- Обмен и функции липидов

Содержание
- 2. Важнейшие липиды тканей человека Жиры – триацилглицеролы (основная масса липидов организма), служащие формой депонирования энергии. Жиры
- 3. Пищевые жиры. Их переваривание и всасывание
- 4. Ресинтез триацилглицерола в стенке кишечника
- 5. Строение транспортных липопротеинов крови
- 6. Липопротеины – транспортные формы липидов в организме человека
- 7. Образование хиломикронов и транспорт жиров
- 8. Нарушение переваривания и всасывания жиров Причины нарушения переваривания и всасывания жиров: нарушение секреции желчи из желчного
- 9. Обмен жирных кислот Жирные кислоты поступают с пищей или синтезируются в организме (кроме полиеновых кислот).Субстраты, необходимые
- 10. Реакция активации жирной кислоты (фермент – ацил-КоА-синтетаза) RCOOH+HS-KoA+АТФ → R-CO-SKoA+АМФ+H4P2O7 Перенос жирных кислот через мембраны митохондрий
- 11. Реакции β-окисления жирных кислот CH3-(CH2)14-CO-SKoA+7ФАД+H2O+7НАД*+7HSKoA → → 8CH3-CO-SKoA+7ФАДH2+7(НАДH+H*)
- 12. Кетоновые тела При голодании, длительной физической работе и в случаях, когда клетки не получают достаточного количества
- 13. Синтез кетоновых тел при голодании
- 14. Синтез кетоновых тел
- 15. Физиологическое значение синтеза ацетоацетата При голодании, длительной физической работе и в случаях, когда клетки не получают
- 16. Использование кетоновых тел
- 17. Перенос ацетильных остатков из митохондрий в цитозоль БИОСИНТЕЗ ВЫСШИХ ЖИРНЫХ КИСЛОТ Превращение ацетил-КоА в малонил-КоА –
- 18. СИНТЕЗ ПАЛЬМИТИНОВОЙ КИСЛОТЫ
- 19. Общее уравнение CH3-CO-SKoA + 7HOOC-CH2-CO-SKoA + 14 (НАДФН+Н*) → → CH3-(CH2)14-COOH + НАДФ* + 6H2O +
- 20. Регуляция ацетил-КоА-карбоксилазы
- 21. Обмен жиров Жиры – наиболее выгодная и основная форма депонирования энергии. Депонированный жир может обеспечивать организм
- 22. Синтез жиров в печени и жировой ткани
- 23. Депонирование жиров в жировой ткани
- 24. Мобилизация жиров из жировой ткани Адипоциты (место депонирования жиров) располагаются в основном под кожей, образуя подкожный
- 25. Гормональная регуляция мобилизации жиров
- 26. Роль резервирования и мобилизации жиров Запасание жиров в жировой ткани – основная форма депонирования источников энергии
- 27. Синтез и секреция ЛПОНП
- 28. Гиперлипротеинемия Гиперлипопротеинемия – повышенный уровень в крови липопротеинов (в основном, ЛПОНП).Является характерным признаком сахарного диабета. При
- 29. Нарушение синтеза и мобилизации жиров при ожирении Избыточное накопление жира в адипоцитах (ожирение) широко распространено. Среди
- 30. Обмен стероидов
- 32. Стероиды - производные восстановленных конденсированных циклических систем - циклопентанпергидрофенантренов. В организме человека основной стероид - холестерол,
- 33. Холестерин как предшественник ряда других стероидов. Холестерин — воскообразное жирорастворимое вещество, присутствующее во всех тканях организма,
- 34. Фонд холестерина в организме и пути его использования и выведения
- 35. Синтез холестерола
- 37. Биосинтез холестерина 1.Ланостерин превращается в мембранах ГЭПР в холестерин. Метильная группа при С14 окисляется, и образуется
- 38. Восстановление гидроксиметилглутарил-КоА (ГМГ) в мевалоновую кислоту
- 39. Регуляция синтеза и активности ГМГ-редуктазы
- 40. Ингибирование синтеза ГМГ КоА-редуктазы Конечный продукт метаболического пути (холестерол) снижает скорость транскрипции гена ГМГ-КоА-редуктазы, подавляя таким
- 41. Превращение холестерола в желчные кислоты
- 42. Выведение холестерола из организма Структурная основа холестерола – кольца циклопентанпергидрофенантрена - не может быть расщеплена до
- 43. Обмен транспортных липопротеинов
- 44. Механизм и маршруты транспорта жиров и холестерина
- 45. Гиперхолестеринемия и её причины Концентрация холестерола в крови взрослых людей составляет 200±50 мг/дл (5,2±1,2 ммоль/л) и,
- 46. Механизм возникновения желчекаменной болезни (холестериновые камни) Желчнокаменная болезнь - патологический процесс, при котором в жёлчном пузыре
- 47. Применение хенодезоксихолевой кислоты для лечения желчекаменной болезни В начальной стадии образования камней можно применять в качестве
- 48. Биохимия атеросклероза Нарушения обмена холестерола чаще всего приводят к гиперхолестеролемии и последующему развитию атеросклероза. При атеросклерозе
- 49. Механизм образования атеросклеротических бляшек
- 50. Дислипопротеинемии
- 51. Биохимические основы лечения гиперхолестеринемии и атеросклероза К лечебным и профилактическим факторам относят обогащение пищи полиеновыми жирными
- 52. Основные фосфолипиды и гликолипиды тканей человека
- 53. Представления о биосинтезе и катаболизме этих соединений
- 56. Функции гликосфинголипидов Взаимодействие между: клетками; клетками и межклеточным матриксом; клетками и микробами. Модуляция: активности протеинкиназ; активности
- 57. Сфинголипидозы Генетические дефекты лизосомных ферментов катаболизма гликосфинголипидов. В норме синтез и катаболизм гликосфинголипидов сбалансированы таким образом,
- 58. Характеристики липидного состава диеты и потребности в липидах детей разного возраста
- 59. Бурая жировая ткань Бурые жировые клетки участвуют в образовании бурой жировой ткани. Эти клетки имеют полигональную
- 60. Функции бурой жировой ткани Главная функция ее - теплопродукция. Бурая жировая ткань поддерживает температуру тела животных
- 61. Особенности процессов переваривания и всасывания липидов в постнатальном периоде У грудных детей и детей младшего возраста
- 62. Окисление кетоновых тел в периферических тканях
- 64. Скачать презентацию

Важнейшие липиды тканей человека
Жиры – триацилглицеролы (основная масса липидов организма), служащие
Важнейшие липиды тканей человека
Жиры – триацилглицеролы (основная масса липидов организма), служащие
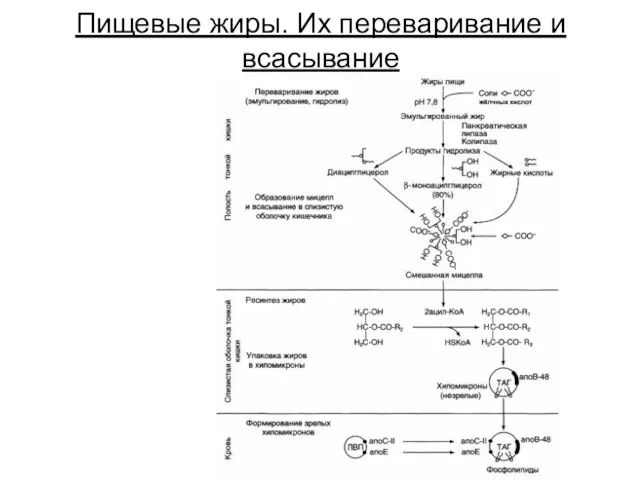
Пищевые жиры. Их переваривание и всасывание
Пищевые жиры. Их переваривание и всасывание
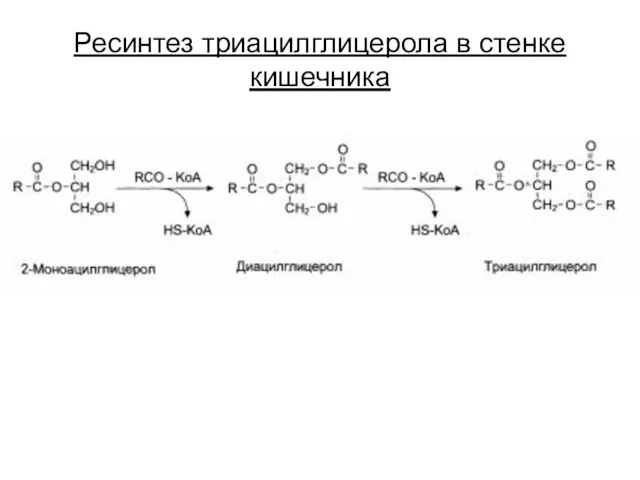
Ресинтез триацилглицерола в стенке кишечника
Ресинтез триацилглицерола в стенке кишечника
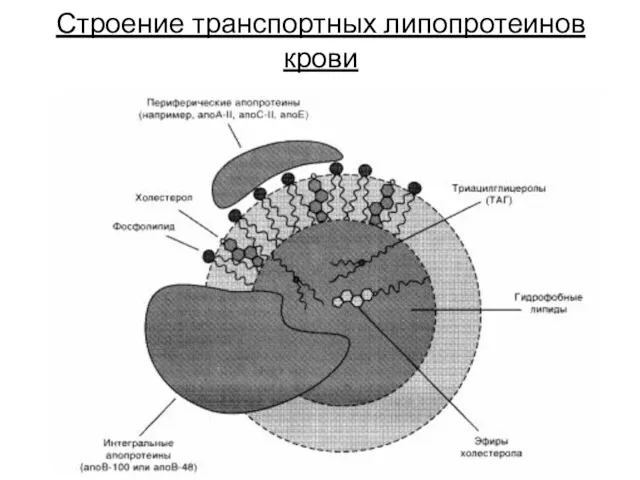
Строение транспортных липопротеинов крови
Строение транспортных липопротеинов крови
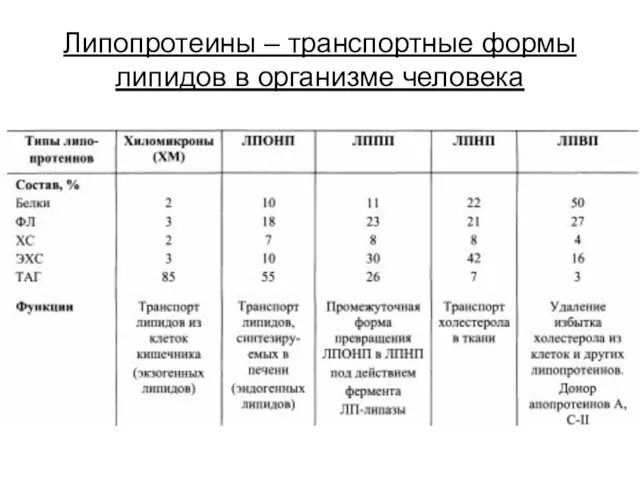
Липопротеины – транспортные формы липидов в организме человека
Липопротеины – транспортные формы липидов в организме человека
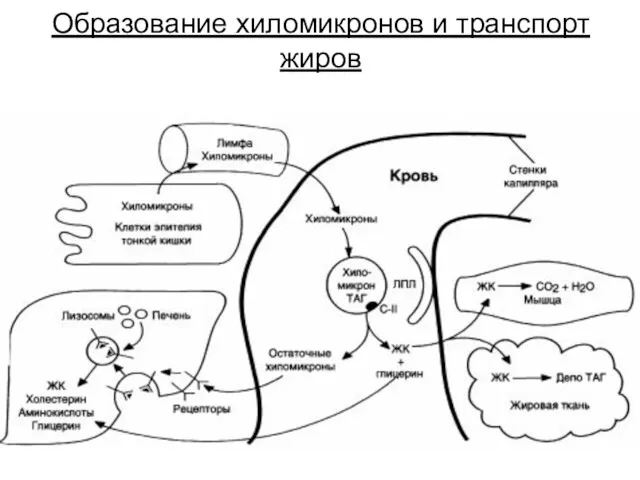
Образование хиломикронов и транспорт жиров
Образование хиломикронов и транспорт жиров

Нарушение переваривания и всасывания жиров
Причины нарушения переваривания и всасывания жиров:
нарушение
Нарушение переваривания и всасывания жиров
Причины нарушения переваривания и всасывания жиров:
нарушение

Обмен жирных кислот
Жирные кислоты поступают с пищей или синтезируются в организме
Обмен жирных кислот
Жирные кислоты поступают с пищей или синтезируются в организме
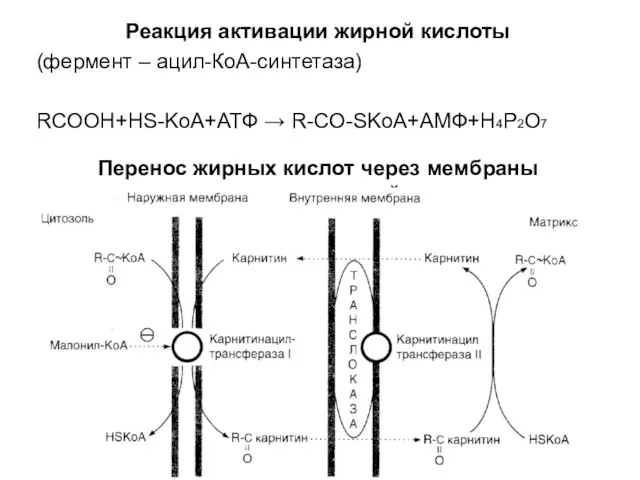
Реакция активации жирной кислоты
(фермент – ацил-КоА-синтетаза)
RCOOH+HS-KoA+АТФ → R-CO-SKoA+АМФ+H4P2O7
Перенос жирных кислот
Реакция активации жирной кислоты
(фермент – ацил-КоА-синтетаза)
RCOOH+HS-KoA+АТФ → R-CO-SKoA+АМФ+H4P2O7
Перенос жирных кислот
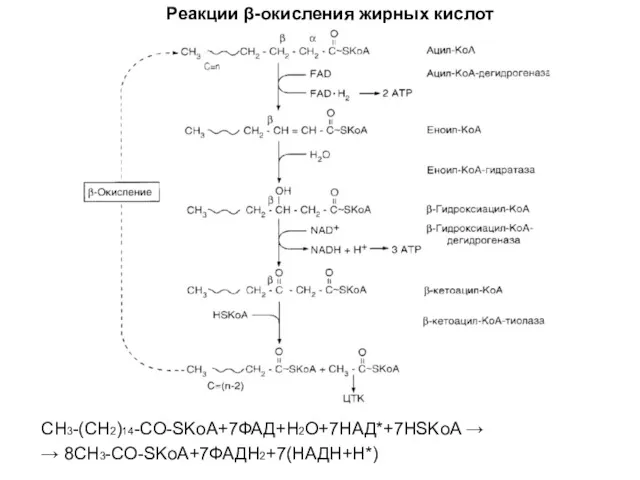
Реакции β-окисления жирных кислот
CH3-(CH2)14-CO-SKoA+7ФАД+H2O+7НАД*+7HSKoA →
→ 8CH3-CO-SKoA+7ФАДH2+7(НАДH+H*)
Реакции β-окисления жирных кислот
CH3-(CH2)14-CO-SKoA+7ФАД+H2O+7НАД*+7HSKoA →
→ 8CH3-CO-SKoA+7ФАДH2+7(НАДH+H*)

Кетоновые тела
При голодании, длительной физической работе и в случаях, когда клетки
Кетоновые тела
При голодании, длительной физической работе и в случаях, когда клетки
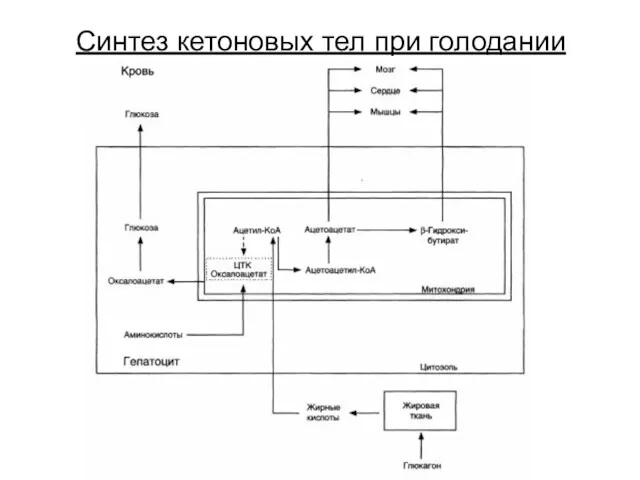
Синтез кетоновых тел при голодании
Синтез кетоновых тел при голодании
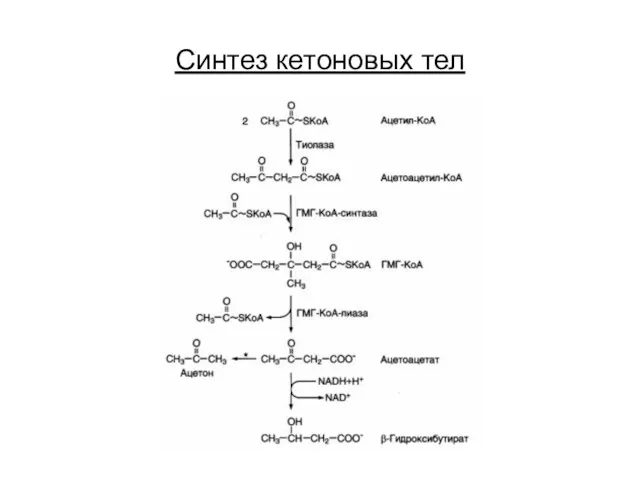
Синтез кетоновых тел
Синтез кетоновых тел

Физиологическое значение синтеза ацетоацетата
При голодании, длительной физической работе и в случаях,
Физиологическое значение синтеза ацетоацетата
При голодании, длительной физической работе и в случаях,

Использование кетоновых тел
Использование кетоновых тел
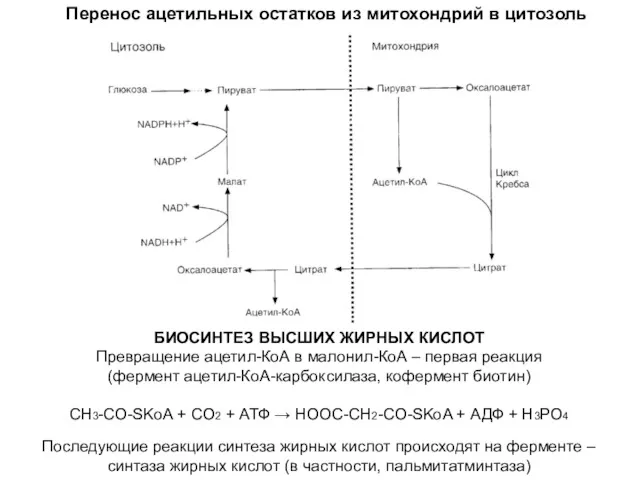
Перенос ацетильных остатков из митохондрий в цитозоль
БИОСИНТЕЗ ВЫСШИХ ЖИРНЫХ КИСЛОТ
Превращение ацетил-КоА
Перенос ацетильных остатков из митохондрий в цитозоль
БИОСИНТЕЗ ВЫСШИХ ЖИРНЫХ КИСЛОТ
Превращение ацетил-КоА
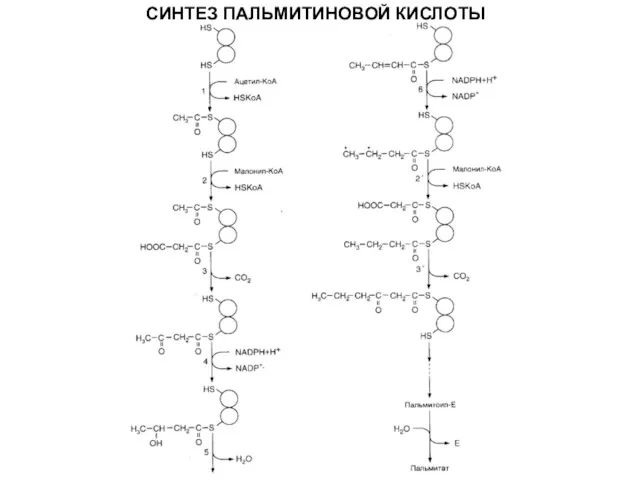
СИНТЕЗ ПАЛЬМИТИНОВОЙ КИСЛОТЫ
СИНТЕЗ ПАЛЬМИТИНОВОЙ КИСЛОТЫ
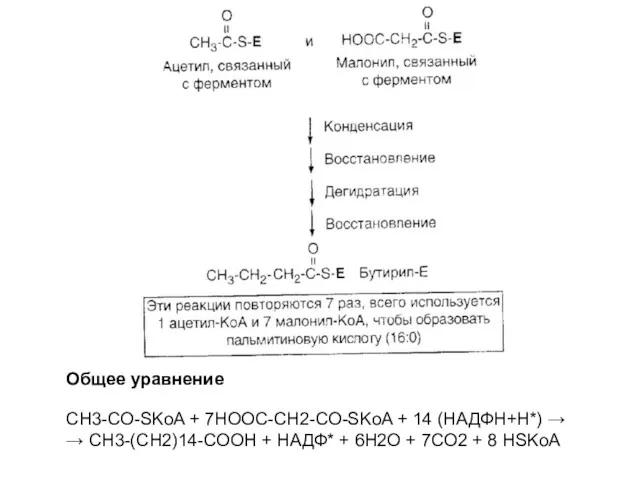
Общее уравнение
CH3-CO-SKoA + 7HOOC-CH2-CO-SKoA + 14 (НАДФН+Н*) →
→ CH3-(CH2)14-COOH +
Общее уравнение
CH3-CO-SKoA + 7HOOC-CH2-CO-SKoA + 14 (НАДФН+Н*) →
→ CH3-(CH2)14-COOH +
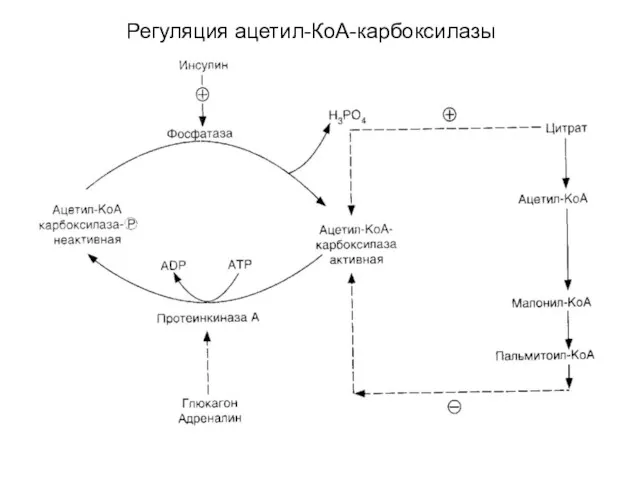
Регуляция ацетил-КоА-карбоксилазы
Регуляция ацетил-КоА-карбоксилазы

Обмен жиров
Жиры – наиболее выгодная и основная форма депонирования энергии. Депонированный
Обмен жиров
Жиры – наиболее выгодная и основная форма депонирования энергии. Депонированный
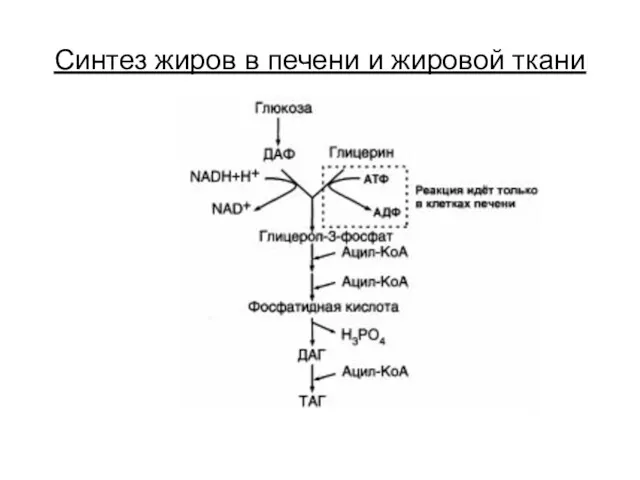
Синтез жиров в печени и жировой ткани
Синтез жиров в печени и жировой ткани
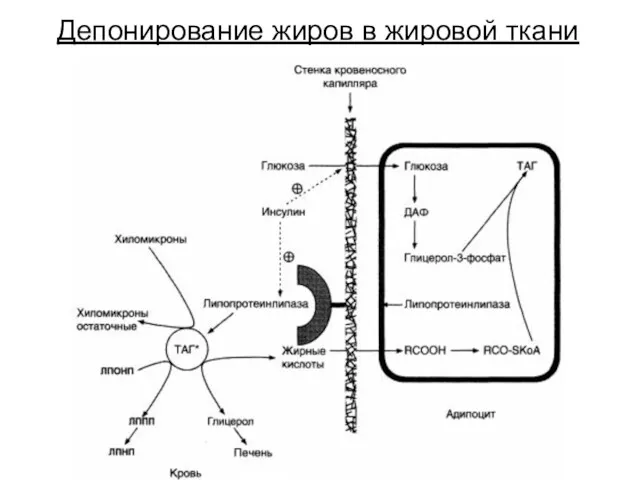
Депонирование жиров в жировой ткани
Депонирование жиров в жировой ткани

Мобилизация жиров из жировой ткани
Адипоциты (место депонирования жиров) располагаются в основном
Мобилизация жиров из жировой ткани
Адипоциты (место депонирования жиров) располагаются в основном
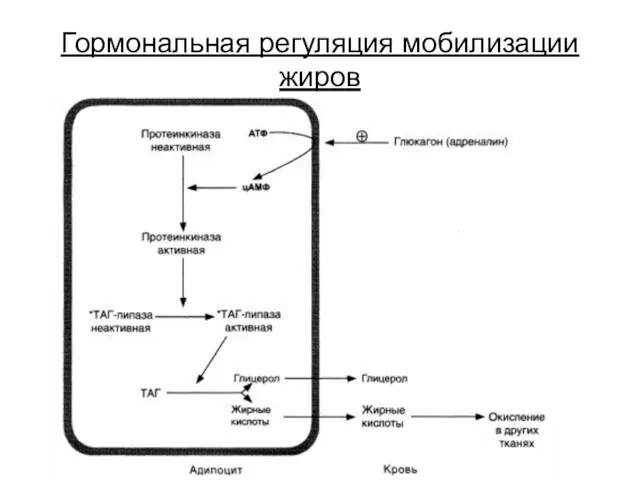
Гормональная регуляция мобилизации жиров
Гормональная регуляция мобилизации жиров

Роль резервирования и мобилизации жиров
Запасание жиров в жировой ткани –
Роль резервирования и мобилизации жиров
Запасание жиров в жировой ткани –
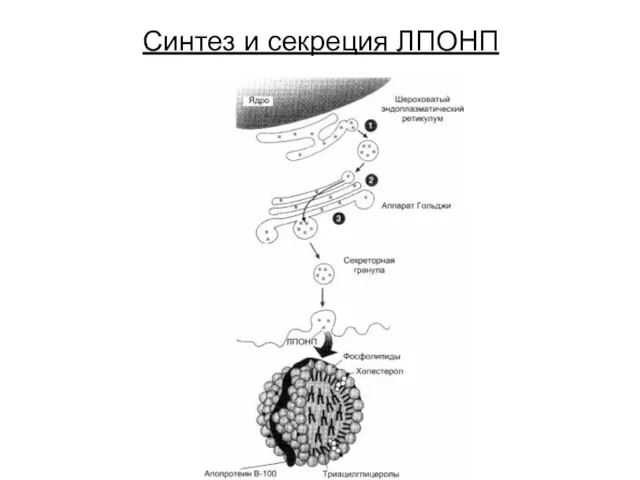
Синтез и секреция ЛПОНП
Синтез и секреция ЛПОНП

Гиперлипротеинемия
Гиперлипопротеинемия – повышенный уровень в крови липопротеинов (в основном, ЛПОНП).Является характерным
Гиперлипротеинемия
Гиперлипопротеинемия – повышенный уровень в крови липопротеинов (в основном, ЛПОНП).Является характерным

Нарушение синтеза и мобилизации жиров при ожирении
Избыточное накопление жира в адипоцитах
Нарушение синтеза и мобилизации жиров при ожирении
Избыточное накопление жира в адипоцитах

Обмен стероидов
Обмен стероидов


Стероиды - производные восстановленных конденсированных циклических систем - циклопентанпергидрофенантренов.
В организме
В организме

Холестерин как предшественник ряда других стероидов.
Холестерин — воскообразное жирорастворимое
Холестерин как предшественник ряда других стероидов.
Холестерин — воскообразное жирорастворимое
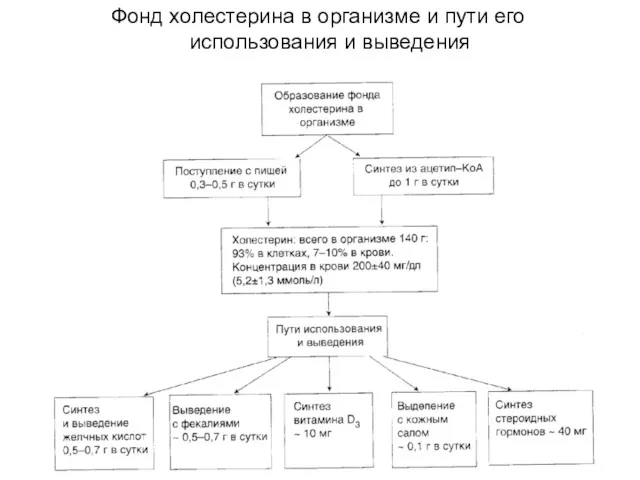
Фонд холестерина в организме и пути его использования и выведения
Фонд холестерина в организме и пути его использования и выведения
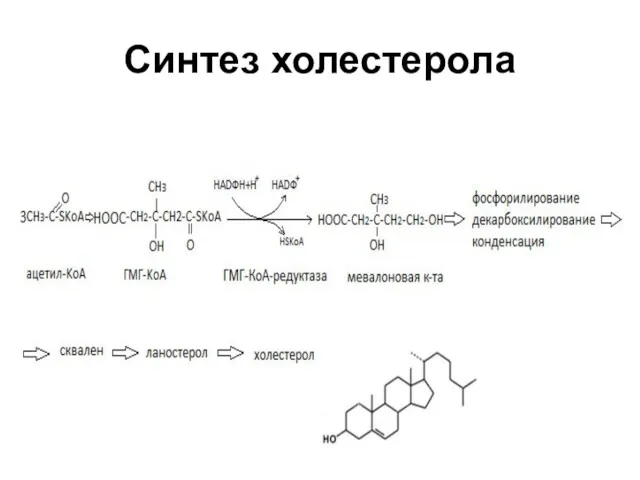
Синтез холестерола
Синтез холестерола
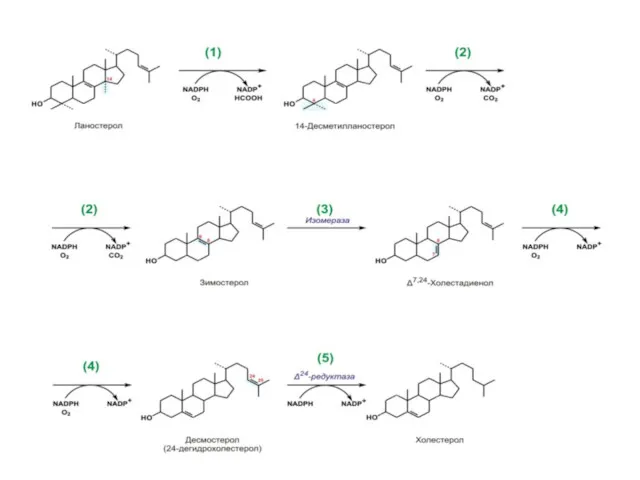

Биосинтез холестерина
1.Ланостерин превращается в мембранах ГЭПР в холестерин.
Метильная группа при С14
Биосинтез холестерина
1.Ланостерин превращается в мембранах ГЭПР в холестерин.
Метильная группа при С14
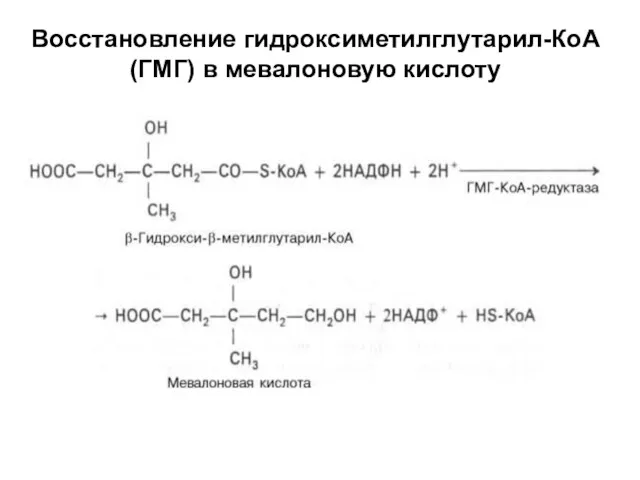
Восстановление гидроксиметилглутарил-КоА (ГМГ) в мевалоновую кислоту
Восстановление гидроксиметилглутарил-КоА (ГМГ) в мевалоновую кислоту
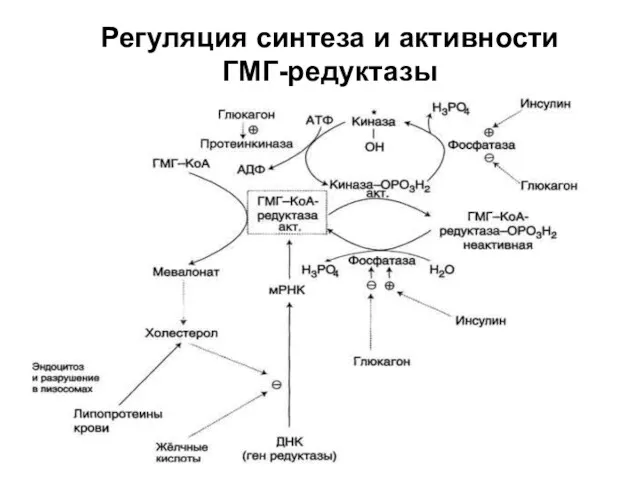
Регуляция синтеза и активности ГМГ-редуктазы
Регуляция синтеза и активности ГМГ-редуктазы

Ингибирование синтеза
ГМГ КоА-редуктазы
Конечный продукт метаболического пути (холестерол) снижает
Ингибирование синтеза
ГМГ КоА-редуктазы
Конечный продукт метаболического пути (холестерол) снижает
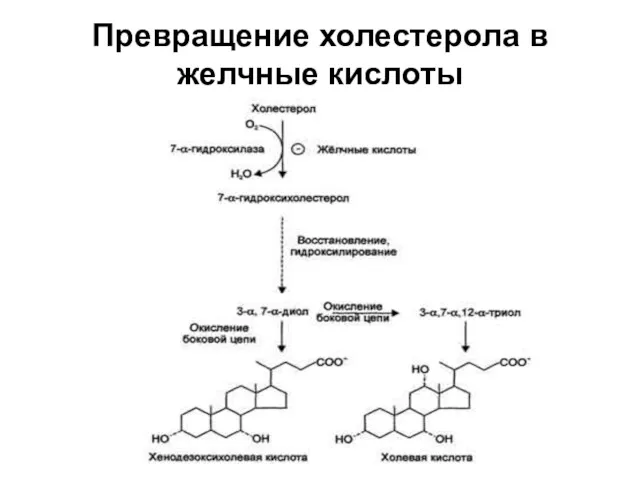
Превращение холестерола в желчные кислоты
Превращение холестерола в желчные кислоты

Выведение холестерола из организма
Структурная основа холестерола – кольца циклопентанпергидрофенантрена
Выведение холестерола из организма
Структурная основа холестерола – кольца циклопентанпергидрофенантрена
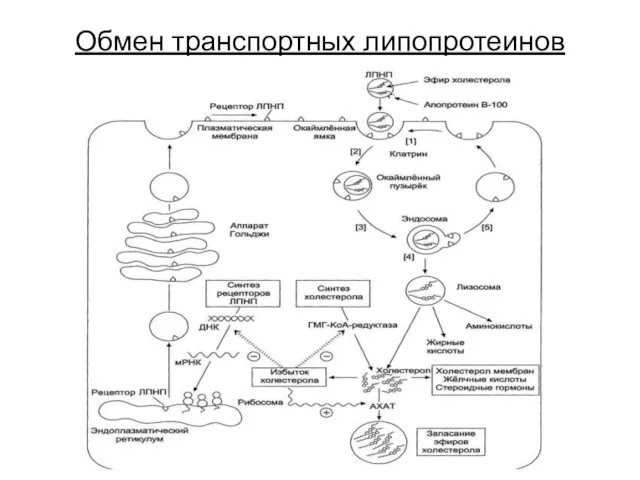
Обмен транспортных липопротеинов
Обмен транспортных липопротеинов
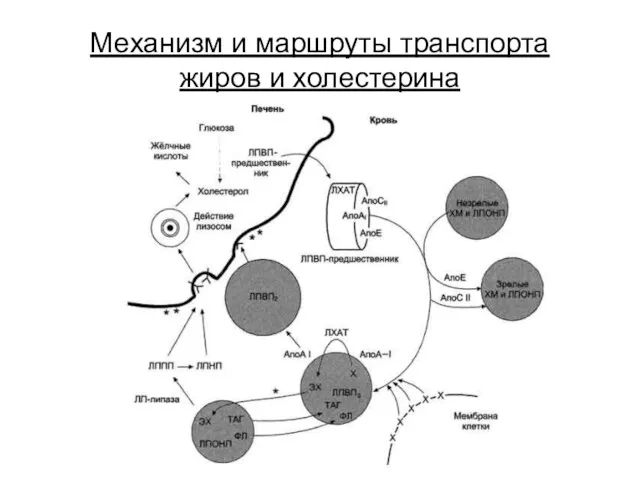
Механизм и маршруты транспорта жиров и холестерина
Механизм и маршруты транспорта жиров и холестерина

Гиперхолестеринемия и её причины
Концентрация холестерола в крови взрослых людей составляет 200±50
Гиперхолестеринемия и её причины
Концентрация холестерола в крови взрослых людей составляет 200±50

Механизм возникновения желчекаменной болезни (холестериновые камни)
Желчнокаменная болезнь - патологический процесс, при
Механизм возникновения желчекаменной болезни (холестериновые камни)
Желчнокаменная болезнь - патологический процесс, при

Применение хенодезоксихолевой кислоты для лечения желчекаменной болезни
В начальной стадии образования камней
Применение хенодезоксихолевой кислоты для лечения желчекаменной болезни
В начальной стадии образования камней

Биохимия атеросклероза
Нарушения обмена холестерола чаще всего приводят к гиперхолестеролемии и последующему
Биохимия атеросклероза
Нарушения обмена холестерола чаще всего приводят к гиперхолестеролемии и последующему
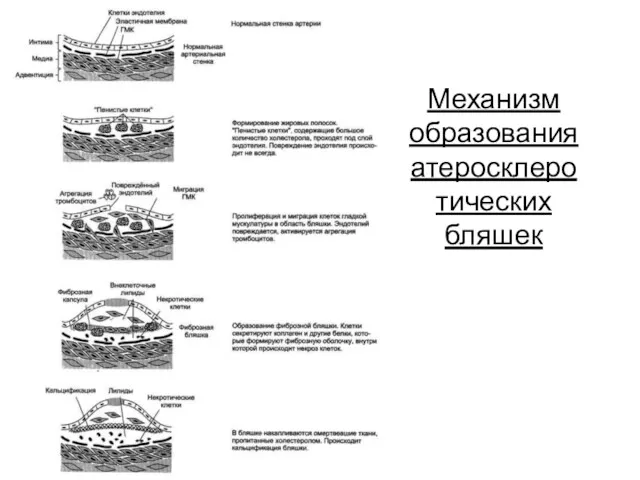
Механизм образования атеросклеротических бляшек
Механизм образования атеросклеротических бляшек
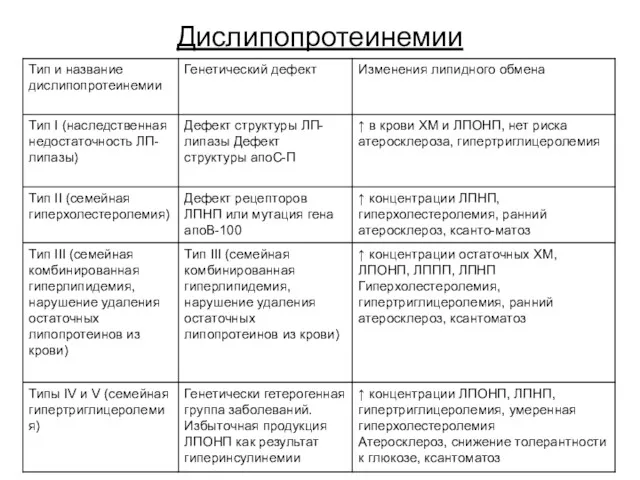
Дислипопротеинемии
Дислипопротеинемии

Биохимические основы лечения гиперхолестеринемии и атеросклероза
К лечебным и профилактическим факторам
Биохимические основы лечения гиперхолестеринемии и атеросклероза
К лечебным и профилактическим факторам
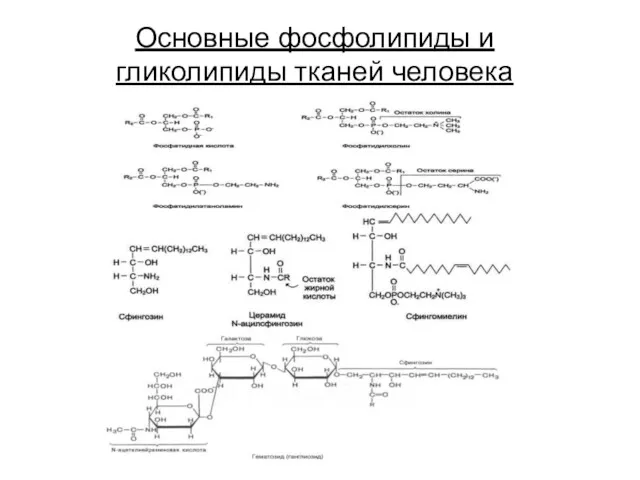
Основные фосфолипиды и гликолипиды тканей человека
Основные фосфолипиды и гликолипиды тканей человека
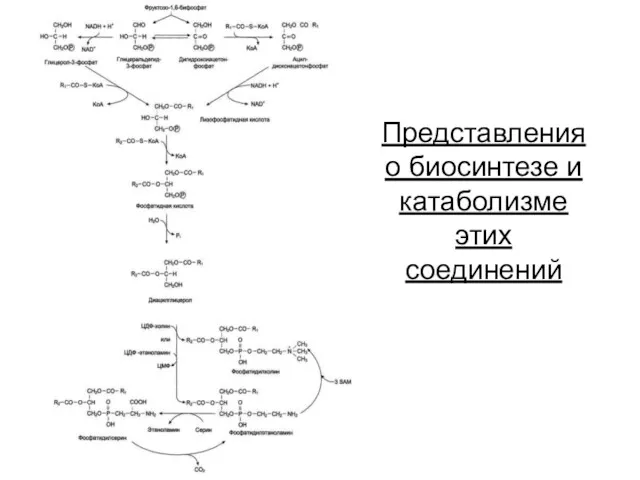
Представления о биосинтезе и катаболизме этих соединений
Представления о биосинтезе и катаболизме этих соединений
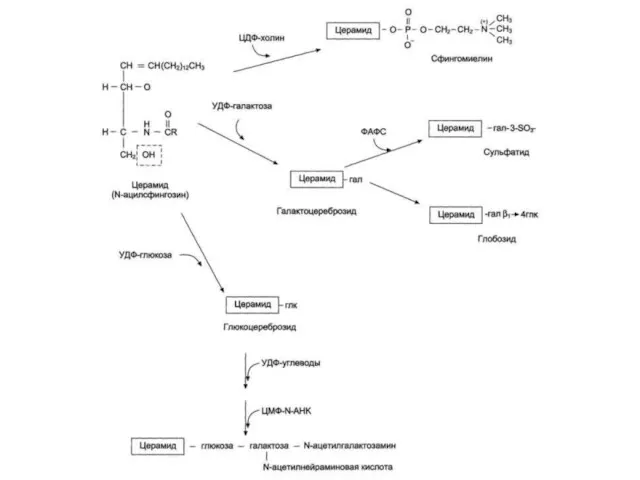

Функции гликосфинголипидов
Взаимодействие между:
клетками;
клетками и межклеточным матриксом;
клетками и микробами.
Модуляция:
активности
Функции гликосфинголипидов
Взаимодействие между:
клетками;
клетками и межклеточным матриксом;
клетками и микробами.
Модуляция:
активности

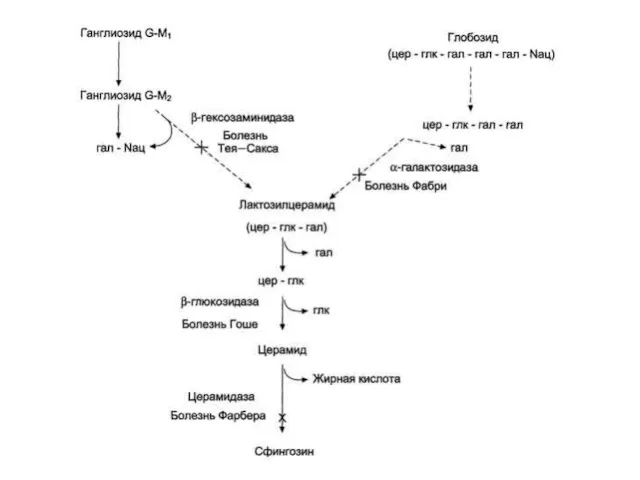
Сфинголипидозы
Генетические дефекты лизосомных ферментов катаболизма гликосфинголипидов. В норме синтез и катаболизм
Сфинголипидозы
Генетические дефекты лизосомных ферментов катаболизма гликосфинголипидов. В норме синтез и катаболизм
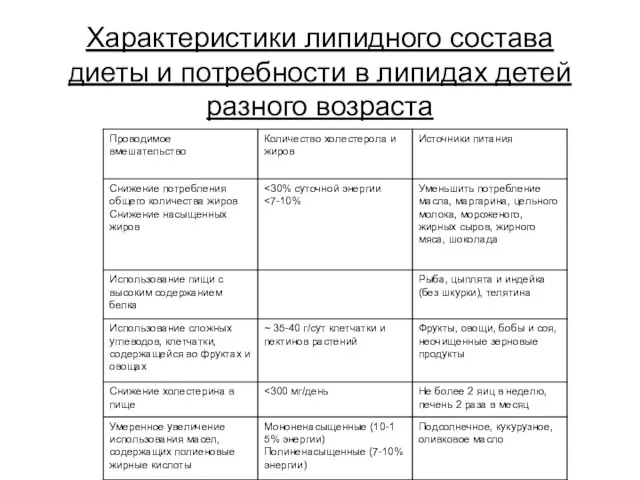
Характеристики липидного состава диеты и потребности в липидах детей разного возраста
Характеристики липидного состава диеты и потребности в липидах детей разного возраста

Бурая жировая ткань
Бурые жировые клетки участвуют в образовании бурой жировой ткани.
Бурая жировая ткань
Бурые жировые клетки участвуют в образовании бурой жировой ткани.

Функции бурой жировой ткани
Главная функция ее - теплопродукция. Бурая жировая ткань
Функции бурой жировой ткани
Главная функция ее - теплопродукция. Бурая жировая ткань

Особенности процессов переваривания и всасывания липидов в постнатальном периоде
У грудных
Особенности процессов переваривания и всасывания липидов в постнатальном периоде
У грудных
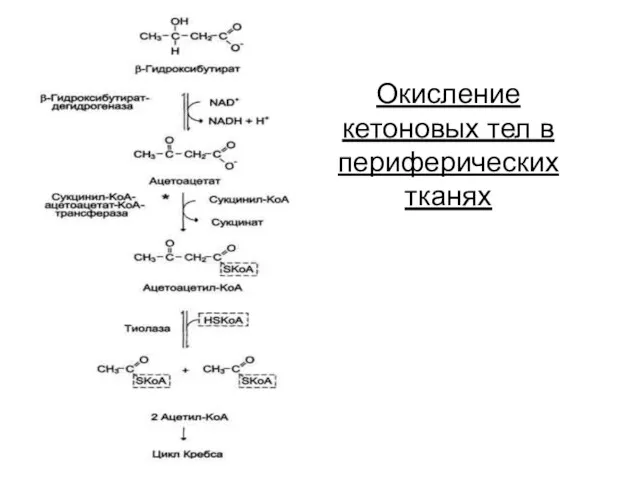
Окисление кетоновых тел в периферических тканях
Окисление кетоновых тел в периферических тканях
 Мышцы/ Основные группы мышц
Мышцы/ Основные группы мышц Введение в биологию. Многообразие живого мира. Уровни организации живой материи
Введение в биологию. Многообразие живого мира. Уровни организации живой материи Специфические пути обмена отдельных аминокислот. Патология. Лекция №12
Специфические пути обмена отдельных аминокислот. Патология. Лекция №12 Многообразие видов как результат эволюции. Урок-экскурсия в зоологический музей
Многообразие видов как результат эволюции. Урок-экскурсия в зоологический музей Плоские черви. Строение и жизненные циклы представителей класса trematoda
Плоские черви. Строение и жизненные циклы представителей класса trematoda Млекопитающие отряд приматы
Млекопитающие отряд приматы Проблема наркомании
Проблема наркомании Chondrichthyes. Хрящевые рыбы
Chondrichthyes. Хрящевые рыбы Птицы. Водно-подводные птицы, водоплавающие птицы, водно-воздушные птицы, наземно-водные птицы
Птицы. Водно-подводные птицы, водоплавающие птицы, водно-воздушные птицы, наземно-водные птицы Деревья. Удивительные факты из жизни деревьев
Деревья. Удивительные факты из жизни деревьев Гаметаларды in vivo культивирлеу
Гаметаларды in vivo культивирлеу Теория антропосоциогенеза Б. Поршнева
Теория антропосоциогенеза Б. Поршнева ВКР: Толерантность гибридов кукурузы к условиям Орловской области
ВКР: Толерантность гибридов кукурузы к условиям Орловской области Биотехнологические основы культивирования микроорганизмов
Биотехнологические основы культивирования микроорганизмов Зоопарк (зоологічний парк)
Зоопарк (зоологічний парк) Особенности строения среднего уха у детей
Особенности строения среднего уха у детей Основные методы селекции растений, животных, микроорганизмов
Основные методы селекции растений, животных, микроорганизмов Сүт және сүт тағамдары микрофлорасы
Сүт және сүт тағамдары микрофлорасы Хромосомалық аурулар
Хромосомалық аурулар Мышечные ткани. Возрастная гистология, регенерация
Мышечные ткани. Возрастная гистология, регенерация Строение органов дыхания
Строение органов дыхания особенности строения, функции и свойства липидов
особенности строения, функции и свойства липидов Різноманітність ссавців
Різноманітність ссавців Лекарственные растения
Лекарственные растения Fyziologie bunky
Fyziologie bunky Вестибулярный аппарат
Вестибулярный аппарат Морфология рыб. Форма рта и плавников
Морфология рыб. Форма рта и плавников Происхождение растений. Основные этапы развития растительного мира
Происхождение растений. Основные этапы развития растительного мира