- Обмін ліпідів

Содержание
- 3. План лекції 1.Травлення ліпідів у кишково-шлунковому тракті 2.Розпад ліпідів у клітинах організму. 3.Окиснення жирних кислот. 4.Утворення
- 5. Запас енергетичних ресурсів у організмі людини Energy is stored in the body in the form of
- 6. Жовчні кислоти
- 7. Uptake of dietary lipid in the intestine of a vertebrate animal, and delivery of fatty acids
- 8. Молекулярна структура хіломікронів Molecular structure of a chylomicron. The surface is covered with a layer of
- 9. Гідроліз триацилгліцеролів у шлунково- кишковому тракті
- 11. Розпад ацилгліцеролів у клітинах
- 12. Вивільнення енергії з триацилгліцеролу Mobilization of triacylglycerols stored in adipose tissue. Low levels of glucose in
- 13. Шляхи включення гліцерину у гліколіз та глюконеогенез
- 14. β-окиснення жирних кислот
- 15. Stages of fatty acid oxidation. Stage 1: A longchain fatty acid is oxidized to yield acetyl
- 16. β-окиснення жирних кислот
- 17. Активація жирних кислот Fatty acid activation by the formation of the fatty acyl-CoA derivative occurs in
- 18. Проникнення жирних кислот через мембрану мітохондрії
- 19. Проникнення жирних кислот через мембрану мітохондрії
- 20. Транспорт жирних кислот через внутрішню мембрану мітохондрій Ферментативні реакції перенесення довголанцюгових жирних кислот з цитозолю клітини
- 21. Карнітин
- 22. Проникнення жирних кислот через мембрану мітохондрії
- 23. Стадія дегідрування
- 24. Стадія гідратації
- 25. Друга стадія дегідрування
- 26. Тіолазна реакція
- 27. β-окислення The fatty acid oxidation (β-oxidation) pathway. (a) In each pass through this sequence, one acetyl
- 28. Енергетичний баланс β-окислення Якщо жирна кислота містить n атомів карбону, то за повного її окиснення утворюється
- 29. Енергетичний баланс β-окислення У випадку пальмітинової кислоти (С 16) відбувається 7 циклів β-окиснення: пальмітоїл-KоА + 7ФАД
- 32. Окислення ненасичених жирних кислот The oxidation of a monounsaturated fatty acyl-CoA, such as oleoyl-CoA (Δ9), requires
- 33. Окислення ненасичених жирних кислот Oxidation of polyunsaturated fatty acids requires a second auxiliary enzyme in addition
- 34. Кетонові тіла
- 35. Утворення кетонових тіл
- 36. Синтез жирних кислот
- 37. Синтез жирних кислот
- 38. Синтаза вищих жирних кислот
- 47. Сумарне рівняння синтезу жирних кислот Із пальмітинової кислоти синтезується стеаринова і інші вищі жирні кислоти шляхом
- 48. Синтез жирних кислот Утворений малоніл~KоА взаємодіє з АТБ-SH під впливом малонілтрансацилази. Ацетил-KоА переноситься на синтетичний комплекс,
- 51. Позамітохондріальна система біосинтезу жирних кислот Як вже зазначалося, будівельним блоком для синтезу жирних кислот в цитозолі
- 52. Утворення ненасичених жирних кислот
- 53. Біосинтез триацилгліцеролів
- 54. Біосинтез триацилгліцеролів
- 55. Біосинтез триацилгліцеролів
- 56. Біосинтез фосфогліцеридів
- 57. Біосинтез фосфогліцеридів
- 58. Біосинтез фосфогліцеридів
- 59. Біосинтез фосфогліцеридів
- 60. Біосинтез холестеролу
- 63. Ketone body formation and export from the liver. Conditions that increase gluconeogenesis (diabetes, fasting) slow the
- 64. The role of β oxidation in the conversion of seed triacylglycerols into glucose in germinating seeds.
- 65. Утворення кетонових тіл Formation of ketone bodies from acetyl-CoA. Under circumstances that cause acetylCoA accumulation (starvation
- 66. Electrons removed from fatty acids during β oxidation pass into the mitochondrial respiratory chain and eventually
- 68. Синтез жирних кислот
- 69. Acyl carrier protein (ACP). The prosthetic group is 4'-phosphopantetheine, which is covalently attached to the hydroxyl
- 71. Beginning of the second round of the fatty acid synthesis cycle. The butyryl group is on
- 72. The acetyl group shuttle for transfer of acetyl groups from mitochondria to the cytosol for fatty
- 73. Regulation of fatty acid synthesis. (a) In the cells of vertebrates, both allosteric regulation and hormone-dependent
- 74. Routes of synthesis of other fatty acids. Palmitate is the precursor of stearate and longer-chain saturated
- 77. Schematic depiction of PL activation in solution by CL and a duodenal mixed micelle. The formation
- 81. Скачать презентацию


План лекції
1.Травлення ліпідів у кишково-шлунковому тракті
2.Розпад ліпідів у клітинах організму.
3.Окиснення жирних
План лекції
1.Травлення ліпідів у кишково-шлунковому тракті
2.Розпад ліпідів у клітинах організму.
3.Окиснення жирних


Запас енергетичних ресурсів у організмі людини
Energy is stored in the body
Запас енергетичних ресурсів у організмі людини
Energy is stored in the body

Жовчні кислоти
Жовчні кислоти

Uptake of dietary lipid in the intestine of a vertebrate animal,
Uptake of dietary lipid in the intestine of a vertebrate animal,

Молекулярна структура хіломікронів
Molecular structure of a chylomicron. The surface is covered
Молекулярна структура хіломікронів
Molecular structure of a chylomicron. The surface is covered

Гідроліз триацилгліцеролів у шлунково- кишковому тракті
Гідроліз триацилгліцеролів у шлунково- кишковому тракті
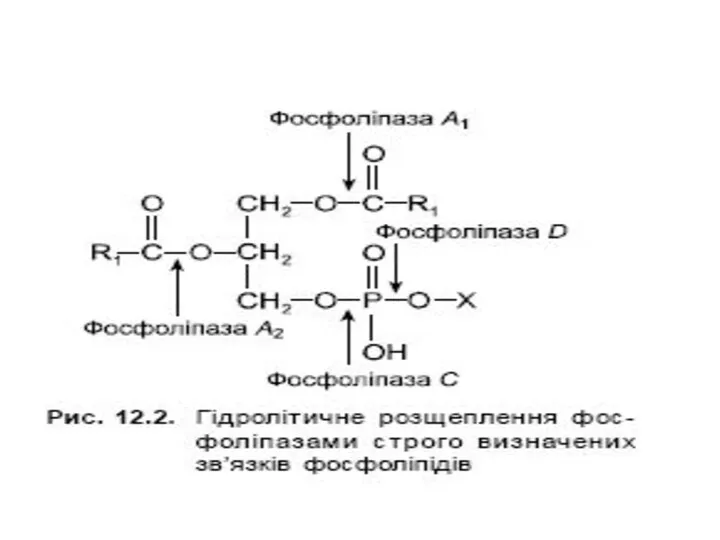
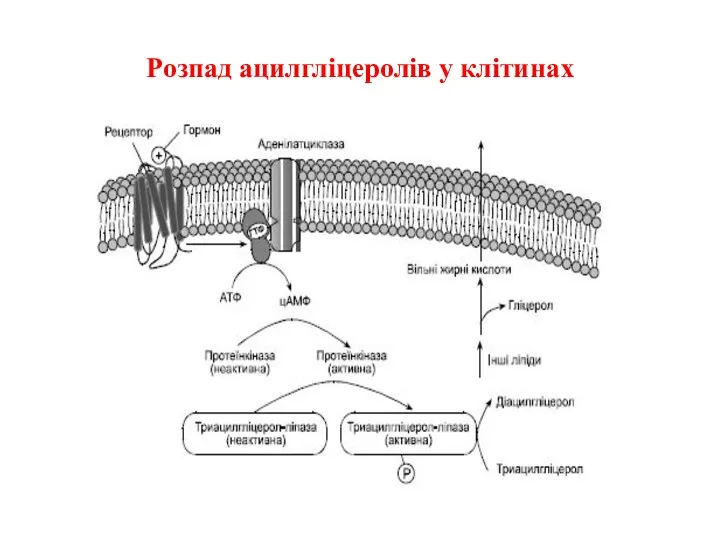
Розпад ацилгліцеролів у клітинах
Розпад ацилгліцеролів у клітинах
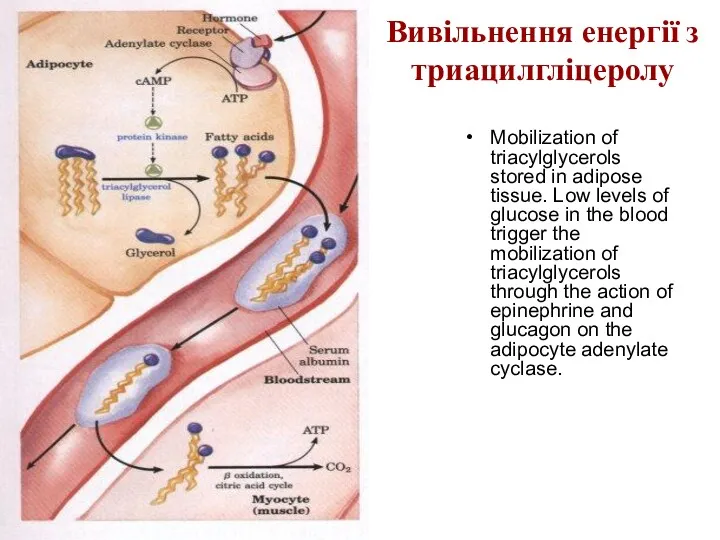
Вивільнення енергії з триацилгліцеролу
Mobilization of triacylglycerols stored in adipose tissue. Low
Вивільнення енергії з триацилгліцеролу
Mobilization of triacylglycerols stored in adipose tissue. Low

Шляхи включення гліцерину у гліколіз та глюконеогенез
Шляхи включення гліцерину у гліколіз та глюконеогенез

β-окиснення жирних кислот
β-окиснення жирних кислот

Stages of fatty acid oxidation.
Stage 1: A longchain fatty acid is
Stages of fatty acid oxidation.
Stage 1: A longchain fatty acid is

β-окиснення жирних кислот
β-окиснення жирних кислот
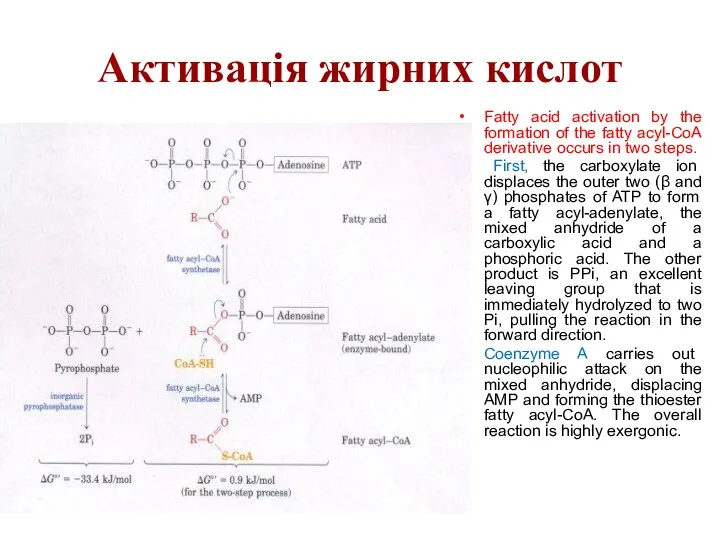
Активація жирних кислот
Fatty acid activation by the formation of the fatty
Активація жирних кислот
Fatty acid activation by the formation of the fatty
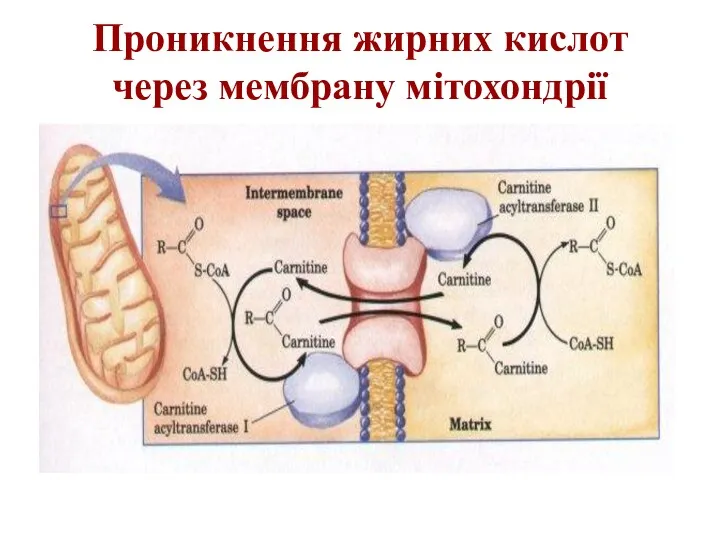
Проникнення жирних кислот через мембрану мітохондрії
Проникнення жирних кислот через мембрану мітохондрії
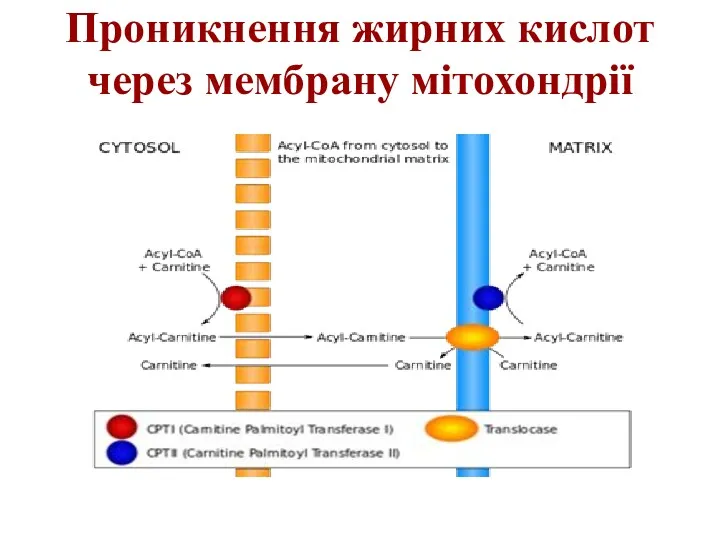
Проникнення жирних кислот через мембрану мітохондрії
Проникнення жирних кислот через мембрану мітохондрії
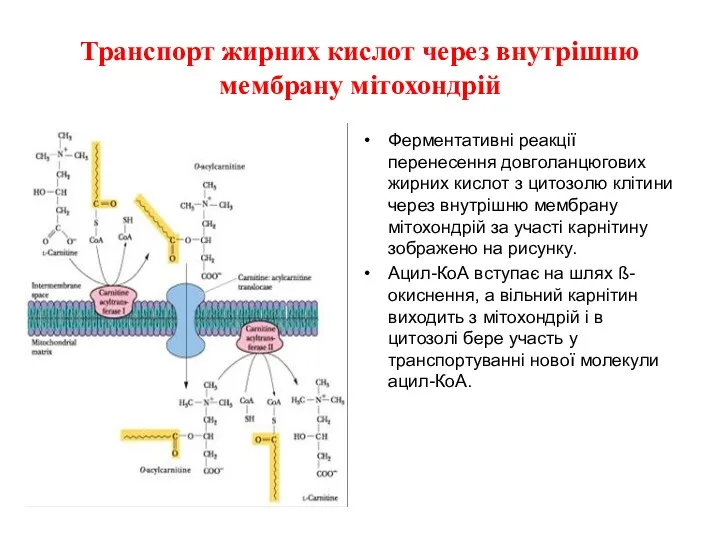
Транспорт жирних кислот через внутрішню мембрану мітохондрій
Ферментативні реакції перенесення довголанцюгових жирних
Транспорт жирних кислот через внутрішню мембрану мітохондрій
Ферментативні реакції перенесення довголанцюгових жирних

Карнітин
Карнітин

Проникнення жирних кислот через мембрану мітохондрії
Проникнення жирних кислот через мембрану мітохондрії

Стадія дегідрування
Стадія дегідрування

Стадія гідратації
Стадія гідратації

Друга стадія дегідрування
Друга стадія дегідрування

Тіолазна реакція
Тіолазна реакція

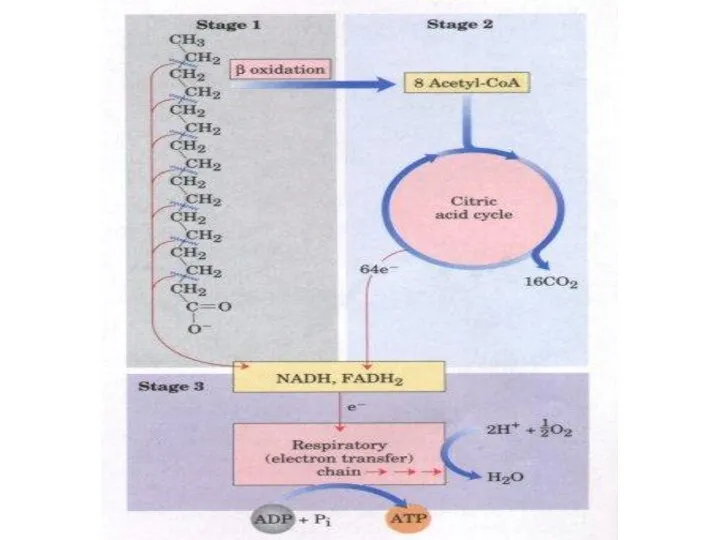
β-окислення
The fatty acid oxidation (β-oxidation) pathway.
(a) In each pass through
β-окислення
The fatty acid oxidation (β-oxidation) pathway.
(a) In each pass through

Енергетичний баланс β-окислення
Якщо жирна кислота містить n атомів карбону, то
Енергетичний баланс β-окислення
Якщо жирна кислота містить n атомів карбону, то

Енергетичний баланс β-окислення
У випадку пальмітинової кислоти (С 16) відбувається 7
Енергетичний баланс β-окислення
У випадку пальмітинової кислоти (С 16) відбувається 7


Окислення ненасичених жирних кислот
The oxidation of a monounsaturated fatty acyl-CoA, such
Окислення ненасичених жирних кислот
The oxidation of a monounsaturated fatty acyl-CoA, such
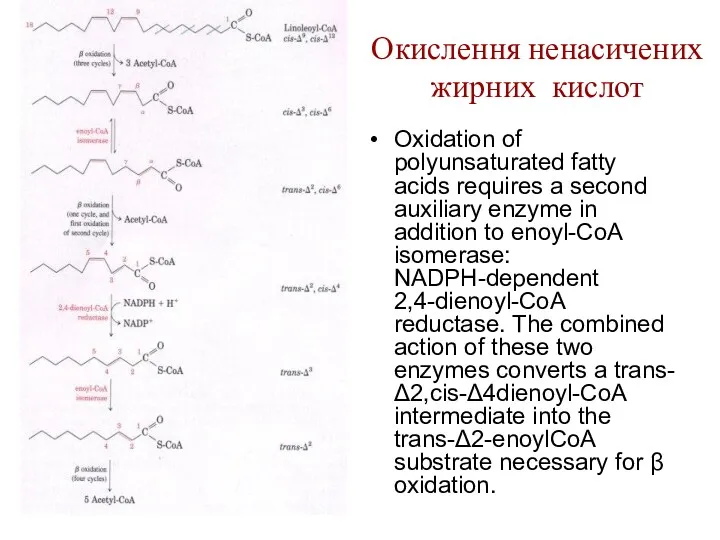
Окислення ненасичених жирних кислот
Oxidation of polyunsaturated fatty acids requires a second
Окислення ненасичених жирних кислот
Oxidation of polyunsaturated fatty acids requires a second

Кетонові тіла
Кетонові тіла
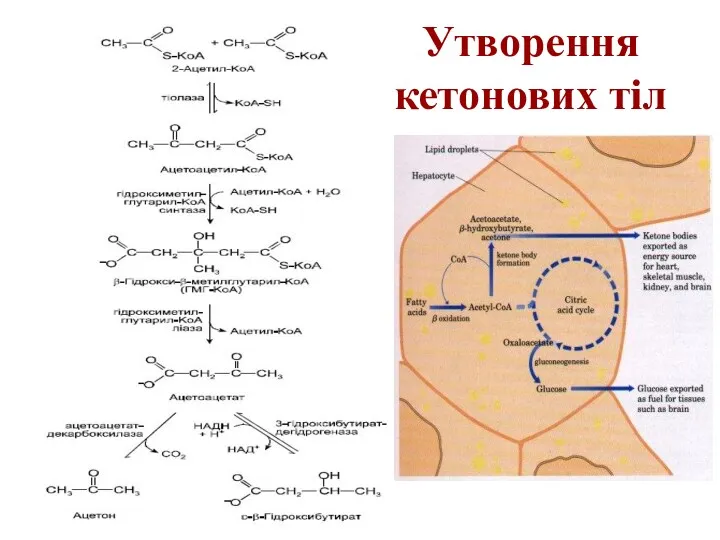
Утворення кетонових тіл
Утворення кетонових тіл

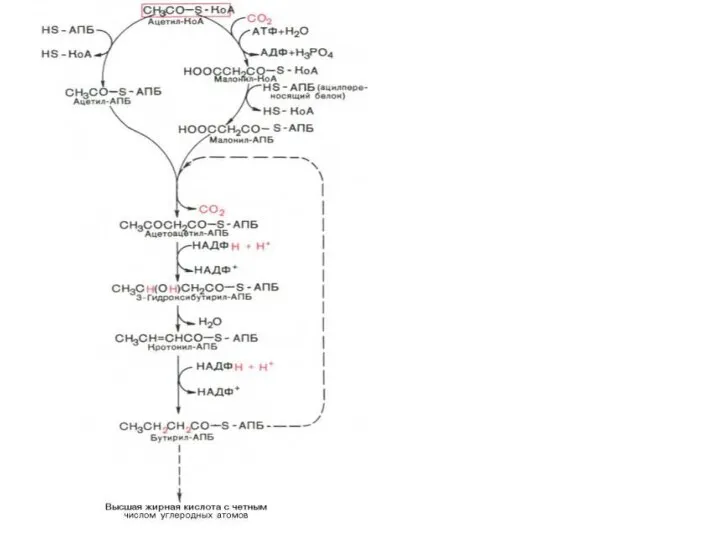
Синтез жирних кислот
Синтез жирних кислот

Синтез жирних кислот
Синтез жирних кислот

Синтаза вищих жирних кислот
Синтаза вищих жирних кислот



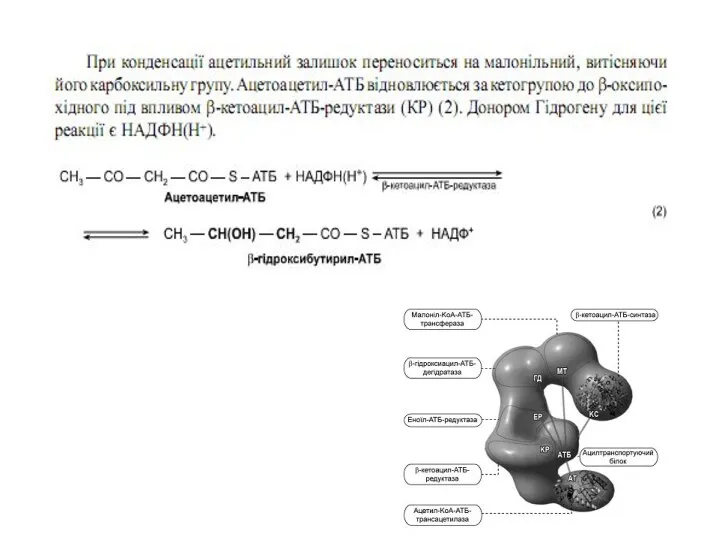



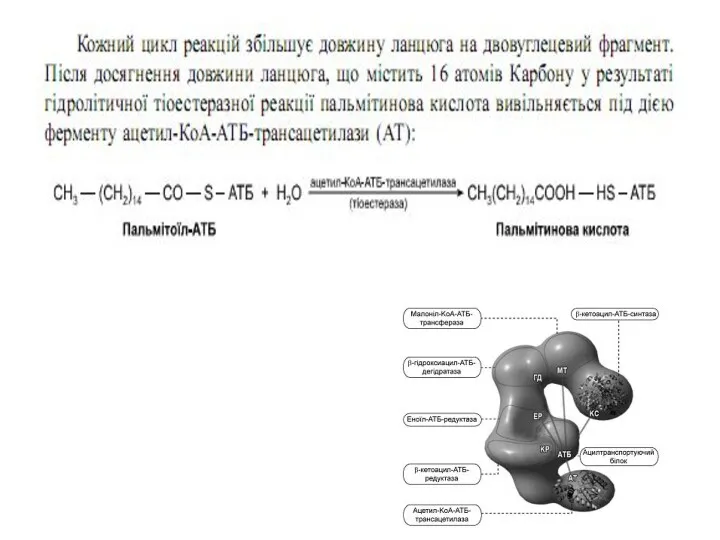

Сумарне рівняння синтезу жирних кислот
Із пальмітинової кислоти синтезується стеаринова і інші
Сумарне рівняння синтезу жирних кислот
Із пальмітинової кислоти синтезується стеаринова і інші

Синтез жирних кислот
Утворений малоніл~KоА взаємодіє з АТБ-SH під впливом малонілтрансацилази.
Синтез жирних кислот
Утворений малоніл~KоА взаємодіє з АТБ-SH під впливом малонілтрансацилази.


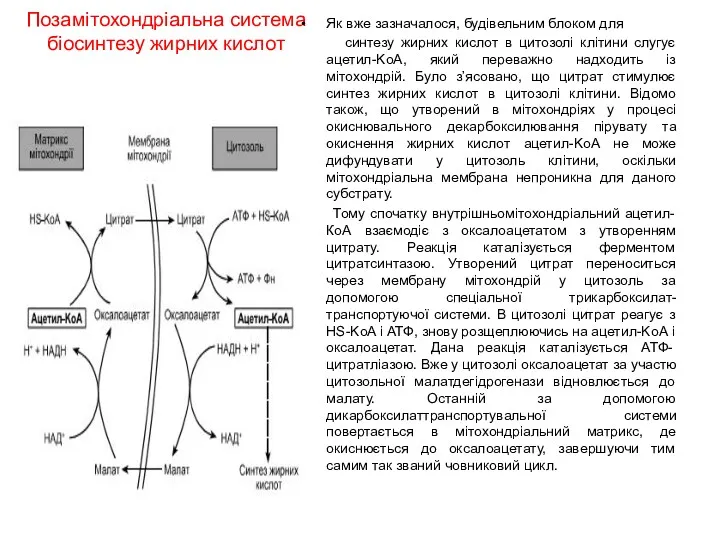
Позамітохондріальна система
біосинтезу жирних кислот
Як вже зазначалося, будівельним блоком для
синтезу
Позамітохондріальна система
біосинтезу жирних кислот
Як вже зазначалося, будівельним блоком для
синтезу

Утворення ненасичених жирних кислот
Утворення ненасичених жирних кислот

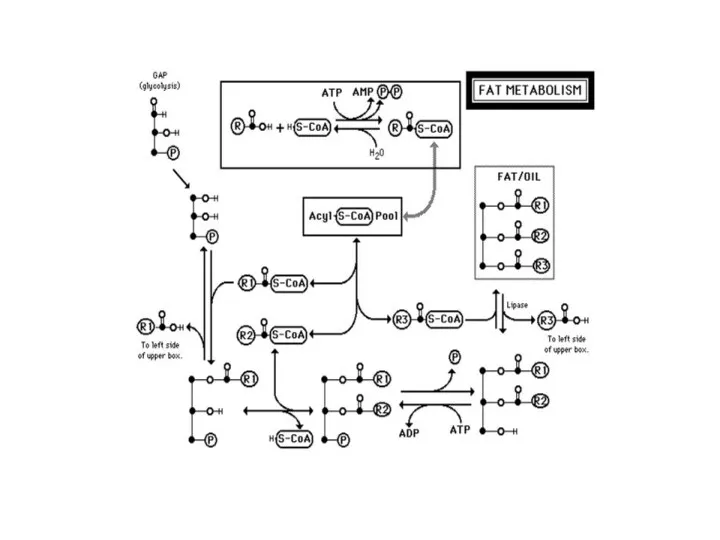
Біосинтез триацилгліцеролів
Біосинтез триацилгліцеролів

Біосинтез триацилгліцеролів
Біосинтез триацилгліцеролів

Біосинтез триацилгліцеролів
Біосинтез триацилгліцеролів
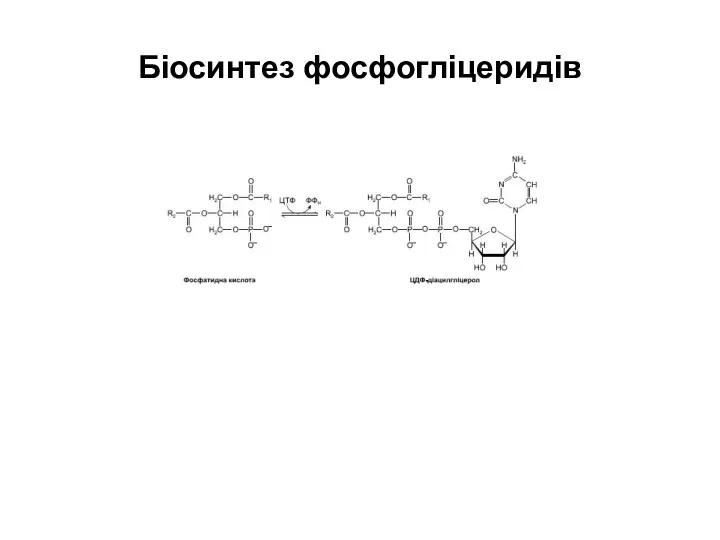
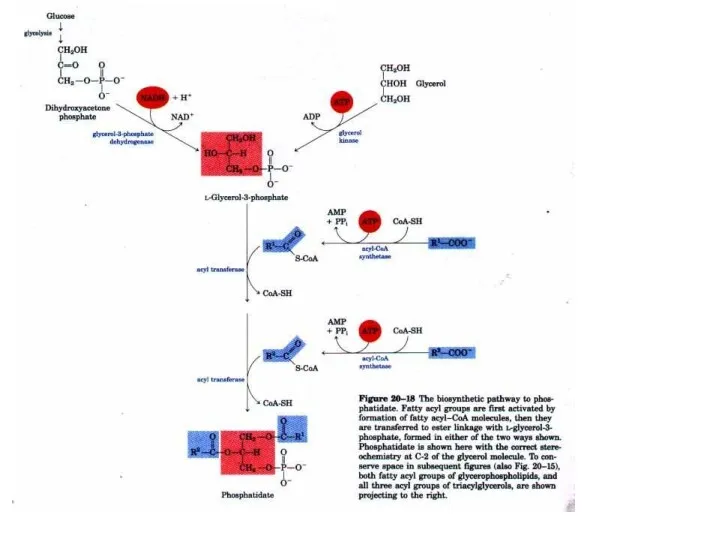
Біосинтез фосфогліцеридів
Біосинтез фосфогліцеридів

Біосинтез фосфогліцеридів
Біосинтез фосфогліцеридів
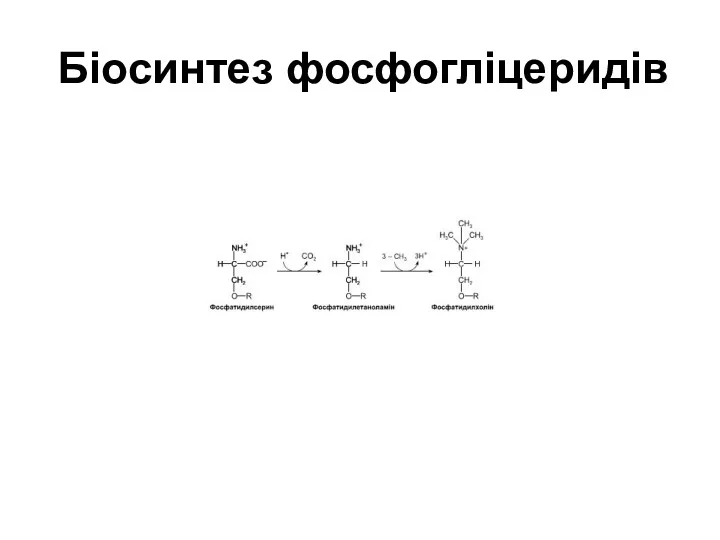
Біосинтез фосфогліцеридів
Біосинтез фосфогліцеридів

Біосинтез фосфогліцеридів
Біосинтез фосфогліцеридів

Біосинтез холестеролу
Біосинтез холестеролу

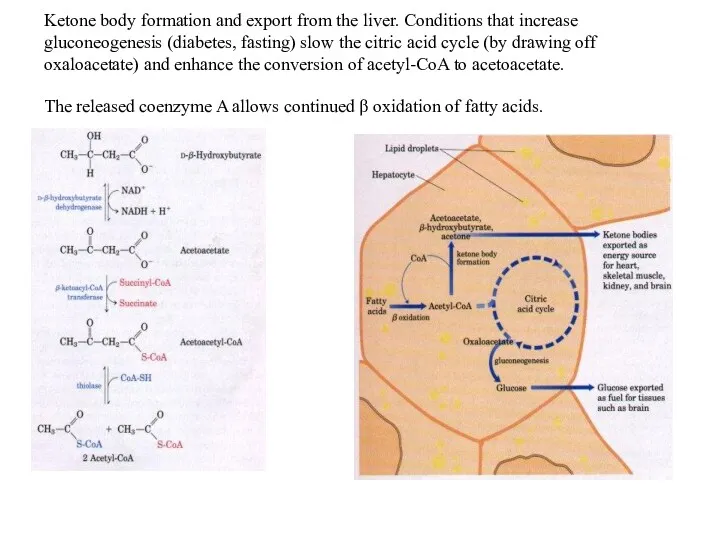
Ketone body formation and export from the liver. Conditions that increase
Ketone body formation and export from the liver. Conditions that increase
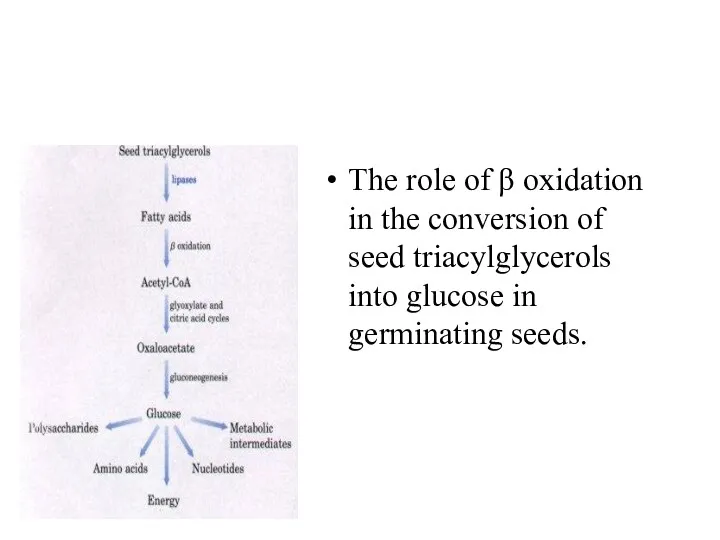
The role of β oxidation in the conversion of seed triacylglycerols
The role of β oxidation in the conversion of seed triacylglycerols

Утворення кетонових тіл
Formation of ketone bodies from acetyl-CoA. Under circumstances that
Утворення кетонових тіл
Formation of ketone bodies from acetyl-CoA. Under circumstances that

Electrons removed from fatty acids during β oxidation pass into the
Electrons removed from fatty acids during β oxidation pass into the


Синтез жирних кислот
Синтез жирних кислот
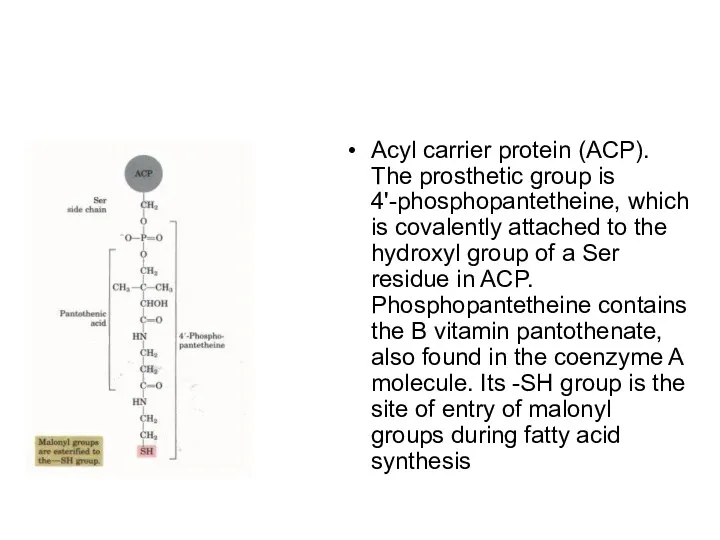
Acyl carrier protein (ACP). The prosthetic group is 4'-phosphopantetheine, which is
Acyl carrier protein (ACP). The prosthetic group is 4'-phosphopantetheine, which is


Beginning of the second round of the fatty acid synthesis cycle.
Beginning of the second round of the fatty acid synthesis cycle.
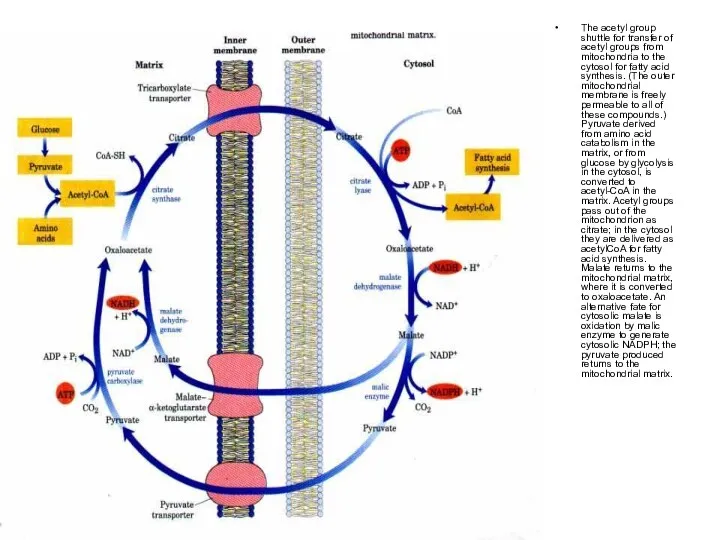
The acetyl group shuttle for transfer of acetyl groups from mitochondria
The acetyl group shuttle for transfer of acetyl groups from mitochondria

Regulation of fatty acid synthesis. (a) In the cells of vertebrates,
Regulation of fatty acid synthesis. (a) In the cells of vertebrates,
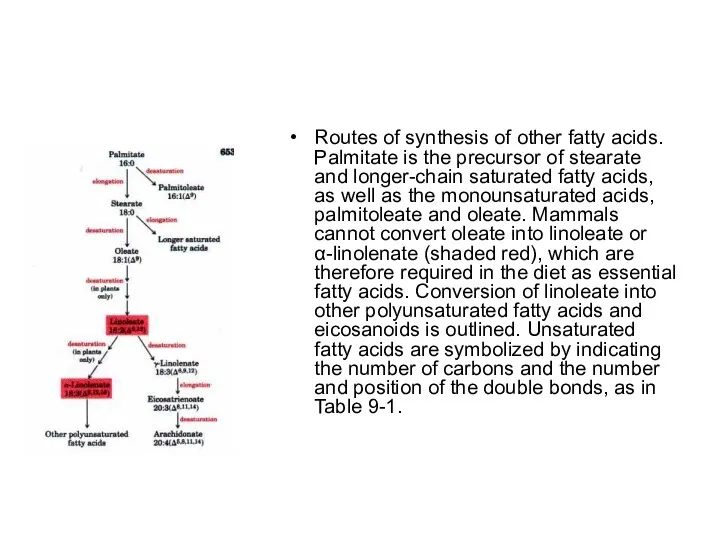
Routes of synthesis of other fatty acids. Palmitate is the precursor
Routes of synthesis of other fatty acids. Palmitate is the precursor
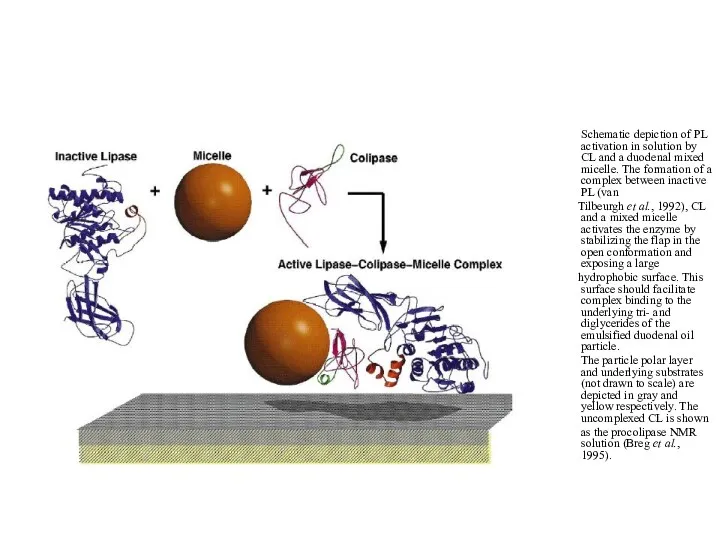
Schematic depiction of PL activation in solution by CL and
Schematic depiction of PL activation in solution by CL and
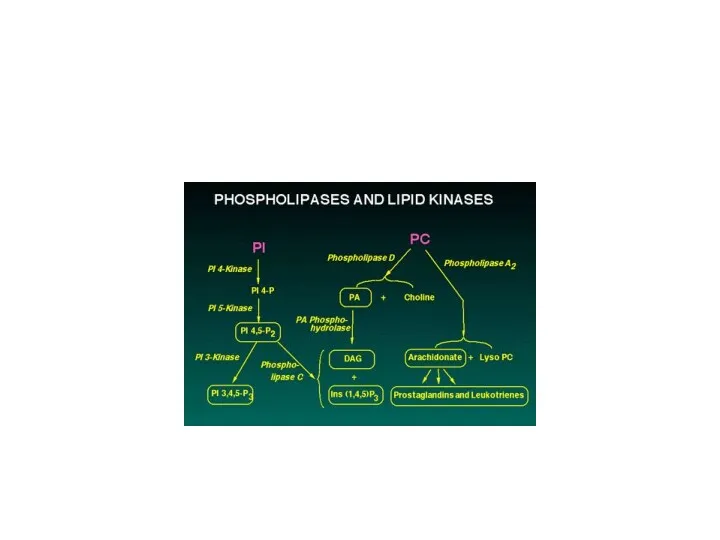
 Мышцы головы: лица (мимические) и жевательные. Фасции головы
Мышцы головы: лица (мимические) и жевательные. Фасции головы Обмен веществ и превращения энергии. Метаболизм
Обмен веществ и превращения энергии. Метаболизм История развития зоологии. Современная зоология
История развития зоологии. Современная зоология Биология как наука. Методы научного познания
Биология как наука. Методы научного познания Использование кейс-технологии на уроках биологии
Использование кейс-технологии на уроках биологии Гормоны и адаптация. (Лекция 15)
Гормоны и адаптация. (Лекция 15) Digestion
Digestion Протеомика. Методы молекулярной биологии и биинформатики в изучении белков
Протеомика. Методы молекулярной биологии и биинформатики в изучении белков Регуляция дыхания
Регуляция дыхания Память и обучение
Память и обучение Фотосинтез и дыхание. Презентация 6 класс
Фотосинтез и дыхание. Презентация 6 класс Класс Млекопитающие. Отряды Китообразные, Ластоногие, Хоботные, Хищные
Класс Млекопитающие. Отряды Китообразные, Ластоногие, Хоботные, Хищные Дигибридное скрещивание. Третий закон Г. Менделя
Дигибридное скрещивание. Третий закон Г. Менделя Черепные нервы I-VI
Черепные нервы I-VI Анатомия и физиология дыхательной системы
Анатомия и физиология дыхательной системы Пищеварение в кишечнике. Пищеварение в тонкой кишке
Пищеварение в кишечнике. Пищеварение в тонкой кишке Презентации
Презентации Организм человека, как единое целое
Организм человека, как единое целое Нуклеиновые кислоты
Нуклеиновые кислоты Низкомолекулярные биорегуляторы терпены
Низкомолекулярные биорегуляторы терпены Електричні явища у мязових волокнах. (Лекція 5)
Електричні явища у мязових волокнах. (Лекція 5) Формы размножения организмов. Митоз
Формы размножения организмов. Митоз Методы исследования в микробиологии. Бактериологический метод исследования
Методы исследования в микробиологии. Бактериологический метод исследования ПрезентацияДомашний зоопарк
ПрезентацияДомашний зоопарк Родительское поведение у животных. (Тема 9)
Родительское поведение у животных. (Тема 9) Полимеразная цепная реакция
Полимеразная цепная реакция Презентация к уроку биологии в 6 классе по программе VIII вида Гранит и его свойства
Презентация к уроку биологии в 6 классе по программе VIII вида Гранит и его свойства Здоровое питание. Технология, 5 класс
Здоровое питание. Технология, 5 класс