- Обмен нуклеиновых кислот. Биосинтез белков

Содержание
- 3. ИСТОРИЯ открытия нуклеиновых кислот Мишер (1869г.) выделение ДНК из ядерного материала тимуса, селезенки и спермиев. Чаргафф
- 4. Строение нуклеиновых кислот Нуклеиновые кислоты – линейные полимеры, состоящие из нуклеотидов, соединенных 3-5 О-Р-О связями. Нуклеотиды
- 9. Пурины и пиримидины Азотистые основания – гетероциклические, плоские структуры, существуют в кето – и энольной форме,
- 10. Пространственная структура нуклеиновых кислот Первичная структура – последовательность нуклеотидов Вторичная структура – двойная спираль ДНК (А,В,С,Д
- 15. Внешний обмен нуклеиновых кислот Нуклеопротеины пищи в кислом желудочном соке распадаются на нуклеиновые кислоты и белки.
- 16. Метаболическая роль нуклеотидов Мономеры для синтеза ДНК и РНК Поддержание энергетического гомеостаза АДФ – АТФ (иногда
- 17. Катаболизм пуринов АМФ ?аденозин ?инозин ? гипоксантин ? ксантин ? мочевая кислота ГМФ ? гуанозин ?
- 19. Катаболизм пиримидинов ЦМФ ? УМФ ? урацил ТМФ ? тимин Восстановление и гидролиз пиримидинов ? раскрытие
- 21. Синтез нуклеотидов Синтез нуклеотидов лимитируется синтезом азотистых оснований de novo. Бьюкенен с помощью меченых атомов показал
- 23. Биосинтез пуринов На основе 5-фосфорибозил -1- пирофосфата строится имидазольное кольцо, затем пуриновое. Общий предшественник пуриновых нуклеотидов
- 29. Биосинтез пиримидинов Биосинтез пиримидинов начинается с построения азотистого гетероцикла с участием NH3,,СО2,глу, асп. Общий предшественник пиримидинов
- 33. Образование нуклеозидтрифосфатов АМФ + АТФ ? 2АДФ ГМФ + АТФ ? ГДФ + АДФ ГДФ +
- 34. Синтез дезоксинуклеотидов Все нуклеотиды образуются с участием фосфорибозилпирофосфата. Дезоксирибонуклеотиды образуются при восстановлении рибозы до дезоксирибозы в
- 37. Репликация ДНК Реакция матричного синтеза. Удвоение цепей ДНК, матрицей служит каждая из одноцепочечных последовательностей «материнской» ДНК.
- 39. Репликация ДНК Этапы: инициация, элонгация, терминация синтеза и созревание дочерней цепи (метилирование). Репарация ошибок и повреждений.
- 40. Репликация ДНК Этап инициации: Сигналом начала репликации служат белковые факторы роста (модифицирующие регуляторные белки?) Формирование одноцепочечных
- 41. Репликация ДНК Механизм реакции: Субстратами служат дезоксинуклеозидтрифосфаты 3-ОН группа дезоксирибозы (рибозы) производит нуклеофильную атаку α атома
- 43. Репликация ДНК Этап элонгации: Направление синтеза 5 ?3 ДНК – полимераза α синтезирует «затравку» (РНК- праймер)
- 44. Репликация ДНК Реплицируются одновременно обе одноцепочечные матрицы (5?3) Одна (лидирующая) цепь реплицируется непрерывно, вторая (отстающая) –
- 47. Репликация ДНК Скорость репликации огромна, т.к. реакция идет в нескольких местах одновременно – ориджины репликации. Сайты
- 50. Репликация ДНК ДНК- полимеразы Δ и ε делают 1 ошибку на 105 - 106 нуклеотидов (ДНК-полимераза
- 53. Репликация ДНК Ошибки в ДНК (мутации) возникают спонтанно (ошибки репликации, дезаминирование нуклеотидов, депуринизация ДНК и т.д.)
- 54. Репликация ДНК Комплекс ферментов репарации узнает и вырезает поврежденные и химически измененные нуклеотиды, ДНК-полимераза β встраивает
- 56. Репликация ДНК Количество раундов репликации ДНК (а значит число возможных делений клетки) зависит от длины теломерных
- 57. Репликация ДНК Созревание молекулы ДНК: Через несколько минут после завершения репликации происходит метилирование аденина (в –GATC-
- 60. Ингибиторы репликации Антибиотики (дауномицин, доксорубицин, рифампицин, актиномицин Д) способны встраиваться (интеркаляция) между основаниями ДНК, ингибируя ее
- 62. Транскрипция Считывание информации с ДНК-матрицы на РНК, синтез тРНК, иРНК, рРНК с помощью одной полимеразы (у
- 63. Транскрипция Механизм РНК – полимеразной реакции тот же, что и ДНК – полимеразной, направление синтеза 5?3,
- 64. Транскрипция В ДНК – матрице выделяют транскиптоны. Участки, ограниченные промоторами и сайтами терминации, между которыми 1
- 65. Транскрипция 3 стадии транскрипции: инициация, элонгация и терминация. Инициация синтеза начинается с «узнавания» полимеразой промоторного сайта
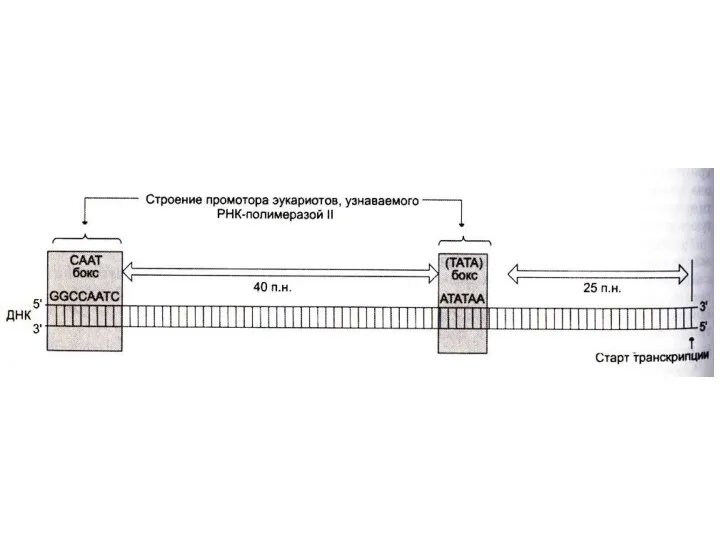
- 66. Инициация транскрипции Для формирование транскрипционной вилки (раскручивание одного витка спирали ДНК-матрицы) к ТАТА-боксу присоединяется белковый фактор
- 69. Элонгация транскрипции Белковые факторы элонгации обеспечивают расплетение ДНК перед продвижением РНК-полимеразы и восстановление двойной спирали позади
- 70. Терминация транскрипции При достижении РНК - полимеразой сайта терминации белковый фактор терминации освобождает пре-РНК из комплекса
- 71. Созревание РНК-транскриптов Процессингу (созреванию) подвергаются все виды РНК (и, т, р). А) Ковалентная модификация 5- и
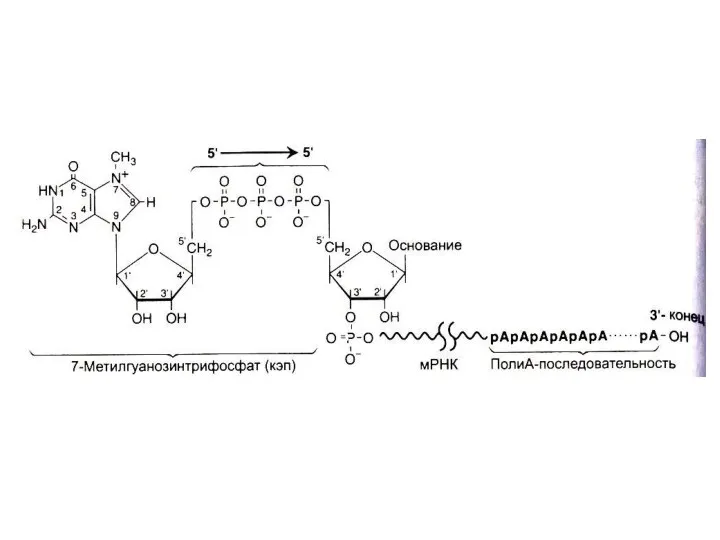
- 72. Ковалентная модификация иРНК Гуанилил-трансфераза присоединяет ГДФ к 5- ОР концу (5-О-Р-О-5 связь), 5 – кэпирование происходит
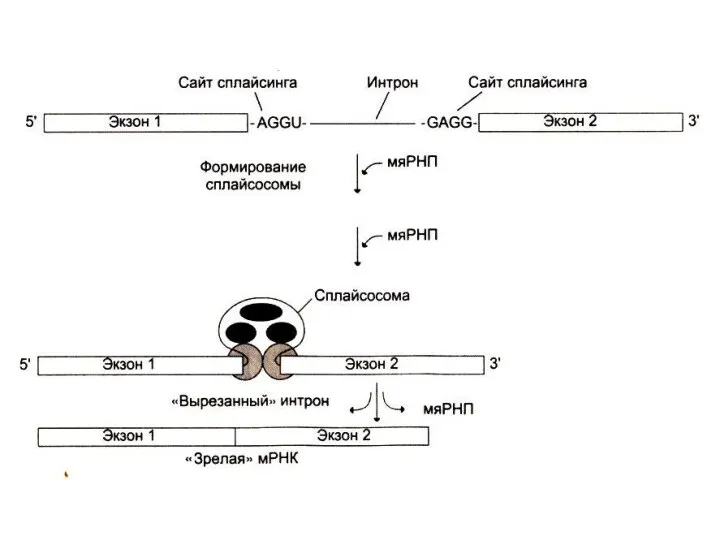
- 74. СПЛАЙСИНГ иРНК Сплайсинг: образование зрелой мРНК: Вырезание интронных последовательностей (ограниченных AGGU- и - GAGG- последовательностями) с
- 76. Процессинг первичных транскриптов тРНК РНК - аза отщепляет нуклеотиды с 3 – ОН конца до 3
- 78. Созревание рибосомальных РНК Образуется множество первичных транскриптов 5 S и 45 S. 45 S транскрипт в
- 79. Ингибиторы транскрипции Рифампицин связывается с β - субъединицей РНК –полимеразы, ингибируя образование первой фосфодиэфирной связи в
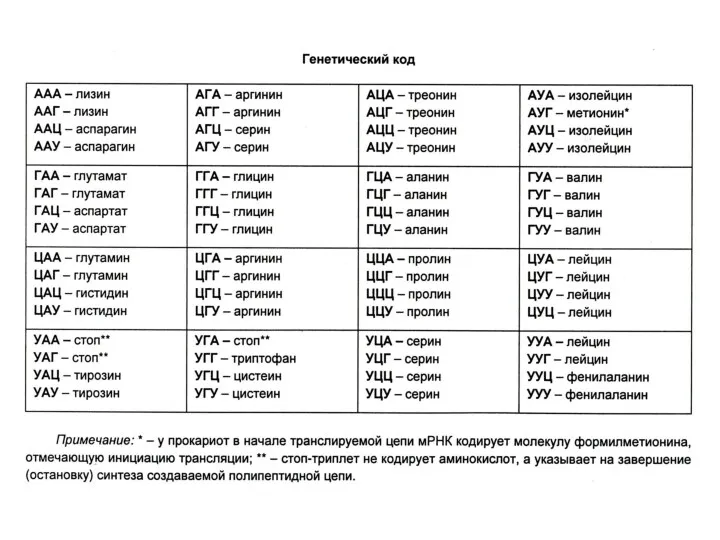
- 80. Трансляция Перевод генетической информации с кодонов мРНК на аминокислотную последовательность белка (экспрессия гена). Генетический код: триплетный,
- 81. Трансляция Что необходимо для синтеза белка? 20 аминокислот м РНК Рибосома АТФ, ГТФ Белковые факторы регуляции
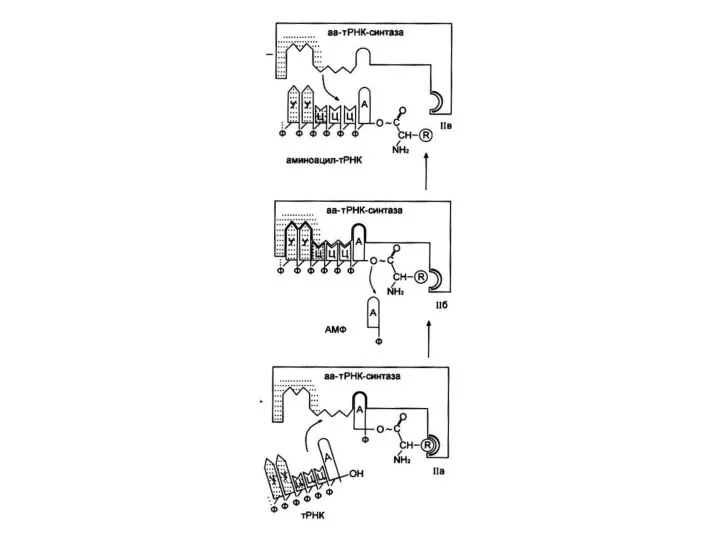
- 82. Узнавание и активация аминокислот в цитоплазме Специфическая для каждой аминокислоты аминоацил-тРНК-синтетаза катализирует реакцию в два этапа:
- 83. Реакция активации аминокислот Аминокислота +АТФ +т РНК ? ? т РНК + АМФ + ФФ. 2
- 86. Инициация трансляции Малая субъединица (40S) + т РНК-мет + ГТФ + eIF -2 (эукариотический инициирующий фактор).
- 89. Элонгация трансляции Поступающие, нагруженные аминокислотами т РНК связываются с кодонами м РНК в аминоацильном центре. Пептидилтрансфераза
- 92. Терминация трансляции В аминоацильном центре оказывается нонсенс – кодон (UAG, UAA, UGA) для которого нет соответствующей
- 93. Созревание белковых молекул Посттрансляционный процессинг осуществляется ферментами ЭПС: Лимитированный протеолиз Ковалентная модификация аминокислот Образование S –
- 94. Ингибиторы трансляции Стрептомицин – препятствует связыванию формилметионин- т РНК с рибосомой, нарушая инициацию трансляции. Связывается с
- 95. Действие токсинов Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой II, блокируя синтез м
- 98. Скачать презентацию

ИСТОРИЯ открытия нуклеиновых кислот
Мишер (1869г.) выделение ДНК из ядерного материала тимуса,
ИСТОРИЯ открытия нуклеиновых кислот
Мишер (1869г.) выделение ДНК из ядерного материала тимуса,

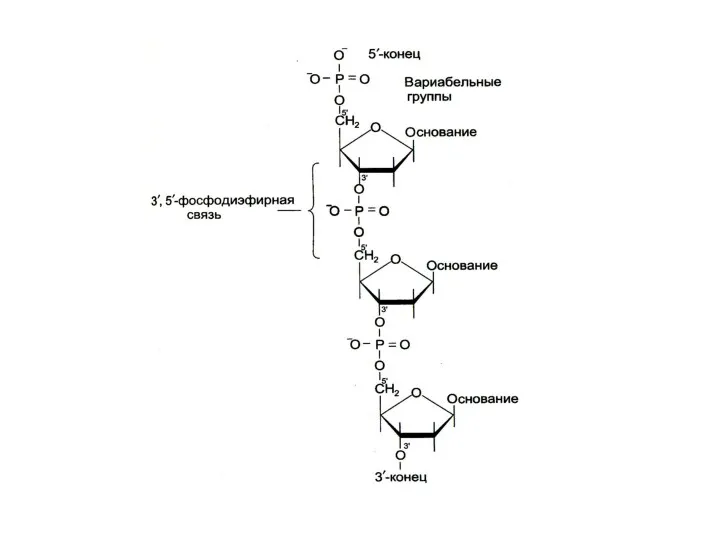
Строение нуклеиновых кислот
Нуклеиновые кислоты – линейные полимеры, состоящие из нуклеотидов, соединенных
Строение нуклеиновых кислот
Нуклеиновые кислоты – линейные полимеры, состоящие из нуклеотидов, соединенных


Пурины и пиримидины
Азотистые основания – гетероциклические, плоские структуры, существуют в кето
Пурины и пиримидины
Азотистые основания – гетероциклические, плоские структуры, существуют в кето

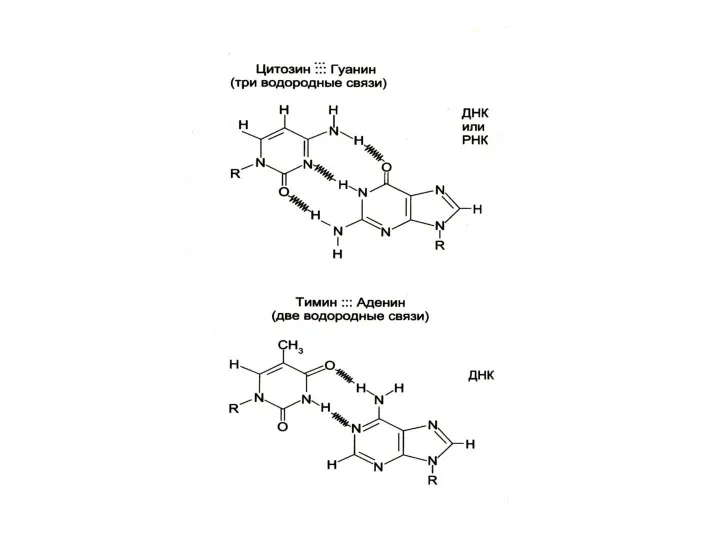
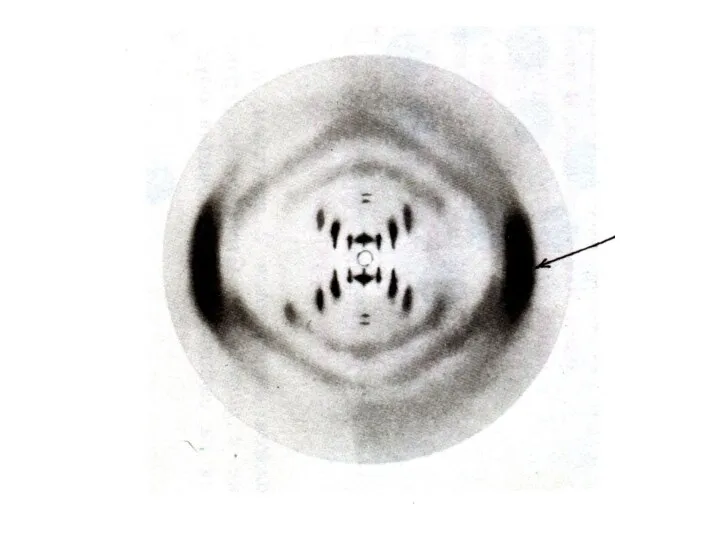
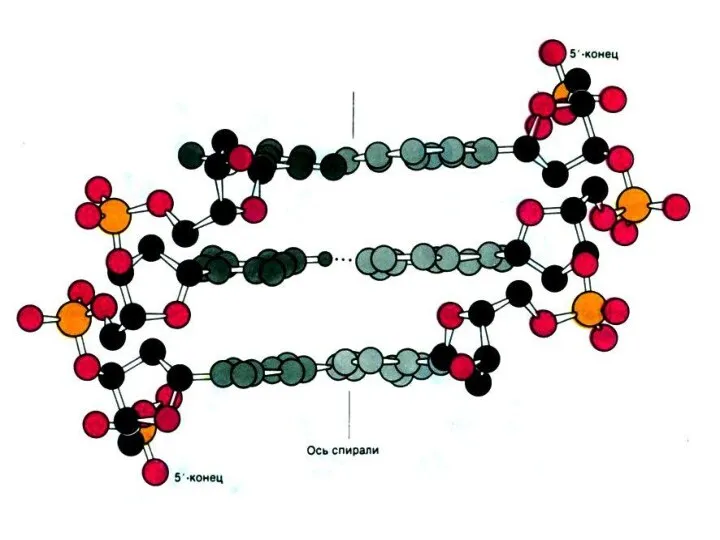
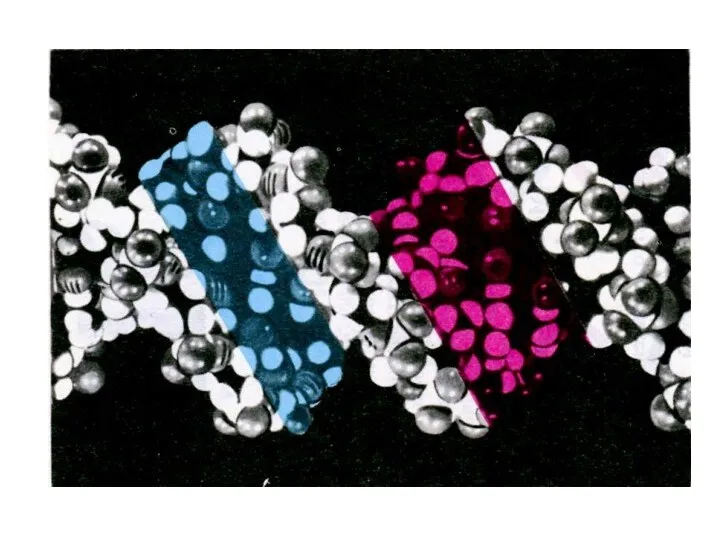
Пространственная структура нуклеиновых кислот
Первичная структура – последовательность нуклеотидов
Вторичная структура – двойная
Пространственная структура нуклеиновых кислот
Первичная структура – последовательность нуклеотидов
Вторичная структура – двойная
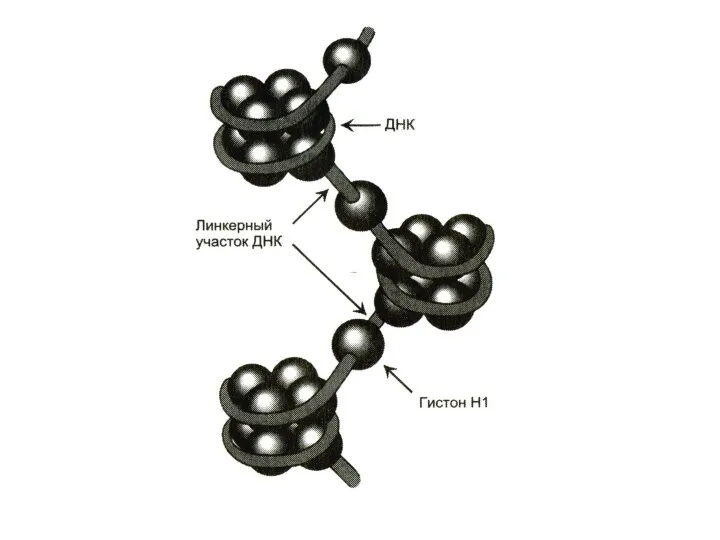

Внешний обмен нуклеиновых кислот
Нуклеопротеины пищи в кислом желудочном соке распадаются на
Внешний обмен нуклеиновых кислот
Нуклеопротеины пищи в кислом желудочном соке распадаются на

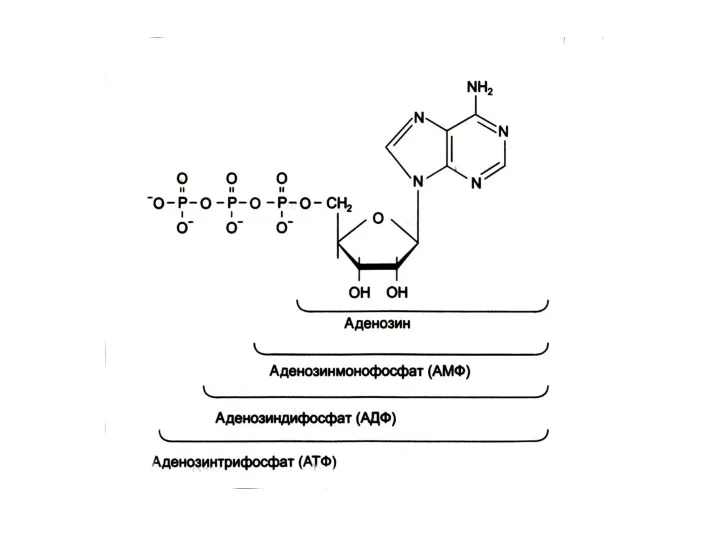
Метаболическая роль нуклеотидов
Мономеры для синтеза ДНК и РНК
Поддержание энергетического гомеостаза АДФ
Метаболическая роль нуклеотидов
Мономеры для синтеза ДНК и РНК
Поддержание энергетического гомеостаза АДФ

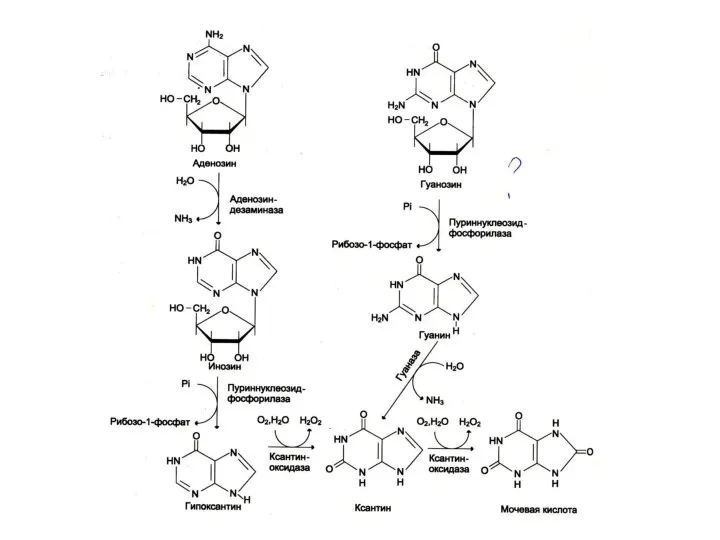
Катаболизм пуринов
АМФ ?аденозин ?инозин ? гипоксантин ? ксантин ? мочевая кислота
ГМФ
Катаболизм пуринов
АМФ ?аденозин ?инозин ? гипоксантин ? ксантин ? мочевая кислота
ГМФ

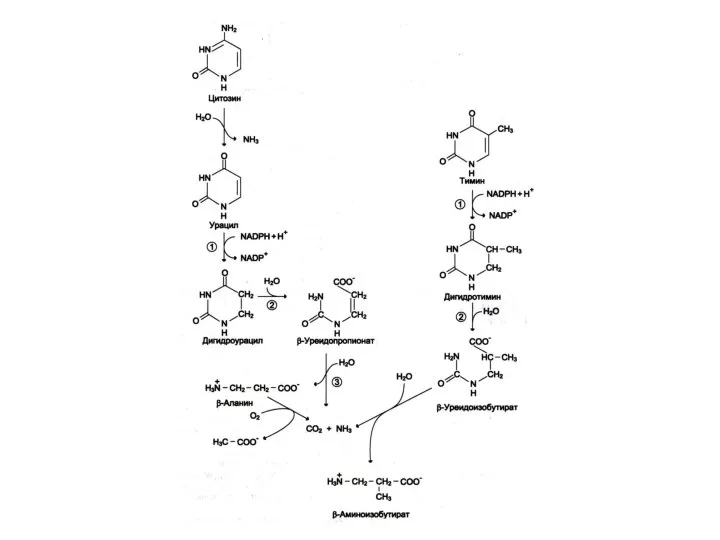
Катаболизм пиримидинов
ЦМФ ? УМФ ? урацил
ТМФ ? тимин
Восстановление и гидролиз пиримидинов
Катаболизм пиримидинов
ЦМФ ? УМФ ? урацил
ТМФ ? тимин
Восстановление и гидролиз пиримидинов

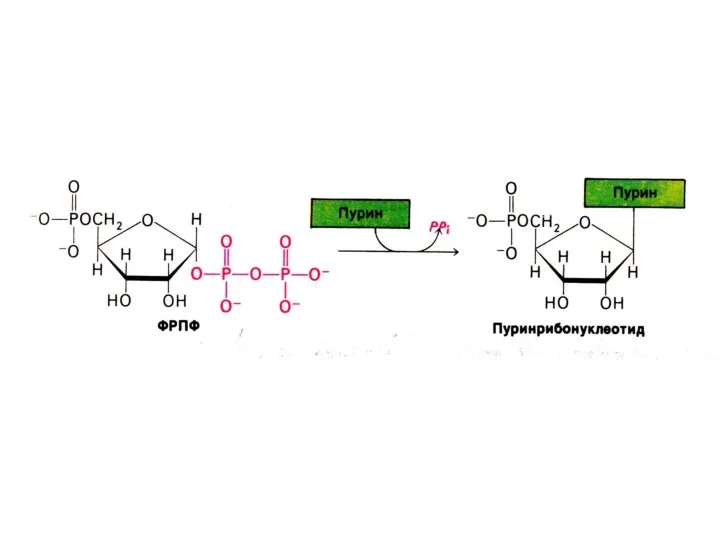
Синтез нуклеотидов
Синтез нуклеотидов лимитируется синтезом азотистых оснований de novo.
Бьюкенен с помощью
Синтез нуклеотидов
Синтез нуклеотидов лимитируется синтезом азотистых оснований de novo.
Бьюкенен с помощью
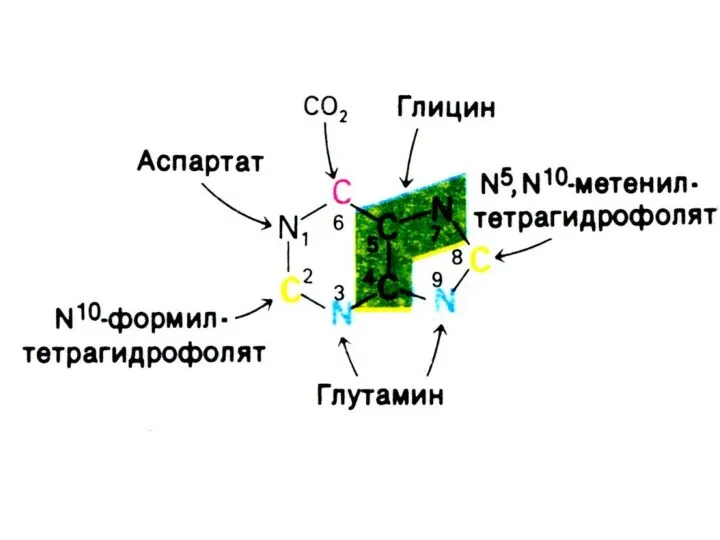

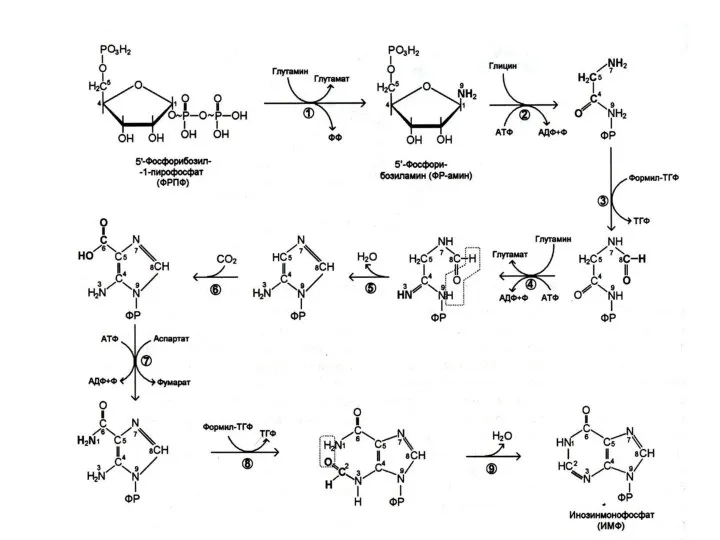
Биосинтез пуринов
На основе 5-фосфорибозил -1- пирофосфата строится имидазольное кольцо, затем пуриновое.
Общий
Биосинтез пуринов
На основе 5-фосфорибозил -1- пирофосфата строится имидазольное кольцо, затем пуриновое.
Общий
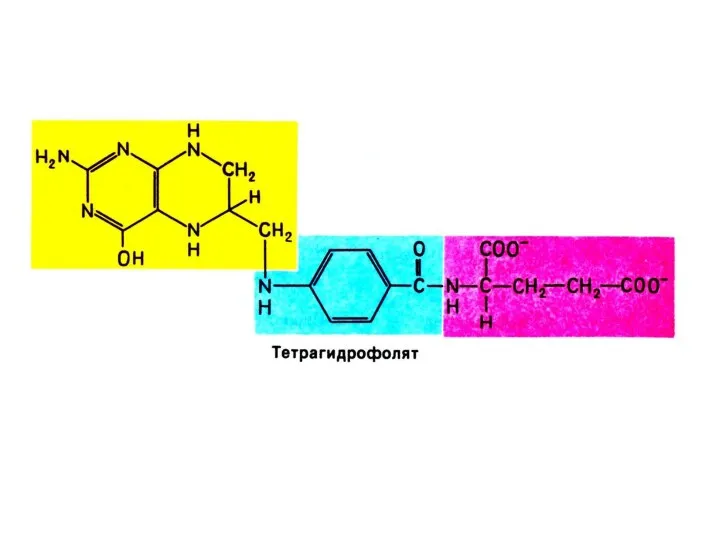
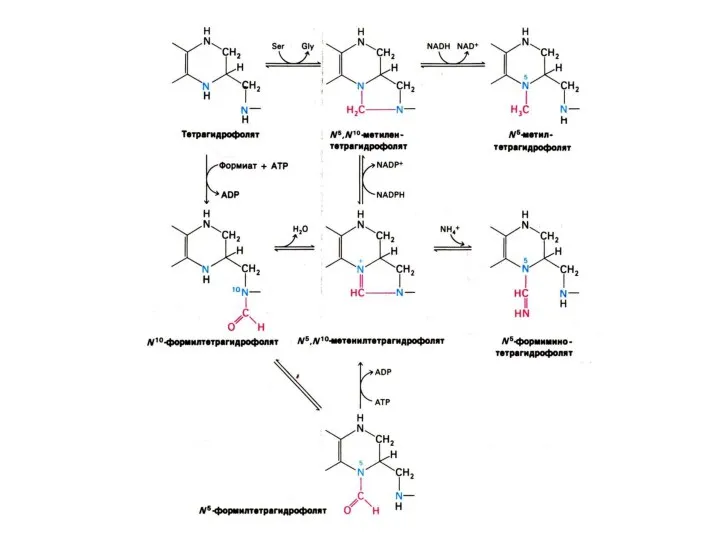

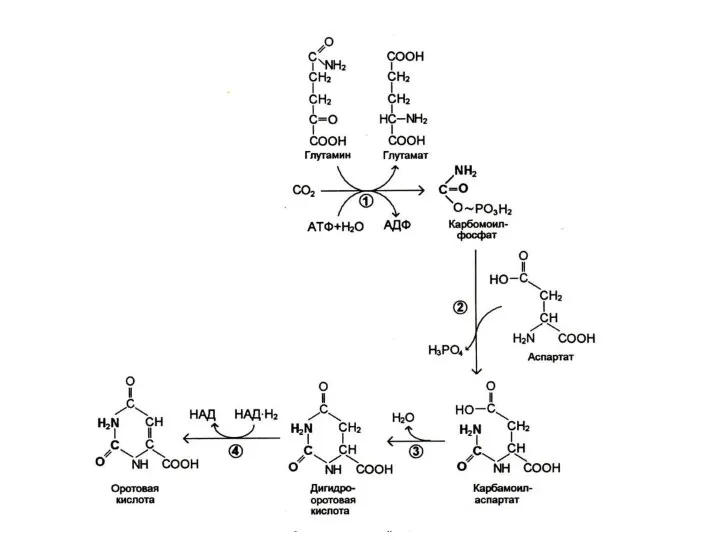
Биосинтез пиримидинов
Биосинтез пиримидинов начинается с построения азотистого гетероцикла с участием NH3,,СО2,глу,
Биосинтез пиримидинов
Биосинтез пиримидинов начинается с построения азотистого гетероцикла с участием NH3,,СО2,глу,
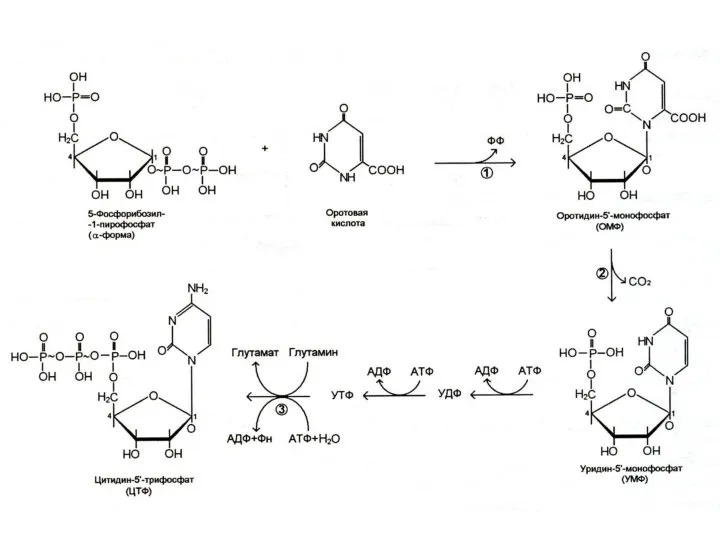
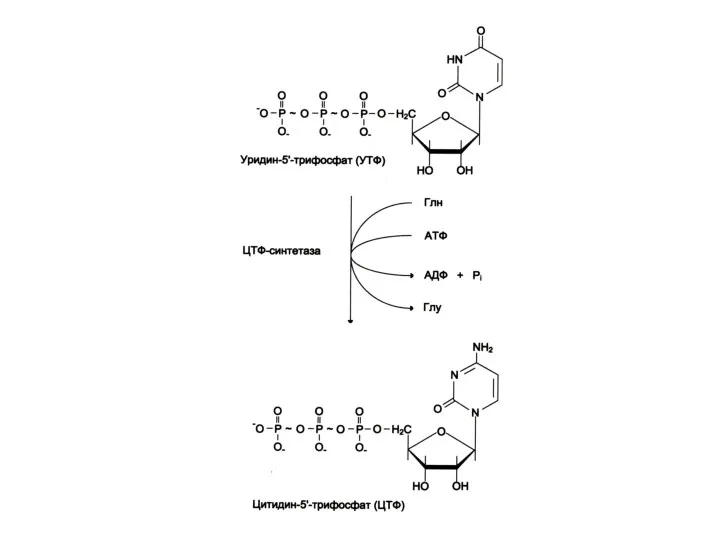

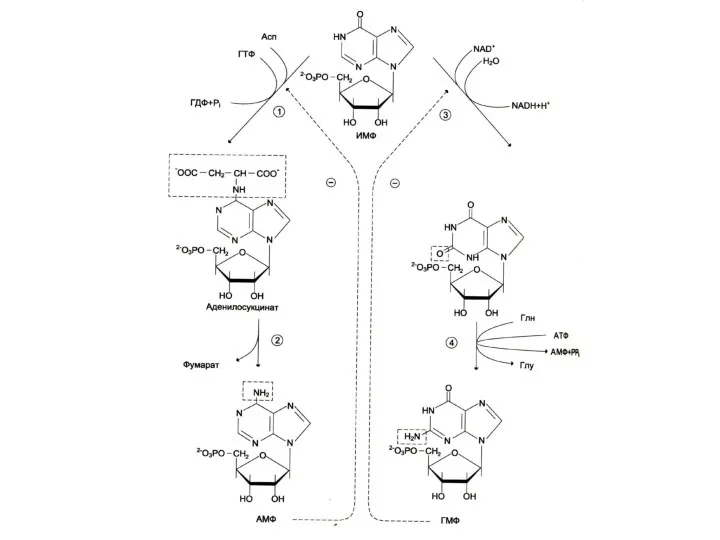
Образование нуклеозидтрифосфатов
АМФ + АТФ ? 2АДФ
ГМФ + АТФ ? ГДФ +
Образование нуклеозидтрифосфатов
АМФ + АТФ ? 2АДФ
ГМФ + АТФ ? ГДФ +

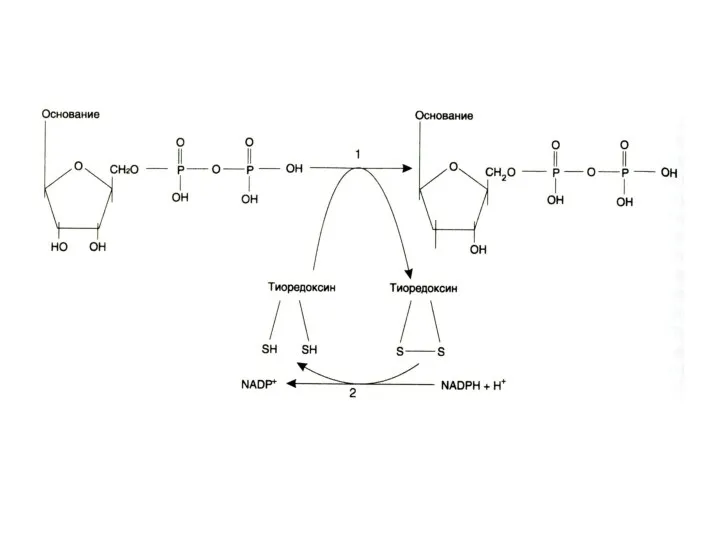
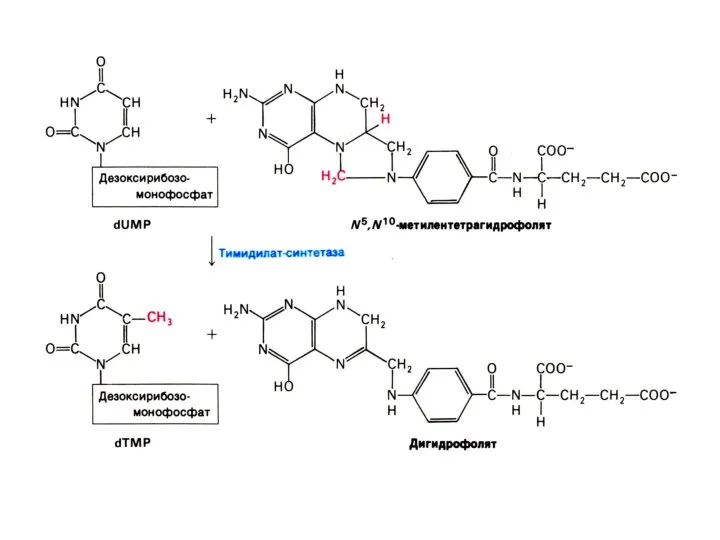
Синтез дезоксинуклеотидов
Все нуклеотиды образуются с участием фосфорибозилпирофосфата.
Дезоксирибонуклеотиды образуются при восстановлении рибозы
Синтез дезоксинуклеотидов
Все нуклеотиды образуются с участием фосфорибозилпирофосфата.
Дезоксирибонуклеотиды образуются при восстановлении рибозы

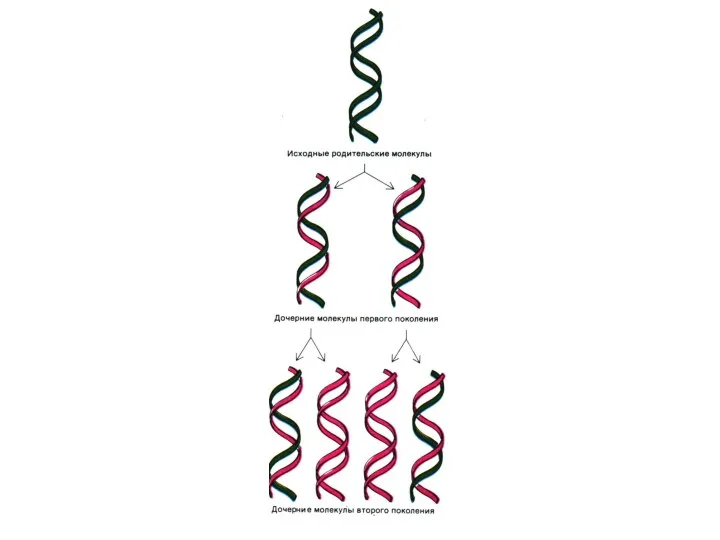
Репликация ДНК
Реакция матричного синтеза. Удвоение цепей ДНК, матрицей служит каждая из
Репликация ДНК
Реакция матричного синтеза. Удвоение цепей ДНК, матрицей служит каждая из

Репликация ДНК
Этапы: инициация, элонгация, терминация синтеза и созревание дочерней цепи (метилирование).
Репарация
Репликация ДНК
Этапы: инициация, элонгация, терминация синтеза и созревание дочерней цепи (метилирование).
Репарация

Репликация ДНК
Этап инициации:
Сигналом начала репликации служат белковые факторы роста (модифицирующие регуляторные
Репликация ДНК
Этап инициации:
Сигналом начала репликации служат белковые факторы роста (модифицирующие регуляторные

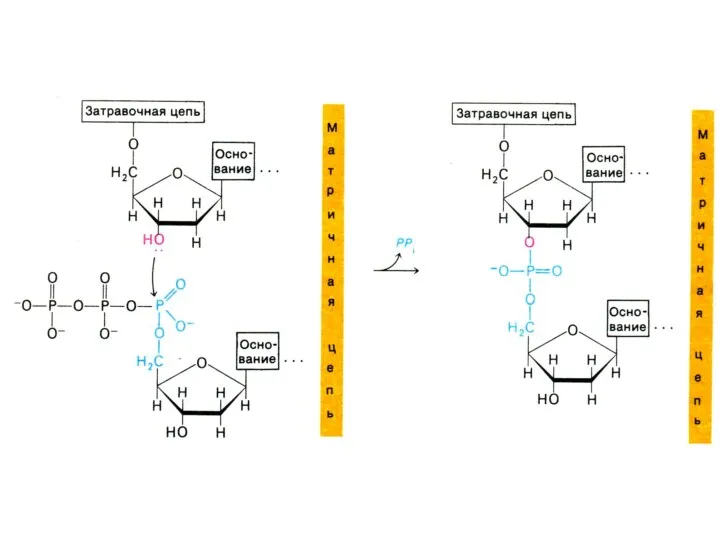
Репликация ДНК
Механизм реакции:
Субстратами служат дезоксинуклеозидтрифосфаты
3-ОН группа дезоксирибозы (рибозы) производит нуклеофильную атаку
Репликация ДНК
Механизм реакции:
Субстратами служат дезоксинуклеозидтрифосфаты
3-ОН группа дезоксирибозы (рибозы) производит нуклеофильную атаку

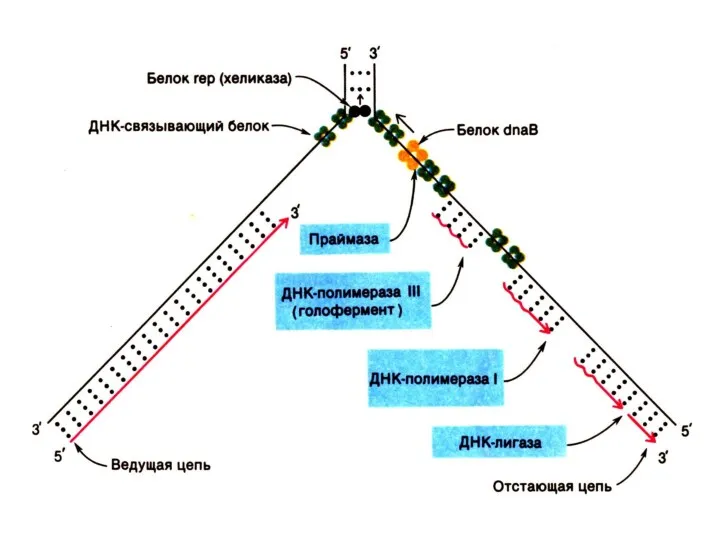
Репликация ДНК
Этап элонгации:
Направление синтеза 5 ?3
ДНК – полимераза α синтезирует «затравку»
Репликация ДНК
Этап элонгации:
Направление синтеза 5 ?3
ДНК – полимераза α синтезирует «затравку»

Репликация ДНК
Реплицируются одновременно обе одноцепочечные матрицы (5?3)
Одна (лидирующая) цепь реплицируется непрерывно,
Репликация ДНК
Реплицируются одновременно обе одноцепочечные матрицы (5?3)
Одна (лидирующая) цепь реплицируется непрерывно,
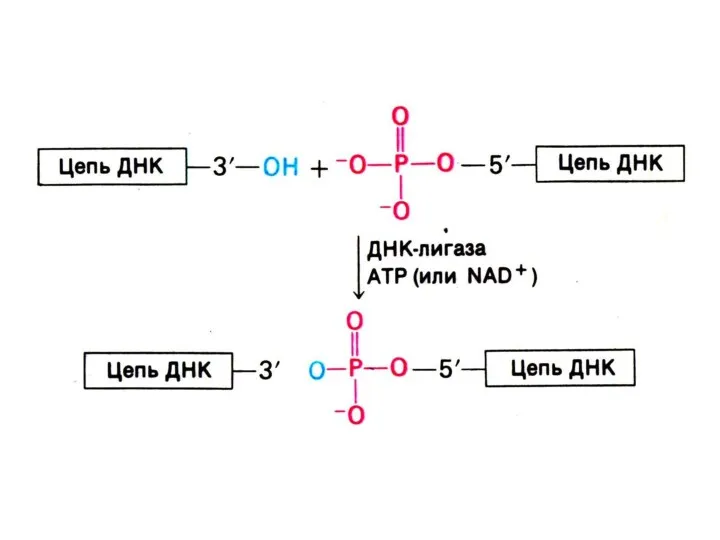

Репликация ДНК
Скорость репликации огромна, т.к. реакция идет в нескольких местах одновременно
Репликация ДНК
Скорость репликации огромна, т.к. реакция идет в нескольких местах одновременно
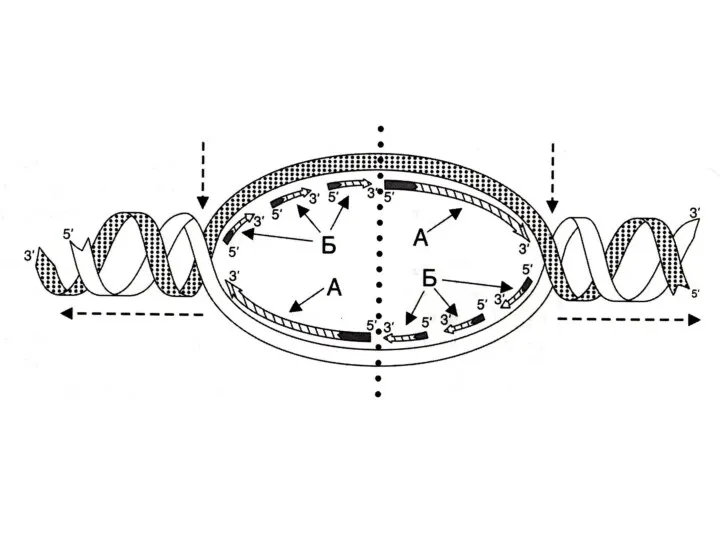
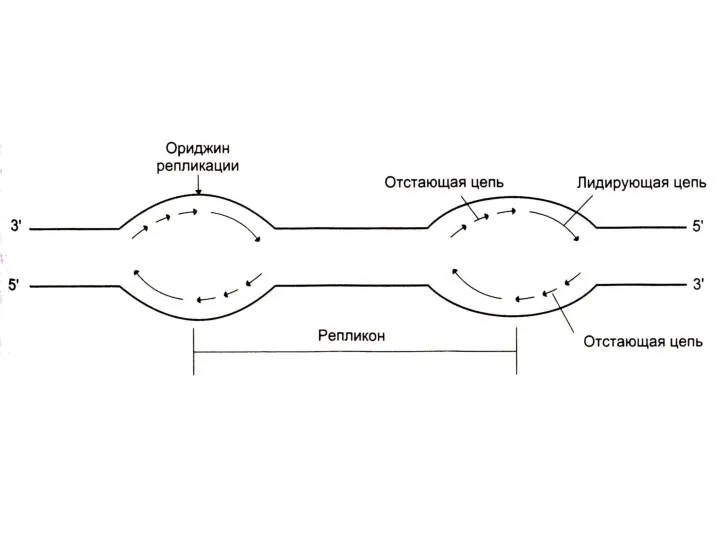

Репликация ДНК
ДНК- полимеразы Δ и ε делают 1 ошибку на 105
Репликация ДНК
ДНК- полимеразы Δ и ε делают 1 ошибку на 105
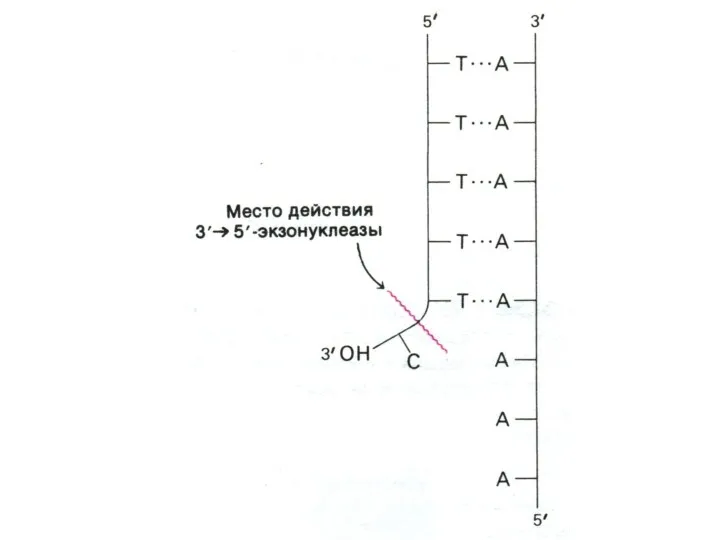
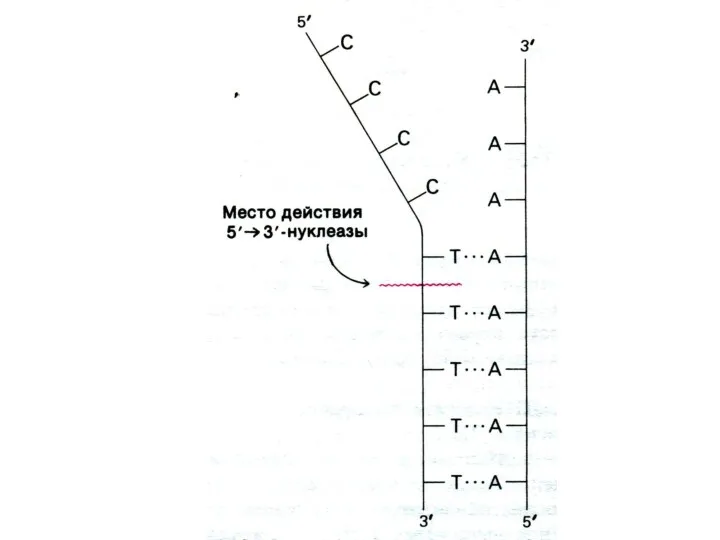

Репликация ДНК
Ошибки в ДНК (мутации) возникают спонтанно (ошибки репликации, дезаминирование нуклеотидов,
Репликация ДНК
Ошибки в ДНК (мутации) возникают спонтанно (ошибки репликации, дезаминирование нуклеотидов,

Репликация ДНК
Комплекс ферментов репарации узнает и вырезает поврежденные и химически измененные
Репликация ДНК
Комплекс ферментов репарации узнает и вырезает поврежденные и химически измененные
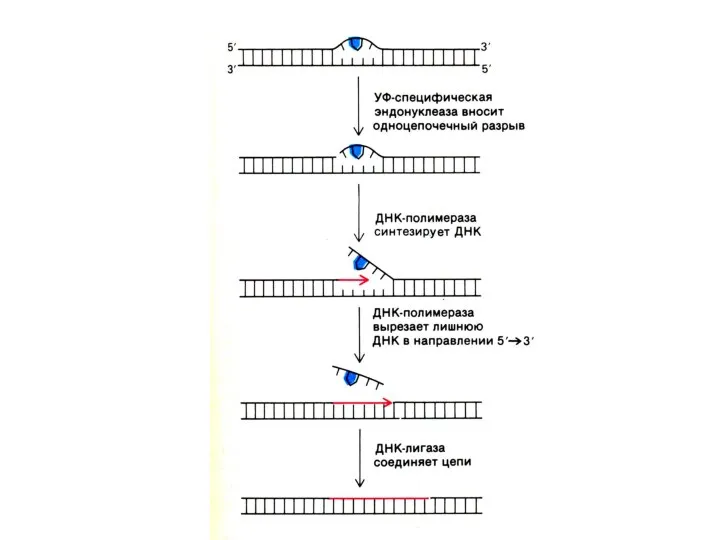

Репликация ДНК
Количество раундов репликации ДНК (а значит число возможных делений клетки)
Репликация ДНК
Количество раундов репликации ДНК (а значит число возможных делений клетки)

Репликация ДНК
Созревание молекулы ДНК:
Через несколько минут после завершения репликации происходит метилирование
Репликация ДНК
Созревание молекулы ДНК:
Через несколько минут после завершения репликации происходит метилирование
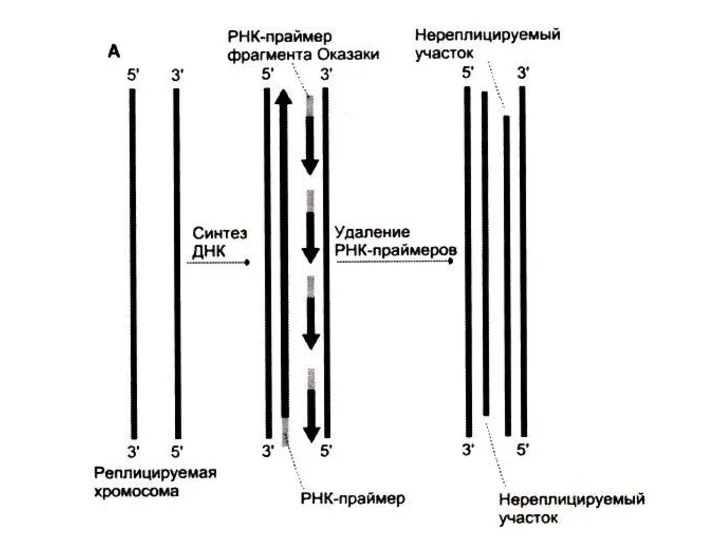
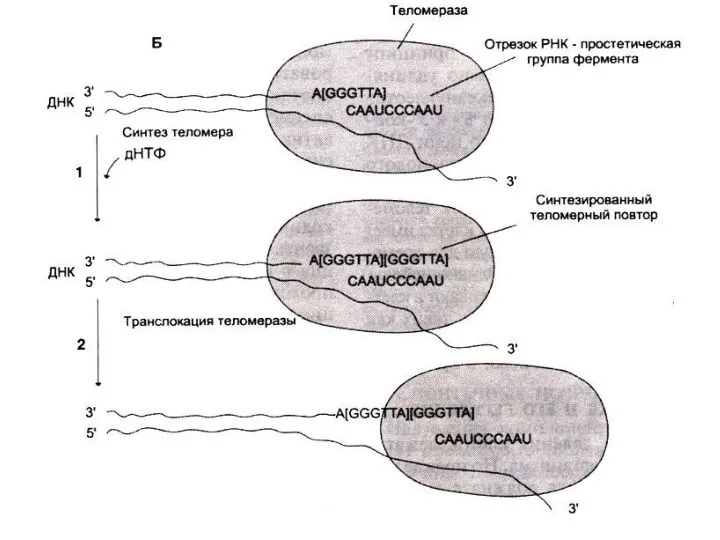

Ингибиторы репликации
Антибиотики (дауномицин, доксорубицин, рифампицин, актиномицин Д) способны встраиваться (интеркаляция)
Ингибиторы репликации
Антибиотики (дауномицин, доксорубицин, рифампицин, актиномицин Д) способны встраиваться (интеркаляция)

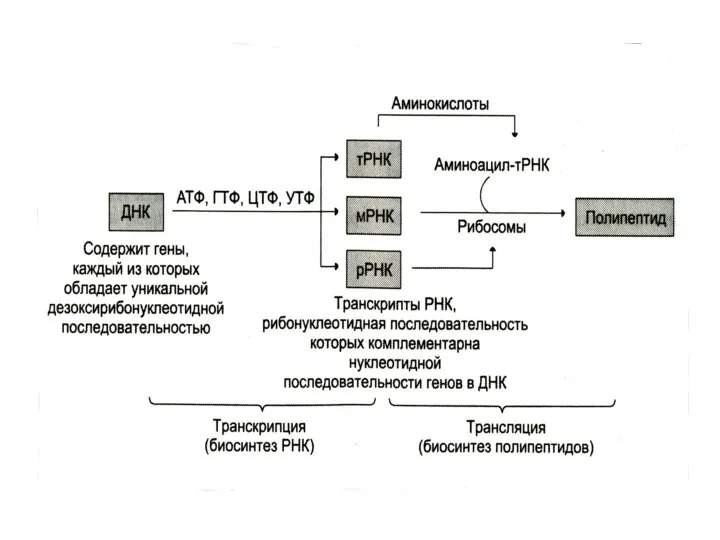
Транскрипция
Считывание информации с ДНК-матрицы на РНК, синтез тРНК, иРНК, рРНК с
Транскрипция
Считывание информации с ДНК-матрицы на РНК, синтез тРНК, иРНК, рРНК с

Транскрипция
Механизм РНК – полимеразной реакции тот же, что и ДНК –
Транскрипция
Механизм РНК – полимеразной реакции тот же, что и ДНК –

Транскрипция
В ДНК – матрице выделяют транскиптоны. Участки, ограниченные промоторами и сайтами
Транскрипция
В ДНК – матрице выделяют транскиптоны. Участки, ограниченные промоторами и сайтами

Транскрипция
3 стадии транскрипции: инициация, элонгация и терминация.
Инициация синтеза начинается с «узнавания»
Транскрипция
3 стадии транскрипции: инициация, элонгация и терминация.
Инициация синтеза начинается с «узнавания»

Инициация транскрипции
Для формирование транскрипционной вилки (раскручивание одного витка спирали ДНК-матрицы) к
Инициация транскрипции
Для формирование транскрипционной вилки (раскручивание одного витка спирали ДНК-матрицы) к
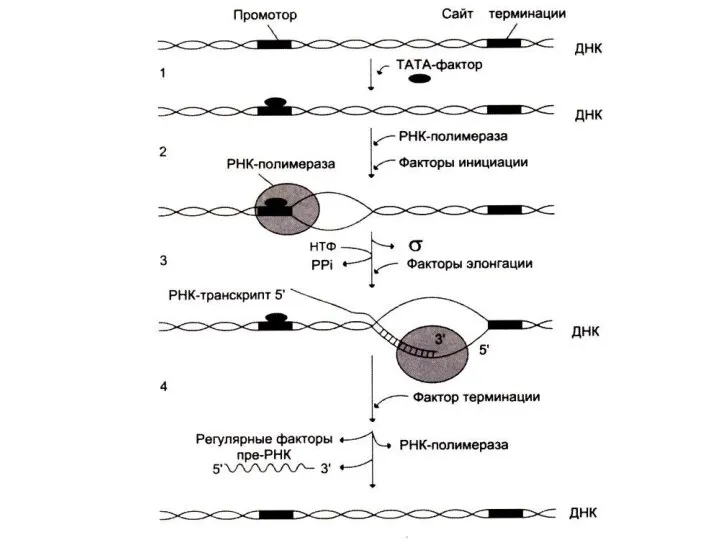

Элонгация транскрипции
Белковые факторы элонгации обеспечивают расплетение ДНК перед продвижением РНК-полимеразы и
Элонгация транскрипции
Белковые факторы элонгации обеспечивают расплетение ДНК перед продвижением РНК-полимеразы и

Терминация транскрипции
При достижении РНК - полимеразой сайта терминации белковый фактор терминации
Терминация транскрипции
При достижении РНК - полимеразой сайта терминации белковый фактор терминации

Созревание РНК-транскриптов
Процессингу (созреванию) подвергаются все виды РНК (и, т, р).
А) Ковалентная
Созревание РНК-транскриптов
Процессингу (созреванию) подвергаются все виды РНК (и, т, р).
А) Ковалентная

Ковалентная модификация иРНК
Гуанилил-трансфераза присоединяет ГДФ к 5- ОР концу (5-О-Р-О-5 связь),
5
Ковалентная модификация иРНК
Гуанилил-трансфераза присоединяет ГДФ к 5- ОР концу (5-О-Р-О-5 связь),
5

СПЛАЙСИНГ иРНК
Сплайсинг: образование зрелой мРНК:
Вырезание интронных последовательностей (ограниченных AGGU- и -
СПЛАЙСИНГ иРНК
Сплайсинг: образование зрелой мРНК:
Вырезание интронных последовательностей (ограниченных AGGU- и -

Процессинг первичных транскриптов тРНК
РНК - аза отщепляет нуклеотиды с 3 –
Процессинг первичных транскриптов тРНК
РНК - аза отщепляет нуклеотиды с 3 –


Созревание рибосомальных РНК
Образуется множество первичных транскриптов 5 S и 45 S.
45
Созревание рибосомальных РНК
Образуется множество первичных транскриптов 5 S и 45 S.
45

Ингибиторы транскрипции
Рифампицин связывается с β - субъединицей РНК –полимеразы, ингибируя образование
Ингибиторы транскрипции
Рифампицин связывается с β - субъединицей РНК –полимеразы, ингибируя образование

Трансляция
Перевод генетической информации с кодонов мРНК на аминокислотную последовательность белка (экспрессия
Трансляция
Перевод генетической информации с кодонов мРНК на аминокислотную последовательность белка (экспрессия

Трансляция
Что необходимо для синтеза белка?
20 аминокислот
м РНК
Рибосома
АТФ, ГТФ
Белковые факторы регуляции инициации,
Трансляция
Что необходимо для синтеза белка?
20 аминокислот
м РНК
Рибосома
АТФ, ГТФ
Белковые факторы регуляции инициации,

Узнавание и активация аминокислот в цитоплазме
Специфическая для каждой аминокислоты аминоацил-тРНК-синтетаза катализирует
Узнавание и активация аминокислот в цитоплазме
Специфическая для каждой аминокислоты аминоацил-тРНК-синтетаза катализирует

Реакция активации аминокислот
Аминокислота +АТФ +т РНК ?
? т РНК + АМФ
Реакция активации аминокислот
Аминокислота +АТФ +т РНК ?
? т РНК + АМФ


Инициация трансляции
Малая субъединица (40S) + т РНК-мет + ГТФ + eIF
Инициация трансляции
Малая субъединица (40S) + т РНК-мет + ГТФ + eIF

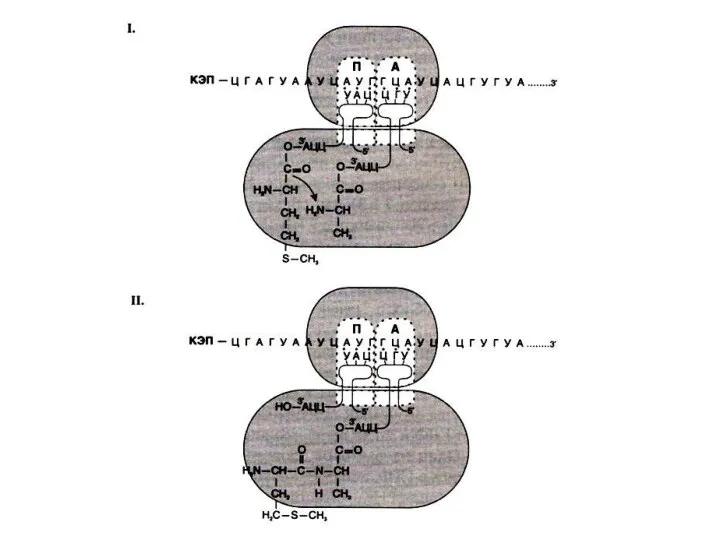

Элонгация трансляции
Поступающие, нагруженные аминокислотами т РНК связываются с кодонами м РНК
Элонгация трансляции
Поступающие, нагруженные аминокислотами т РНК связываются с кодонами м РНК
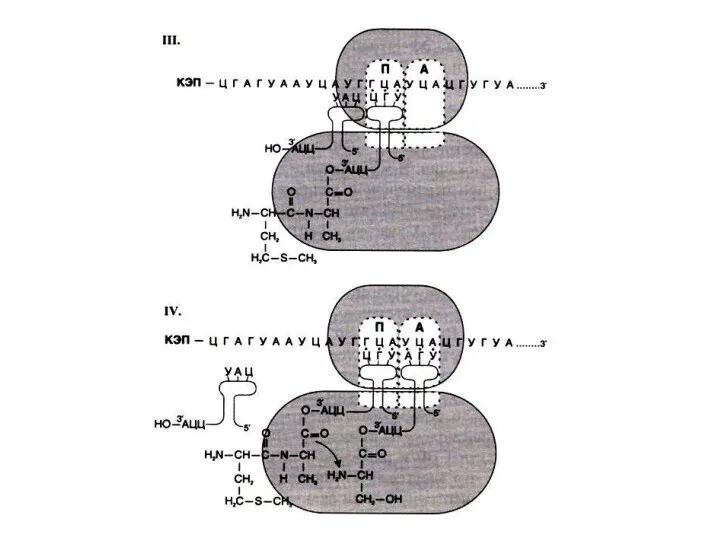


Терминация трансляции
В аминоацильном центре оказывается нонсенс – кодон (UAG, UAA, UGA)
Терминация трансляции
В аминоацильном центре оказывается нонсенс – кодон (UAG, UAA, UGA)

Созревание белковых молекул
Посттрансляционный процессинг осуществляется ферментами ЭПС:
Лимитированный протеолиз
Ковалентная модификация аминокислот
Образование
Созревание белковых молекул
Посттрансляционный процессинг осуществляется ферментами ЭПС:
Лимитированный протеолиз
Ковалентная модификация аминокислот
Образование

Ингибиторы трансляции
Стрептомицин – препятствует связыванию формилметионин- т РНК с рибосомой, нарушая
Ингибиторы трансляции
Стрептомицин – препятствует связыванию формилметионин- т РНК с рибосомой, нарушая

Действие токсинов
Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой
Действие токсинов
Аманитин (токсин бледной поганки), циклический пептид, связывается с эукариотической РНК-полимеразой

 Класс земноводные
Класс земноводные Презентация по теме Анализаторы
Презентация по теме Анализаторы Квітка як орган статевого розмноження рослин
Квітка як орган статевого розмноження рослин Электронный атлас по паразитологии. Раздел 2 Тип Ленточные черви. Часть 1 Сосальщики
Электронный атлас по паразитологии. Раздел 2 Тип Ленточные черви. Часть 1 Сосальщики Нервная система
Нервная система Лабораторное исследование. Зубы млекопитающих
Лабораторное исследование. Зубы млекопитающих Учение об анализаторах. Орган зрения
Учение об анализаторах. Орган зрения сон и сновидения
сон и сновидения Класс Паукообразные
Класс Паукообразные Селекция. Селекция туралы жалпы түсінік
Селекция. Селекция туралы жалпы түсінік Биологическая продукция, скорость накопления биомассы в экосистеме
Биологическая продукция, скорость накопления биомассы в экосистеме Вегетативные органы. Корень
Вегетативные органы. Корень Цитологические основы наследования признаков при моногибридном скрещивании
Цитологические основы наследования признаков при моногибридном скрещивании Сущность жизни и свойства живого
Сущность жизни и свойства живого Цитотехнологія. Клітинна інженерія
Цитотехнологія. Клітинна інженерія Презентация к уроку биологии 8 кл по теме Кожа
Презентация к уроку биологии 8 кл по теме Кожа Морские свинки
Морские свинки Закономерности наследования на клеточном уровне. Генетика пола. Хромосомная теория наследственности
Закономерности наследования на клеточном уровне. Генетика пола. Хромосомная теория наследственности Отличия прокариотической клетки от эукариотической
Отличия прокариотической клетки от эукариотической О чем может рассказать сонатная форма
О чем может рассказать сонатная форма Су-тұз алмасу бұзылыстарының клиникалық биохимиясы
Су-тұз алмасу бұзылыстарының клиникалық биохимиясы Презентация к уроку биологии 7 класс
Презентация к уроку биологии 7 класс Животные луга
Животные луга Ядовитые грибы Нижегородской области
Ядовитые грибы Нижегородской области Строение клетки. Кроссворд
Строение клетки. Кроссворд Онтогенез. 10 класс
Онтогенез. 10 класс Як тварини спілкуються
Як тварини спілкуються Закономірності впливу екологічних чинників на організми та їхні угруповання
Закономірності впливу екологічних чинників на організми та їхні угруповання