- Обмен простых белков. Пути обмена аминокислот. (Тема 5)

Содержание
- 2. Общие пути превращения аминокислот включают реакции дезаминирования, трансаминирования, декарбоксилирования, биосинтеза и рацемизации. Первые четыре реакции имеют
- 3. Дезаминирование аминокислот Существует 4 типа дезаминирования аминокислот (отщепление аминогруппы). Во всех случаях NH2-группа аминокислоты освобождается в
- 5. Продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты. Для животных тканей, растений и большинства аэробных микроорганизмов
- 6. Окислительное дезаминирование аминокислот, протекает в две стадии Первая стадия является ферментативной и завершается образованием неустойчивого промежуточного
- 7. Оксидазы аминокислот (L- и D-изомеров) являются сложными флавопротеинами, содержащими в качестве кофермента ФМН или ФАД, которые
- 8. Схема реакции окислительного дезаминирования аминокислот с участием коферментов:
- 9. Восстановленные флавиннуклеотиды оксидаз L- и D-аминокислот могут непосредственно окисляться молекулярным кислородом. При этом образуется перекись водорода,
- 10. В животных тканях Г. Эйлером открыт высокоактивный специфический фермент (глутаматдегидрогеназа), катализирующий окислительное дезаминирование L-глутаминовой кислоты. Он
- 12. Первая стадия окисления глутаминовой кислоты аналогична реакции окислительного дезаминирования. Восстановленный НАДН далее окисляется при участии флавиновых
- 13. Различают три разных типа глутаматдегидрогеназ: один из них использует в качестве кофермента как НАД, так и
- 14. Трансаминирование аминокислот Трансаминирование - реакции межмолекулярного переноса аминогруппы (-NH2) от аминокислоты на α-кетокислоту без промежуточного образования
- 16. Реакции трансаминирования являются обратимыми и универсальными для всех живых организмов. Эти реакции протекают при участии специфических
- 17. Специфичность трансаминаз обеспечивается белковым компонентом. Ферменты трансаминирования катализируют перенос NH2-группы не на α-кетокислоту, а сначала на
- 18. Опуская промежуточные стадии образования шиффовых оснований, обе стадии реакции трансаминирования можно представить в виде общей схемы:
- 19. Клиническое значение определения активности трансаминаз Широкое распространение и активность трансаминаз в органах и тканях человека, а
- 20. Аспартат + α-Кетоглутарат = = Оксалоацетат + Глутамат Аланин + α-Кетоглутарат = = Пируват + Глутамат
- 21. Декарбоксилирование аминокислот Это процесс отщепления карбоксильной группы аминокислот в виде СО2 . Несмотря на ограниченный круг
- 22. В животных тканях установлено декарбоксилирование следующих аминокислот и их производных: тирозина, триптофана, 5-окситриптофана, валина, серина, гистидина,
- 23. В живых организмах открыты 4 типа декарбоксилирования аминокислот: 1. α-Декарбоксилирование, характерное для тканей животных, при котором
- 24. 2. ω-Декарбоксилирование, свойственное микроорганизмам. Например, из аспарагиновой кислоты этим путем образуется α-аланин:
- 25. 3. Декарбоксилирование, связанное с реакцией трансаминирования: В этой реакции образуются альдегид и новая аминокислота, соответствующая исходной
- 26. 4. Декарбоксилирование, связанное с реакцией конденсации двух молекул: Эта реакция в тканях животных осуществляется при синтезе
- 28. Скачать презентацию

Общие пути превращения аминокислот включают реакции дезаминирования, трансаминирования, декарбоксилирования, биосинтеза
Общие пути превращения аминокислот включают реакции дезаминирования, трансаминирования, декарбоксилирования, биосинтеза

Дезаминирование аминокислот
Существует 4 типа дезаминирования аминокислот (отщепление аминогруппы).
Во всех
Дезаминирование аминокислот
Существует 4 типа дезаминирования аминокислот (отщепление аминогруппы).
Во всех
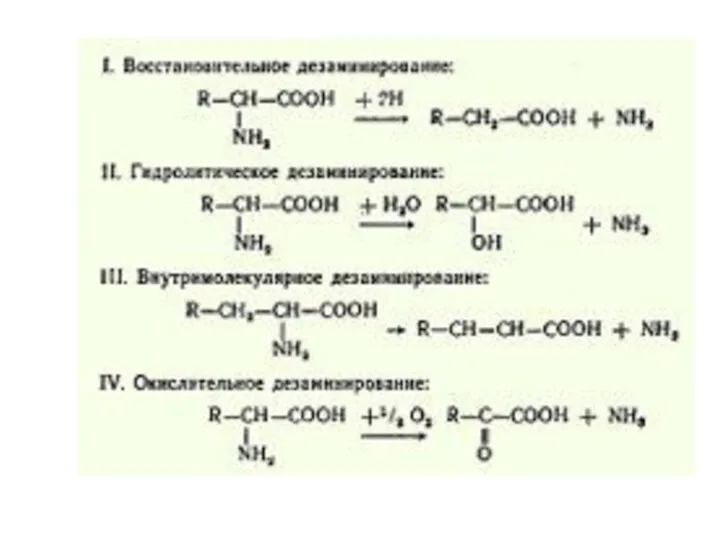

Продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты.
Для
Продуктами дезаминирования являются жирные кислоты, оксикислоты и кетокислоты.
Для
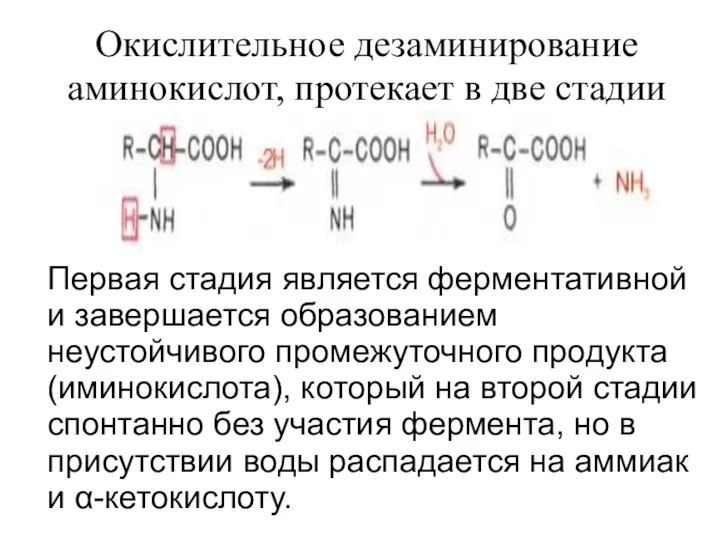
Окислительное дезаминирование аминокислот, протекает в две стадии
Первая стадия является ферментативной и
Окислительное дезаминирование аминокислот, протекает в две стадии
Первая стадия является ферментативной и

Оксидазы аминокислот (L- и D-изомеров) являются сложными флавопротеинами, содержащими в
Оксидазы аминокислот (L- и D-изомеров) являются сложными флавопротеинами, содержащими в
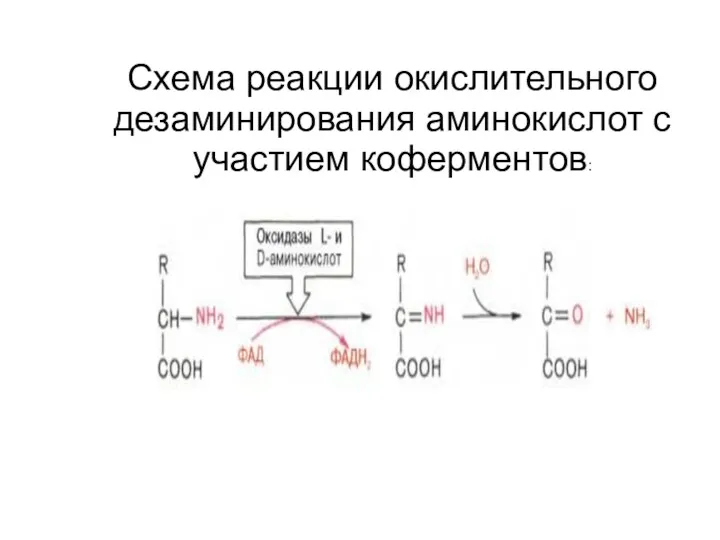
Схема реакции окислительного дезаминирования аминокислот с участием коферментов:
Схема реакции окислительного дезаминирования аминокислот с участием коферментов:

Восстановленные флавиннуклеотиды оксидаз L- и D-аминокислот могут непосредственно окисляться молекулярным
Восстановленные флавиннуклеотиды оксидаз L- и D-аминокислот могут непосредственно окисляться молекулярным

В животных тканях Г. Эйлером открыт высокоактивный специфический фермент (глутаматдегидрогеназа),
В животных тканях Г. Эйлером открыт высокоактивный специфический фермент (глутаматдегидрогеназа),


Первая стадия окисления глутаминовой кислоты аналогична реакции окислительного дезаминирования. Восстановленный
Первая стадия окисления глутаминовой кислоты аналогична реакции окислительного дезаминирования. Восстановленный

Различают три разных типа глутаматдегидрогеназ: один из них использует в
Различают три разных типа глутаматдегидрогеназ: один из них использует в

Трансаминирование аминокислот
Трансаминирование - реакции межмолекулярного переноса аминогруппы
(-NH2) от аминокислоты на
Трансаминирование аминокислот
Трансаминирование - реакции межмолекулярного переноса аминогруппы
(-NH2) от аминокислоты на


Реакции трансаминирования являются обратимыми и универсальными для всех живых организмов.
Реакции трансаминирования являются обратимыми и универсальными для всех живых организмов.

Специфичность трансаминаз обеспечивается белковым компонентом. Ферменты трансаминирования катализируют перенос NH2-группы
Специфичность трансаминаз обеспечивается белковым компонентом. Ферменты трансаминирования катализируют перенос NH2-группы

Опуская промежуточные стадии образования шиффовых оснований, обе стадии реакции трансаминирования можно
Опуская промежуточные стадии образования шиффовых оснований, обе стадии реакции трансаминирования можно

Клиническое значение определения активности трансаминаз
Широкое распространение и активность трансаминаз в
Клиническое значение определения активности трансаминаз
Широкое распространение и активность трансаминаз в

Аспартат + α-Кетоглутарат =
= Оксалоацетат + Глутамат
Аланин + α-Кетоглутарат
Аспартат + α-Кетоглутарат =
= Оксалоацетат + Глутамат
Аланин + α-Кетоглутарат

Декарбоксилирование аминокислот
Это процесс отщепления карбоксильной группы аминокислот в виде СО2 .
Декарбоксилирование аминокислот
Это процесс отщепления карбоксильной группы аминокислот в виде СО2 .

В животных тканях установлено декарбоксилирование следующих аминокислот и их производных:
В животных тканях установлено декарбоксилирование следующих аминокислот и их производных:

В живых организмах открыты 4 типа декарбоксилирования аминокислот:
1. α-Декарбоксилирование, характерное для
В живых организмах открыты 4 типа декарбоксилирования аминокислот:
1. α-Декарбоксилирование, характерное для

2. ω-Декарбоксилирование, свойственное микроорганизмам. Например, из аспарагиновой кислоты этим путем образуется
2. ω-Декарбоксилирование, свойственное микроорганизмам. Например, из аспарагиновой кислоты этим путем образуется
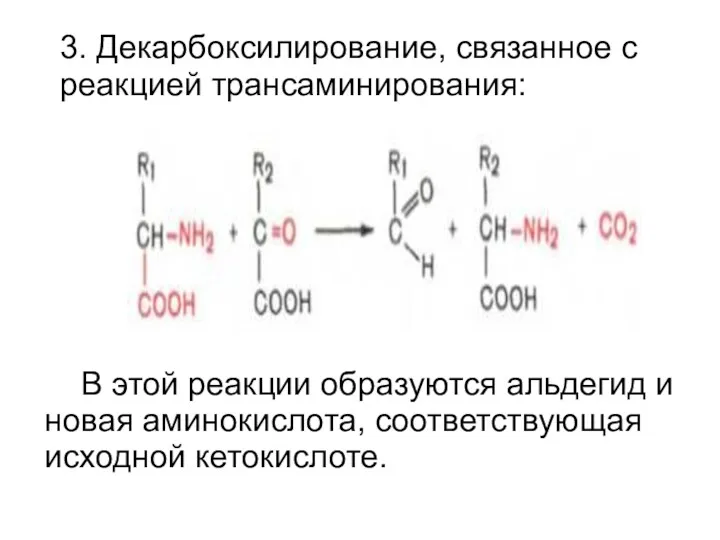
3. Декарбоксилирование, связанное с реакцией трансаминирования:
В этой реакции образуются альдегид
3. Декарбоксилирование, связанное с реакцией трансаминирования:
В этой реакции образуются альдегид
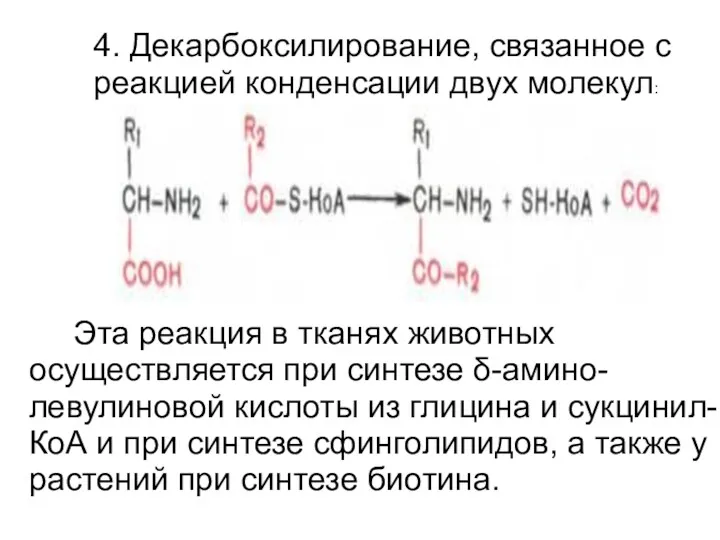
4. Декарбоксилирование, связанное с реакцией конденсации двух молекул:
Эта реакция в
4. Декарбоксилирование, связанное с реакцией конденсации двух молекул:
Эта реакция в
 Этические проблемы использования животных
Этические проблемы использования животных Пищеварение 1.1
Пищеварение 1.1 Общие признаки животных. 5 класс
Общие признаки животных. 5 класс Ретикулярная формация. Лимбическая система. Оболочки мозга
Ретикулярная формация. Лимбическая система. Оболочки мозга Хемосинтез. Хемосинтезирующие организмы
Хемосинтез. Хемосинтезирующие организмы Визначення типу шкіри на різних ділянках обличчя та складання правил догляду за власною шкірою
Визначення типу шкіри на різних ділянках обличчя та складання правил догляду за власною шкірою Прокариоты на службе по защите почвенной среды жизни от тяжёлых металлов
Прокариоты на службе по защите почвенной среды жизни от тяжёлых металлов Прісноводна гідра
Прісноводна гідра Zebra is a striped member of the horse family
Zebra is a striped member of the horse family Голоса птиц
Голоса птиц Імунітет рослин до інфекційних хвороб
Імунітет рослин до інфекційних хвороб Морфологическое описание растения
Морфологическое описание растения Презентация по биологии для учащихся 9 класса по теме Клеточная мембрана
Презентация по биологии для учащихся 9 класса по теме Клеточная мембрана Про кошек
Про кошек Презентация к уроку биологии в 7 классе Отряд Жесткокрылые, или Жуки
Презентация к уроку биологии в 7 классе Отряд Жесткокрылые, или Жуки Транскрипция и трансляция. Генетическая трансформация
Транскрипция и трансляция. Генетическая трансформация Китообразные и ластоногие
Китообразные и ластоногие Породы кошек
Породы кошек Классификация и характеристика рыб
Классификация и характеристика рыб Строение клеток прокариот и эукариот
Строение клеток прокариот и эукариот Органы и системы органов
Органы и системы органов Мед - пчелиная заслуга
Мед - пчелиная заслуга Микробиология - наука о микроорганизмах
Микробиология - наука о микроорганизмах Карл Линней - основоположник биологической систематики
Карл Линней - основоположник биологической систематики Движение крови в организме. Органы кровообращения
Движение крови в организме. Органы кровообращения Приглашение на семинар по управлению инкубаторием компании Пас Реформ
Приглашение на семинар по управлению инкубаторием компании Пас Реформ Витамины красоты
Витамины красоты Внеклассное мероприятие Красная книга Оренбургской области
Внеклассное мероприятие Красная книга Оренбургской области