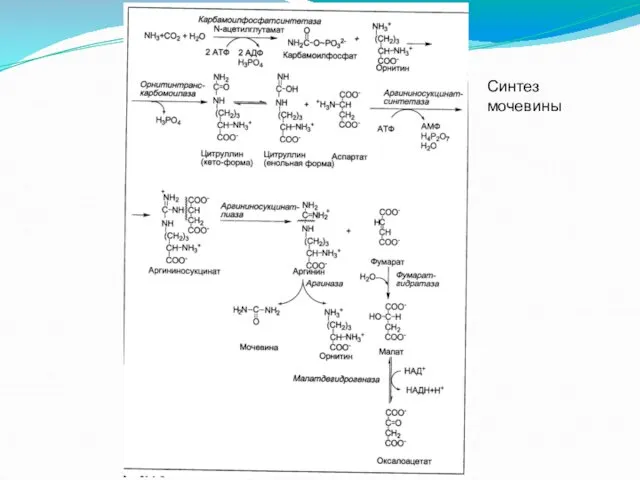
Синтез мочевины
Синтез мочевины
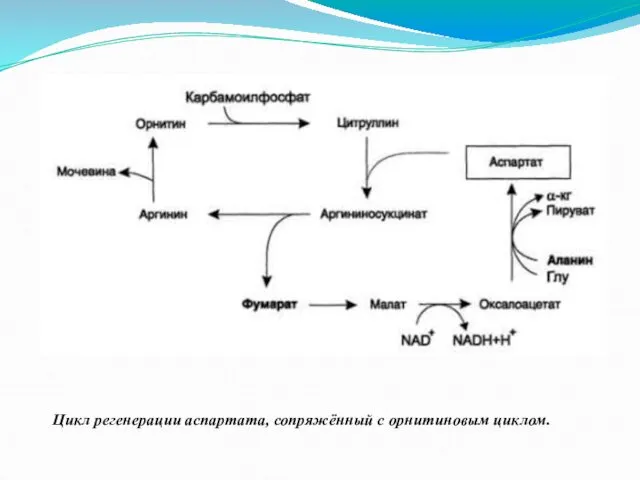
Цикл регенерации аспартата, сопряжённый с орнитиновым циклом.
Цикл регенерации аспартата, сопряжённый с орнитиновым циклом.

Орнитиновый цикл в печени выполняет 2 функции:
превращение азота аминокислот в
Орнитиновый цикл в печени выполняет 2 функции:
превращение азота аминокислот в
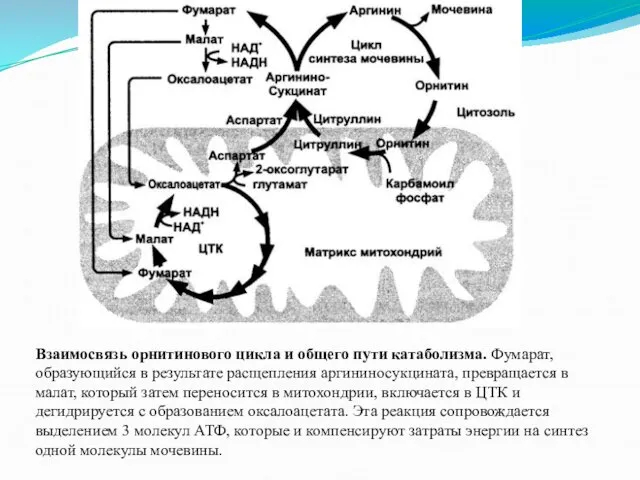
Взаимосвязь орнитинового цикла и общего пути катаболизма. Фумарат, образующийся в результате
Взаимосвязь орнитинового цикла и общего пути катаболизма. Фумарат, образующийся в результате
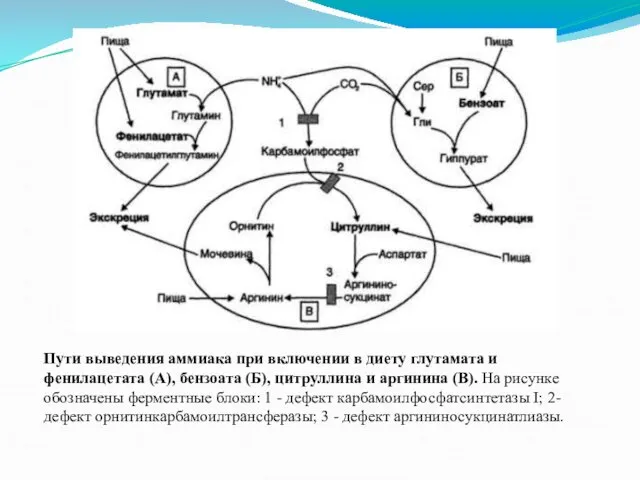
Пути выведения аммиака при включении в диету глутамата и фенилацетата (А),
Пути выведения аммиака при включении в диету глутамата и фенилацетата (А),
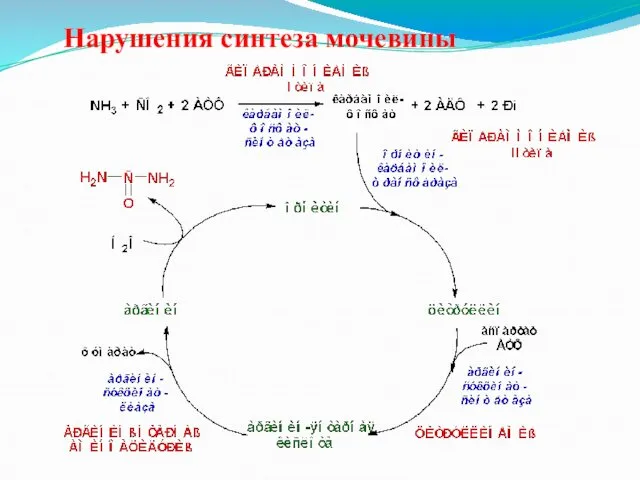
Нарушения синтеза мочевины
Нарушения синтеза мочевины
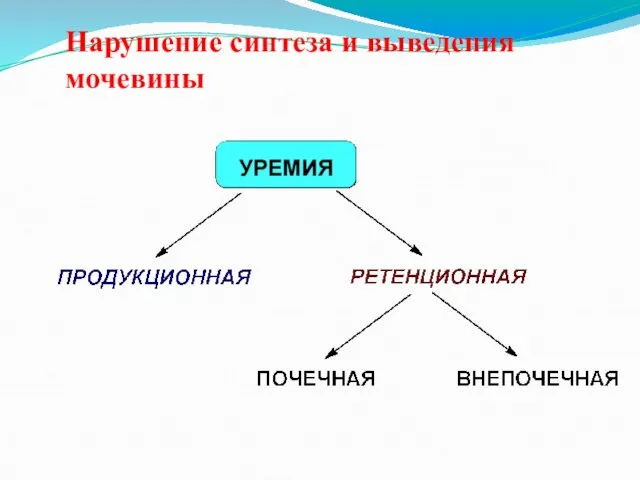
Нарушение синтеза и выведения мочевины
Нарушение синтеза и выведения мочевины

Количество азотсодержащих веществ в моче (%) при нормальном белковом питании
Количество азотсодержащих веществ в моче (%) при нормальном белковом питании

Синтез и распад пуриновых и пиримидиновых оснований
Синтез и распад пуриновых и пиримидиновых оснований
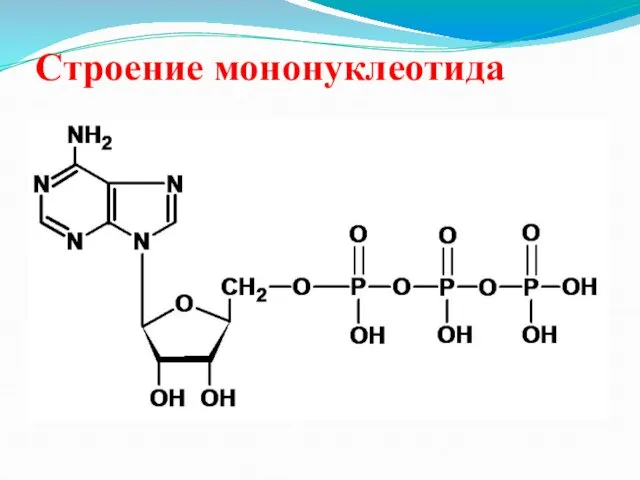
Строение мононуклеотида
Строение мононуклеотида
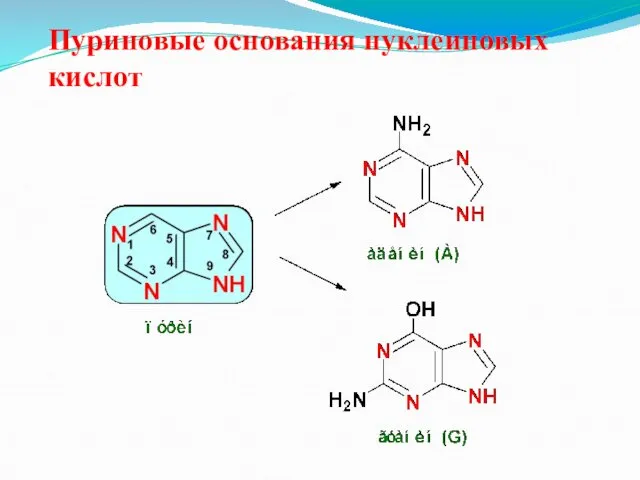
Пуриновые основания нуклеиновых кислот
Пуриновые основания нуклеиновых кислот
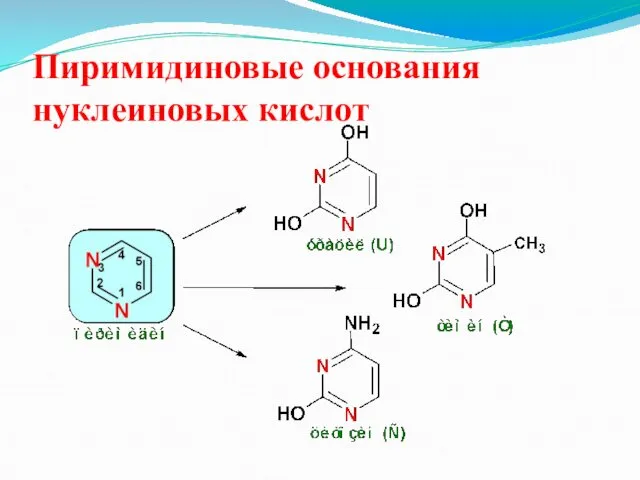
Пиримидиновые основания нуклеиновых кислот
Пиримидиновые основания нуклеиновых кислот
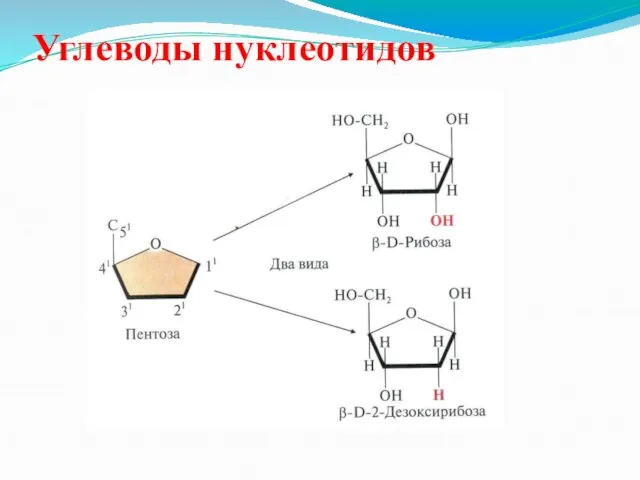
Углеводы нуклеотидов
Углеводы нуклеотидов
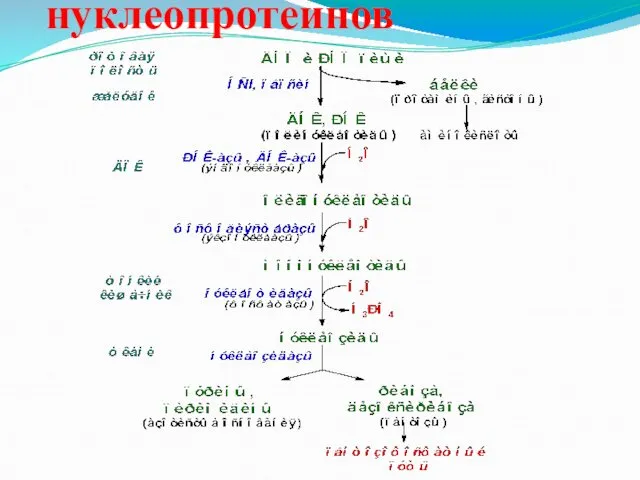
Гидролиз нуклеопротеинов
Гидролиз нуклеопротеинов
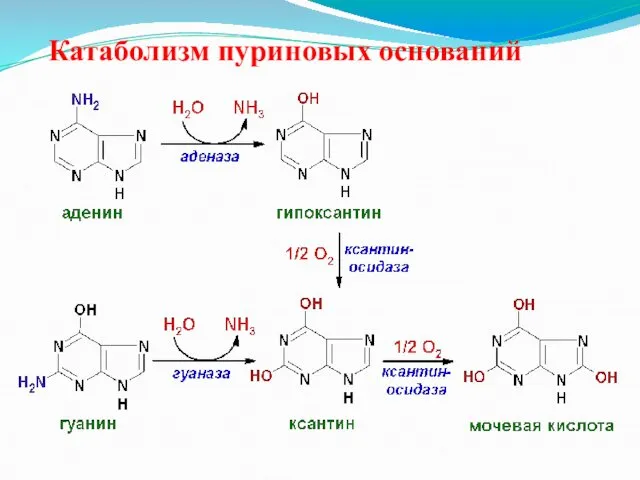
Катаболизм пуриновых оснований
Катаболизм пуриновых оснований
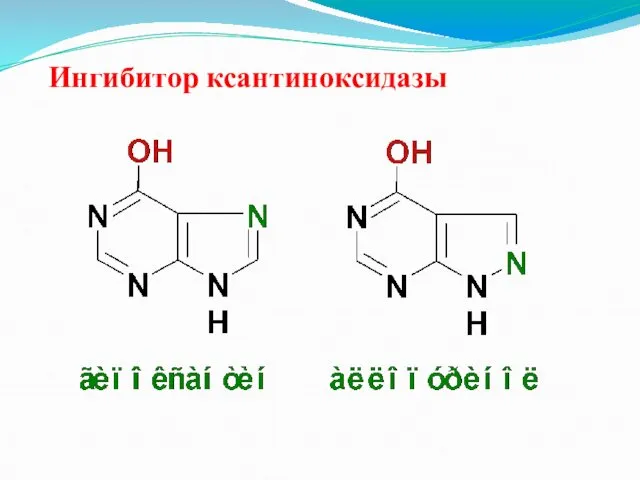
Ингибитор ксантиноксидазы
Ингибитор ксантиноксидазы
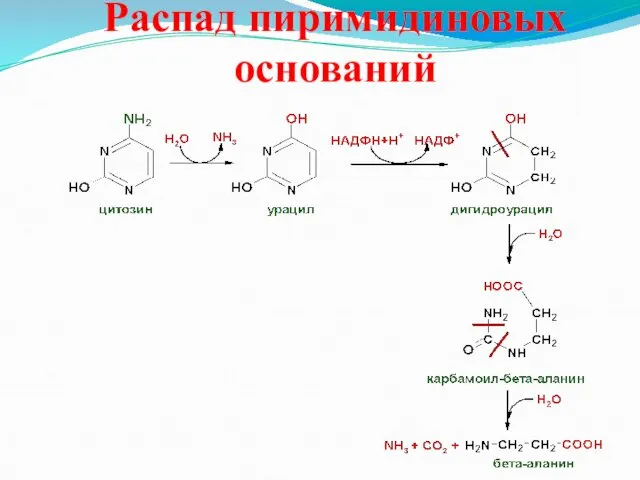
Распад пиримидиновых оснований
Распад пиримидиновых оснований
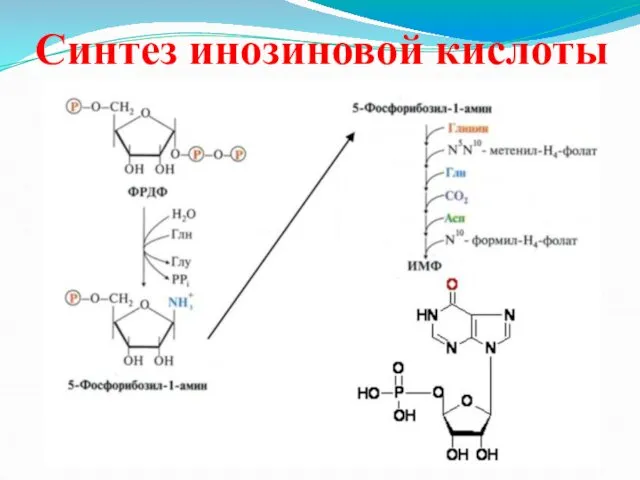
Синтез инозиновой кислоты
Синтез инозиновой кислоты
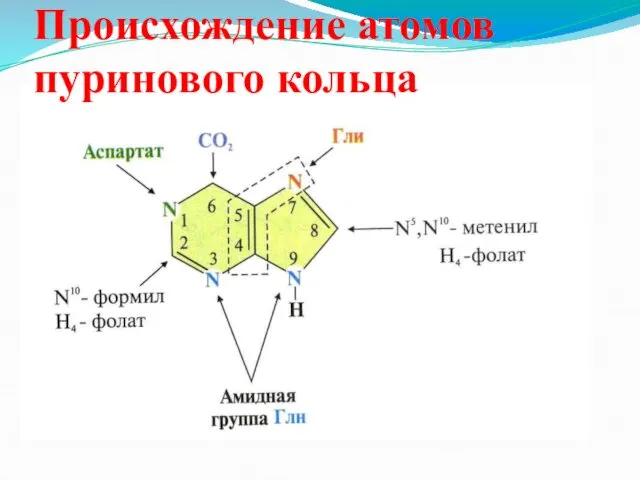
Происхождение атомов пуринового кольца
Происхождение атомов пуринового кольца
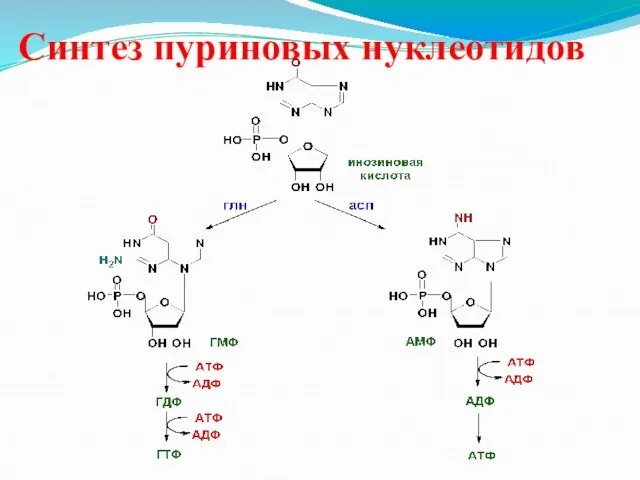
Синтез пуриновых нуклеотидов
Синтез пуриновых нуклеотидов
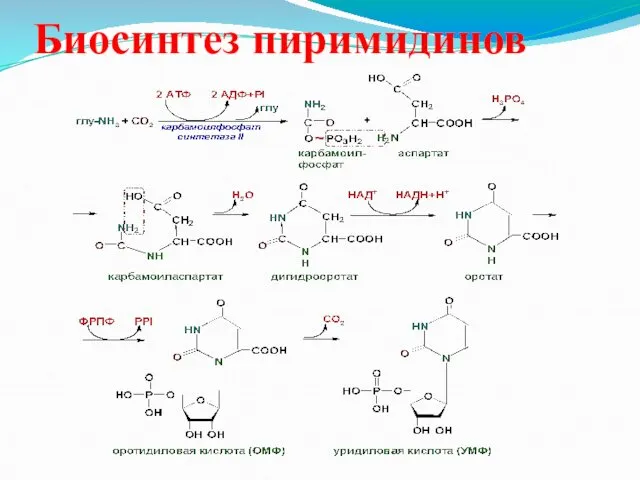
Биосинтез пиримидинов
Биосинтез пиримидинов
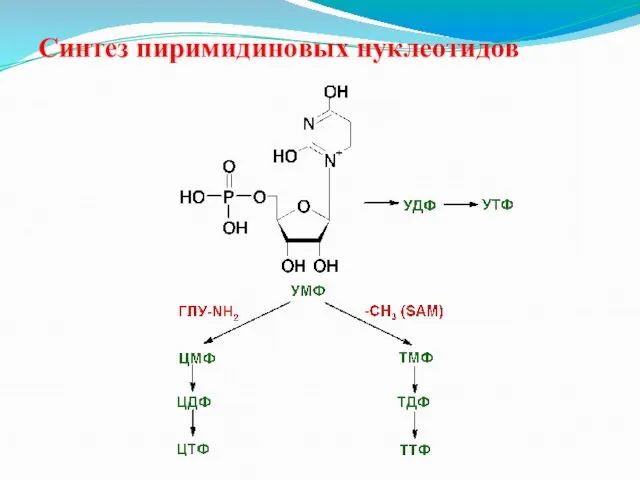
Синтез пиримидиновых нуклеотидов
Синтез пиримидиновых нуклеотидов
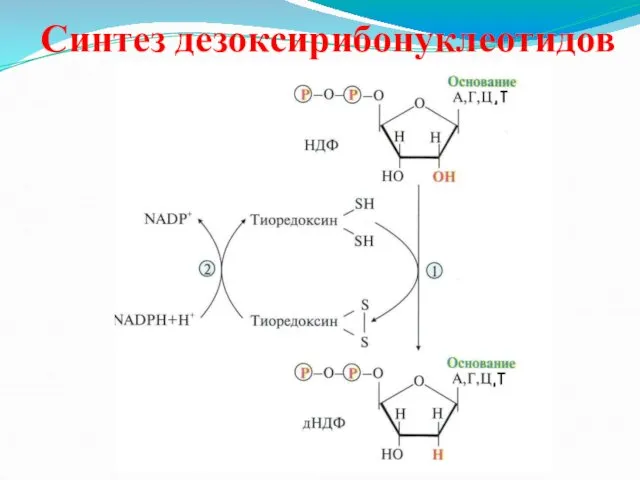
Синтез дезоксирибонуклеотидов
Синтез дезоксирибонуклеотидов
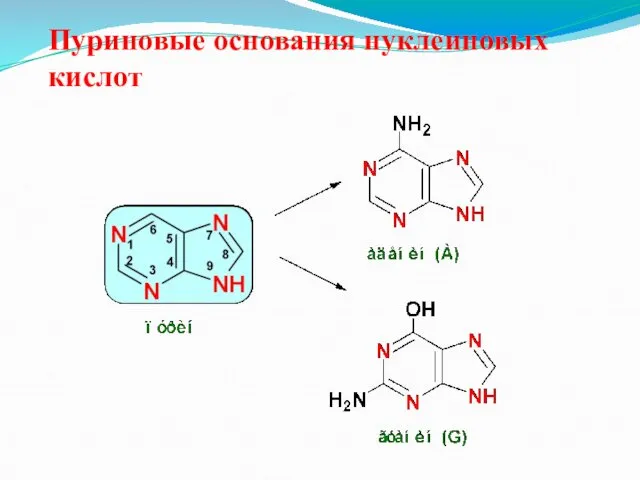
Пуриновые основания нуклеиновых кислот
Пуриновые основания нуклеиновых кислот
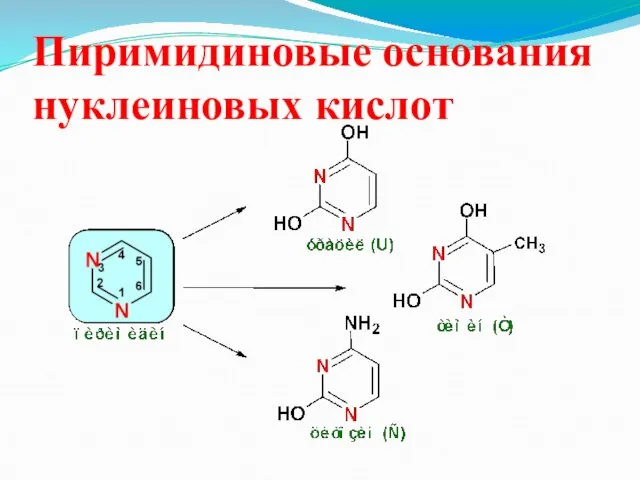
Пиримидиновые основания нуклеиновых кислот
Пиримидиновые основания нуклеиновых кислот
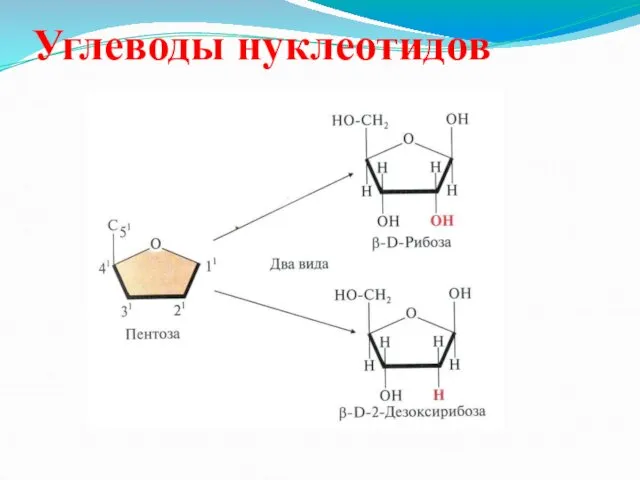
Углеводы нуклеотидов
Углеводы нуклеотидов
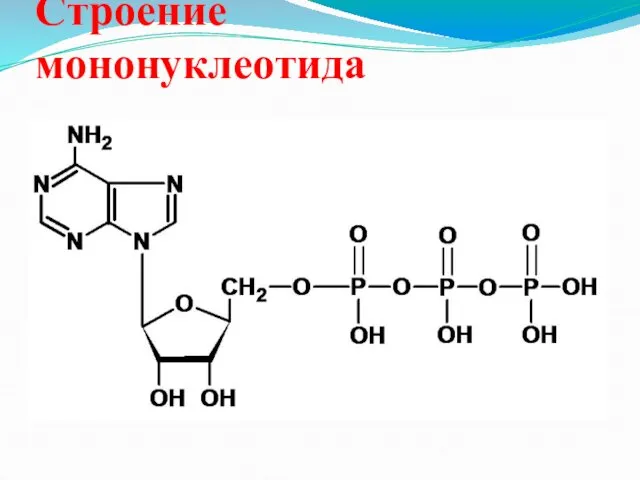
Строение мононуклеотида
Строение мононуклеотида
 Проект Выращивание микрозелени
Проект Выращивание микрозелени Приспособленность организмов к среде обитания
Приспособленность организмов к среде обитания Зоопсихология. Методы зоопсихологических исследований
Зоопсихология. Методы зоопсихологических исследований Животные и растения России. Белка и пион
Животные и растения России. Белка и пион Corynebacterium Erysipelothrix & Listeria
Corynebacterium Erysipelothrix & Listeria Углеводы. Функции углеводов
Углеводы. Функции углеводов Грибы-паразиты
Грибы-паразиты Роль пчелы
Роль пчелы Стихи, загадки, песни про птиц
Стихи, загадки, песни про птиц Сүйектің байланысу түрлері
Сүйектің байланысу түрлері ГМО в нашей жизни
ГМО в нашей жизни Мышечная и нервная ткани. Лекция № 4
Мышечная и нервная ткани. Лекция № 4 Родительское поведение животных
Родительское поведение животных Плодовые овощи
Плодовые овощи Семейство куньих. Горностай
Семейство куньих. Горностай Половая система человека
Половая система человека Подцарство Простейшие
Подцарство Простейшие Значение и биология пчёл
Значение и биология пчёл Микроорганизімдер систематикасы
Микроорганизімдер систематикасы Биологический словарь
Биологический словарь Генетика микроорганизмов
Генетика микроорганизмов о вреде мобильных телефонов
о вреде мобильных телефонов Череп. Общий обзор
Череп. Общий обзор Организм как целостная система
Организм как целостная система Пингвины
Пингвины Опасная флора и фауна
Опасная флора и фауна Стрекозы. Виды стрекоз
Стрекозы. Виды стрекоз Методика фитоценотических или геоботанических исследований
Методика фитоценотических или геоботанических исследований