- Обмін речовин та енергії (метаболізм)

Содержание
- 2. Обмін речовин (метаболізм) – сукупність всіх хімічних реакцій і всіх видів перетворень речовин та енергії в
- 3. Хімічні речовини, що беруть участь в обміні речовин називаються метаболітами. Поза клітиною майже всі ці перетворення
- 4. Метаболізм забезпечує: 1. Розвиток 2. Життєдіяльність 3. Самовідтворення організмів 4. Зв'язок з оточуючим середовищем 5. Адаптацію
- 5. Метаболізм складається з анаболізму (процеси синтезу) і катаболізму (процеси розпаду речовин)
- 7. Основа обміну речовин Асиміляція (анаболізм): хімічні перетворення, що приводять до використання органічних речовин, які надходять із
- 8. Ці процеси взаємопов'язані і направлені на безперервне поновлення живого матеріалу і забезпечення його необхідною енергією. Організм
- 9. Живі клітини постійно потребують органічні і неорганічні речовини, а також хімічну енергію, яку вони отримують переважно
- 10. Із багаточисельних метаболітів виділяють 3 найбільш важливих – піруват (ПВК), ацетил-КоА і гліцерин. Вони є ланкою,
- 12. Ні вуглеводи, ні жири, ні білки не можуть безпосередньо бути «паливом» для клітинних процесів. Клітини тварин
- 13. Макроергічні сполуки - це сполуки, що містять макроергічні (багаті на енергію) фосфатні зв'язки. В результаті їх
- 14. Центральне місце серед макроергічних сполук займає АТФ – молекула, багата енергією, оскільки вона містить 2 фосфоефірних
- 15. Структура АТФ
- 16. Аденозинтрифосфорна кислота (АТФ)
- 17. АТФ – універсальна «енергетична валюта» всіх живих організмів. АТФ мобільна, потрапляє в любу частину клітини, де
- 18. В клітині молекула АТФ використовується протягом 1 хвилини після її утворення. У людини кількість АТФ, рівне
- 19. Біологічне окиснення може відбуватися кількома шляхами: - відщеплення від субстратів атомів Гідрогену і електронів (дегідрування): на
- 20. Біологічне окиснення Це сукупність окисно-відновних процесів у біологічних об'єктах, які відбуваються в клітинах організму в період
- 21. Функції біологічного окиснення: Енергетичне забезпечення процесів життєдіяльності: підтримання температури тіла, хімічні синтези, електричні процеси, механічна робота.
- 22. Окиснення органічних речовин Оксигеном до СО2 і Н2О називається тканинним диханням або біологічним окисненням.
- 23. Тканинне дихання відбувається за участі ланцюгу перенесення електронів (ЛПЕ) або дихального ланцюгу, який утворений окисно-відновними ферментами,
- 26. Уперше мітохондрії (МХ) були виявлені у вигляді гранул у м'язових клітинах (1850 р.). Кількість МХ у
- 27. Дихальний ланцюг Сукупність ферментів, що здійснюють транспорт електронів від речовини, що окиснюється, на О2 називається дихальним
- 28. В ЛПЕ приймають участь 4 групи ферментів класу оксидоредуктаз: піридинзалежні дегідрогенази, флавінзалежні дегідрогенази, убіхінон або коензим
- 33. Піридинзалежні дегідрогенази (двохкомпонентні) Специфічний білок (апофермент) + кофермент НАД або НАДФ. Є універсальними акцепторами Гідрогену для
- 34. NAD - залежна дегідрогеназа каталізує реакції окиснення безпосередньо субстрату (α-кетоглутарат, ізоцитрат, ПВК, малат, глутамат та ін.).
- 36. Флавінзалежні дегідрогенази (двохкомпонентні) Специфічний білок (апофермент) + кофермент ФМН або ФАД. Флавінові коферменти щільно зв'язані з
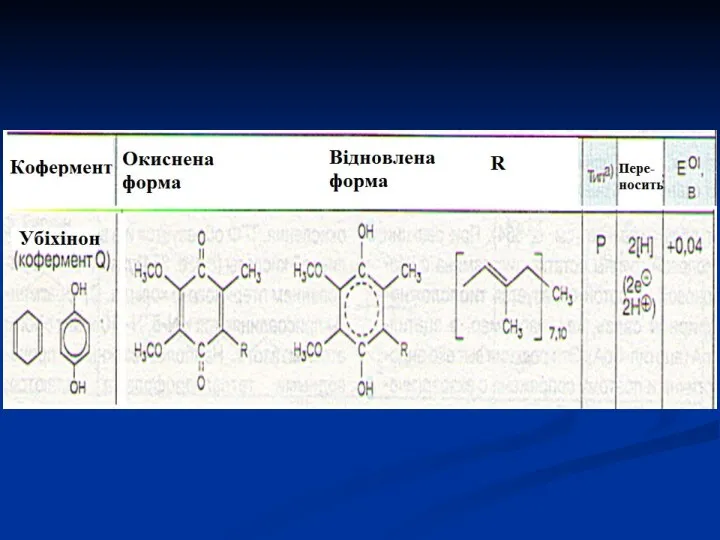
- 38. УБІХІНОН або коензим Q – проміжний переносник атомів Гідрогену, тобто електронів і протонів в мітохондріальній мембрані,
- 40. Цитохроми Відносяться до гемвмісних хромопротеїнів. Їх простетичною групою є гем, у якого атом заліза має здатність
- 42. В природі існують сімейства цитохромів (в, с, а), які відрізняються між собою величиною електричного потенціалу, який
- 43. Редокс-потенціал Напрямок потоку електронів при спряженні однієї окисної системи з іншою визначається їх стандартними окисно-відновними потенціалами
- 44. Зміна значень окисно-відновних потенціалів компонентів дихального ланцюга мітохондрій
- 45. Ендогенна вода Убіхінон передає на систему цитохромів тільки електрони, а протони (Н+) надходять в середовище, де
- 46. Інгібітори дихального ланцюгу Речовини, що блокують певні етапи цього процесу. Для НАД-залежної дегідрогенази: - ротенон –
- 47. Окисне фосфорилування Синтез АТФ з АДФ відбувається в результаті окисного фосфорилування, яке сполучене з процесом біологічного
- 49. створюється протонний градієнт, при якому концентрація протонів у міжмембранному просторі більше, а рН менше, ніж у
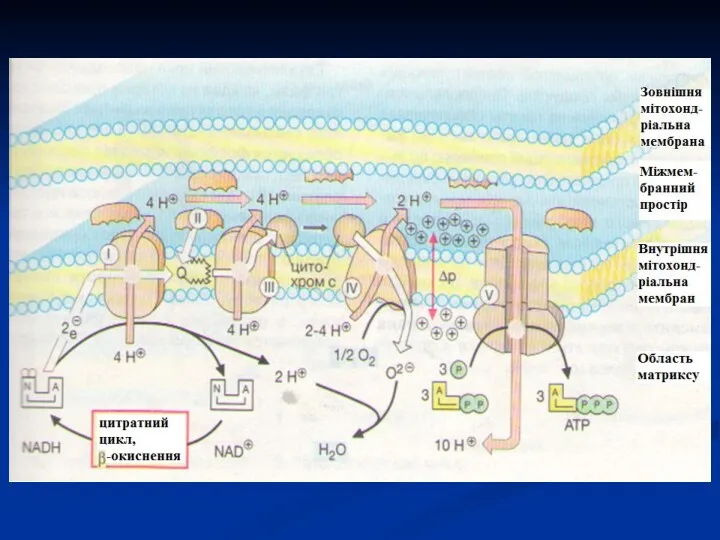
- 50. В дихальному ланцюгу можна виділити 3 ділянки, в яких перенесення електронів супроводжується зниженням вільної енергії. Ці
- 51. Енергія електрохімічного потенціалу використовується для синтезу АТФ, якщо протони повертаються в матрикс через іонні канали АТФ-синтази.
- 52. АТФ-синтаза Складається з 2-х білкових комплексів. Гідрофобний комплекс занурений в мембрану. Він є основою, яка фіксує
- 53. Для синтезу 1 моля АТФ з АДФ необхідно 7,5 ккал енергії, що є рівносильним різниці редокс-потенціалів
- 54. Мітохондріальний ланцюг перенесення електронів
- 55. В НАД-залежних реакціях при окисненні 1 моля субстрату утворюється 3 АТФ, у ФАД-залежних реакціях – 2
- 56. Роз'єднання дихання і фосфорилування Деякі хімічні речовини (протонофори) можуть переносити протони або інші іони (іонофори) з
- 57. В результаті роз'єднання кількість АТФ зменшується, а АДФ збільшується. Швидкість окиснення НАДН і ФАДН2 підвищується, збільшується
- 58. МІКРОСОМАЛЬНЕ ОКИСНЕННЯ Це ще один тип біологічного окиснення, який відбувається в мікросомах (переважно в печінці і
- 59. До ферментів мікросомального окиснення відтносяться МОНООКСИГЕНАЗИ і ДИОКСИГЕНАЗИ. Монооксигенази (гідроксилази) при окисненні речовини включають 1 атом
- 60. ФУНКЦІЇ МІКРОСОМАЛЬНОГО ОКИСНЕННЯ: Окиснення речовин циклічної структури, які не можуть окиснюватися шляхом дегідрування (триптофан, гомогентизинова кислота
- 61. Утворення пероксиду водню. При тканинному диханні можливе утворення пероксиду водню (Н2О2), який є сильним окисником сульфгідрильних
- 63. Скачать презентацию

Обмін речовин (метаболізм) – сукупність всіх хімічних реакцій і всіх видів
Обмін речовин (метаболізм) – сукупність всіх хімічних реакцій і всіх видів

Хімічні речовини, що беруть участь в обміні речовин називаються метаболітами.
Поза
Хімічні речовини, що беруть участь в обміні речовин називаються метаболітами.
Поза

Метаболізм забезпечує:
1. Розвиток
2. Життєдіяльність
3. Самовідтворення організмів
4. Зв'язок з оточуючим середовищем
5. Адаптацію
Метаболізм забезпечує: 1. Розвиток 2. Життєдіяльність 3. Самовідтворення організмів 4. Зв'язок з оточуючим середовищем 5. Адаптацію

Метаболізм складається з анаболізму (процеси синтезу) і катаболізму (процеси розпаду речовин)
Метаболізм складається з анаболізму (процеси синтезу) і катаболізму (процеси розпаду речовин)


Основа обміну речовин
Асиміляція (анаболізм):
хімічні перетворення, що приводять до використання органічних речовин,
Основа обміну речовин
Асиміляція (анаболізм):
хімічні перетворення, що приводять до використання органічних речовин,

Ці процеси взаємопов'язані і направлені на безперервне поновлення живого матеріалу і
Ці процеси взаємопов'язані і направлені на безперервне поновлення живого матеріалу і

Живі клітини постійно потребують органічні і неорганічні речовини, а також хімічну
Живі клітини постійно потребують органічні і неорганічні речовини, а також хімічну

Із багаточисельних метаболітів виділяють 3 найбільш важливих – піруват (ПВК), ацетил-КоА
Із багаточисельних метаболітів виділяють 3 найбільш важливих – піруват (ПВК), ацетил-КоА


Ні вуглеводи, ні жири, ні білки не можуть безпосередньо бути «паливом»

Макроергічні сполуки
- це сполуки, що містять макроергічні (багаті на енергію) фосфатні
Макроергічні сполуки
- це сполуки, що містять макроергічні (багаті на енергію) фосфатні

Центральне місце серед макроергічних сполук займає АТФ – молекула, багата енергією,
Центральне місце серед макроергічних сполук займає АТФ – молекула, багата енергією,
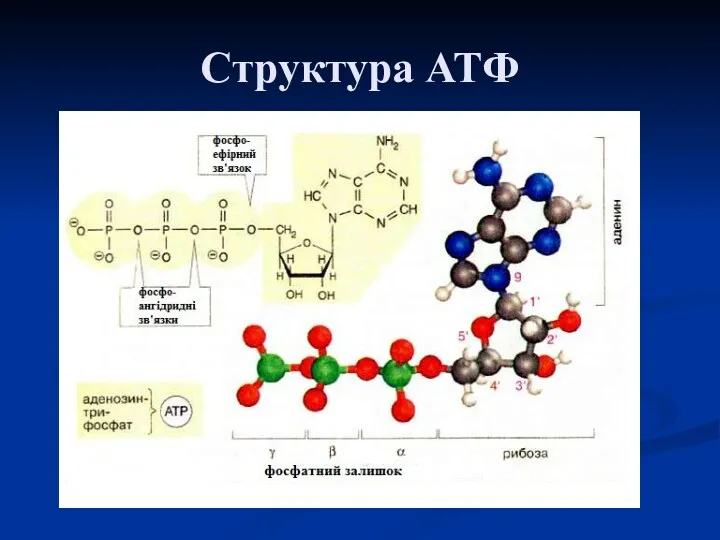
Структура АТФ
Структура АТФ

Аденозинтрифосфорна кислота (АТФ)
Аденозинтрифосфорна кислота (АТФ)

АТФ – універсальна «енергетична валюта» всіх живих організмів.
АТФ мобільна, потрапляє
АТФ – універсальна «енергетична валюта» всіх живих організмів.
АТФ мобільна, потрапляє

В клітині молекула АТФ використовується протягом 1 хвилини після її утворення.
В клітині молекула АТФ використовується протягом 1 хвилини після її утворення.

Біологічне окиснення
може відбуватися кількома шляхами:
- відщеплення від субстратів атомів Гідрогену
Біологічне окиснення
може відбуватися кількома шляхами:
- відщеплення від субстратів атомів Гідрогену

Біологічне окиснення
Це сукупність окисно-відновних процесів у біологічних об'єктах, які відбуваються в
Біологічне окиснення
Це сукупність окисно-відновних процесів у біологічних об'єктах, які відбуваються в

Функції біологічного окиснення:
Енергетичне забезпечення процесів життєдіяльності: підтримання температури тіла, хімічні синтези,
Функції біологічного окиснення:
Енергетичне забезпечення процесів життєдіяльності: підтримання температури тіла, хімічні синтези,

Окиснення органічних речовин Оксигеном до СО2 і Н2О називається тканинним диханням
Окиснення органічних речовин Оксигеном до СО2 і Н2О називається тканинним диханням

Тканинне дихання відбувається за участі
ланцюгу перенесення електронів (ЛПЕ) або
Тканинне дихання відбувається за участі
ланцюгу перенесення електронів (ЛПЕ) або



Уперше мітохондрії (МХ) були виявлені у вигляді гранул у м'язових клітинах
Уперше мітохондрії (МХ) були виявлені у вигляді гранул у м'язових клітинах

Дихальний ланцюг
Сукупність ферментів, що здійснюють транспорт електронів від речовини, що окиснюється,
Дихальний ланцюг
Сукупність ферментів, що здійснюють транспорт електронів від речовини, що окиснюється,

В ЛПЕ приймають участь
4 групи ферментів класу оксидоредуктаз:
піридинзалежні дегідрогенази,
флавінзалежні
В ЛПЕ приймають участь
4 групи ферментів класу оксидоредуктаз:
піридинзалежні дегідрогенази,
флавінзалежні




Піридинзалежні дегідрогенази (двохкомпонентні)
Специфічний білок (апофермент) + кофермент НАД або НАДФ.
Є
Піридинзалежні дегідрогенази (двохкомпонентні)
Специфічний білок (апофермент) + кофермент НАД або НАДФ.
Є
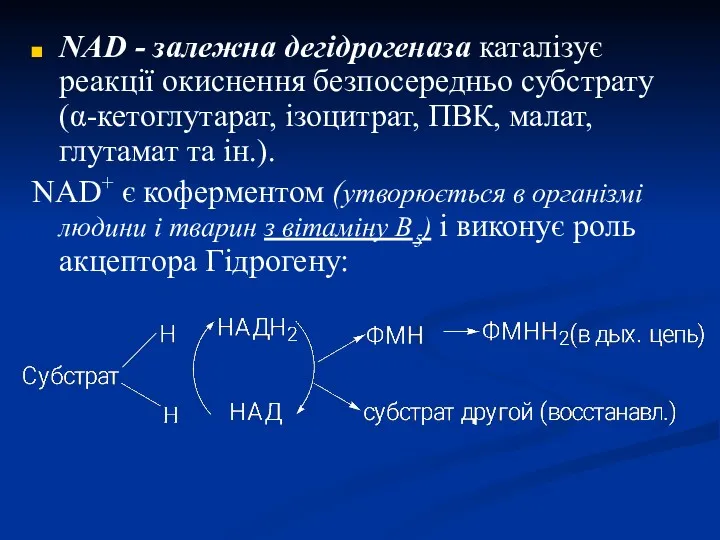
NAD - залежна дегідрогеназа каталізує реакції окиснення безпосередньо субстрату (α-кетоглутарат, ізоцитрат,
NAD - залежна дегідрогеназа каталізує реакції окиснення безпосередньо субстрату (α-кетоглутарат, ізоцитрат,


Флавінзалежні дегідрогенази (двохкомпонентні)
Специфічний білок (апофермент) + кофермент ФМН або ФАД.
Флавінові коферменти
Флавінзалежні дегідрогенази (двохкомпонентні)
Специфічний білок (апофермент) + кофермент ФМН або ФАД.
Флавінові коферменти


УБІХІНОН або коензим Q – проміжний переносник атомів Гідрогену, тобто електронів
УБІХІНОН або коензим Q – проміжний переносник атомів Гідрогену, тобто електронів

Цитохроми
Відносяться до гемвмісних хромопротеїнів. Їх простетичною групою є гем, у
Цитохроми
Відносяться до гемвмісних хромопротеїнів. Їх простетичною групою є гем, у


В природі існують сімейства цитохромів (в, с, а), які відрізняються між
В природі існують сімейства цитохромів (в, с, а), які відрізняються між

Редокс-потенціал
Напрямок потоку електронів при спряженні однієї окисної системи з іншою визначається
Редокс-потенціал
Напрямок потоку електронів при спряженні однієї окисної системи з іншою визначається
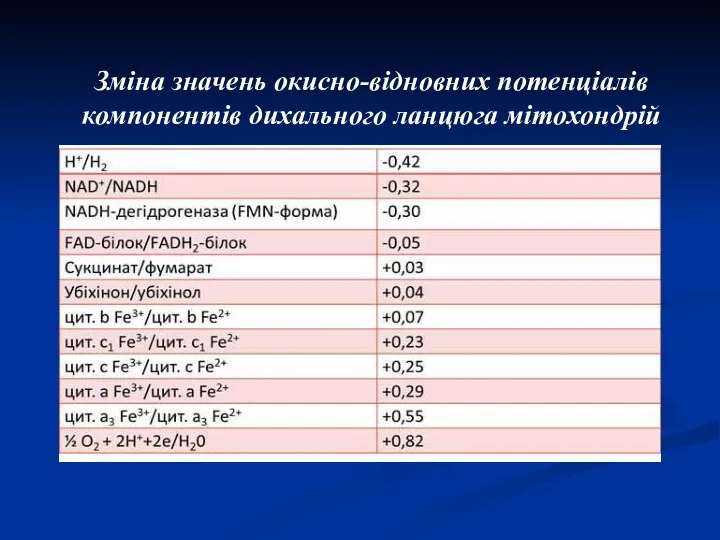
Зміна значень окисно-відновних потенціалів компонентів дихального ланцюга мітохондрій
Зміна значень окисно-відновних потенціалів компонентів дихального ланцюга мітохондрій

Ендогенна вода
Убіхінон передає на систему цитохромів тільки електрони, а протони (Н+)
Ендогенна вода
Убіхінон передає на систему цитохромів тільки електрони, а протони (Н+)

Інгібітори дихального ланцюгу
Речовини, що блокують певні етапи цього процесу.
Інгібітори дихального ланцюгу
Речовини, що блокують певні етапи цього процесу.

Окисне фосфорилування
Синтез АТФ з АДФ відбувається в результаті окисного фосфорилування, яке
Окисне фосфорилування
Синтез АТФ з АДФ відбувається в результаті окисного фосфорилування, яке
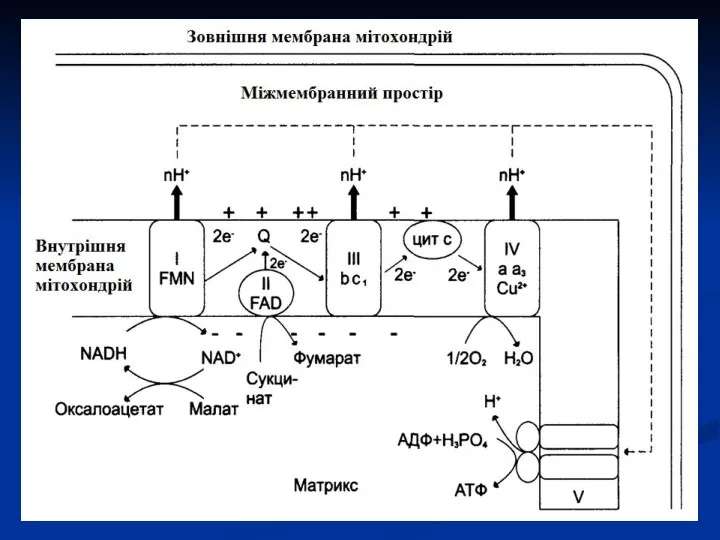

створюється протонний градієнт, при якому концентрація протонів у міжмембранному просторі більше,
створюється протонний градієнт, при якому концентрація протонів у міжмембранному просторі більше,

В дихальному ланцюгу можна виділити 3 ділянки, в яких перенесення електронів
В дихальному ланцюгу можна виділити 3 ділянки, в яких перенесення електронів

Енергія електрохімічного потенціалу використовується для синтезу АТФ, якщо протони повертаються в
Енергія електрохімічного потенціалу використовується для синтезу АТФ, якщо протони повертаються в

АТФ-синтаза
Складається з 2-х білкових комплексів.
Гідрофобний комплекс занурений в мембрану. Він
АТФ-синтаза
Складається з 2-х білкових комплексів.
Гідрофобний комплекс занурений в мембрану. Він

Для синтезу 1 моля АТФ з АДФ необхідно
7,5 ккал енергії, що
Для синтезу 1 моля АТФ з АДФ необхідно
7,5 ккал енергії, що
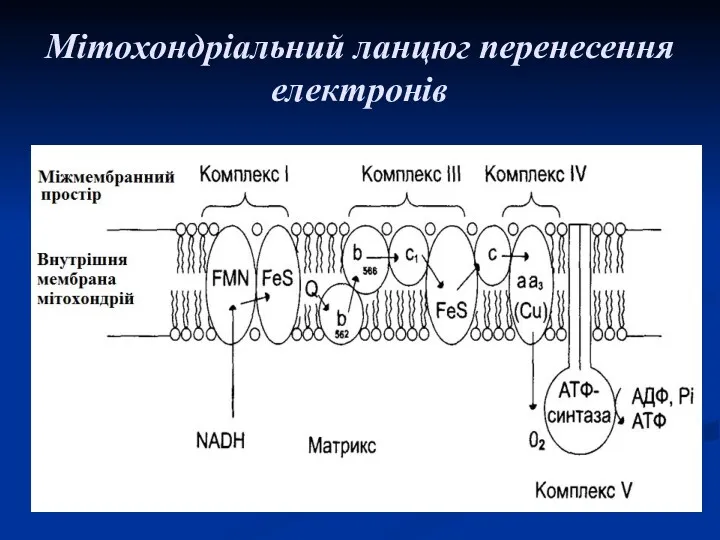
Мітохондріальний ланцюг перенесення електронів
Мітохондріальний ланцюг перенесення електронів

В НАД-залежних реакціях при окисненні 1 моля субстрату утворюється 3 АТФ,
у
В НАД-залежних реакціях при окисненні 1 моля субстрату утворюється 3 АТФ,
у

Роз'єднання дихання і фосфорилування
Деякі хімічні речовини (протонофори) можуть переносити протони або
Роз'єднання дихання і фосфорилування
Деякі хімічні речовини (протонофори) можуть переносити протони або

В результаті роз'єднання кількість АТФ зменшується, а АДФ збільшується. Швидкість окиснення
В результаті роз'єднання кількість АТФ зменшується, а АДФ збільшується. Швидкість окиснення

МІКРОСОМАЛЬНЕ ОКИСНЕННЯ
Це ще один тип біологічного окиснення, який відбувається в мікросомах
МІКРОСОМАЛЬНЕ ОКИСНЕННЯ
Це ще один тип біологічного окиснення, який відбувається в мікросомах

До ферментів мікросомального окиснення відтносяться МОНООКСИГЕНАЗИ і ДИОКСИГЕНАЗИ.
Монооксигенази (гідроксилази) при
До ферментів мікросомального окиснення відтносяться МОНООКСИГЕНАЗИ і ДИОКСИГЕНАЗИ.
Монооксигенази (гідроксилази) при

ФУНКЦІЇ МІКРОСОМАЛЬНОГО ОКИСНЕННЯ:
Окиснення речовин циклічної структури, які не можуть окиснюватися шляхом
ФУНКЦІЇ МІКРОСОМАЛЬНОГО ОКИСНЕННЯ:
Окиснення речовин циклічної структури, які не можуть окиснюватися шляхом

Утворення пероксиду водню.
При тканинному диханні можливе утворення пероксиду водню (Н2О2), який
Утворення пероксиду водню.
При тканинному диханні можливе утворення пероксиду водню (Н2О2), який
 Общее учение о нервной системе (неврология). Развитие нервной системы. Спинной мозг
Общее учение о нервной системе (неврология). Развитие нервной системы. Спинной мозг Индивидуальное развитие животных
Индивидуальное развитие животных Заказник Урочище Берикпара, Южный Алтай
Заказник Урочище Берикпара, Южный Алтай Общая характеристика и разнообразие хрящевых рыб. Их роль в водной экосистеме
Общая характеристика и разнообразие хрящевых рыб. Их роль в водной экосистеме Круглоротые (бесчелюстные) cyclostomata
Круглоротые (бесчелюстные) cyclostomata Физиология центральной нервной системы. Нейроны, синапсы, медиаторы. (Часть 1)
Физиология центральной нервной системы. Нейроны, синапсы, медиаторы. (Часть 1) Комнатные растения
Комнатные растения Скелет туловища. Соединение позвонков
Скелет туловища. Соединение позвонков Вегетативное размножение растений и его использование человеком. 6 класс
Вегетативное размножение растений и его использование человеком. 6 класс Творческий проект. Тема: Вода
Творческий проект. Тема: Вода Введение в физиологию. Возбудимые ткани
Введение в физиологию. Возбудимые ткани Особенности движения веществ в организмах животных
Особенности движения веществ в организмах животных Исследовательская работа на тему Эрозия почвы у супермаркета О_кей
Исследовательская работа на тему Эрозия почвы у супермаркета О_кей Моллюски
Моллюски Великий круговорот жизни (окружающий мир, 3 класс)
Великий круговорот жизни (окружающий мир, 3 класс) Сбор и методы сохранения беспозвоночных животных
Сбор и методы сохранения беспозвоночных животных Современное состояние и охрана растительности
Современное состояние и охрана растительности Происхождение человека
Происхождение человека Эволюция Вселенной
Эволюция Вселенной Роль воды и воздуха для прорастания семян гороха
Роль воды и воздуха для прорастания семян гороха Удивительные овощи. Фото-отчет по выставке
Удивительные овощи. Фото-отчет по выставке Тест органоиды клетки
Тест органоиды клетки Семя и его строение
Семя и его строение Строение цветка
Строение цветка Бактерии. Строение клетки
Бактерии. Строение клетки Зимний сад
Зимний сад Царство Вирусы
Царство Вирусы Урок биологии на тему Плесневые грибы и дрожжи, 6 класс
Урок биологии на тему Плесневые грибы и дрожжи, 6 класс