- Общая схема организации двигательной активности

Содержание
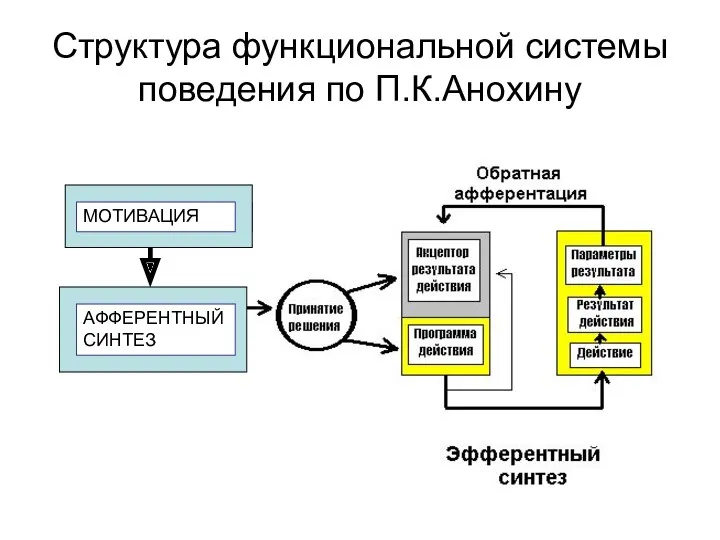
- 2. Структура функциональной системы поведения по П.К.Анохину МОТИ ВАЦИЯ МОТИВАЦИЯ АФФЕРЕНТНЫЙ СИНТЕЗ
- 3. ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ АФФЕРЕНТНЫЙ СИНТЕЗ ПРИНЯТИЕ РЕШЕНИЯ ЧТО ДЕЛАТЬ? ЭФФЕРЕНТНЫЙ СИНТЕЗ ПРИНЯТИЕ РЕШЕНИЯ КАК ДЕЛАТЬ?
- 4. Схема организации двигательной системы. В иерархическом порядке представлены связи между центрами нервной системы, участвующими в регуляции
- 5. Схема связей в двигательной системе
- 6. ЛИМБИЧЕСКАЯ СИСТЕМА
- 7. Лимбическая система Функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее эмоционально-мотивационные компоненты поведения и
- 8. Афферентные входы От различных областей головного мозга Через гипоталамус от ретикулярной формации ствола (один из главных
- 9. Лимбическая система
- 10. ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ 1. Организация вегетативно-соматических компонентов эмоций 2. Организация кратковременной и долговременной памяти 3. Участие
- 11. Кольцевые нейрональные связи Дают возможность длительной циркуляции (реверберации) возбуждения, что создает условия для сохранения единого функционального
- 12. 2 ЛИМБИЧЕСКИХ КРУГА БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела - мамиллярно-таламический пучок Вик-д’Азира -
- 13. Функциональная структура лимбики по МакЛину (1970) 1. Нижний отдел - миндалина и гиппокамп - центры эмоций
- 14. Функции лимбической системы После обработки информации от внешней среды и внутренних органах запускает вегетативные, соматические и
- 15. Физиология гиппокампа
- 17. Физиология гиппокампа Гиппокамп (hippocampus) является основной структурой лимбической системы. Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными
- 18. Физиология гиппокампа Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа, а тем самым —
- 19. Физиология миндалевидного тела
- 21. Физиология миндалевидного тела Миндалевидное тело, миндалина — подкорковая структура лимбической системы, расположенная в глубине височной доли
- 22. Физиология миндалевидного тела При искусственной активации миндалины появляются реакции: принюхивания, облизывания, жевания, глотания, саливации, изменения перистальтики
- 23. Влияние миндалин на иерархические отношения в стае До операции После удаления миндалин у Дейва
- 24. Физиология гипоталамуса Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга, расположенный ниже таламуса, или «зрительных бугров»,
- 25. Основные структуры гипоталамуса
- 26. ФУНКЦИИ ГИПОТАЛАМУСА Высший центр регуляции вегетативной нервной системы Высший центр регуляции эндокринных функций Регуляция мотиваций пищевого
- 27. Поведенческие функции гипоталамуса Эффекты стимуляции гипоталамуса Латеральный гипоталамус: жажда, аппетит, увеличение активности организма, ярость, агрессия. Вентромедиальное
- 28. Центры поощрения В различные области мозга обезьяны вводили электроды, и обезьяна, нажимая на рычаг, включала контакты
- 29. Центры поощрения и наказания Личные оценки могут характеризовать ощущения как приятные или неприятные (поощрительные или наказывающие,
- 30. Главные (первичные) центры поощрения располагаются по ходу медиального пучка переднего мозга и особенно в латеральном и
- 31. Центры наказания и реакции избегания расположены в центральном сером веществе, окружающем сильвиев водопровод, в среднем мозге
- 32. Ярость возникает при активации центров наказания. В этом состоянии даже легкая провокация вызывает нападение. Противоположная эмоциональная
- 33. Роль поощрения и наказания в поведении, обучении и памяти Всё или почти все, что делает человек,
- 34. Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших полушарий. Повторение этих же стимулов
- 35. Регуляция вегетативных (висцеральных) функций Осуществляется преимущественно через гипоталамус, куда информация поступает от различных участков лимбической системы
- 36. Роль лимбической системы в обучении Связана с кругом Пейпеца, где главную роль играют гиппокамп и связанные
- 37. ЭМОЦИОНАЛЬНЫЙ МОЗГ
- 38. ЦИТАТА «Является ли эмоция продуктом волшебства или физиологическим процессом, который зависит от анатомического механизма? Я думаю
- 39. Формирование эмоций (пережеваний) Наиболее важная функция лимбической системы. В свою очередь, эмоции, являются субъективным компонентом мотиваций,
- 40. Эмоция — специфическое состояние психической сферы, обусловленная рефлекторной реакцией организма на внешние и внутренние раздражения, характеризующееся
- 41. Эмоциональное возбуждение как результат определенной мотивационной деятельности теснейшим образом связано с удовлетворением трех основных потребностей человека:
- 42. Удовлетворенная потребность вызывает эмоциональное переживание положительного характера и определяет направление поведенческой деятельности. Положительные эмоции, закрепляясь в
- 43. Эмоции, проявляются при недостатке точных сведений и путей достижения жизненных потребностей. Такое представление о природе эмоции
- 44. По Г. И. Косицкому, оценка величины эмоционального напряжения определяется по формуле: СН = Ц (Ин∙Вн∙Эн— Ис∙Вс∙Эс),
- 45. Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности. Эта стадия имеет тренирующее значение,
- 46. Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности. Эта стадия имеет тренирующее значение,
- 47. Третья стадия (СНШ) — астеническая отрицательная реакция, характеризующаяся истощением ресурсов организма и находящая свое психологическое выражение
- 48. В теории функциональной системы по П. К. Анохину нейрофизиологическая природа эмоций связывается с представлениями о функциональной
- 49. Регуляция вегетативных (висцеральных) функций Осуществляется преимущественно через гипоталамус, куда информация поступает от различных участков лимбической системы
- 50. Формирование эмоций (пережеваний) Наиболее важная функция лимбической системы. В свою очередь, эмоции, являются субъективным компонентом мотиваций,
- 51. Роль лимбической системы в обучении Связана с кругом Пейпеца, где главную роль играют гиппокамп и связанные
- 53. План лекции: - Характеристика вегетативной нервной системы - Общий план строения ВНС - Центры ВНС -
- 54. 1801 - М. Биша – «ВЕГЕТАТИВНЫЕ ПРОЦЕССЫ» 1807- Г. Рейл – «ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА» 1903 -
- 55. Цитата « Мы не являемся хозяевами, а лишь свидетелями частоты сердцебиений, сокращений желудка и кишечника. Их
- 56. Работа ВНС осуществляется рефлекторно (по принципу обратной связи) и независимо (автономно) от сознания, но не от
- 57. Вегетативная нервная система Симпатическая нервная система Парасимпатическая нервная система Метасимпатическая (энтеральная) нервная система ?
- 61. Центральная часть - симпатическое ядро бокового рога серого вещества спинного мозга. Оно тянется от первых грудных
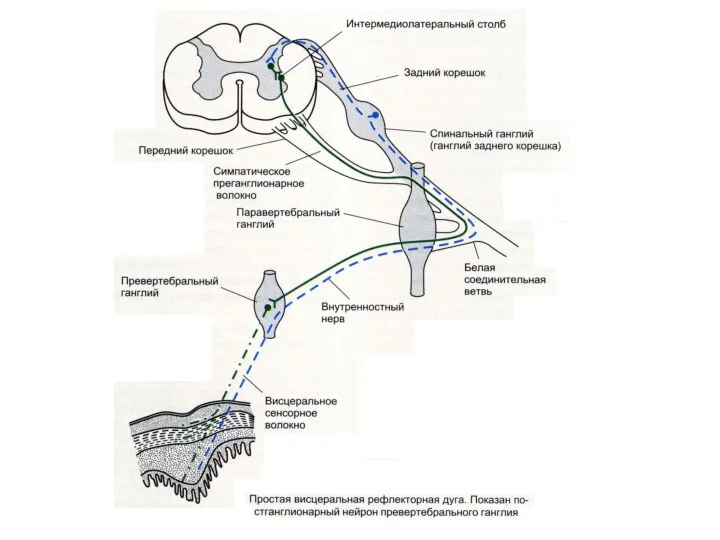
- 62. Схема дуги симпатического рефлекса 1 — спинной мозг, 2 — чувствительный нейрон, 3 — спинальный ганглий
- 63. имеет собственные чувствительные пути: клетки, тела которых локализуются в превертебральных симпатических ганглиях. Один из длинных отростков
- 64. Спино – цилиарный центр (ц. Будге) На уровне последнего шейного (С8) и двух верхних грудных сегментов
- 65. Спино – цилиарный центр (ц. Будге) Раздражение этих волокон вызывает: расширение зрачка – мидраз раскрытие глазной
- 68. в среднем мозгу - ядро глазодвигательного нерва. Преганглионарные волокна к глазным мышцам, железам и другим образованиям
- 69. Периферическая часть парасимпатической НС образована чувствительными и эфферентными нейронами Главным коллектором чувствительных путей парасимпатической нервной системы
- 70. СОМАТИЧЕСКАЯ И ВЕГЕТАТИВНАЯ РЕФЛЕКТОРНЫЕ ДУГИ
- 72. Вегетативный ганглий Преганглионарное волокно Постганглионарные волокна Преганглионарный синапс (АХ) Никотиновый рецептор
- 73. Отличия вегетативной и соматической нервной системы
- 74. МЕДИАТОРНЫЕ МЕХАНИЗМЫ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
- 75. МЕДИАТОРЫ ПЕРИФЕРИЧЕСКОЙ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
- 76. Висцеральные сигналы По сравнению с соматическими характеризуются: низкой скоростью проведения, менее развитой системой пространственной локализации восприятия
- 77. Нервные окончания Аксоны постганглионарных вегетативных нейронов образуют многочисленные варикозные расширения - содержащие синаптические пузырьки. Эти утолщения
- 78. ВЗАИМОДЕЙСТВИЕ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ НЕРВНОЙ РЕГУЛЯЦИИ АНТАГОНИЗМ СИНЕРГИЗМ ОТСУТСТВИЕ ВЗАИМОДЕЙСТВИЯ
- 79. Симпатические и парасимпатические эффекты
- 80. Моносимпатическая регуляция
- 81. Монопарасимпатическая регуляция
- 82. 1. Иннервирует только внутренние органы, наделенные собственной моторной активностью. 2. Получает синаптические входы от СНС и
- 84. Функциональный модуль метасимпатической нервной системы 1 — чувствительный нейрон, 2 — интернейрон, 3 — эфферентный нейрон,
- 85. Модель организации метасимпатической НС
- 86. Аденозинтрифосфат (АТФ) АТФ локализуется в пресинаптических терминалях эффекторных нейронов метасимпатической нервной системы. Преобладающим действием АТФ на
- 87. Трансдукторы Клетки, возбуждающиеся обычным путем, а отвечающие эндокринным способом, называют трансдукторами. Аксоны этих клеток не образуют
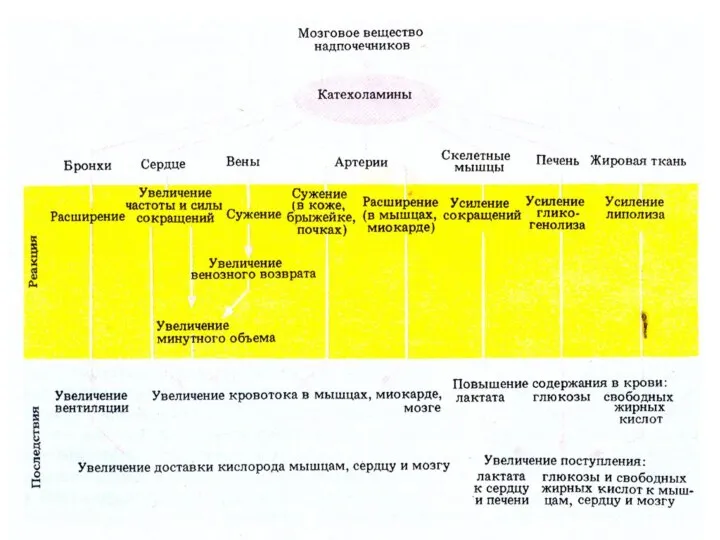
- 88. Эффекты ВНС
- 92. Только ацетилхолин, норадреналин и серотонин удовлетворяют всем критериям, предъявляемым к нейромедиаторам в разных отделах ВНС. НЕЙРОМЕДИАТОРЫ
- 93. НЕЙРОМЕДИАТОРЫ Холинергические нейроны Все преганглионарные нейроны как в симпатическом, так и в парасимпатическом отделах холинергические. Почти
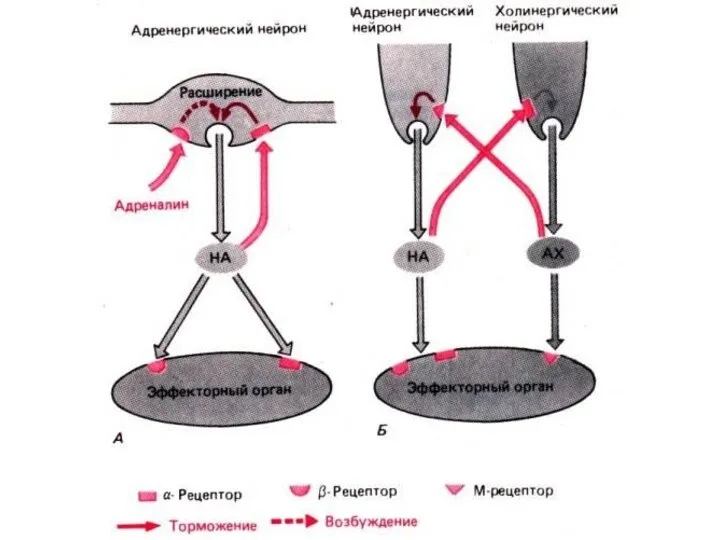
- 94. АДРЕНОРЕЦЕПТОРЫ
- 95. ХОЛИНОРЕЦЕПТОРЫ
- 96. Вещества действующие на адренергические эффекторы Симпатомиметики: естественные агонисты адренорецепторов — норадреналин и адреналин, вещества стимулирующие определенный
- 97. Вещества действующие на холинергические эффекторы Парасимпатомиметики – агонисты холинорецепторов (пилокарпин и метахолин) (ацетилхолин после введения в
- 98. Вещества действующие на постганглионарные нейроны ВНС Возбуждающие Ацетилхолин секретируется в преганглионарных нейронах и стимулирует постганглионарные. Никотин
- 99. Серотонин Почти 90% образуется в хромаффинных клетках слизистой оболочки пищеварительного тракта. Во время пищеварения часть вещества
- 100. Эффекты серотонина На сосуды: Вазоконстрикция проявляется на денервированных сосудах. В скелетных мышцах и кожных покровах -вазодилатация,
- 101. Серотонинергические рецепторы
- 102. Основные трансмиттеры в химической передаче возбуждения в периферической нервной системе млекопитающих АХ — ацетилхолин, АД —
- 103. Типы. метасимпатических нейронов млекопитающих А —мотонейроны, идущие к гладкой мышце; Б — интернейроны; В — парасимпатические
- 107. ВИДЫ ВЕГЕТАТИВНЫХ РЕФЛЕКСОВ Висцеро-висцеральный рефлекс - аксон-рефлекс Висцеро-соматический рефлекс Висцеро-сенсорный рефлекс Висцеро-дермальный рефлекс Соматовисцеральный рефлекс Дермо-висцеральный
- 108. Висцеро-висцеральный рефлекс Возбуждение возникает и заканчивается во внутренних органах. Эффектор способен отвечать либо усилением, либо торможением
- 109. Аксон - рефлекс Это местная ответная реакция ткани на раздражитель без участия ЦНС возбуждение интероцептора является
- 110. Висцеро-соматический рефлекс В дополнение к висцеральным вызывает также соматические ответы в виде, например, усиления (сокращения) или
- 111. Висцеросенсорный рефлекс В ответ на раздражение автономных чувствительных волокон возникают реакции не только во внутренних органах,
- 112. РЕАКЦИИ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ Симпатическая нервная система в зависимости от характера и силы раздражений отвечает либо
- 113. РЕАКЦИИ ПАРАСИМПАТИЧЕСКОЙ СИСТЕМЫ Парасимпатическая система осуществляет локальный и более специфический контроль функций эффекторных органов. Цель парасимпатической
- 114. Иерархия в управлении деятельностью внутренних органов
- 115. Спинальные центры
- 116. Стволовые центры
- 117. Высший вегетативный орган – гипоталамус
- 118. Ядерные зоны гипоталамуса на схеме сагиттального разреза через третий желудочек. 1 –преоптическое ядро (преоптическая область); 2–паравентрикулярное
- 121. Двусторонние связи гипоталамуса с: Таламусом Лимбической системой КБП Центральным серым веществом среднего мозга Соматическими ядрами ствола
- 122. Нейроны гипоталамуса Обладают рецепторной функцией и способны улавливать изменения химического состава крови и цериброспинальной жидкости, что
- 123. Гипоталамус: чувствительное звено Чувствительная информация от внутренних органов поступает в гипоталамус по восходящим спинно-бульбарным путям. Одни
- 124. Гипоталамус: эффекторное звено Гипоталамус играет ведущую роль в поддержании гомеостаза. Стимуляция задних ядер гипоталамуса сопровождается эффектами,
- 125. Гипоталамус: поведенческая функция Гипоталамус участвует в формировании поведенческих реакций организма, необходимых для сохранения гомеостаза. Стимуляция его
- 127. Схема функциональной организации поведенческих программ, заложенных в гипоталамусе
- 128. Вегетативные реакции, сопровождающие пищевое и оборонительное поведение при электрическом раздражении гипоталамуса у кошки
- 129. Слева: нисходящие пути от ствола мозга и гипоталамуса, конвергирующие на преганглионарных нейронах промежуточной зоны пояснично–грудного отдела
- 130. Дуга рефлекса мочеиспускания у кошки с интактным головным мозгом (рефлекторная дуга 1) и у хронической спинальной
- 131. Иннервация мочевого пузыря
- 136. Афферентные и эфферентные пути спинального рефлекса дефекации. Вставочные спинномозговые нейроны между афферентными и эфферентными волокнами не
- 137. Иннервация мужских половых органов. Вставочные спинномозговые нейроны между афферентными и эфферентными волокнами не показаны
- 138. Таламус - ограничивает поступление афферентной импульсации в кору большого мозга от внутренних органов, обеспечивая ее большую
- 139. Ретикулярная формация Ее нейроны формируют центры дыхания и кровообращения и реализуют свое влияние через активацию симпатической
- 140. Голубое пятно Его норадренергические нейроны иннервируют артериолы и капилляры коры больших полушарий и рассматриваются как центральный
- 141. Лимбическая система Лимбическая система обеспечивает взаимодействие экстероцептивных (обонятельных, слуховых и др.) и интероцептивных воздействий. Занимая в
- 142. Лимбическая система Участвует в приспособительной регуляции деятельности систем: Сердечнососудистой Дыхательной Пищеварительной и др. систем Что проявляется
- 144. Мозжечок Мозжечок благодаря наличию двойного (активирующего и тормозного) механизма действия способен оказывать стабилизирующее влияние на функции
- 145. Мозжечок Реализует свое влияние через симпатическую нервную систему и эндокринные железы, вызывая: Сужение кровеносных сосудов Расширение
- 146. Карта представительства афферентных систем внутренних органов в коре 1—6, 13—16, 19 — блуждающий нерв (1—4, 19
- 147. Кора большого мозга. КБП — высший интегративный центр регуляции всех функций организма, в том числе вегетативных.
- 148. Стимуляция двигательной зоны коры вызывает такие же изменения деятельности сердечно-сосудистой системы (увеличение минутного объема сердца, усиление
- 149. Тонус вегетативных центров Многие преганглионарные и ганглионарные вегетативные нейроны обладают постоянной активностью, называемой тонусом. Ее происхождение:
- 150. Значение тонуса вегетативных центров. Тонус вегетативных центров играет важную роль в приспособительном регулировании функций внутренних органов.
- 151. Вегетативный портрет. Симпатикотоники и парасимпатикотоники. Деление лиц на симпатикотоников и парасимпатикотоников на основании интенсивности деятельности внутренних
- 152. Трофическое действие нервной системы Идею о трофическом действии НС сформулировал И. П. Павлов. В опытах на
- 153. Повышение работоспособности утомленной изолированной икроножной мышцы лягушки (1) при раздражении симпатических волокон (2) (феномен Орбели —
- 154. Такое действие симпатической нервной системы Л.А. Орбели назвал адаптационно - трофическим. Трофическое действие на ткань присуще
- 155. Вегетативная нервная система детей
- 156. Вегетативная нервная система детей В онтогенезе ВНС претерпевает существенные структурные и функциональные изменения; меняется и доля
- 157. ВНС у детей Медиатором преганглионарных симпатических нейронов является адреноподобное вещество (у взрослых — ацетилхолин), т.е. дифференцировка
- 158. ВНС у детей В процессе онтогенеза число холинергических синапсов в структурах ВНС постепенно увеличивается. Специализация медиаторов
- 159. ВНС у детей Автоматизм клеток симпатических ганглиев и низкий мембранный потенциал симпатических нейронов новорожденных объясняются функциональными
- 160. ВНС у детей Характерными особенностями ВНС в первые годы жизни ребенка являются также: повышенная возбудимость непостоянство
- 161. ВНС у детей Различная скорость созревания симпатической и парасимпатической нервной системы в ганглиях интрамуральной системы конечные
- 162. ВНС у детей Парасимпатический отдел начинает включаться в рефлекторные реакции сердца с 3-го месяца жизни. Однако,
- 163. ВНС у детей Максимальное замедление пульса от 150 до 30 ударов было отмечено у них при
- 164. ВНС у детей В регуляцию ЖКТ также сначала включается парасимпатическая нервная система. Симпатическая регуляция начинает осуществлятся
- 165. ВНС у детей Механизм формирования тонуса вегетативных центров в онтогенезе. В формировании тонуса блуждаюшего нерва важную
- 166. ВНС у детей Для оценки степени выраженности тонуса блуждающего нерва в детском возрасте используют глазосердечный рефлекс
- 167. ВНС у детей Важное значение в становлении тонуса блуждающего нерва играет импульсация от баро- и хеморецепторов
- 168. Иерархия в управлении деятельностью внутренних органов
- 169. Физиология эндокринной системы
- 170. Классические эндокринные железы (голубовато-зеленые) и некоторые органы диффузной эндокринной системы (фиолетовые точки). С – клетки ЩЖ
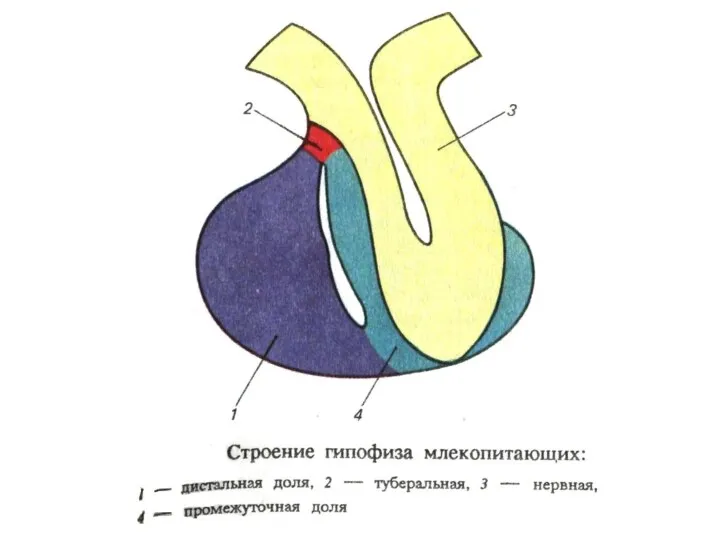
- 171. Эндокринная система 1. Эндокринные железы ГИПОФИЗ (аденогипофиз и нейрогипофиз) НАДПОЧЕЧНИКИ (кора и мозговое в-во) ЩИТОВИДНАЯ ЖЕЛЕЗА
- 172. Железы внутренней секреции и их гормоны
- 175. Химическая природа и пути действия гормонов Простые и сложные белки, - через мембранные пептиды - гормоны
- 177. Виды действия гормонов Метаболическое Морфогенетическое Кинетическое Корригирующее
- 178. Нейроэндокринные системы регуляции гормональной системы по принципу механизма обратной отрицательной связи, тормозящих секрецию гипоталамических и гипофизарных
- 179. Динамики секреции в гормональных системах происходит в соответствии с характерными ритмами (от минут до периодов жизни),
- 181. Нейроны гипоталамуса Обладают рецепторной функцией и способны улавливать изменения химического состава крови и цереброспинальной жидкости, что
- 182. Двусторонние связи гипоталамуса с: Таламусом Лимбической системой КБП Центральным серым веществом среднего мозга Соматическими ядрами ствола
- 183. Гипоталамус Вырабатывает собственные гормоны Эффекторные: окситоцин и вазопрессин (супраоптическое и паравентрикулярное) Релизинг-гормоны: либерины и статины
- 186. Гормоны гипофизотропной части гипоталамуса выделяются в области срединного возвышения и через воротную систему гипофиза транспортируются в
- 187. Топография циркумвентрикулярных органов (ЦВО), которые находятся вне гематоэнцефалического барьера и гормоны могут действовать здесь как гуморальные
- 188. Гуморальные связи ЦНС с периферией организма. Слева: варианты выделения гормонов 1- гипофизтропные гормоны определяют высвобождение гормонов
- 190. Система гормональной регуляции организма позвоночных — гипоталамо—гипофизарная система; — эндокринные железы—мишени гормонов аденогипофиза. Висперотропные нейрогормоны: АДГ
- 191. Задняя доля гипофиза состоит из терминалей аксонов крупных клеток супраоптического и паравентрикулярного ядер (СЯ и ПЯ
- 193. Нейрогормональный контроль висцеральных органов 1 — пептидергические нейросекреторные клетки (Нск), 2 — пути к различного типа
- 196. Основные принципы регуляции секреции аденогипофизарных гормонов (нижние прямоугольники) со стороны гипоталамуса (верхний прямоугольник).
- 197. Прямые и обратные связи в системе нейроэндокринной регуляции 1 — медленно развивающееся и продолжительное ингибирование секреции
- 198. Регуляция активности эндокринных желез ЦНС при участии гипоталамуса и гипофиза ТЛ — тиреолиберин; СЛ — соматолиберин;
- 200. Нейро-гуморальная рефлекторная регуляция Выделения молока при кормлении грудью (рефлекс Фергесона). Может вызываться условнорефлекторно (крик младенца). Лежит
- 201. Пролактин, его функции и регуляция. Прямой контроль секреции пролактина с помощью механизма отрицательной обратной связи не
- 202. Гормон роста СТГ, его функции и регуляция. СТГ стимулирует в печени секрецию ИФР-1 (соматомедин С) и
- 203. Множественные функции соматостатина. Соматостатин присутствует во многих тканях. Он действует: Через кровь как гормон Паракринным путем
- 204. Множественное действие кортикотропин-РГ (КРГ) КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует также в управлении центрами
- 205. Функции антидиуретического гормона (вазопрессина). АДГ действует на ЦНС как нейротрансмиттер и нейромодулятор, на ПОМК-клетки аденогипофиза –
- 206. Взаимодействие гипоталамуса и гипофиза
- 207. Гормоны гипофиза и их функции
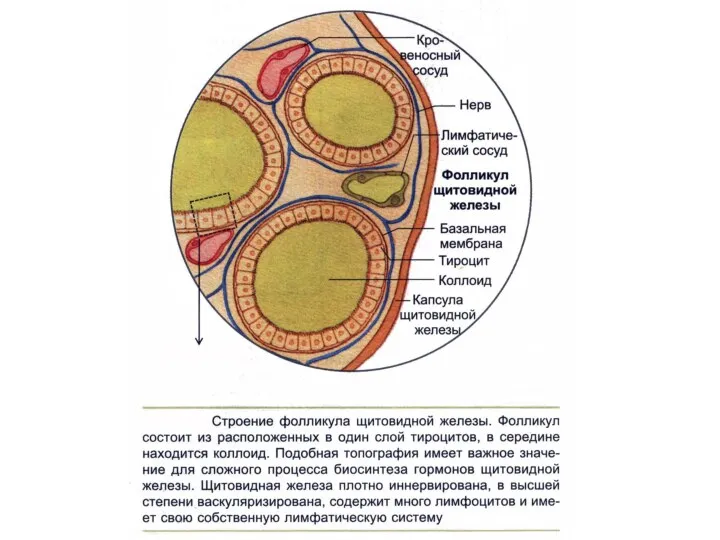
- 208. ЩИТОВИДНАЯ ЖЕЛЕЗА
- 209. Фолликулы щитовидной железы при неактивном (А) и активном (Б) состояниях железы А — эпителий плоский, фолликулы
- 211. Молекула тиреоглобулина и строение основных гормонов щитовидной железы. Представленные три- или тетрайодтиронины, а также моно- и
- 212. Уровень гормонов щитовидной железы и связывающих белков в плазме и функциональные тесты. А) приведены характерные для
- 213. Связи щитовидной железы у млекопитающих
- 215. Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иоде покрывается за счет всасывания из в
- 218. Транспорт тироидного гормона в сыворотке крови (тироксин связывающий глобулин, транстиретин, альбумин)
- 219. Гормоны щитовидной (1) и паращитовидных (2) желез и их функции
- 220. Регуляция содержания кальция в крови кальцитонином и паратгормоном
- 222. НАДПОЧЕЧНИКИ
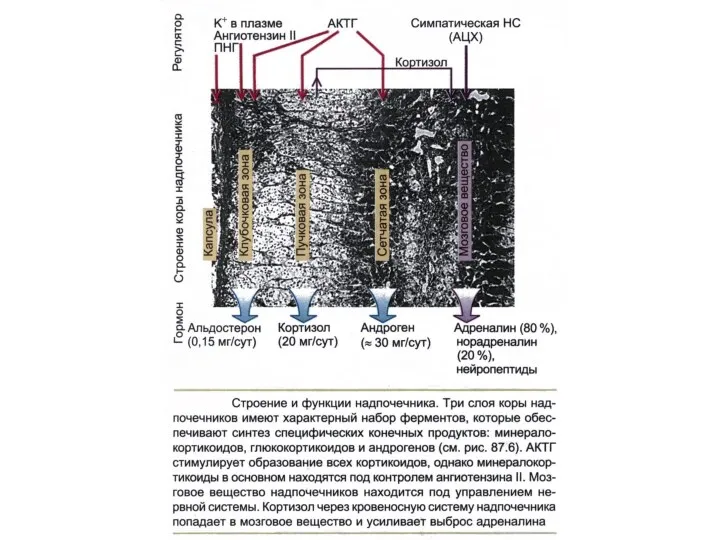
- 223. Схема зон надпочечника и вырабатываемые ими гормоны 1 — пучковая зона, 2 — сетчатая зона, 3
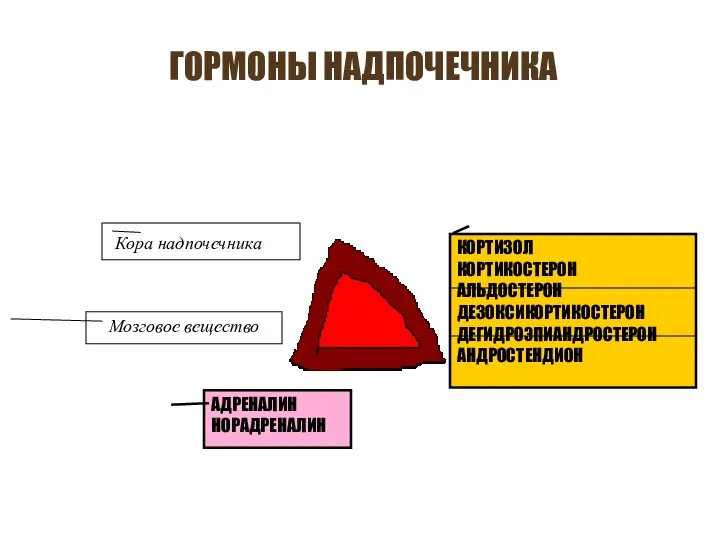
- 225. ГОРМОНЫ НАДПОЧЕЧНИКА Кора надпочечника Мозговое вещество КОРТИЗОЛ КОРТИКОСТЕРОН АЛЬДОСТЕРОН ДЕЗОКСИКОРТИКОСТЕРОН ДЕГИДРОЭПИАНДРОСТЕРОН АНДРОСТЕНДИОН АДРЕНАЛИН НОРАДРЕНАЛИН
- 226. Гормоны надпочечников
- 228. Система гипоталамус гипофиз–кора надпочечников.
- 229. Паракринное действие кортизола на синтез катехоламинов в мозговом веществе надпочечников. Важный этап синтеза от тирозина к
- 230. Регуляция оси «гипоталамус – гипофиз – кора надпочечников», обратная связь с участием кортизола, а также взаимодействия
- 231. Множественное действие КРГ. КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует в управлении центрами ВНС. Действуя
- 232. МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ СЛИЗИСТОЙ ОБОЛОЧКИ
- 233. СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ Реакция тревоги СТАДИЯ РЕЗИСТЕНТНОСТИ СТАДИЯ ИСТОЩЕНИЯ ИСХОДНЫЙ СРЕДНИЙ УРОВЕНЬ УСТОЙЧИВОСТИ
- 234. ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
- 235. Гормоны поджелудочной железы и их функции: α- альфа-клетки, продуцирующие глюкогон, β – бетта-клетки, продуцирующие инсулин, D
- 236. Регуляция активности островковых клеток поджелудочной железы
- 237. Спасибо за ваше внимание
- 245. Скачать презентацию
Структура функциональной системы поведения по П.К.Анохину
МОТИ ВАЦИЯ
МОТИВАЦИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
Структура функциональной системы поведения по П.К.Анохину
МОТИ ВАЦИЯ
МОТИВАЦИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
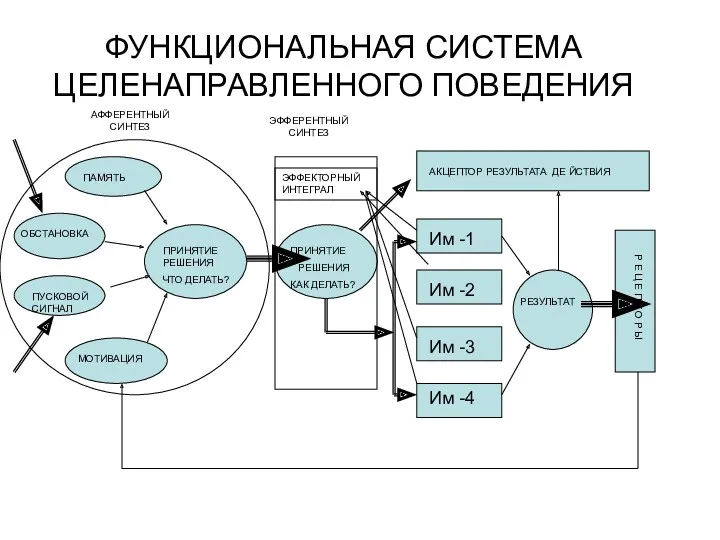
ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ РЕШЕНИЯ
ЧТО ДЕЛАТЬ?
ЭФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ
ФУНКЦИОНАЛЬНАЯ СИСТЕМА ЦЕЛЕНАПРАВЛЕННОГО ПОВЕДЕНИЯ
АФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ РЕШЕНИЯ
ЧТО ДЕЛАТЬ?
ЭФФЕРЕНТНЫЙ СИНТЕЗ
ПРИНЯТИЕ
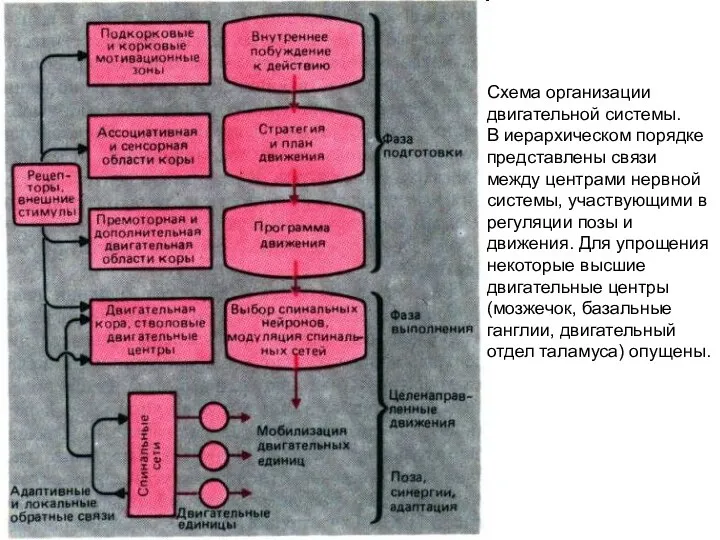
Схема организации двигательной системы.
В иерархическом порядке представлены связи между центрами
Схема организации двигательной системы.
В иерархическом порядке представлены связи между центрами
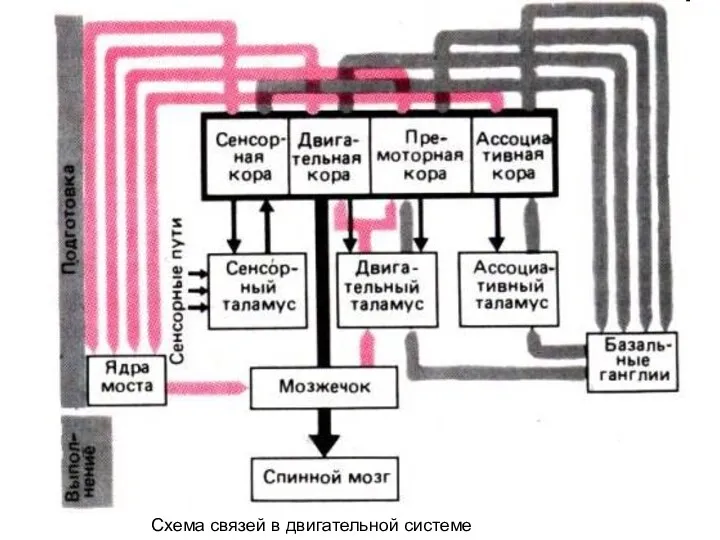
Схема связей в двигательной системе
Схема связей в двигательной системе
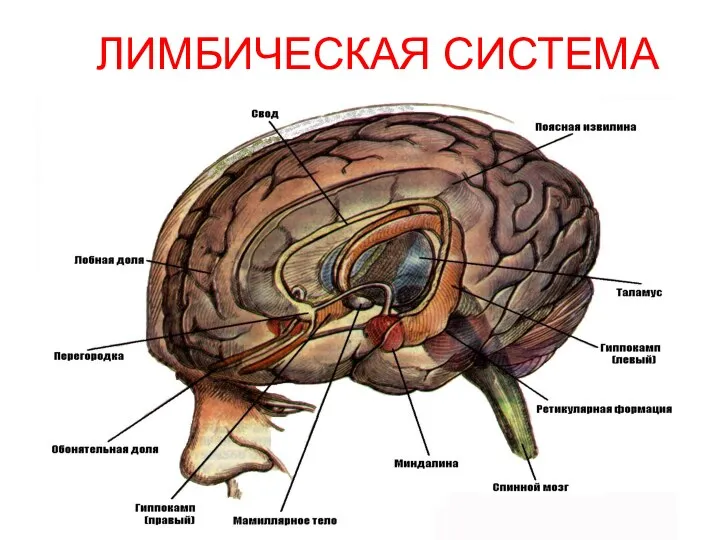
ЛИМБИЧЕСКАЯ СИСТЕМА
ЛИМБИЧЕСКАЯ СИСТЕМА

Лимбическая система
Функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее
Лимбическая система
Функциональное объединение различных структур конечного, промежуточного и среднего мозга, обеспечивающее

Афферентные входы
От различных областей головного мозга
Через гипоталамус от ретикулярной формации
Афферентные входы
От различных областей головного мозга
Через гипоталамус от ретикулярной формации
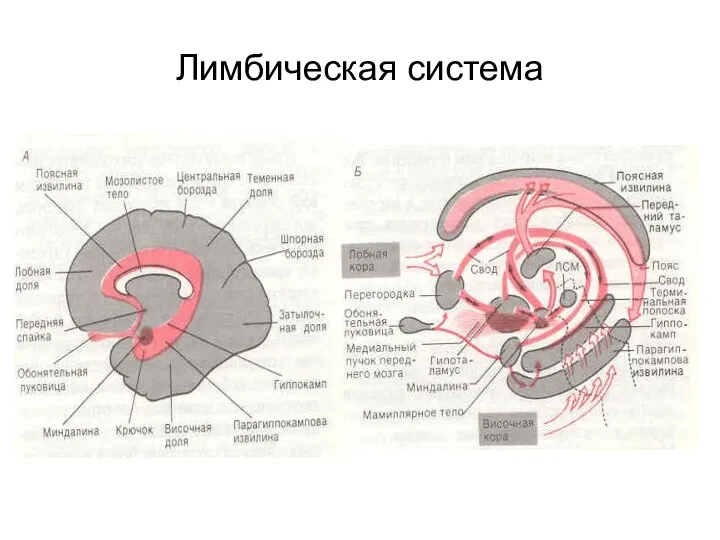
Лимбическая система
Лимбическая система

ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ
1. Организация вегетативно-соматических компонентов эмоций
2. Организация кратковременной и долговременной
ОСНОВНЫЕ ФУНКЦИИ ЛИМБИКИ
1. Организация вегетативно-соматических компонентов эмоций
2. Организация кратковременной и долговременной

Кольцевые нейрональные связи
Дают возможность длительной циркуляции (реверберации) возбуждения, что создает условия
Кольцевые нейрональные связи
Дают возможность длительной циркуляции (реверберации) возбуждения, что создает условия
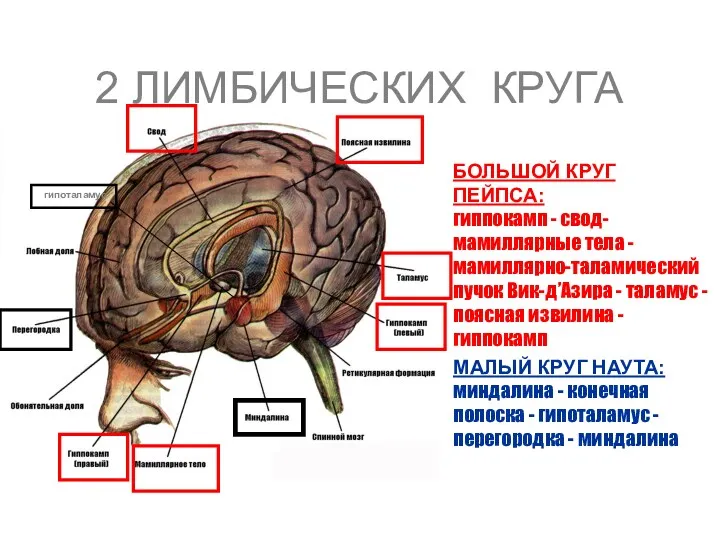
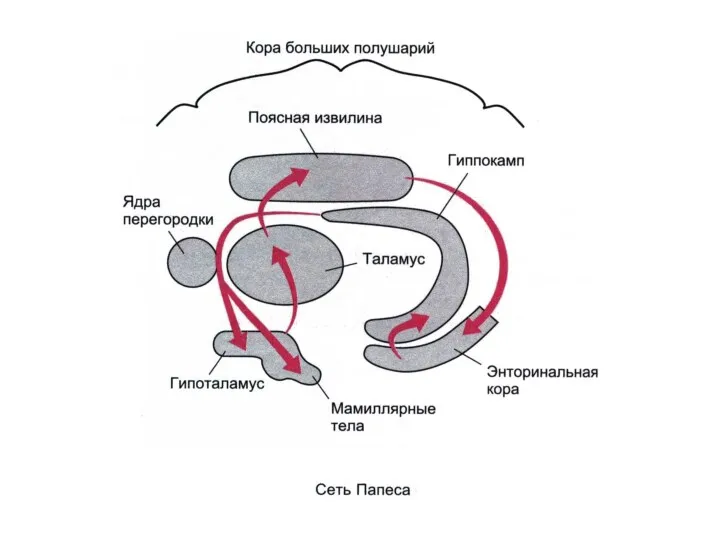
2 ЛИМБИЧЕСКИХ КРУГА
БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела -
2 ЛИМБИЧЕСКИХ КРУГА
БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела -

Функциональная структура лимбики по МакЛину (1970)
1. Нижний отдел - миндалина и
Функциональная структура лимбики по МакЛину (1970)
1. Нижний отдел - миндалина и

Функции
лимбической системы
После обработки информации от внешней среды и внутренних органах
Функции
лимбической системы
После обработки информации от внешней среды и внутренних органах

Физиология гиппокампа
Физиология гиппокампа
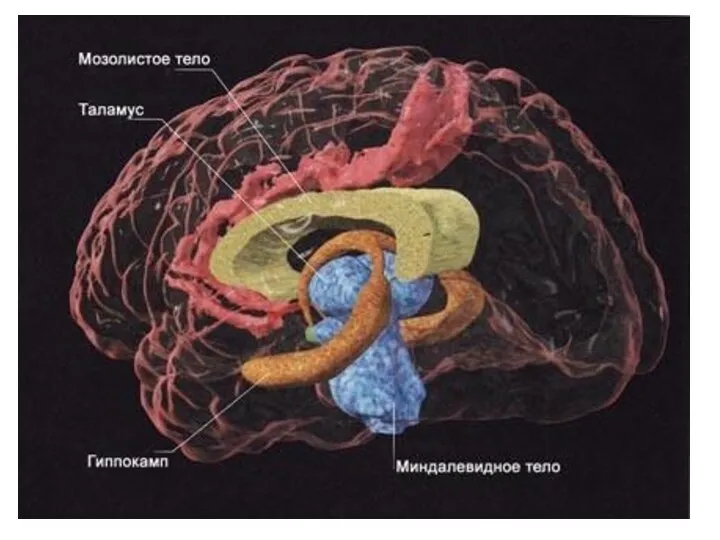

Физиология гиппокампа
Гиппокамп (hippocampus) является основной структурой лимбической системы. Морфологически гиппокамп
Физиология гиппокампа
Гиппокамп (hippocampus) является основной структурой лимбической системы. Морфологически гиппокамп

Физиология гиппокампа
Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа,
Физиология гиппокампа
Значение тета-ритма заключается в том, что он отражает реакцию гиппокампа,

Физиология миндалевидного тела
Физиология миндалевидного тела
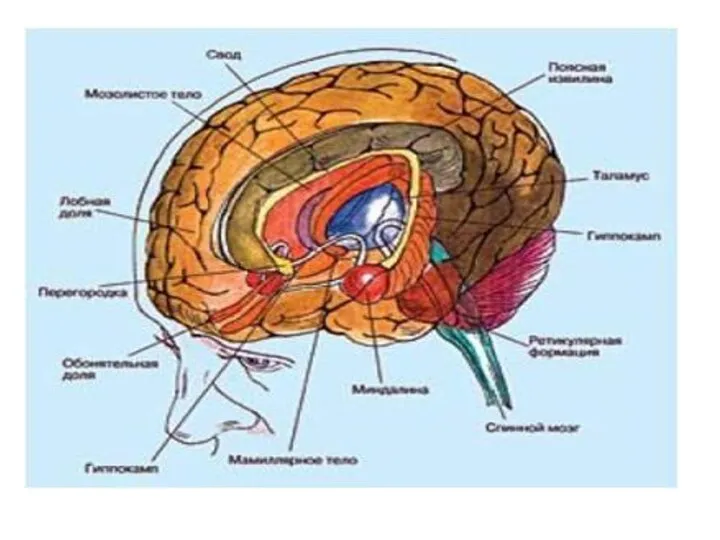

Физиология миндалевидного тела
Миндалевидное тело, миндалина — подкорковая структура лимбической системы,
Физиология миндалевидного тела
Миндалевидное тело, миндалина — подкорковая структура лимбической системы,

Физиология миндалевидного тела
При искусственной активации миндалины появляются реакции:
принюхивания,
облизывания,
жевания, глотания,
саливации,
Физиология миндалевидного тела
При искусственной активации миндалины появляются реакции:
принюхивания,
облизывания,
жевания, глотания,
саливации,
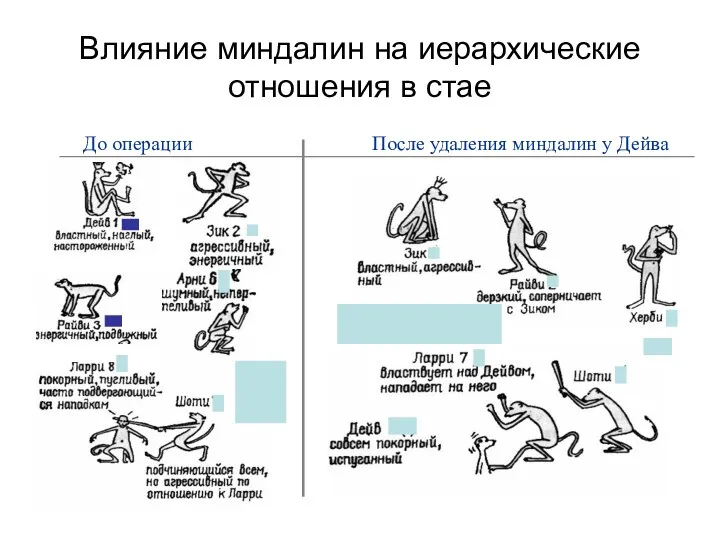
Влияние миндалин на иерархические отношения в стае
До операции
После удаления миндалин у
Влияние миндалин на иерархические отношения в стае
До операции
После удаления миндалин у
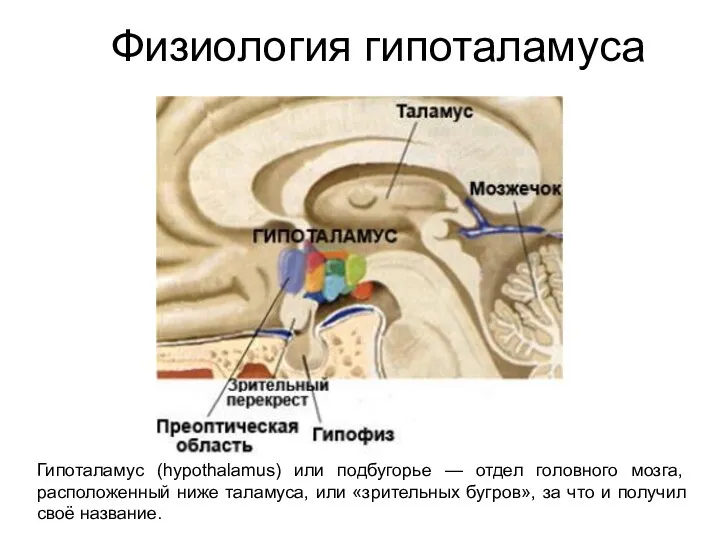
Физиология гипоталамуса
Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга, расположенный
Физиология гипоталамуса
Гипоталамус (hypothalamus) или подбугорье — отдел головного мозга, расположенный
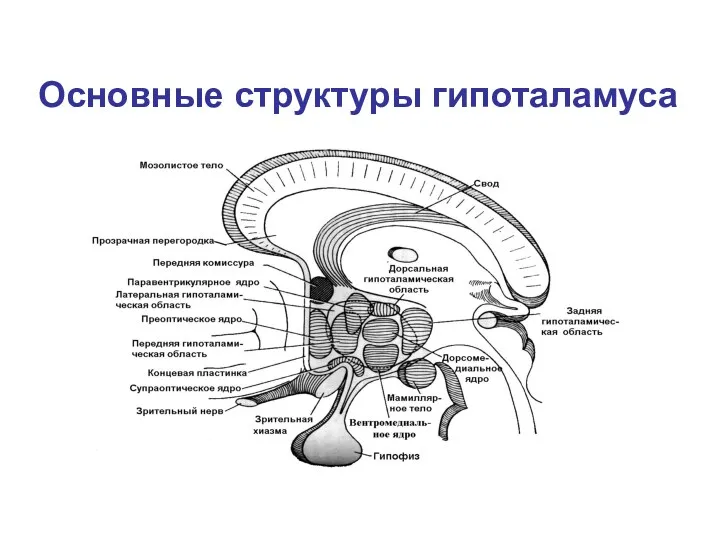
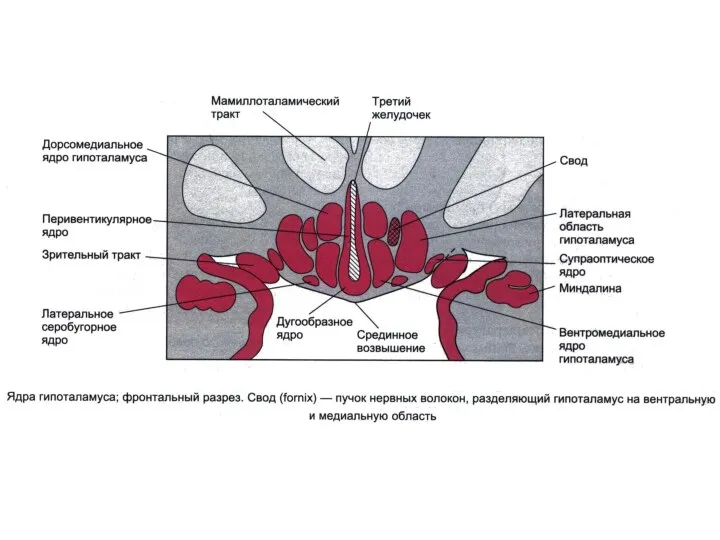
Основные структуры гипоталамуса
Основные структуры гипоталамуса

ФУНКЦИИ ГИПОТАЛАМУСА
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция
ФУНКЦИИ ГИПОТАЛАМУСА
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция

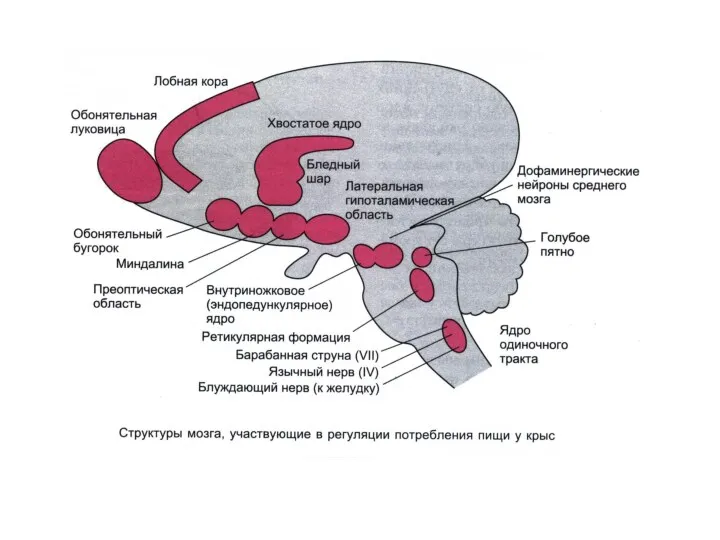
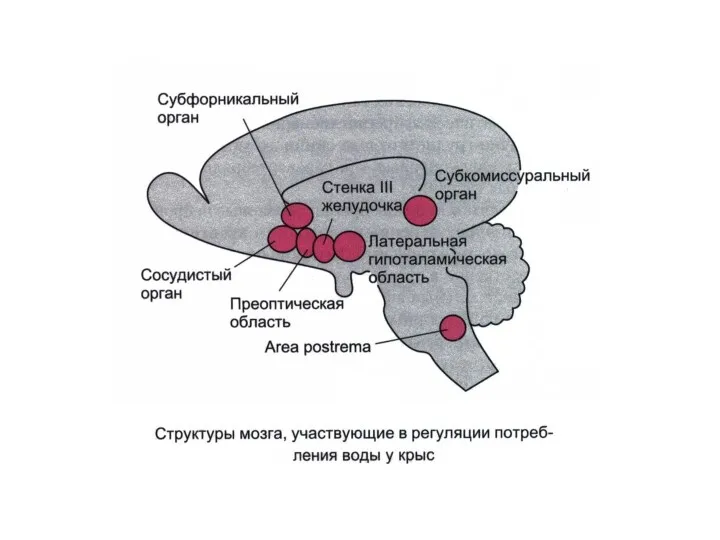
Поведенческие функции гипоталамуса
Эффекты стимуляции гипоталамуса
Латеральный гипоталамус: жажда, аппетит, увеличение активности
Поведенческие функции гипоталамуса
Эффекты стимуляции гипоталамуса
Латеральный гипоталамус: жажда, аппетит, увеличение активности

Центры поощрения
В различные области мозга обезьяны вводили электроды, и обезьяна,
Центры поощрения
В различные области мозга обезьяны вводили электроды, и обезьяна,

Центры поощрения и наказания
Личные оценки могут характеризовать ощущения как приятные
Центры поощрения и наказания
Личные оценки могут характеризовать ощущения как приятные

Главные (первичные) центры поощрения располагаются по ходу медиального пучка переднего мозга
Главные (первичные) центры поощрения располагаются по ходу медиального пучка переднего мозга

Центры наказания
и реакции избегания расположены в центральном сером веществе, окружающем
Центры наказания
и реакции избегания расположены в центральном сером веществе, окружающем

Ярость возникает при активации центров наказания. В этом состоянии даже легкая
Ярость возникает при активации центров наказания. В этом состоянии даже легкая

Роль поощрения и наказания в поведении, обучении и памяти
Всё или
Роль поощрения и наказания в поведении, обучении и памяти
Всё или

Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших
Привыкание. Новые сенсорные стимулы почти всегда возбуждают значительные области коры больших

Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает
Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает

Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль
Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль
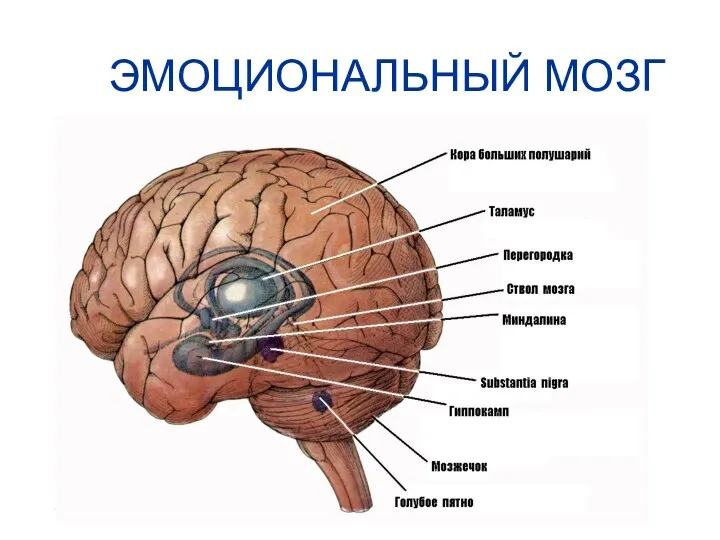
ЭМОЦИОНАЛЬНЫЙ МОЗГ
ЭМОЦИОНАЛЬНЫЙ МОЗГ

ЦИТАТА
«Является ли эмоция продуктом волшебства или физиологическим процессом, который зависит от
ЦИТАТА
«Является ли эмоция продуктом волшебства или физиологическим процессом, который зависит от

Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,
Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,

Эмоция — специфическое состояние психической сферы, обусловленная рефлекторной реакцией организма на
Эмоция — специфическое состояние психической сферы, обусловленная рефлекторной реакцией организма на

Эмоциональное возбуждение как результат определенной мотивационной деятельности теснейшим образом связано с
Эмоциональное возбуждение как результат определенной мотивационной деятельности теснейшим образом связано с

Удовлетворенная потребность вызывает эмоциональное переживание положительного характера и определяет направление поведенческой
Удовлетворенная потребность вызывает эмоциональное переживание положительного характера и определяет направление поведенческой

Эмоции, проявляются при недостатке точных сведений и путей достижения жизненных потребностей.
Эмоции, проявляются при недостатке точных сведений и путей достижения жизненных потребностей.

По Г. И. Косицкому, оценка величины эмоционального напряжения определяется по формуле:
СН
По Г. И. Косицкому, оценка величины эмоционального напряжения определяется по формуле:
СН

Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.
Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.

Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.
Первая стадия напряжения (CHI) — состояние внимания, мобилизация активности, повышение работоспособности.

Третья стадия (СНШ) — астеническая отрицательная реакция, характеризующаяся истощением ресурсов организма
Третья стадия (СНШ) — астеническая отрицательная реакция, характеризующаяся истощением ресурсов организма

В теории функциональной системы по П. К. Анохину нейрофизиологическая природа эмоций
В теории функциональной системы по П. К. Анохину нейрофизиологическая природа эмоций

Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает
Регуляция вегетативных (висцеральных) функций
Осуществляется преимущественно через гипоталамус, куда информация поступает

Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,
Формирование эмоций (пережеваний)
Наиболее важная функция лимбической системы. В свою очередь, эмоции,

Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль
Роль лимбической системы в обучении
Связана с кругом Пейпеца, где главную роль


План лекции:
- Характеристика вегетативной нервной системы
- Общий план строения ВНС
- Центры
План лекции:
- Характеристика вегетативной нервной системы
- Общий план строения ВНС
- Центры

1801 - М. Биша – «ВЕГЕТАТИВНЫЕ ПРОЦЕССЫ»
1807- Г. Рейл – «ВЕГЕТАТИВНАЯ
1801 - М. Биша – «ВЕГЕТАТИВНЫЕ ПРОЦЕССЫ»
1807- Г. Рейл – «ВЕГЕТАТИВНАЯ

Цитата
« Мы не являемся хозяевами, а лишь свидетелями частоты сердцебиений, сокращений
Цитата
« Мы не являемся хозяевами, а лишь свидетелями частоты сердцебиений, сокращений

Работа ВНС осуществляется
рефлекторно (по принципу обратной связи) и независимо
Работа ВНС осуществляется
рефлекторно (по принципу обратной связи) и независимо

Вегетативная нервная система
Симпатическая нервная система
Парасимпатическая нервная система
Метасимпатическая (энтеральная) нервная система ?
Вегетативная нервная система
Симпатическая нервная система
Парасимпатическая нервная система
Метасимпатическая (энтеральная) нервная система ?
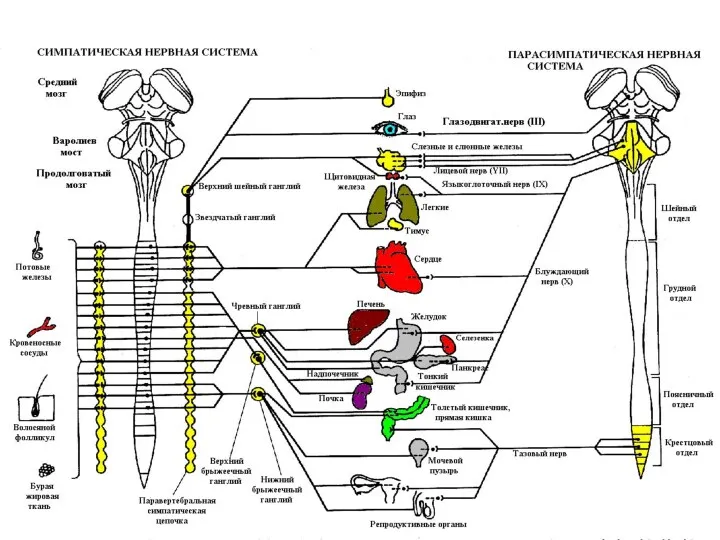
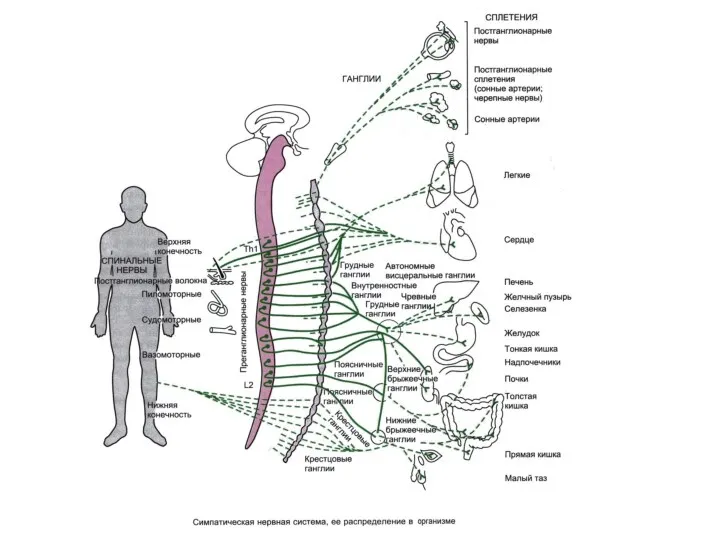
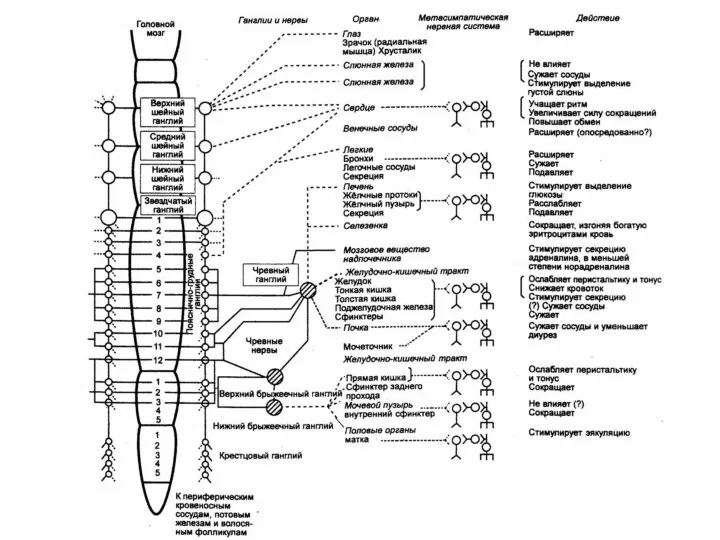

Центральная часть - симпатическое ядро бокового рога
серого вещества спинного мозга.
Центральная часть - симпатическое ядро бокового рога
серого вещества спинного мозга.
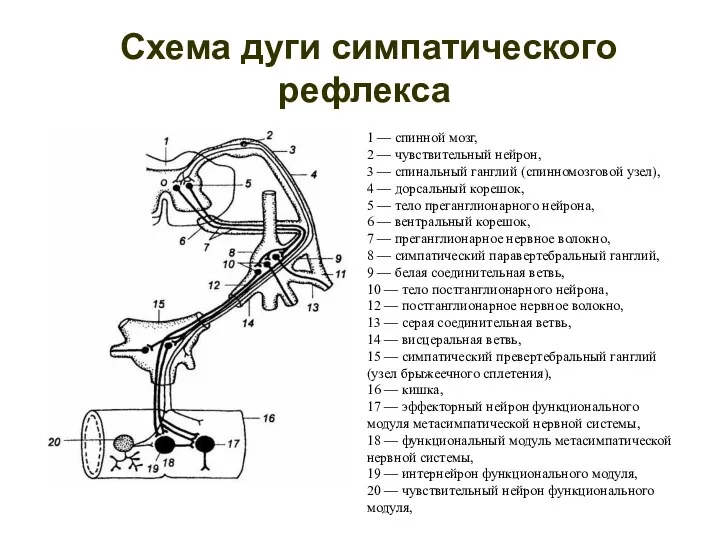
Схема дуги симпатического рефлекса
1 — спинной мозг,
2 — чувствительный
Схема дуги симпатического рефлекса
1 — спинной мозг,
2 — чувствительный

имеет собственные чувствительные пути:
клетки, тела которых локализуются в превертебральных симпатических
имеет собственные чувствительные пути:
клетки, тела которых локализуются в превертебральных симпатических

Спино – цилиарный центр (ц. Будге)
На уровне последнего шейного (С8) и
Спино – цилиарный центр (ц. Будге)
На уровне последнего шейного (С8) и

Спино – цилиарный центр (ц. Будге)
Раздражение этих волокон вызывает:
расширение зрачка
Спино – цилиарный центр (ц. Будге)
Раздражение этих волокон вызывает:
расширение зрачка
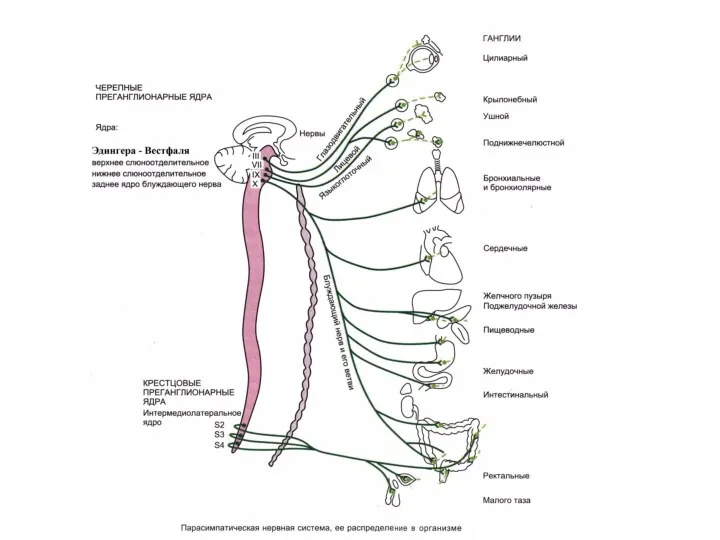
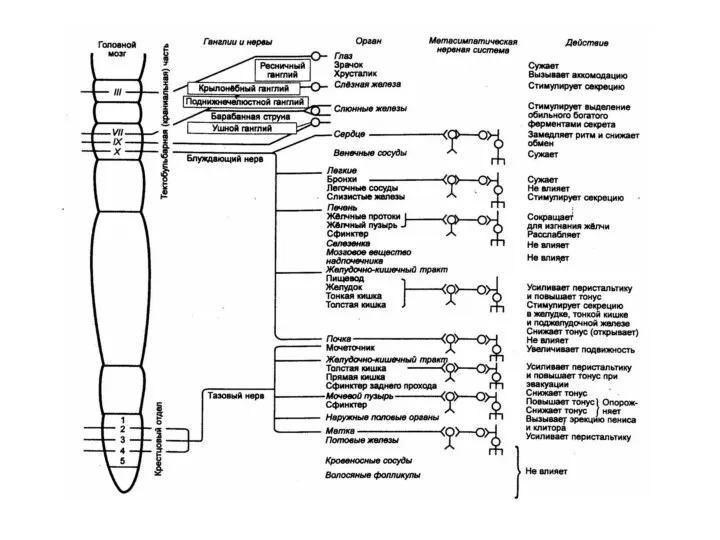

в среднем мозгу - ядро глазодвигательного нерва. Преганглионарные волокна к глазным
в среднем мозгу - ядро глазодвигательного нерва. Преганглионарные волокна к глазным

Периферическая часть парасимпатической НС
образована чувствительными и эфферентными нейронами
Главным коллектором чувствительных путей
Периферическая часть парасимпатической НС
образована чувствительными и эфферентными нейронами
Главным коллектором чувствительных путей
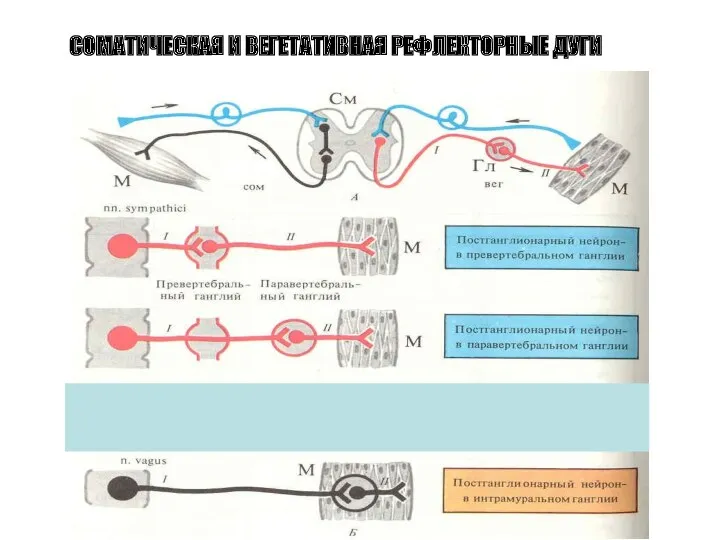
СОМАТИЧЕСКАЯ И ВЕГЕТАТИВНАЯ РЕФЛЕКТОРНЫЕ ДУГИ
СОМАТИЧЕСКАЯ И ВЕГЕТАТИВНАЯ РЕФЛЕКТОРНЫЕ ДУГИ
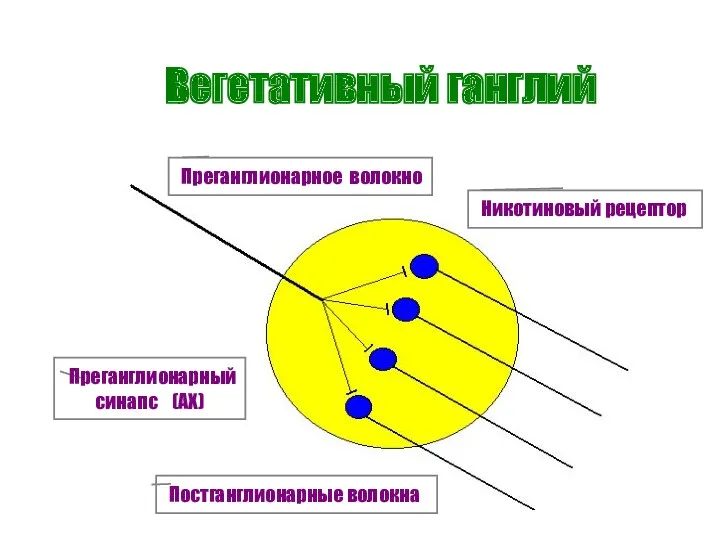
Вегетативный ганглий
Преганглионарное волокно
Постганглионарные волокна
Преганглионарный
синапс (АХ)
Никотиновый рецептор
Вегетативный ганглий
Преганглионарное волокно
Постганглионарные волокна
Преганглионарный
синапс (АХ)
Никотиновый рецептор

Отличия вегетативной и соматической нервной системы
Отличия вегетативной и соматической нервной системы
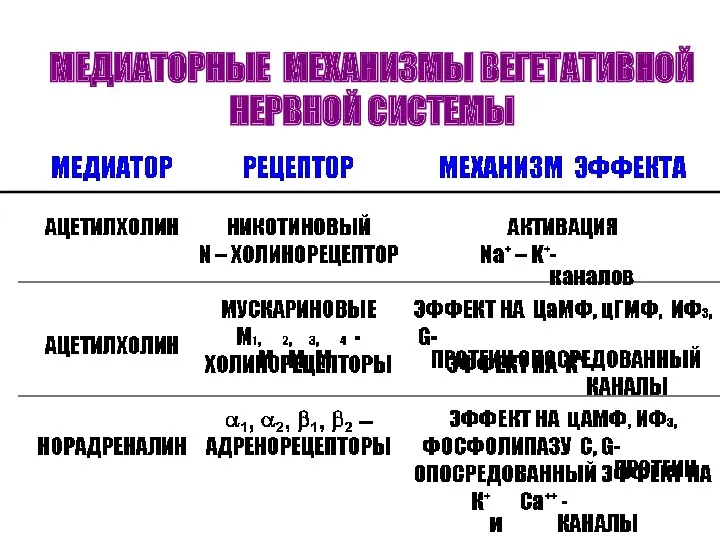
МЕДИАТОРНЫЕ МЕХАНИЗМЫ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
МЕДИАТОРНЫЕ МЕХАНИЗМЫ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
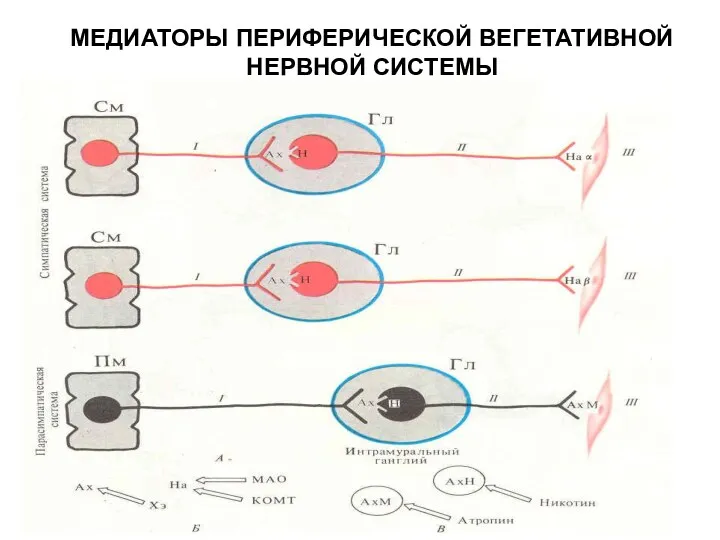
МЕДИАТОРЫ ПЕРИФЕРИЧЕСКОЙ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
МЕДИАТОРЫ ПЕРИФЕРИЧЕСКОЙ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ

Висцеральные сигналы
По сравнению с соматическими характеризуются:
низкой скоростью проведения,
менее развитой системой
Висцеральные сигналы
По сравнению с соматическими характеризуются:
низкой скоростью проведения,
менее развитой системой
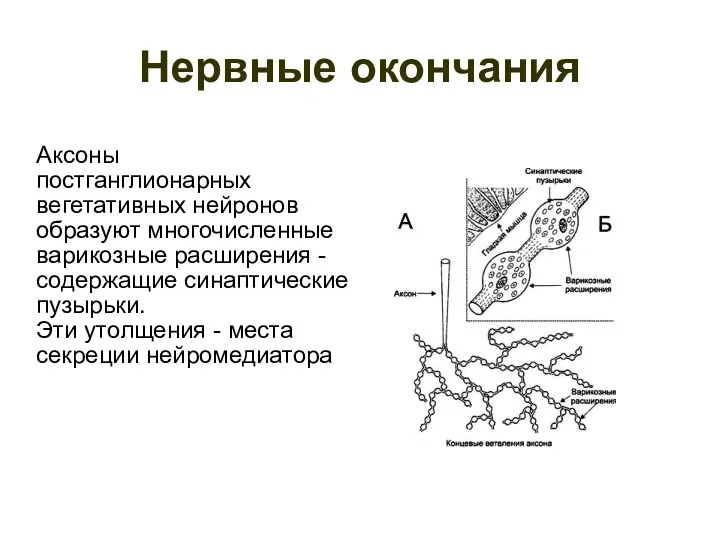
Нервные окончания
Аксоны
постганглионарных
вегетативных нейронов
образуют многочисленные
варикозные расширения -
содержащие синаптические
пузырьки.
Эти утолщения - места
секреции
Нервные окончания
Аксоны
постганглионарных
вегетативных нейронов
образуют многочисленные
варикозные расширения -
содержащие синаптические
пузырьки.
Эти утолщения - места
секреции

ВЗАИМОДЕЙСТВИЕ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ НЕРВНОЙ РЕГУЛЯЦИИ
АНТАГОНИЗМ
СИНЕРГИЗМ
ОТСУТСТВИЕ ВЗАИМОДЕЙСТВИЯ
ВЗАИМОДЕЙСТВИЕ СИМПАТИЧЕСКОЙ И ПАРАСИМПАТИЧЕСКОЙ НЕРВНОЙ РЕГУЛЯЦИИ
АНТАГОНИЗМ
СИНЕРГИЗМ
ОТСУТСТВИЕ ВЗАИМОДЕЙСТВИЯ

Симпатические и парасимпатические эффекты
Симпатические и парасимпатические эффекты

Моносимпатическая регуляция
Моносимпатическая регуляция
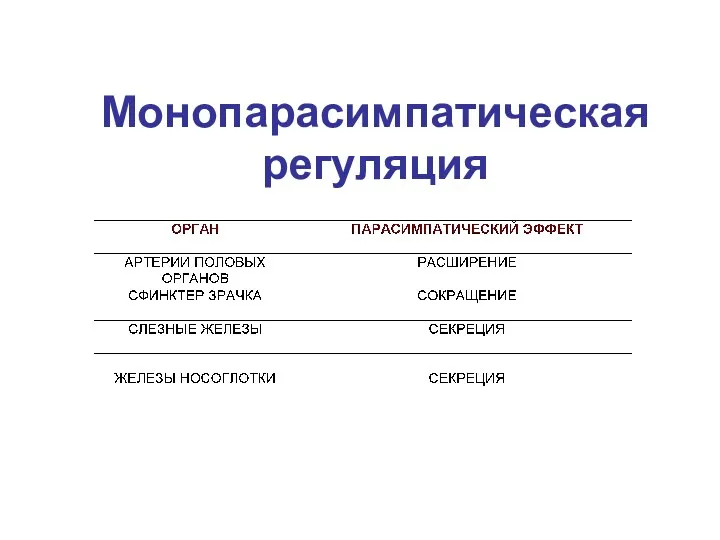
Монопарасимпатическая регуляция
Монопарасимпатическая регуляция

1. Иннервирует только внутренние органы, наделенные собственной моторной активностью.
2. Получает синаптические
1. Иннервирует только внутренние органы, наделенные собственной моторной активностью.
2. Получает синаптические
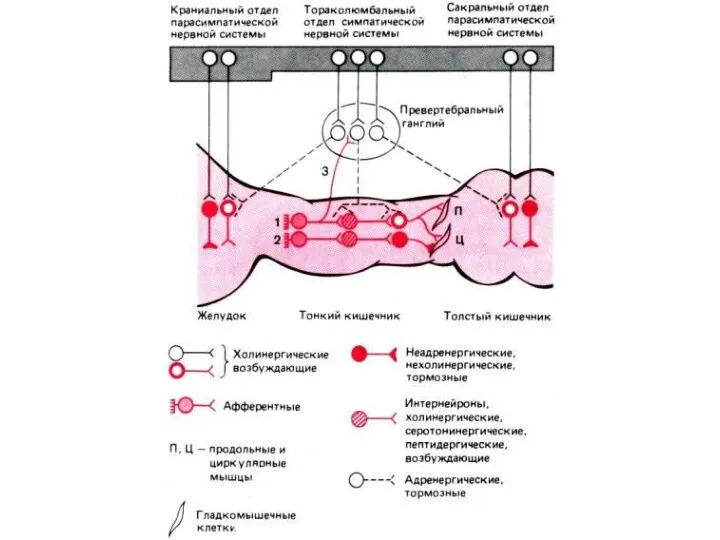
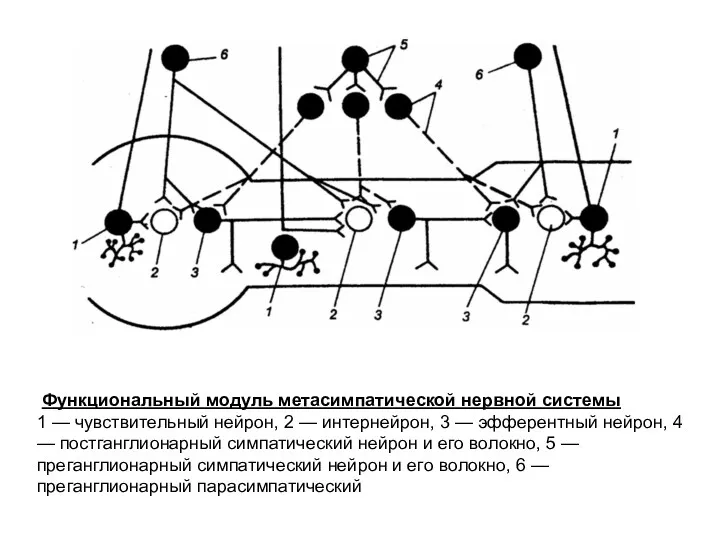
Функциональный модуль метасимпатической нервной системы
1 — чувствительный нейрон, 2 —
Функциональный модуль метасимпатической нервной системы
1 — чувствительный нейрон, 2 —
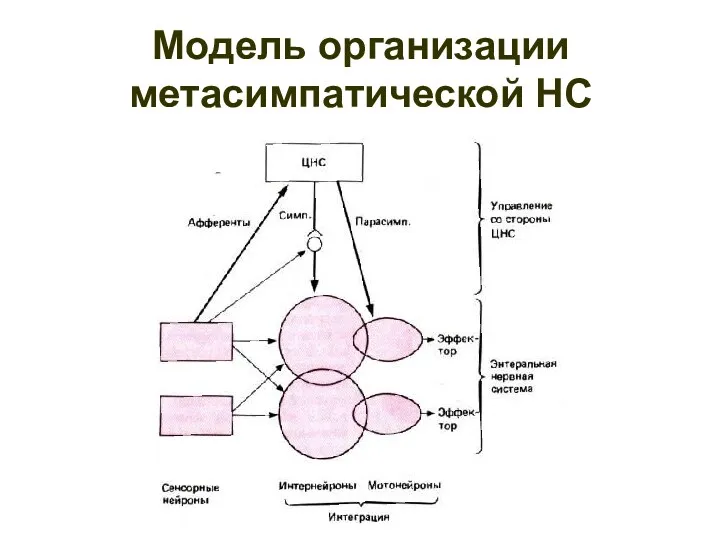
Модель организации метасимпатической НС
Модель организации метасимпатической НС

Аденозинтрифосфат (АТФ)
АТФ локализуется в пресинаптических терминалях эффекторных нейронов метасимпатической нервной системы.
Аденозинтрифосфат (АТФ)
АТФ локализуется в пресинаптических терминалях эффекторных нейронов метасимпатической нервной системы.

Трансдукторы
Клетки, возбуждающиеся обычным путем, а отвечающие эндокринным способом, называют трансдукторами. Аксоны
Трансдукторы
Клетки, возбуждающиеся обычным путем, а отвечающие эндокринным способом, называют трансдукторами. Аксоны
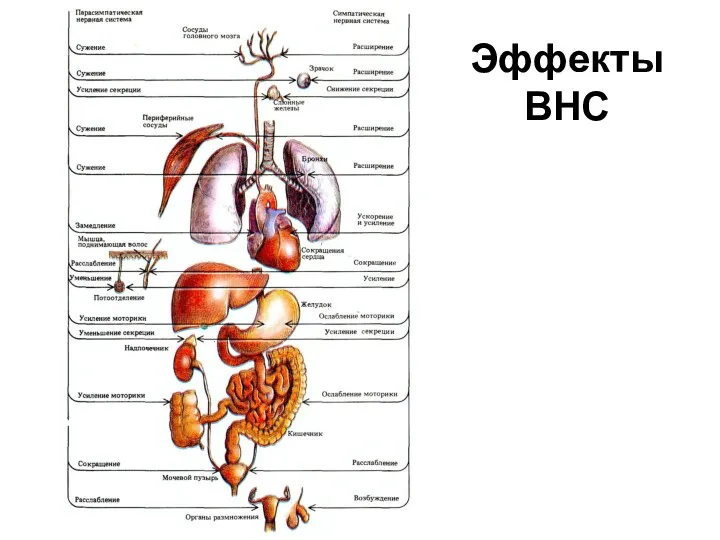
Эффекты ВНС
Эффекты ВНС
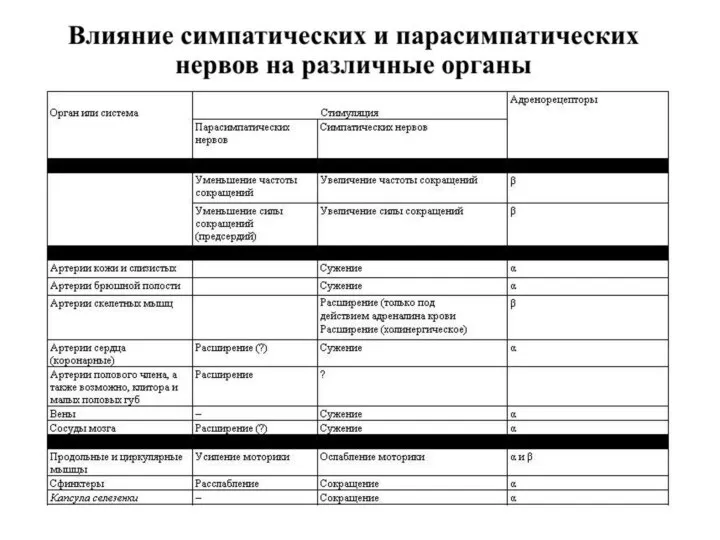
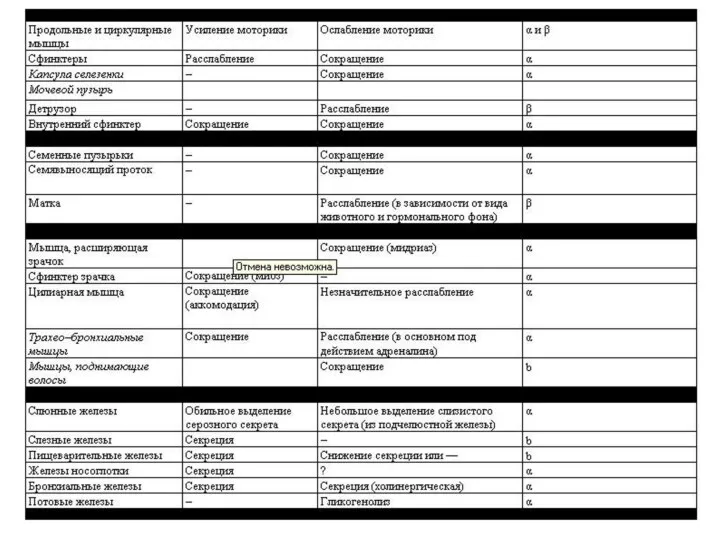
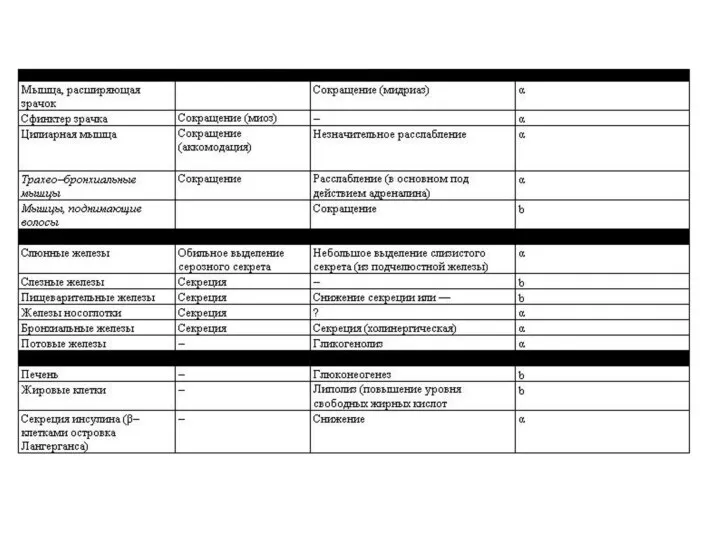

Только ацетилхолин, норадреналин и серотонин удовлетворяют всем критериям, предъявляемым к
Только ацетилхолин, норадреналин и серотонин удовлетворяют всем критериям, предъявляемым к

НЕЙРОМЕДИАТОРЫ
Холинергические нейроны
Все преганглионарные нейроны как в симпатическом, так и в
НЕЙРОМЕДИАТОРЫ
Холинергические нейроны
Все преганглионарные нейроны как в симпатическом, так и в
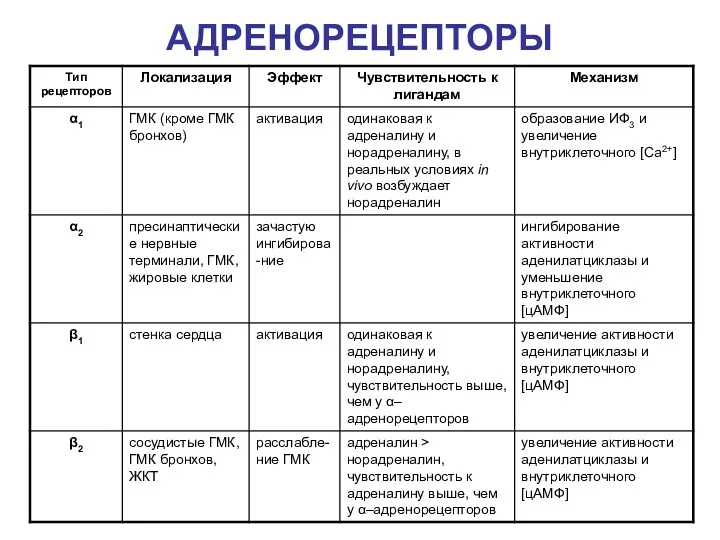
АДРЕНОРЕЦЕПТОРЫ
АДРЕНОРЕЦЕПТОРЫ
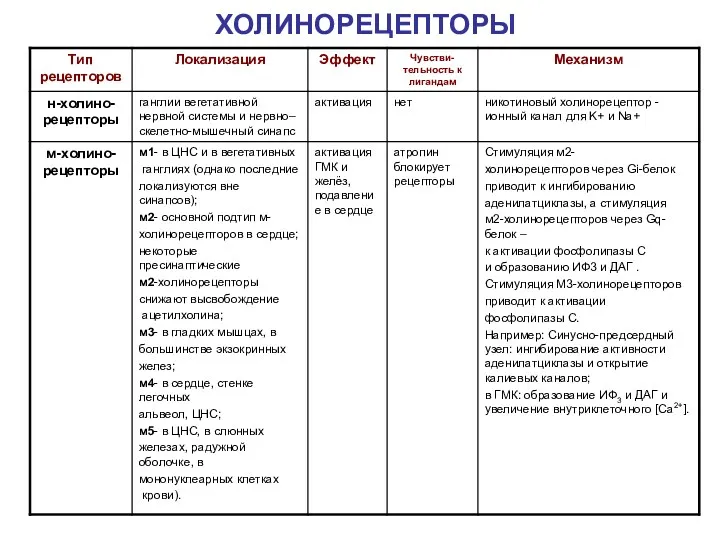
ХОЛИНОРЕЦЕПТОРЫ
ХОЛИНОРЕЦЕПТОРЫ

Вещества действующие на адренергические эффекторы
Симпатомиметики:
естественные агонисты адренорецепторов — норадреналин и адреналин,
вещества
Вещества действующие на адренергические эффекторы
Симпатомиметики:
естественные агонисты адренорецепторов — норадреналин и адреналин,
вещества

Вещества действующие на холинергические эффекторы
Парасимпатомиметики – агонисты холинорецепторов (пилокарпин и метахолин)
Вещества действующие на холинергические эффекторы
Парасимпатомиметики – агонисты холинорецепторов (пилокарпин и метахолин)

Вещества действующие на постганглионарные нейроны ВНС
Возбуждающие
Ацетилхолин секретируется в преганглионарных нейронах и
Вещества действующие на постганглионарные нейроны ВНС
Возбуждающие
Ацетилхолин секретируется в преганглионарных нейронах и

Серотонин
Почти 90% образуется в хромаффинных клетках слизистой оболочки пищеварительного тракта.
Во
Серотонин
Почти 90% образуется в хромаффинных клетках слизистой оболочки пищеварительного тракта.
Во

Эффекты серотонина
На сосуды:
Вазоконстрикция проявляется на денервированных сосудах.
В скелетных мышцах
Эффекты серотонина
На сосуды:
Вазоконстрикция проявляется на денервированных сосудах.
В скелетных мышцах

Серотонинергические рецепторы
Серотонинергические рецепторы
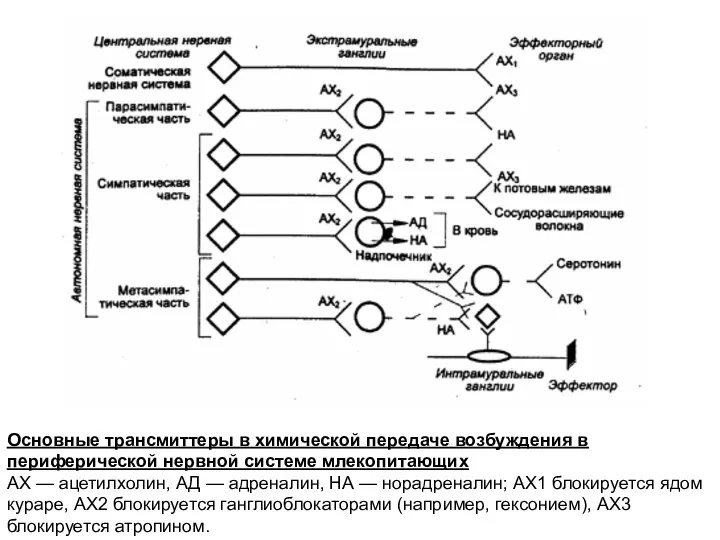
Основные трансмиттеры в химической передаче возбуждения в периферической нервной системе млекопитающих
АХ
Основные трансмиттеры в химической передаче возбуждения в периферической нервной системе млекопитающих
АХ
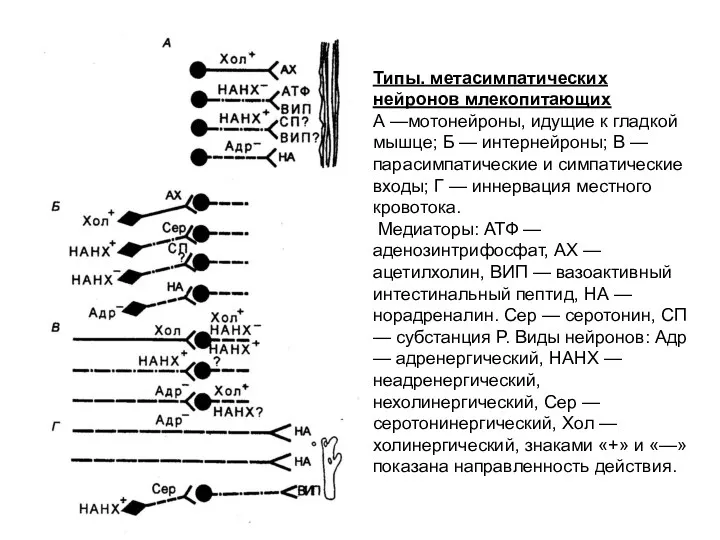
Типы. метасимпатических нейронов млекопитающих
А —мотонейроны, идущие к гладкой мышце; Б —
Типы. метасимпатических нейронов млекопитающих
А —мотонейроны, идущие к гладкой мышце; Б —


ВИДЫ ВЕГЕТАТИВНЫХ РЕФЛЕКСОВ
Висцеро-висцеральный рефлекс
- аксон-рефлекс
Висцеро-соматический рефлекс
Висцеро-сенсорный рефлекс
Висцеро-дермальный рефлекс
Соматовисцеральный рефлекс
Дермо-висцеральный рефлекс
ВИДЫ ВЕГЕТАТИВНЫХ РЕФЛЕКСОВ
Висцеро-висцеральный рефлекс
- аксон-рефлекс
Висцеро-соматический рефлекс
Висцеро-сенсорный рефлекс
Висцеро-дермальный рефлекс
Соматовисцеральный рефлекс
Дермо-висцеральный рефлекс

Висцеро-висцеральный рефлекс
Возбуждение возникает и заканчивается во внутренних органах.
Эффектор способен отвечать
Висцеро-висцеральный рефлекс
Возбуждение возникает и заканчивается во внутренних органах.
Эффектор способен отвечать

Аксон - рефлекс
Это местная ответная реакция ткани на раздражитель без участия
Аксон - рефлекс
Это местная ответная реакция ткани на раздражитель без участия
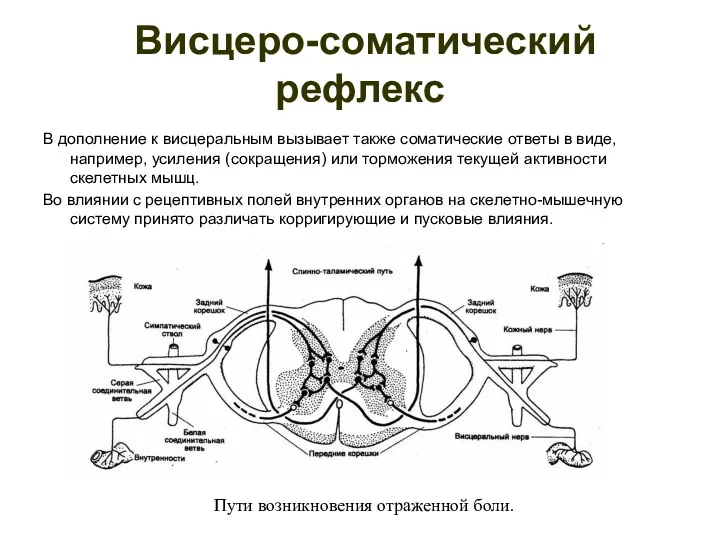
Висцеро-соматический рефлекс
В дополнение к висцеральным вызывает также соматические ответы в
Висцеро-соматический рефлекс
В дополнение к висцеральным вызывает также соматические ответы в
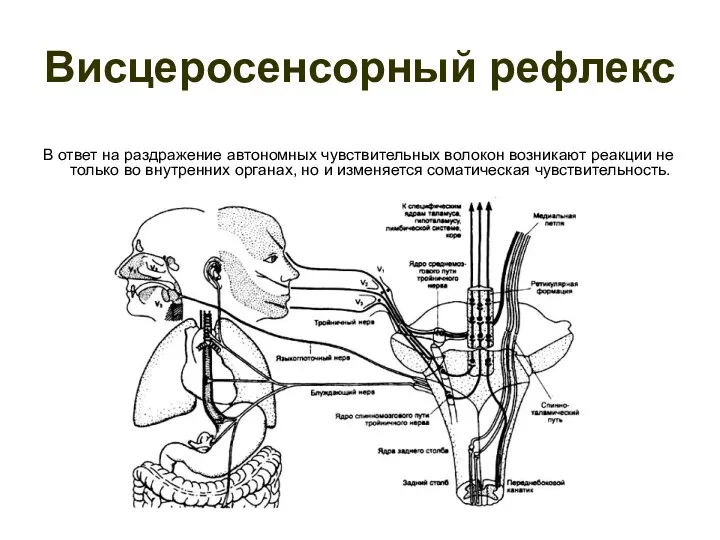
Висцеросенсорный рефлекс
В ответ на раздражение автономных чувствительных волокон возникают реакции не
Висцеросенсорный рефлекс
В ответ на раздражение автономных чувствительных волокон возникают реакции не

РЕАКЦИИ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Симпатическая нервная система в зависимости от характера и
РЕАКЦИИ СИМПАТИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Симпатическая нервная система в зависимости от характера и

РЕАКЦИИ ПАРАСИМПАТИЧЕСКОЙ СИСТЕМЫ
Парасимпатическая система осуществляет локальный и более специфический контроль функций
РЕАКЦИИ ПАРАСИМПАТИЧЕСКОЙ СИСТЕМЫ
Парасимпатическая система осуществляет локальный и более специфический контроль функций
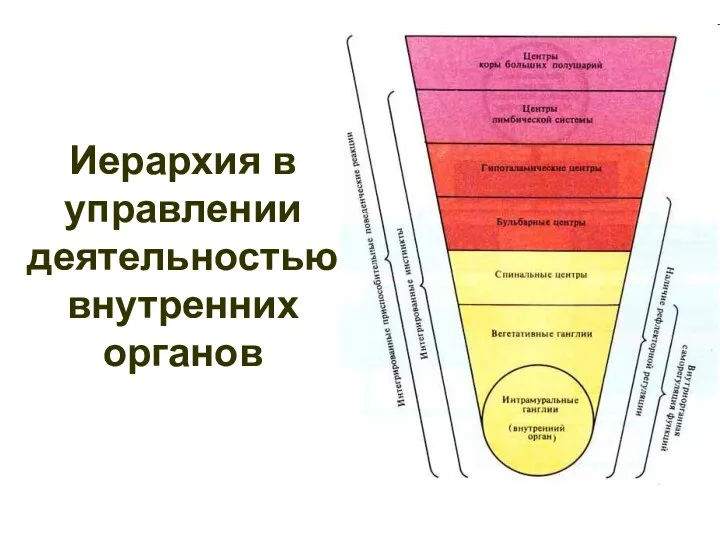
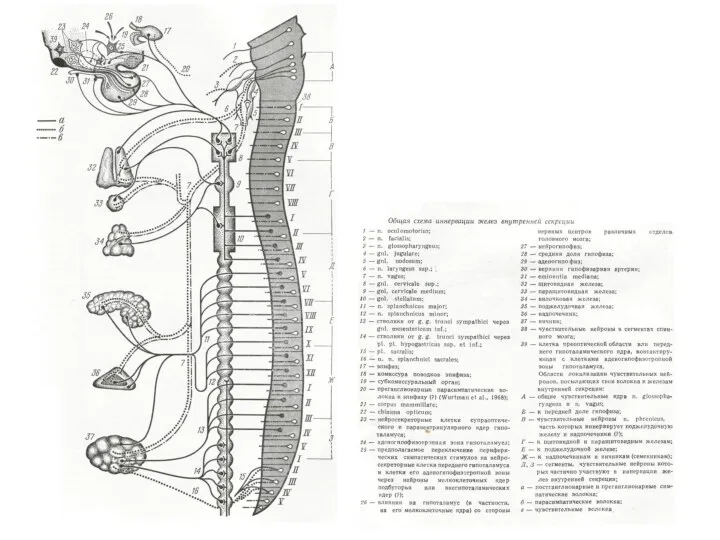
Иерархия в управлении деятельностью внутренних органов
Иерархия в управлении деятельностью внутренних органов

Спинальные центры
Спинальные центры
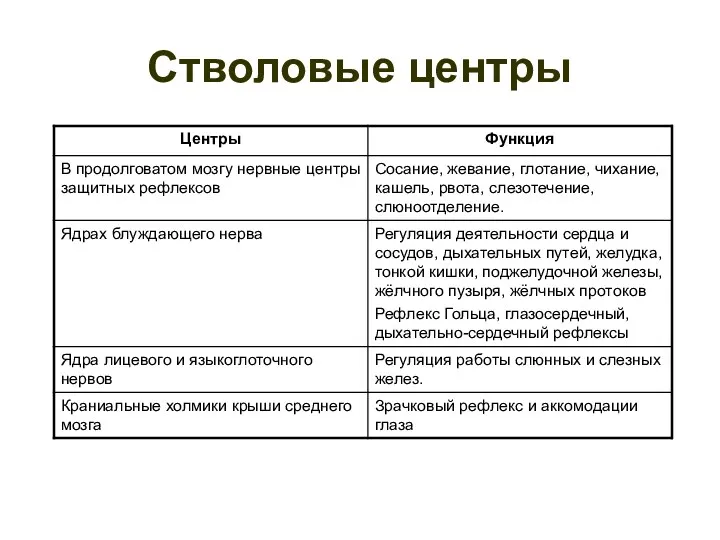
Стволовые центры
Стволовые центры
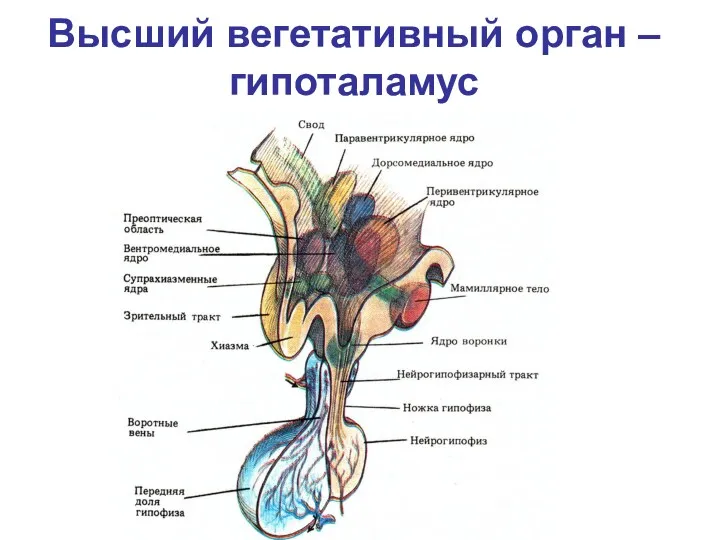
Высший вегетативный орган – гипоталамус
Высший вегетативный орган – гипоталамус
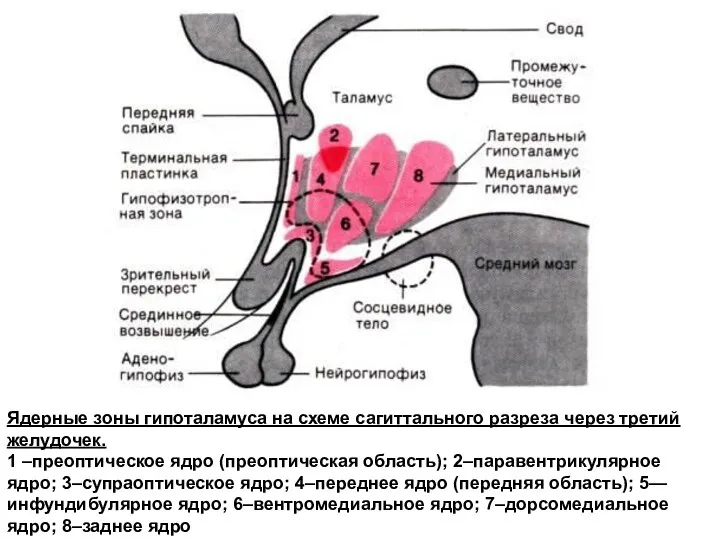
Ядерные зоны гипоталамуса на схеме сагиттального разреза через третий желудочек.
1
Ядерные зоны гипоталамуса на схеме сагиттального разреза через третий желудочек.
1
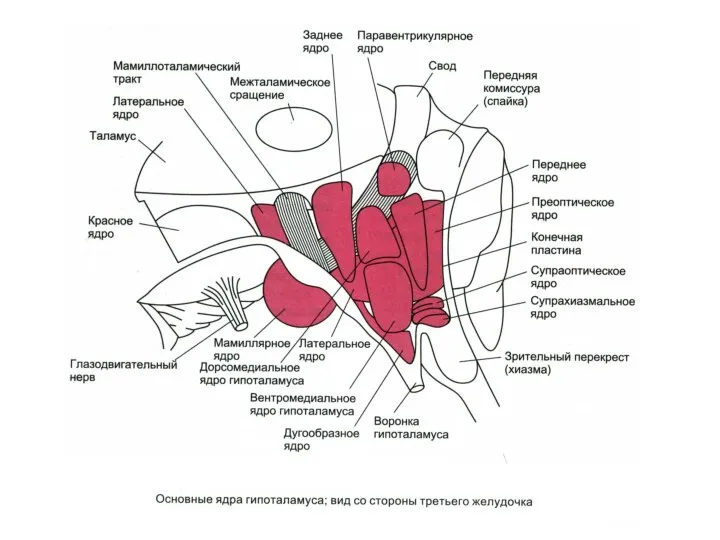

Двусторонние связи гипоталамуса с:
Таламусом
Лимбической системой
КБП
Двусторонние связи гипоталамуса с:
Таламусом
Лимбической системой
КБП

Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливать изменения химического состава крови
Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливать изменения химического состава крови

Гипоталамус:
чувствительное звено
Чувствительная информация от внутренних органов поступает в гипоталамус по восходящим
Гипоталамус:
чувствительное звено
Чувствительная информация от внутренних органов поступает в гипоталамус по восходящим

Гипоталамус:
эффекторное звено
Гипоталамус играет ведущую роль в поддержании гомеостаза.
Стимуляция задних ядер гипоталамуса
Гипоталамус:
эффекторное звено
Гипоталамус играет ведущую роль в поддержании гомеостаза.
Стимуляция задних ядер гипоталамуса

Гипоталамус:
поведенческая функция
Гипоталамус участвует в формировании поведенческих реакций организма, необходимых для
Гипоталамус:
поведенческая функция
Гипоталамус участвует в формировании поведенческих реакций организма, необходимых для
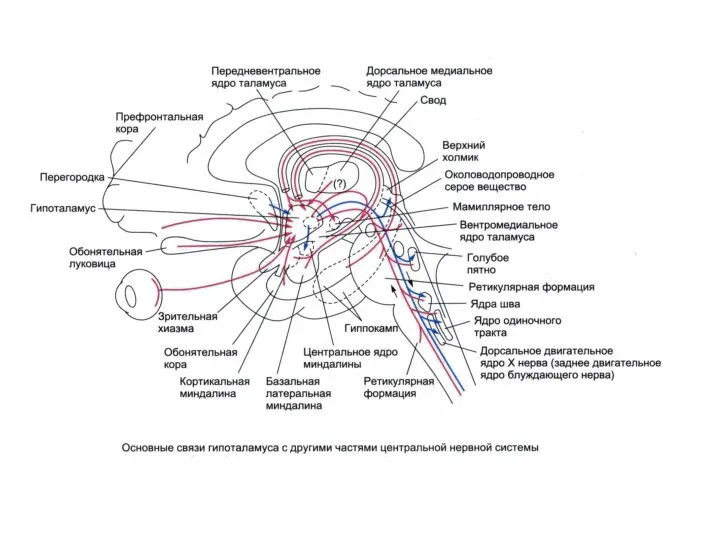
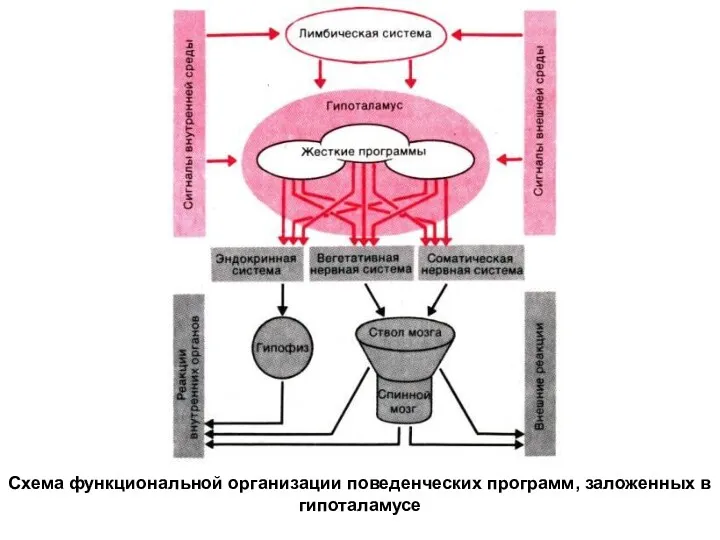
Схема функциональной организации поведенческих программ, заложенных в гипоталамусе
Схема функциональной организации поведенческих программ, заложенных в гипоталамусе
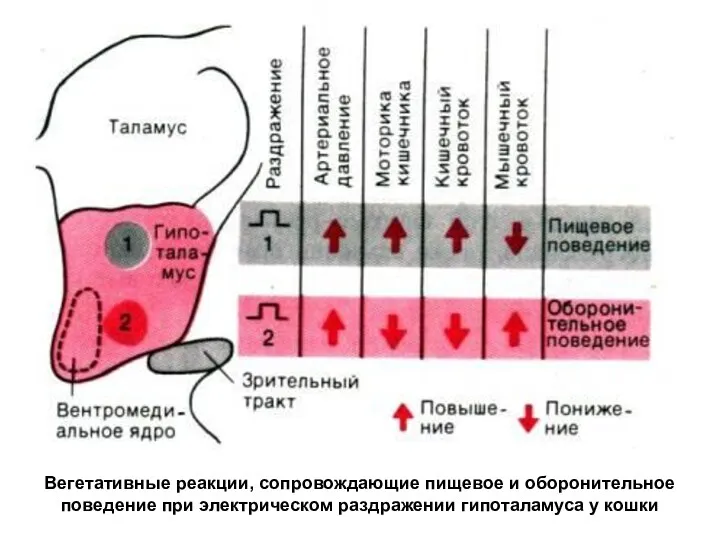
Вегетативные реакции, сопровождающие пищевое и оборонительное поведение при электрическом раздражении гипоталамуса
Вегетативные реакции, сопровождающие пищевое и оборонительное поведение при электрическом раздражении гипоталамуса
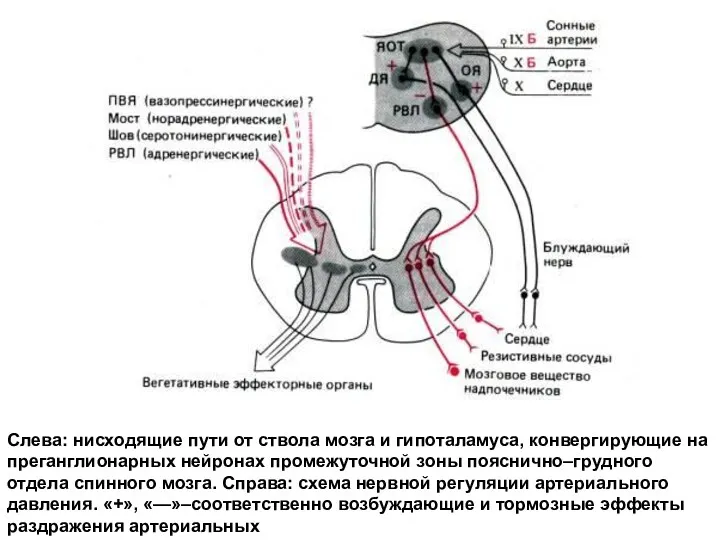
Слева: нисходящие пути от ствола мозга и гипоталамуса, конвергирующие на преганглионарных
Слева: нисходящие пути от ствола мозга и гипоталамуса, конвергирующие на преганглионарных
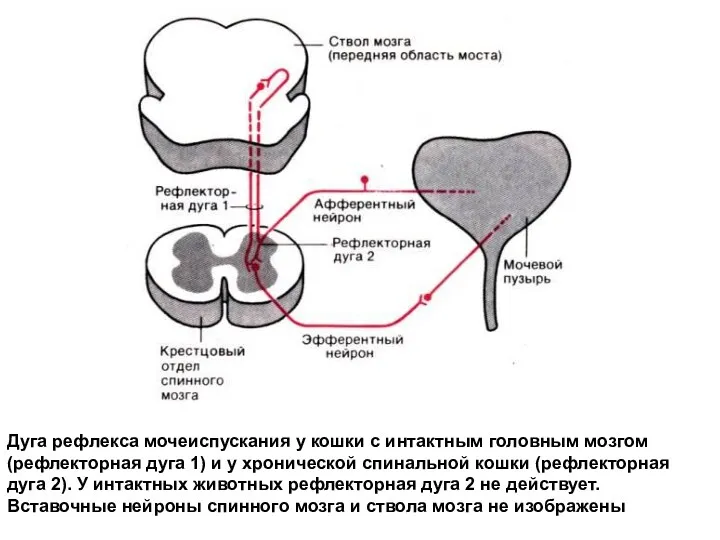
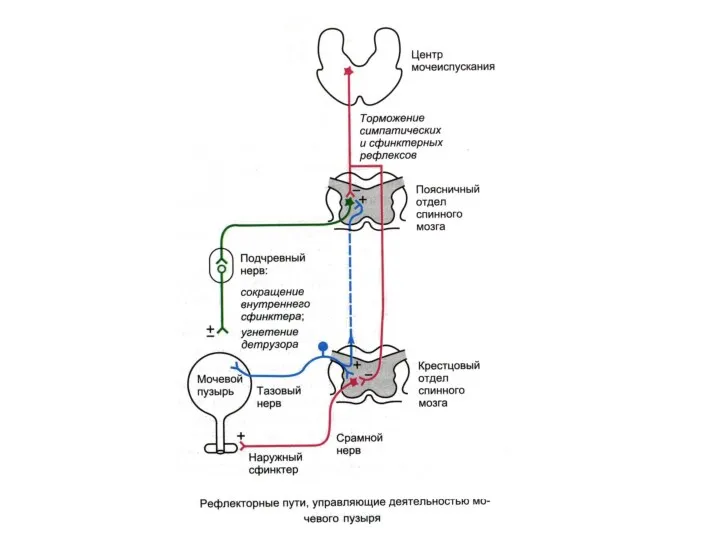
Дуга рефлекса мочеиспускания у кошки с интактным головным мозгом (рефлекторная дуга
Дуга рефлекса мочеиспускания у кошки с интактным головным мозгом (рефлекторная дуга
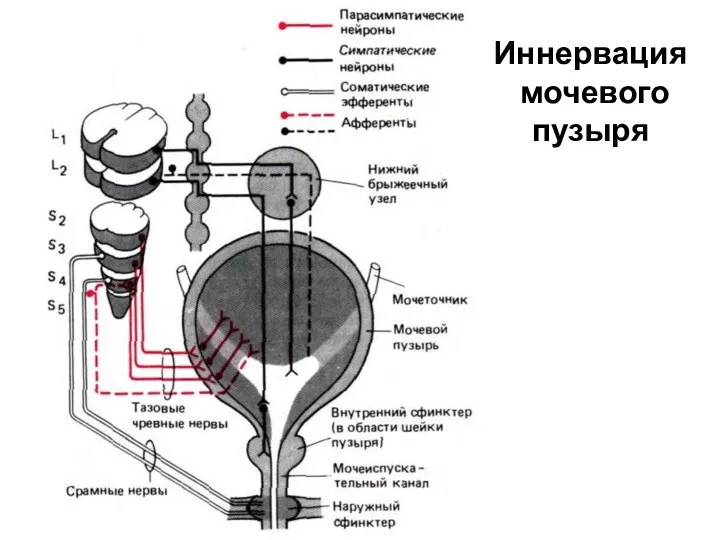
Иннервация
мочевого
пузыря
Иннервация
мочевого
пузыря
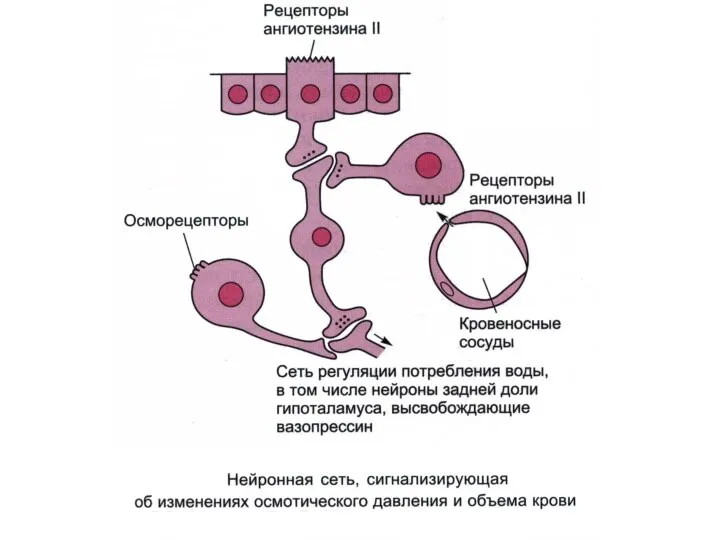
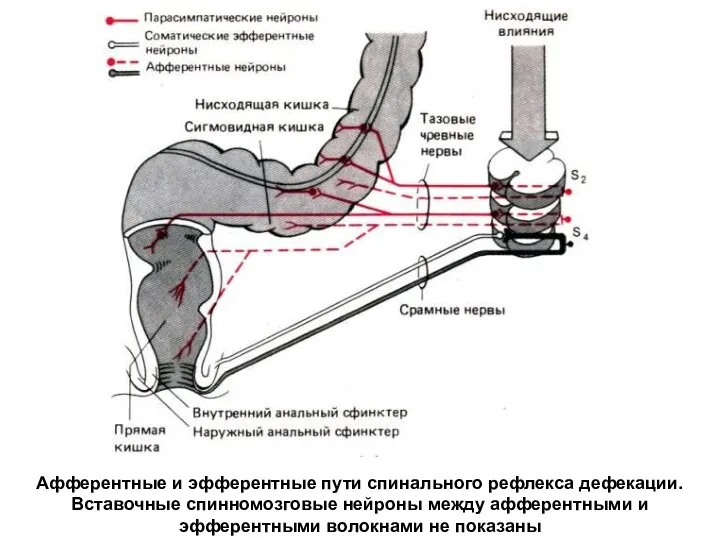
Афферентные и эфферентные пути спинального рефлекса дефекации. Вставочные спинномозговые нейроны между
Афферентные и эфферентные пути спинального рефлекса дефекации. Вставочные спинномозговые нейроны между
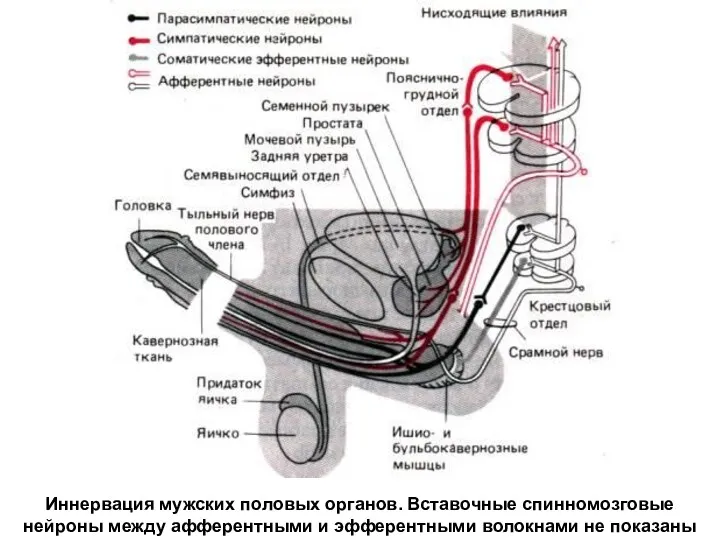
Иннервация мужских половых органов. Вставочные спинномозговые нейроны между афферентными и эфферентными
Иннервация мужских половых органов. Вставочные спинномозговые нейроны между афферентными и эфферентными

Таламус
- ограничивает поступление афферентной импульсации в кору большого мозга от
Таламус
- ограничивает поступление афферентной импульсации в кору большого мозга от

Ретикулярная формация
Ее нейроны формируют центры дыхания и кровообращения и реализуют свое
Ретикулярная формация
Ее нейроны формируют центры дыхания и кровообращения и реализуют свое

Голубое пятно
Его норадренергические нейроны иннервируют артериолы и капилляры коры больших полушарий
Голубое пятно
Его норадренергические нейроны иннервируют артериолы и капилляры коры больших полушарий

Лимбическая система
Лимбическая система обеспечивает взаимодействие экстероцептивных (обонятельных, слуховых и др.) и
Лимбическая система
Лимбическая система обеспечивает взаимодействие экстероцептивных (обонятельных, слуховых и др.) и

Лимбическая система
Участвует в приспособительной регуляции деятельности систем:
Сердечнососудистой
Дыхательной
Пищеварительной
Лимбическая система
Участвует в приспособительной регуляции деятельности систем:
Сердечнососудистой
Дыхательной
Пищеварительной

Мозжечок
Мозжечок благодаря наличию двойного (активирующего и тормозного) механизма действия способен оказывать
Мозжечок
Мозжечок благодаря наличию двойного (активирующего и тормозного) механизма действия способен оказывать

Мозжечок
Реализует свое влияние через симпатическую нервную систему и эндокринные железы,
Мозжечок
Реализует свое влияние через симпатическую нервную систему и эндокринные железы,
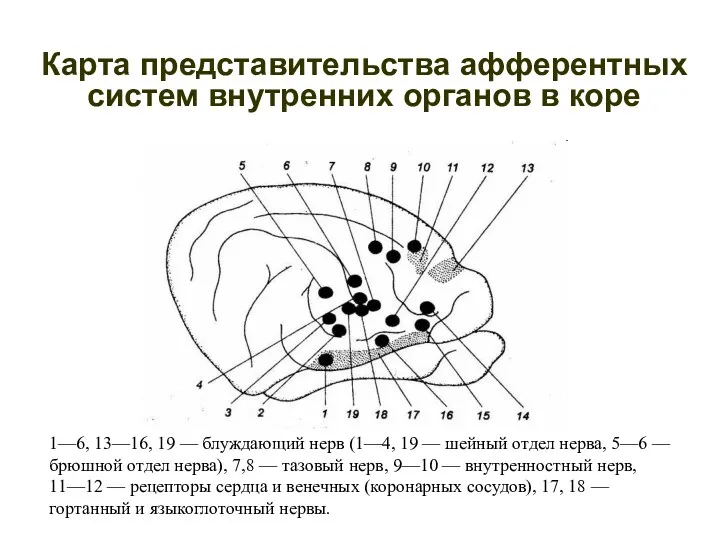
Карта представительства афферентных систем внутренних органов в коре
1—6, 13—16, 19 —
Карта представительства афферентных систем внутренних органов в коре
1—6, 13—16, 19 —

Кора большого мозга.
КБП — высший интегративный центр регуляции всех функций
Кора большого мозга.
КБП — высший интегративный центр регуляции всех функций

Стимуляция двигательной зоны коры вызывает такие же изменения деятельности сердечно-сосудистой системы
Стимуляция двигательной зоны коры вызывает такие же изменения деятельности сердечно-сосудистой системы

Тонус вегетативных центров
Многие преганглионарные и ганглионарные вегетативные нейроны обладают постоянной активностью,
Тонус вегетативных центров
Многие преганглионарные и ганглионарные вегетативные нейроны обладают постоянной активностью,

Значение тонуса вегетативных центров.
Тонус вегетативных центров играет важную роль в
Значение тонуса вегетативных центров.
Тонус вегетативных центров играет важную роль в

Вегетативный портрет.
Симпатикотоники и парасимпатикотоники.
Деление лиц на симпатикотоников и парасимпатикотоников на основании
Вегетативный портрет.
Симпатикотоники и парасимпатикотоники.
Деление лиц на симпатикотоников и парасимпатикотоников на основании

Трофическое действие нервной системы
Идею о трофическом действии НС сформулировал И. П.
Трофическое действие нервной системы
Идею о трофическом действии НС сформулировал И. П.

Повышение работоспособности утомленной изолированной икроножной мышцы лягушки (1) при раздражении симпатических
Повышение работоспособности утомленной изолированной икроножной мышцы лягушки (1) при раздражении симпатических

Такое действие симпатической нервной системы Л.А. Орбели назвал адаптационно - трофическим.
Такое действие симпатической нервной системы Л.А. Орбели назвал адаптационно - трофическим.

Вегетативная нервная система детей
Вегетативная нервная система детей

Вегетативная нервная система детей
В онтогенезе ВНС претерпевает существенные структурные и функциональные
Вегетативная нервная система детей
В онтогенезе ВНС претерпевает существенные структурные и функциональные

ВНС у детей
Медиатором преганглионарных симпатических нейронов является адреноподобное вещество (у взрослых
ВНС у детей
Медиатором преганглионарных симпатических нейронов является адреноподобное вещество (у взрослых

ВНС у детей
В процессе онтогенеза число холинергических синапсов в структурах ВНС
ВНС у детей
В процессе онтогенеза число холинергических синапсов в структурах ВНС

ВНС у детей
Автоматизм клеток симпатических ганглиев и низкий мембранный потенциал симпатических
ВНС у детей
Автоматизм клеток симпатических ганглиев и низкий мембранный потенциал симпатических

ВНС у детей
Характерными особенностями ВНС в первые годы жизни ребенка являются
ВНС у детей
Характерными особенностями ВНС в первые годы жизни ребенка являются

ВНС у детей
Различная скорость созревания симпатической и парасимпатической нервной системы в
ВНС у детей
Различная скорость созревания симпатической и парасимпатической нервной системы в

ВНС у детей
Парасимпатический отдел начинает включаться в рефлекторные реакции сердца с
ВНС у детей
Парасимпатический отдел начинает включаться в рефлекторные реакции сердца с

ВНС у детей
Максимальное замедление пульса от 150 до 30 ударов было
ВНС у детей
Максимальное замедление пульса от 150 до 30 ударов было

ВНС у детей
В регуляцию ЖКТ также сначала включается парасимпатическая нервная система.
ВНС у детей
В регуляцию ЖКТ также сначала включается парасимпатическая нервная система.

ВНС у детей
Механизм формирования тонуса вегетативных центров в онтогенезе.
В формировании тонуса
ВНС у детей
Механизм формирования тонуса вегетативных центров в онтогенезе.
В формировании тонуса

ВНС у детей
Для оценки степени выраженности тонуса блуждающего нерва в детском
ВНС у детей
Для оценки степени выраженности тонуса блуждающего нерва в детском

ВНС у детей
Важное значение в становлении тонуса блуждающего нерва играет импульсация
ВНС у детей
Важное значение в становлении тонуса блуждающего нерва играет импульсация
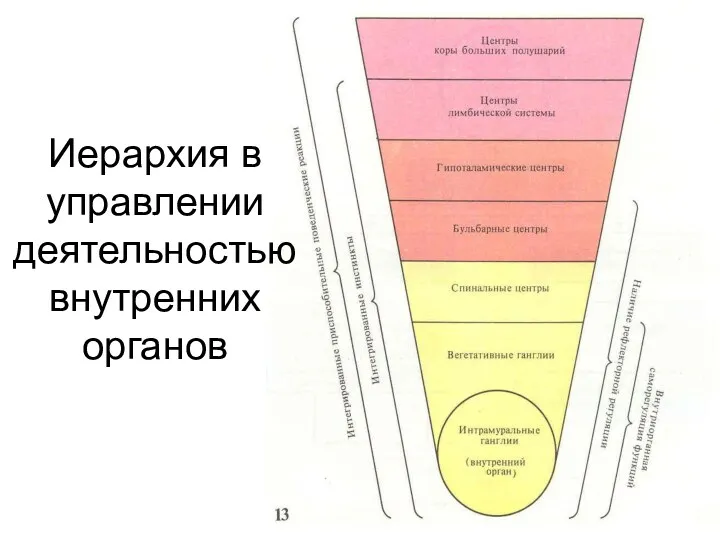
Иерархия в управлении деятельностью внутренних органов
Иерархия в управлении деятельностью внутренних органов
Физиология
эндокринной
системы
Физиология
эндокринной
системы
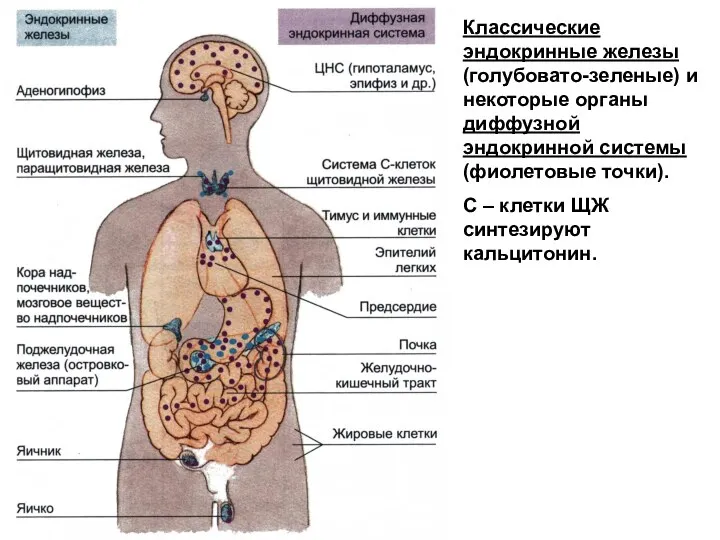
Классические эндокринные железы (голубовато-зеленые) и некоторые органы диффузной эндокринной системы (фиолетовые
Классические эндокринные железы (голубовато-зеленые) и некоторые органы диффузной эндокринной системы (фиолетовые

Эндокринная система
1. Эндокринные железы
ГИПОФИЗ (аденогипофиз и нейрогипофиз)
НАДПОЧЕЧНИКИ (кора и мозговое в-во)
ЩИТОВИДНАЯ
Эндокринная система
1. Эндокринные железы
ГИПОФИЗ (аденогипофиз и нейрогипофиз)
НАДПОЧЕЧНИКИ (кора и мозговое в-во)
ЩИТОВИДНАЯ
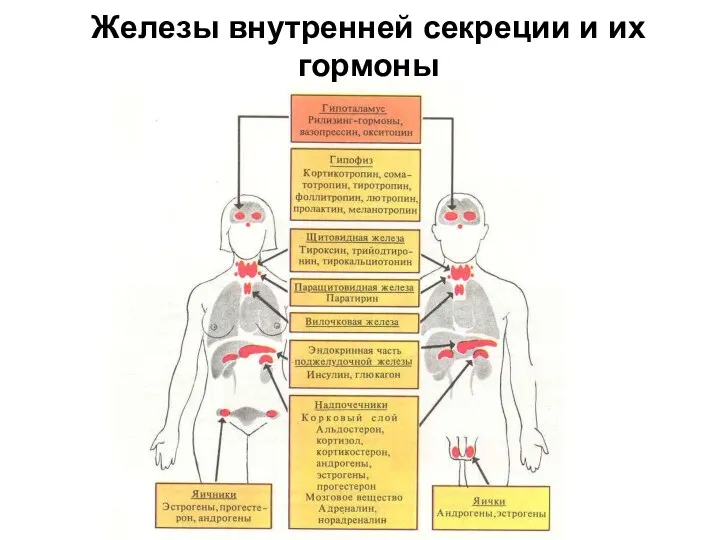
Железы внутренней секреции и их гормоны
Железы внутренней секреции и их гормоны

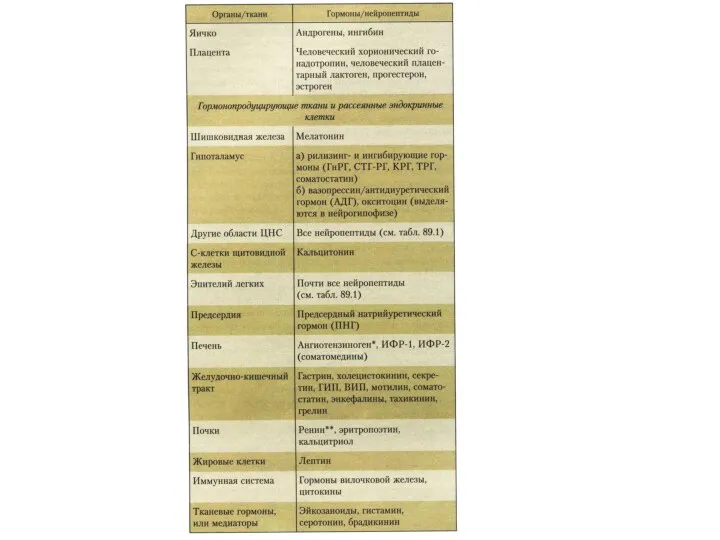

Химическая природа и пути действия гормонов
Простые и сложные белки, - через
Химическая природа и пути действия гормонов
Простые и сложные белки, - через
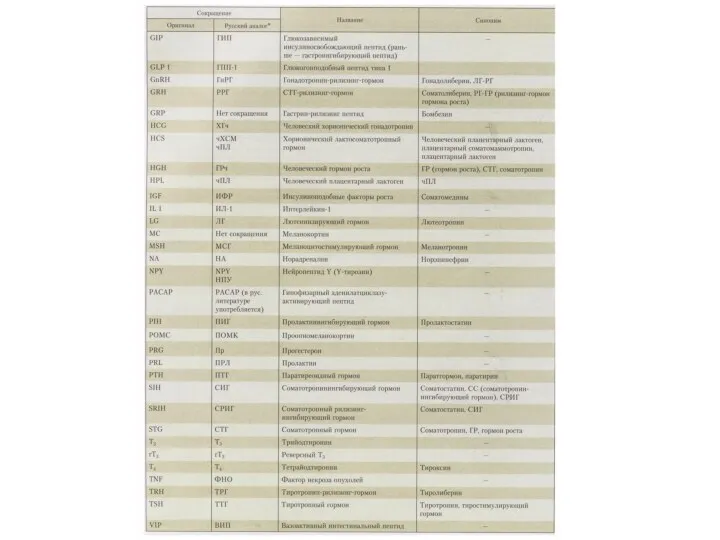

Виды действия гормонов
Метаболическое
Морфогенетическое
Кинетическое
Корригирующее
Виды действия гормонов
Метаболическое
Морфогенетическое
Кинетическое
Корригирующее
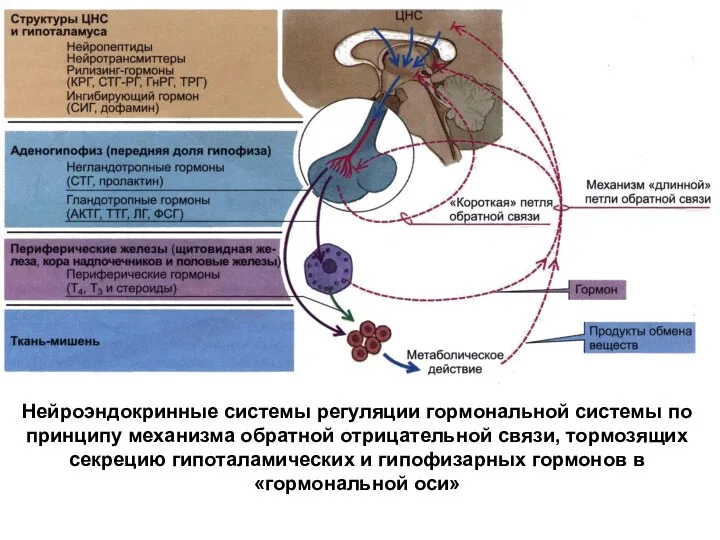
Нейроэндокринные системы регуляции гормональной системы по принципу механизма обратной отрицательной связи,
Нейроэндокринные системы регуляции гормональной системы по принципу механизма обратной отрицательной связи,
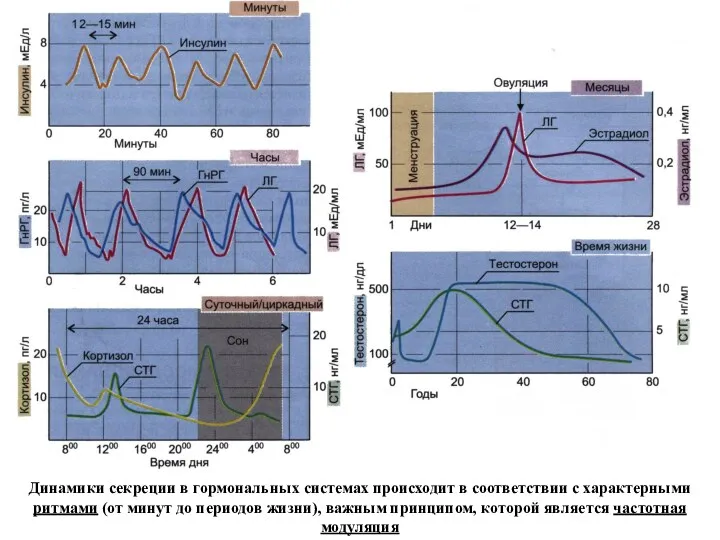
Динамики секреции в гормональных системах происходит в соответствии с характерными ритмами
Динамики секреции в гормональных системах происходит в соответствии с характерными ритмами
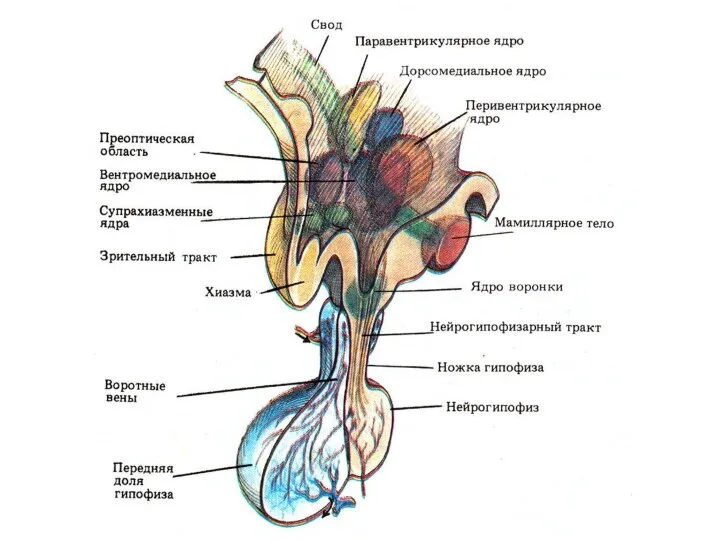

Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливать изменения химического состава крови
Нейроны гипоталамуса
Обладают рецепторной функцией и способны улавливать изменения химического состава крови

Двусторонние связи гипоталамуса с:
Таламусом
Лимбической системой
КБП
Двусторонние связи гипоталамуса с:
Таламусом
Лимбической системой
КБП

Гипоталамус
Вырабатывает собственные гормоны
Эффекторные: окситоцин и вазопрессин (супраоптическое и паравентрикулярное)
Гипоталамус
Вырабатывает собственные гормоны
Эффекторные: окситоцин и вазопрессин (супраоптическое и паравентрикулярное)
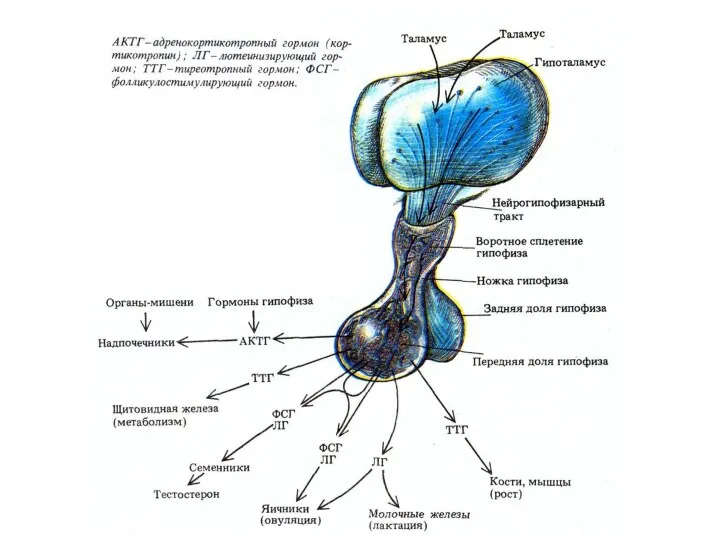
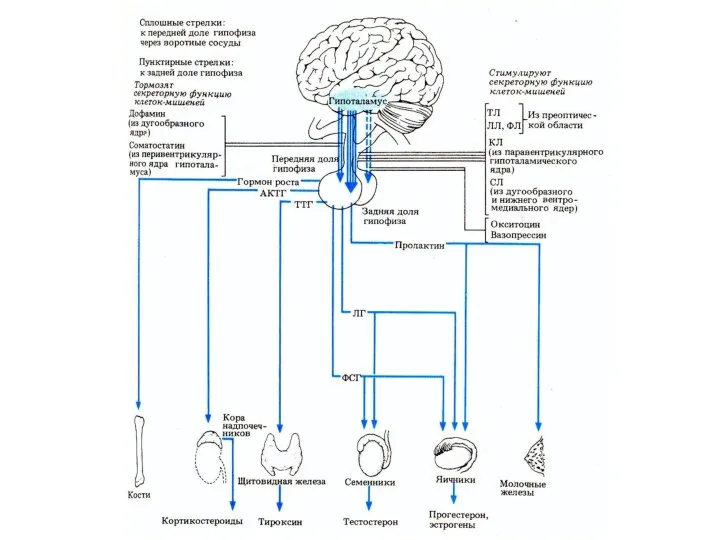
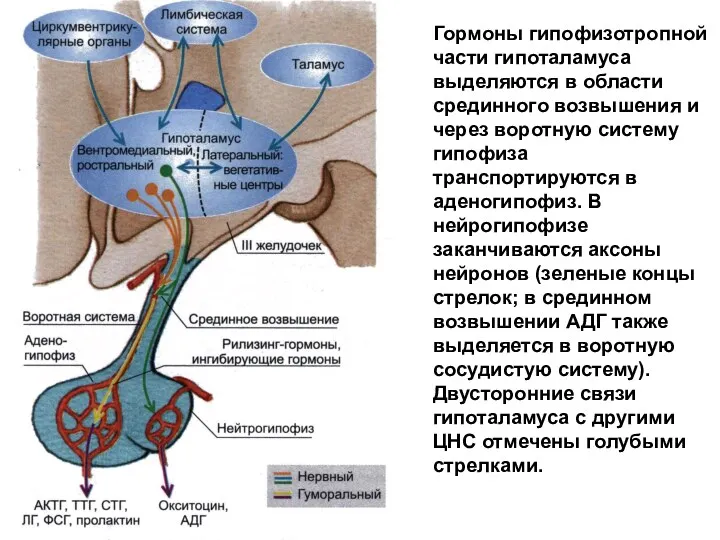
Гормоны гипофизотропной части гипоталамуса выделяются в области срединного возвышения и через
Гормоны гипофизотропной части гипоталамуса выделяются в области срединного возвышения и через
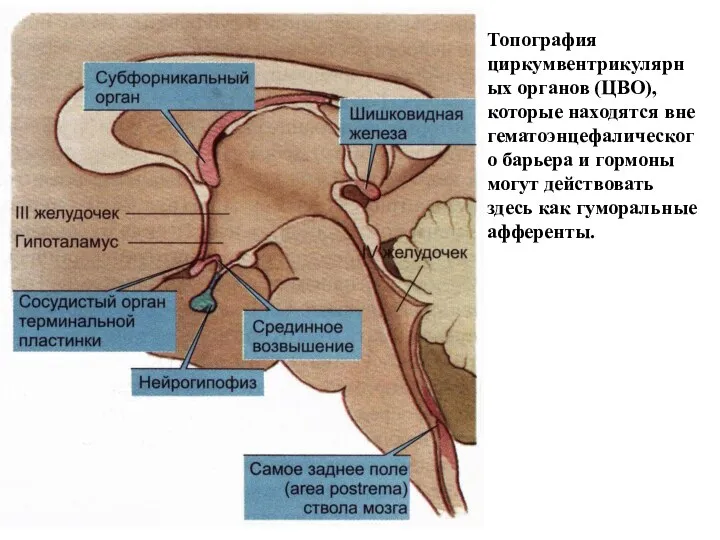
Топография циркумвентрикулярных органов (ЦВО), которые находятся вне гематоэнцефалического барьера и гормоны
Топография циркумвентрикулярных органов (ЦВО), которые находятся вне гематоэнцефалического барьера и гормоны
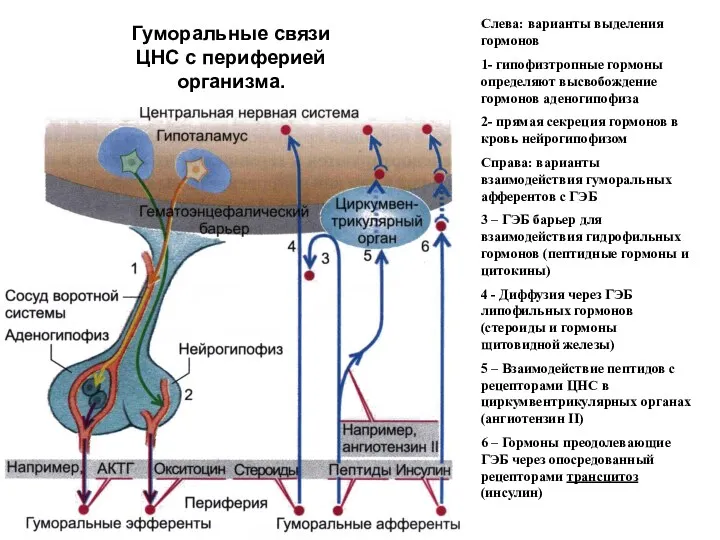
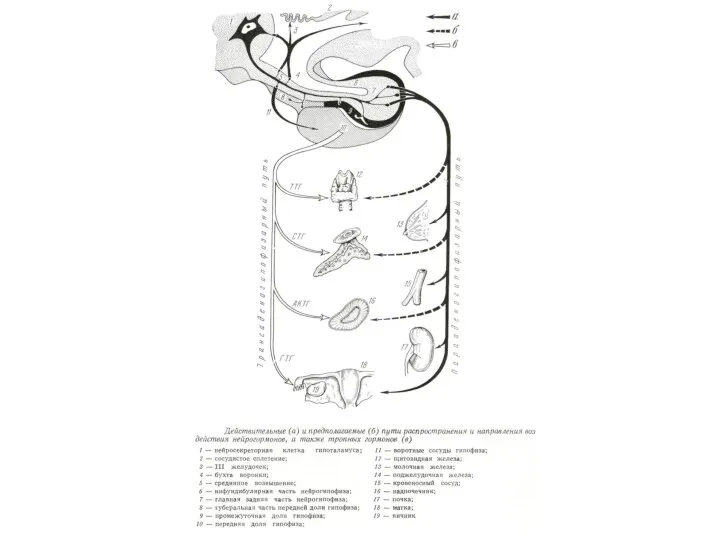
Гуморальные связи ЦНС с периферией организма.
Слева: варианты выделения гормонов
1- гипофизтропные гормоны
Гуморальные связи ЦНС с периферией организма.
Слева: варианты выделения гормонов
1- гипофизтропные гормоны
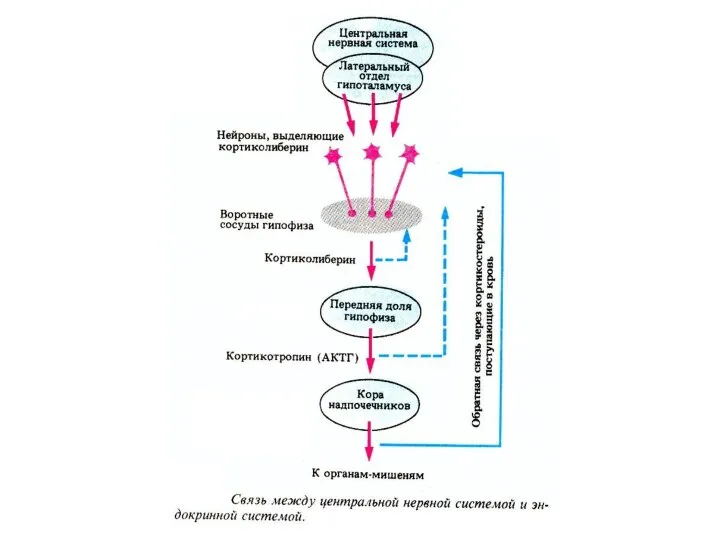
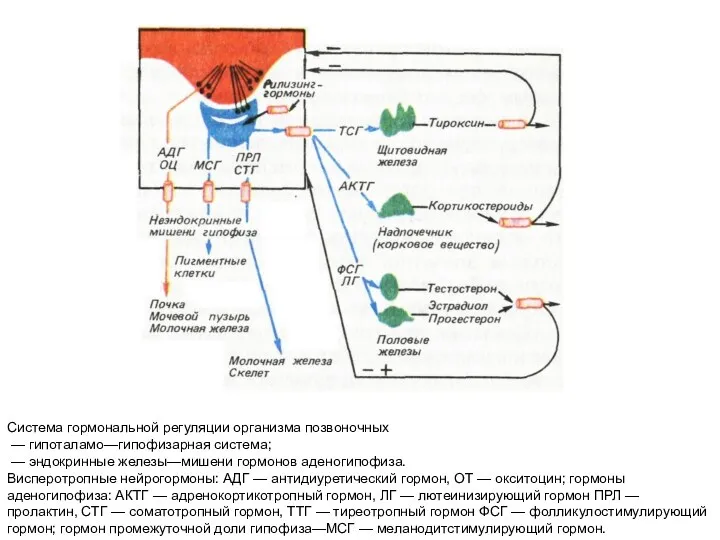
Система гормональной регуляции организма позвоночных
— гипоталамо—гипофизарная система;
—
Система гормональной регуляции организма позвоночных
— гипоталамо—гипофизарная система;
—
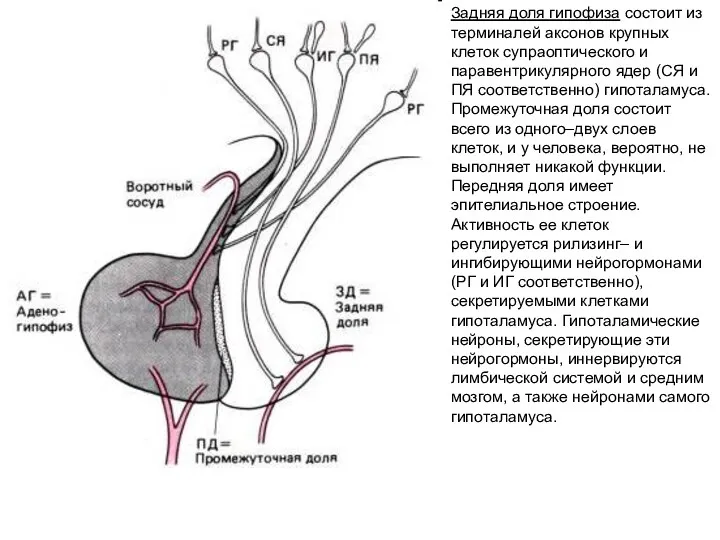
Задняя доля гипофиза состоит из терминалей аксонов крупных клеток супраоптического и
Задняя доля гипофиза состоит из терминалей аксонов крупных клеток супраоптического и
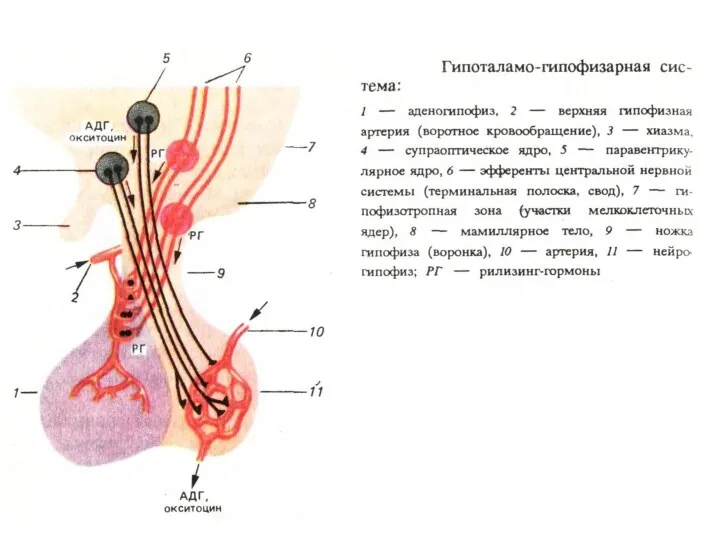
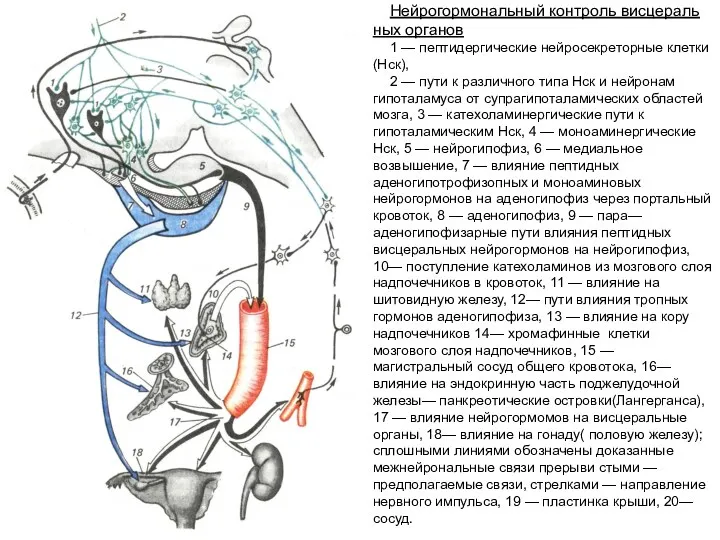
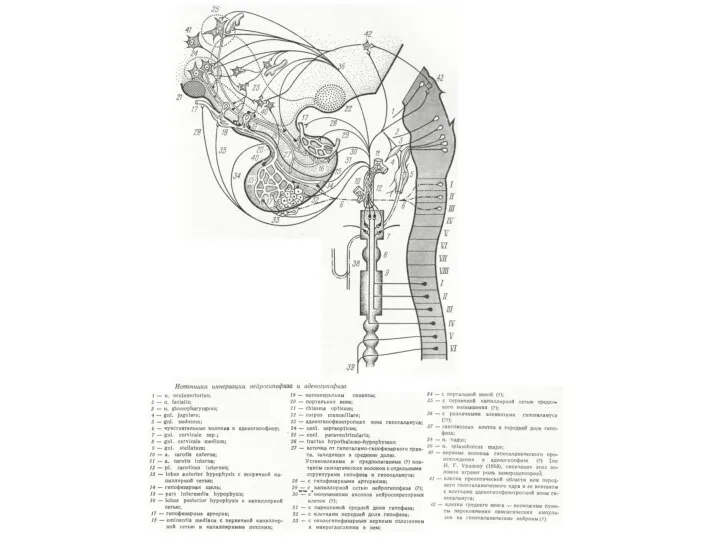
Нейрогормональный контроль висцеральных органов
1 — пептидергические нейросекреторные клетки (Нск),
2 — пути
Нейрогормональный контроль висцеральных органов
1 — пептидергические нейросекреторные клетки (Нск),
2 — пути


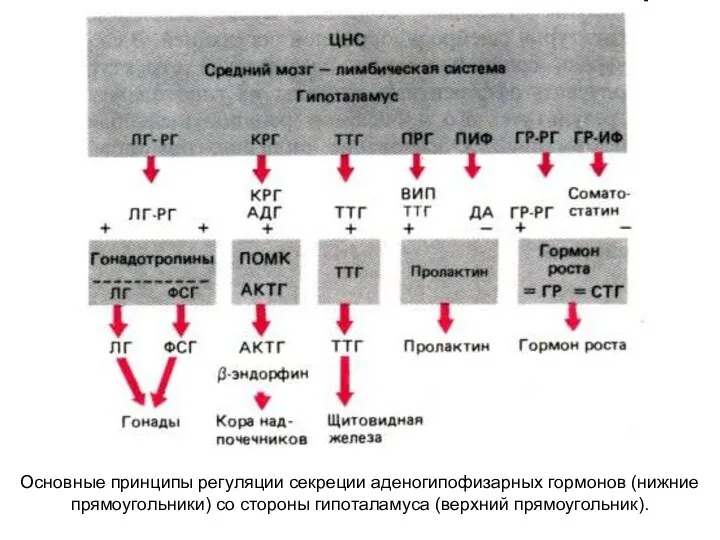
Основные принципы регуляции секреции аденогипофизарных гормонов (нижние прямоугольники) со стороны гипоталамуса
Основные принципы регуляции секреции аденогипофизарных гормонов (нижние прямоугольники) со стороны гипоталамуса
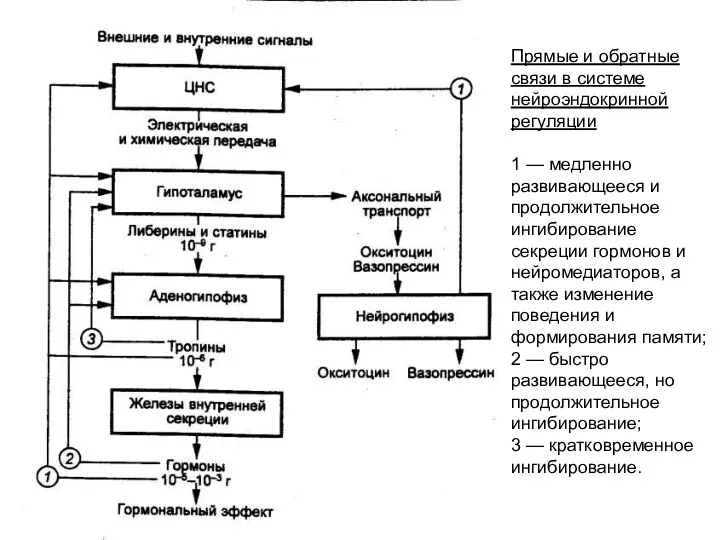
Прямые и обратные связи в системе нейроэндокринной регуляции
1 — медленно
Прямые и обратные связи в системе нейроэндокринной регуляции
1 — медленно
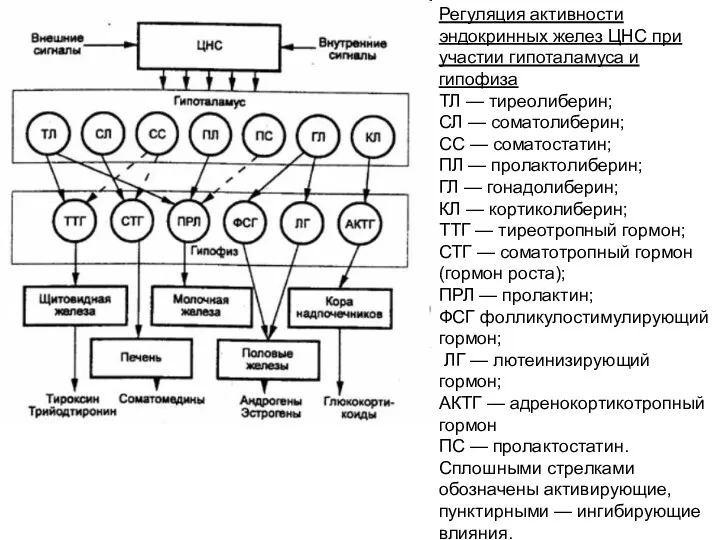
Регуляция активности эндокринных желез ЦНС при участии гипоталамуса и гипофиза
ТЛ —
Регуляция активности эндокринных желез ЦНС при участии гипоталамуса и гипофиза
ТЛ —
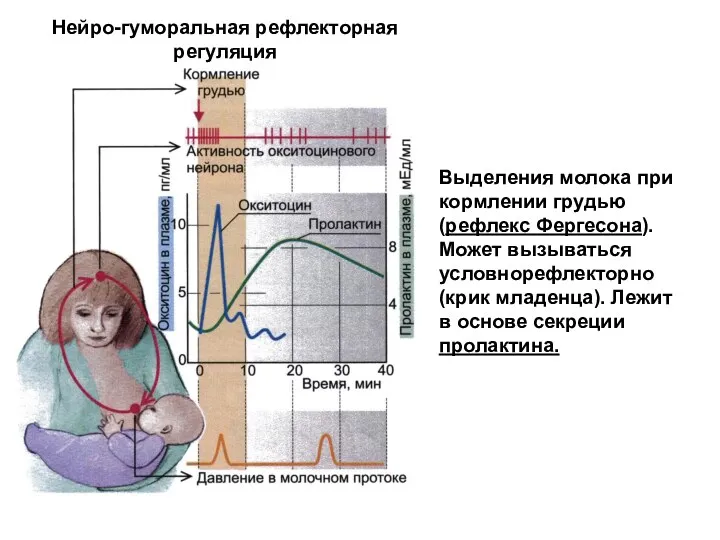
Нейро-гуморальная рефлекторная регуляция
Выделения молока при кормлении грудью (рефлекс Фергесона). Может вызываться
Нейро-гуморальная рефлекторная регуляция
Выделения молока при кормлении грудью (рефлекс Фергесона). Может вызываться
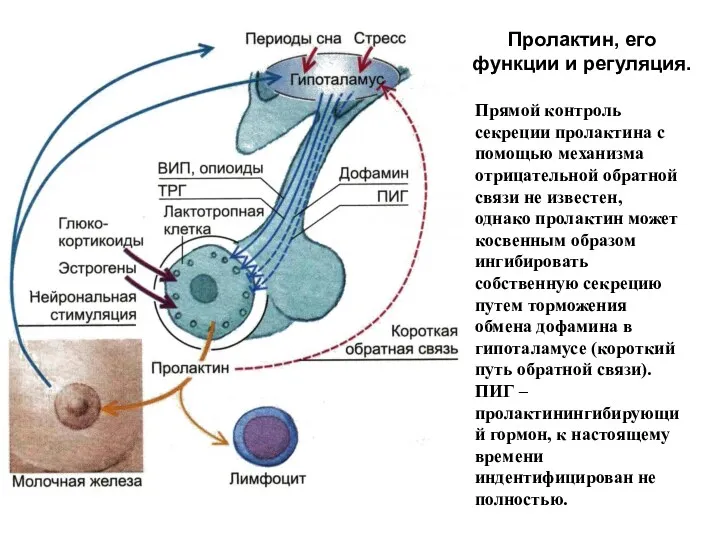
Пролактин, его функции и регуляция.
Прямой контроль секреции пролактина с помощью механизма
Пролактин, его функции и регуляция.
Прямой контроль секреции пролактина с помощью механизма
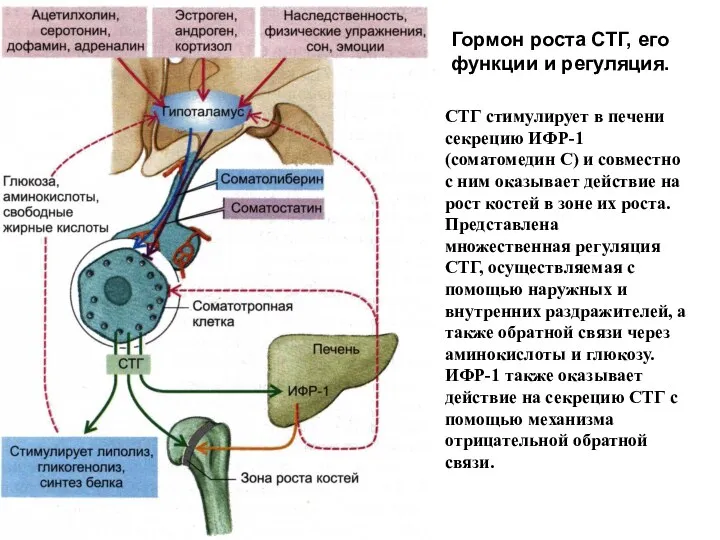
Гормон роста СТГ, его функции и регуляция.
СТГ стимулирует в печени
Гормон роста СТГ, его функции и регуляция.
СТГ стимулирует в печени
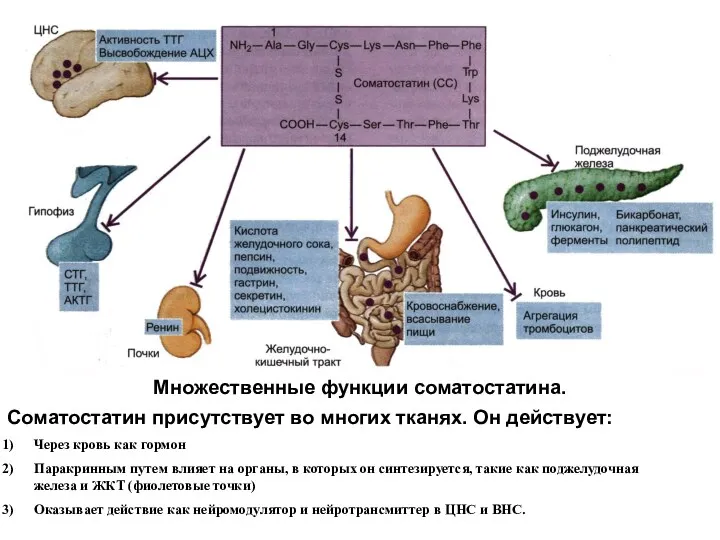
Множественные функции соматостатина.
Соматостатин присутствует во многих тканях. Он действует:
Через кровь
Множественные функции соматостатина.
Соматостатин присутствует во многих тканях. Он действует:
Через кровь
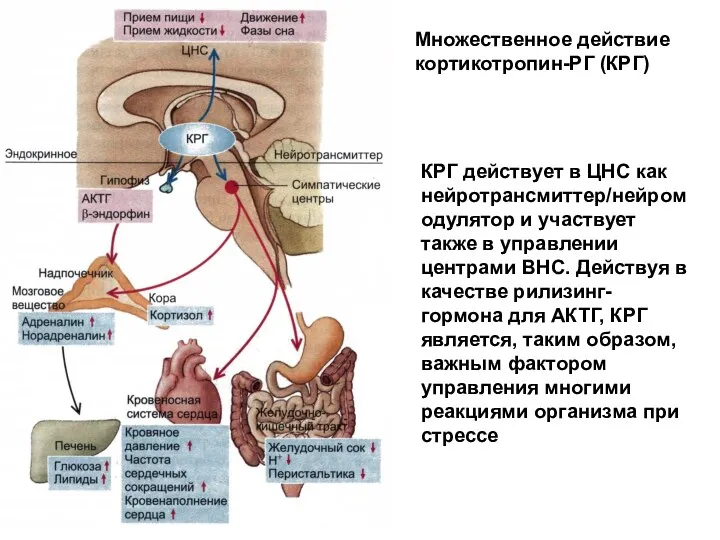
Множественное действие кортикотропин-РГ (КРГ)
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует
Множественное действие кортикотропин-РГ (КРГ)
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует
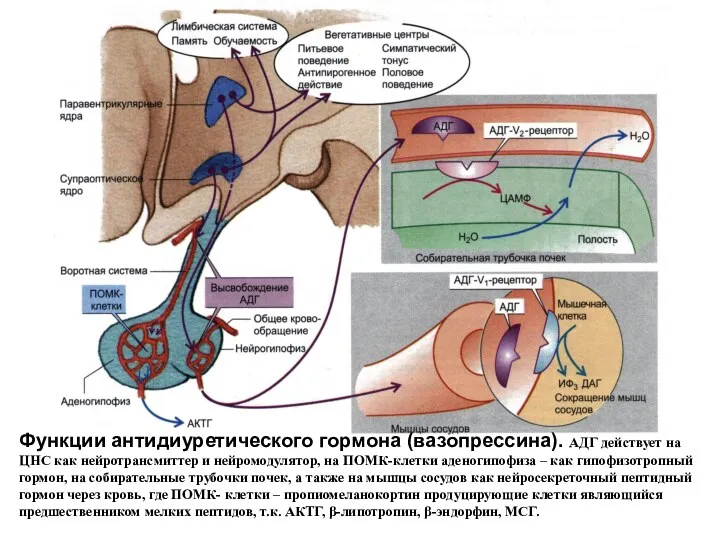
Функции антидиуретического гормона (вазопрессина). АДГ действует на ЦНС как нейротрансмиттер и
Функции антидиуретического гормона (вазопрессина). АДГ действует на ЦНС как нейротрансмиттер и
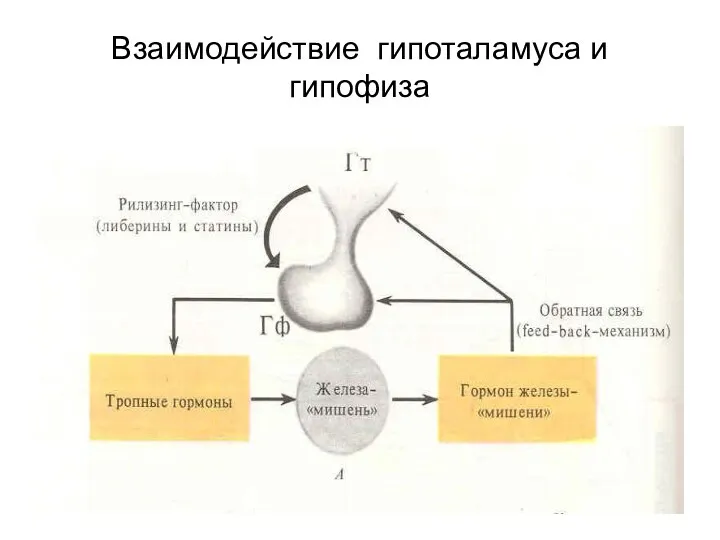
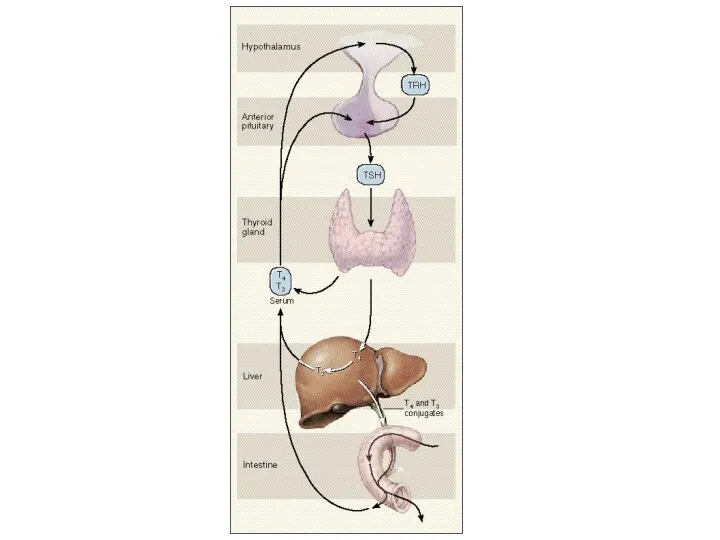
Взаимодействие гипоталамуса и гипофиза
Взаимодействие гипоталамуса и гипофиза
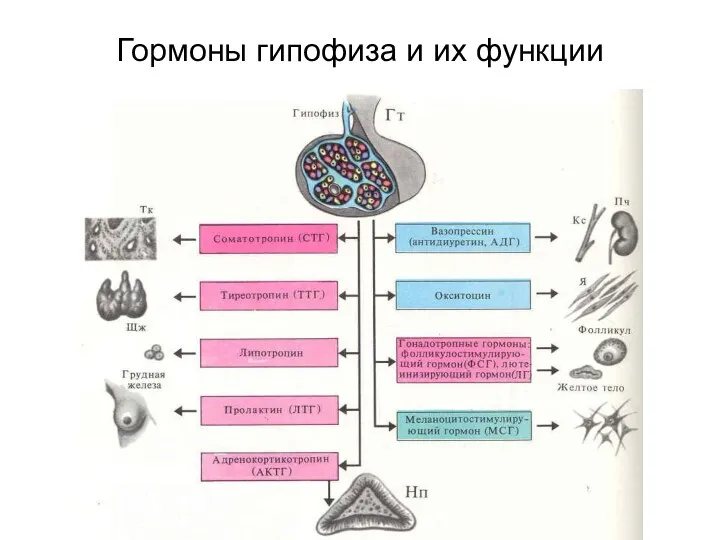
Гормоны гипофиза и их функции
Гормоны гипофиза и их функции

ЩИТОВИДНАЯ ЖЕЛЕЗА
ЩИТОВИДНАЯ ЖЕЛЕЗА
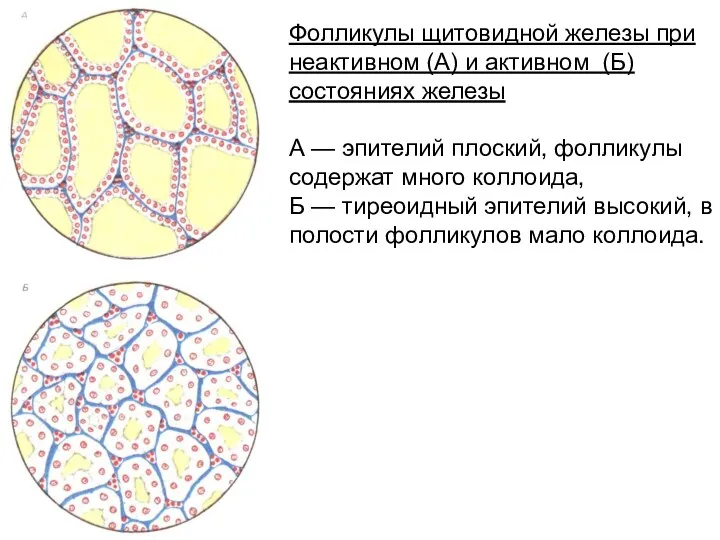
Фолликулы щитовидной железы при неактивном (А) и активном (Б) состояниях железы
А
Фолликулы щитовидной железы при неактивном (А) и активном (Б) состояниях железы
А
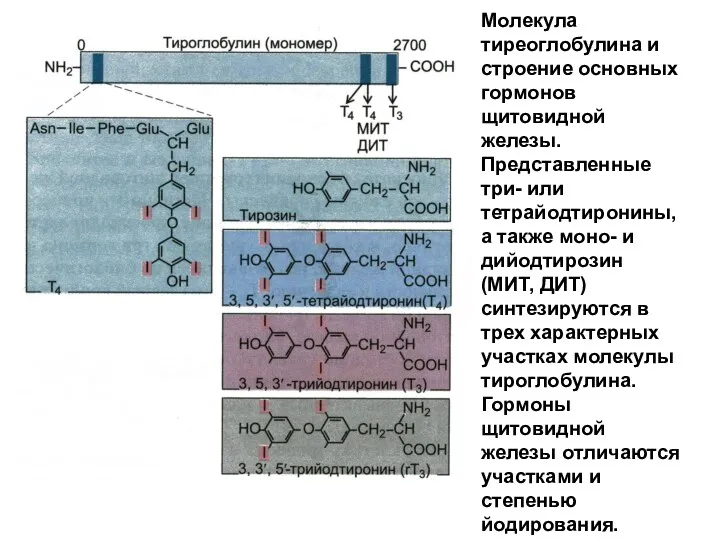
Молекула тиреоглобулина и строение основных гормонов щитовидной железы. Представленные три- или
Молекула тиреоглобулина и строение основных гормонов щитовидной железы. Представленные три- или

Уровень гормонов щитовидной железы и связывающих белков в плазме и функциональные
Уровень гормонов щитовидной железы и связывающих белков в плазме и функциональные
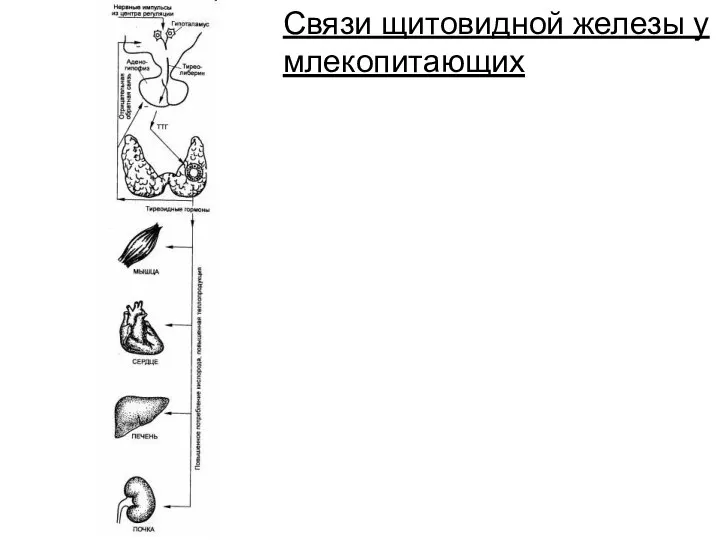
Связи щитовидной железы у млекопитающих
Связи щитовидной железы у млекопитающих
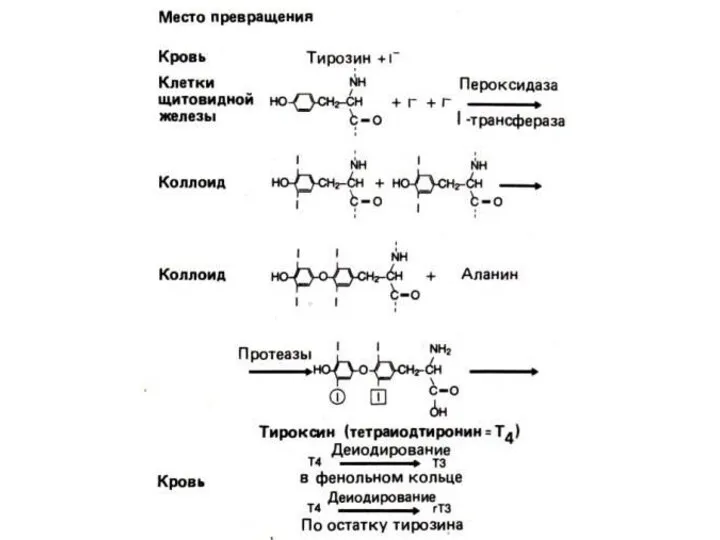
Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иоде покрывается
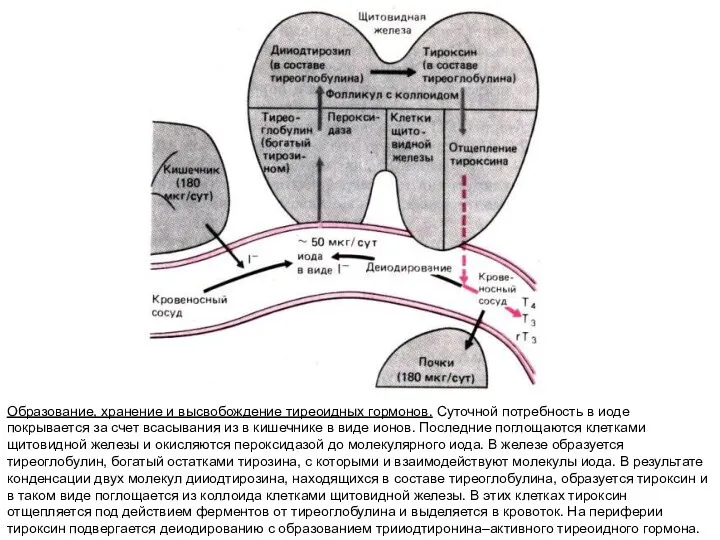
Образование, хранение и высвобождение тиреоидных гормонов. Cуточной потребность в иоде покрывается

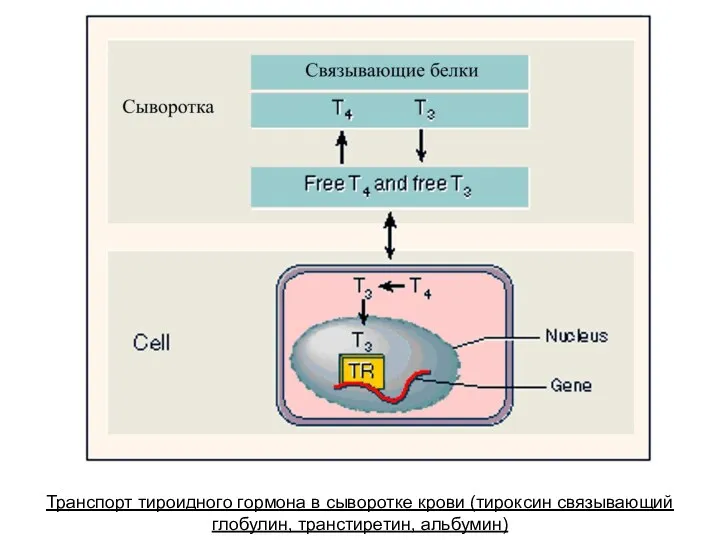
Транспорт тироидного гормона в сыворотке крови (тироксин связывающий глобулин, транстиретин, альбумин)
Транспорт тироидного гормона в сыворотке крови (тироксин связывающий глобулин, транстиретин, альбумин)
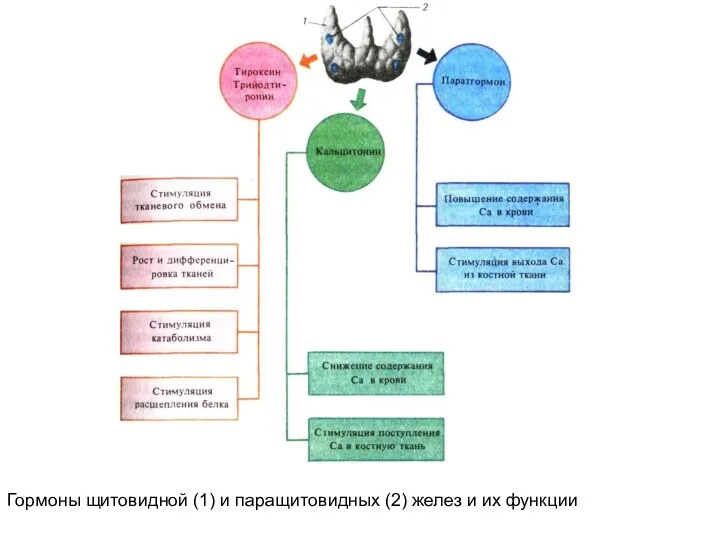
Гормоны щитовидной (1) и паращитовидных (2) желез и их функции
Гормоны щитовидной (1) и паращитовидных (2) желез и их функции
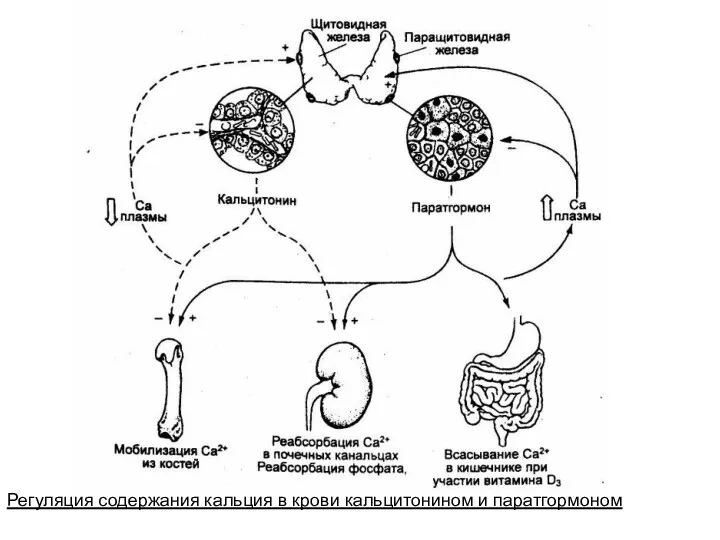
Регуляция содержания кальция в крови кальцитонином и паратгормоном
Регуляция содержания кальция в крови кальцитонином и паратгормоном
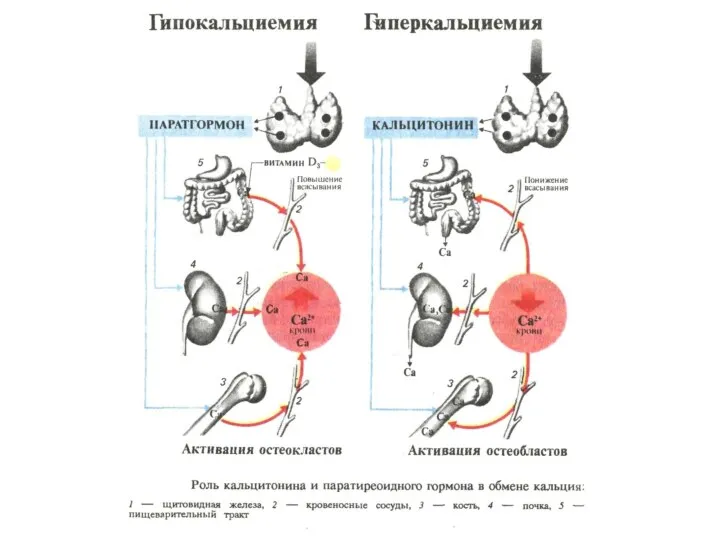

НАДПОЧЕЧНИКИ
НАДПОЧЕЧНИКИ
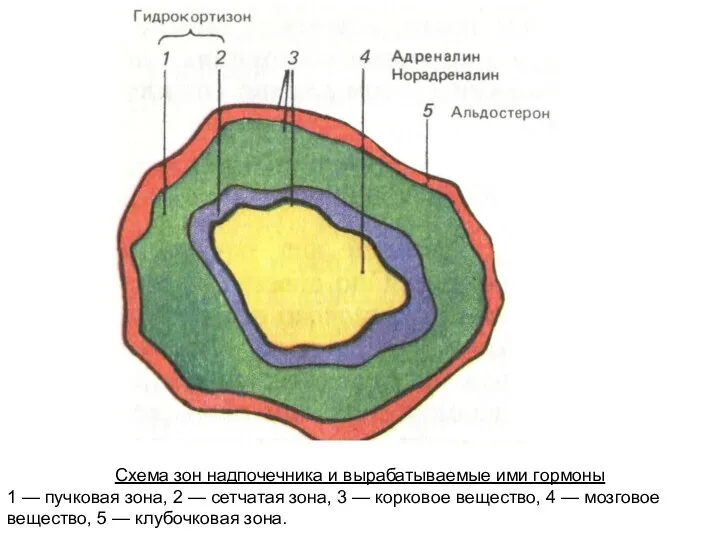
Схема зон надпочечника и вырабатываемые ими гормоны
1 — пучковая зона, 2
Схема зон надпочечника и вырабатываемые ими гормоны
1 — пучковая зона, 2
ГОРМОНЫ НАДПОЧЕЧНИКА
Кора надпочечника
Мозговое вещество
КОРТИЗОЛ
КОРТИКОСТЕРОН
АЛЬДОСТЕРОН
ДЕЗОКСИКОРТИКОСТЕРОН
ДЕГИДРОЭПИАНДРОСТЕРОН
АНДРОСТЕНДИОН
АДРЕНАЛИН
НОРАДРЕНАЛИН
ГОРМОНЫ НАДПОЧЕЧНИКА
Кора надпочечника
Мозговое вещество
КОРТИЗОЛ
КОРТИКОСТЕРОН
АЛЬДОСТЕРОН
ДЕЗОКСИКОРТИКОСТЕРОН
ДЕГИДРОЭПИАНДРОСТЕРОН
АНДРОСТЕНДИОН
АДРЕНАЛИН
НОРАДРЕНАЛИН
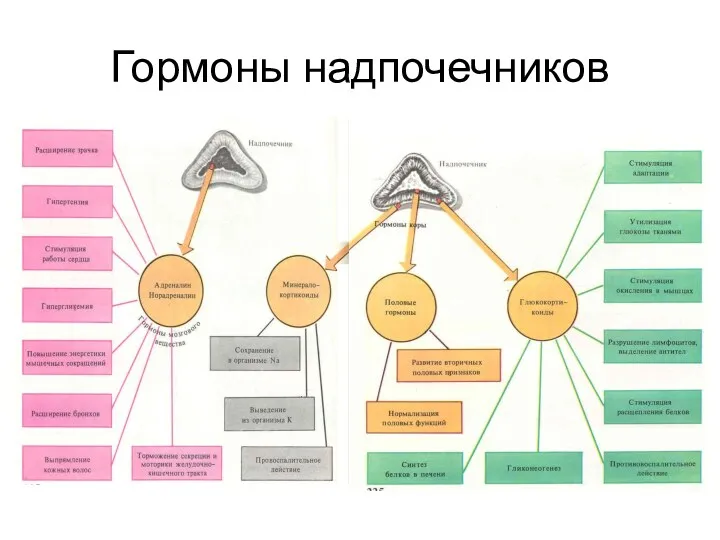
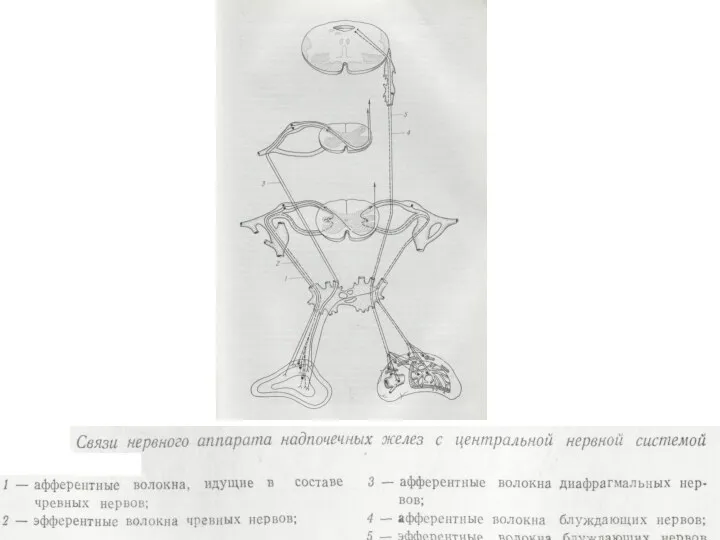
Гормоны надпочечников
Гормоны надпочечников

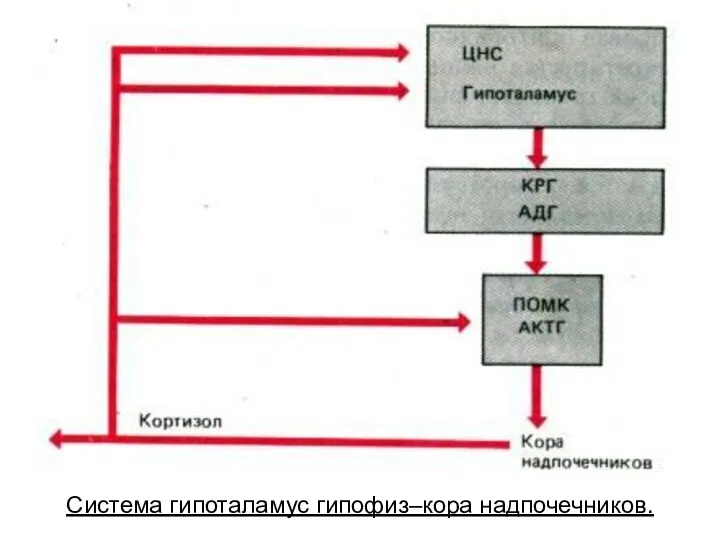
Система гипоталамус гипофиз–кора надпочечников.
Система гипоталамус гипофиз–кора надпочечников.
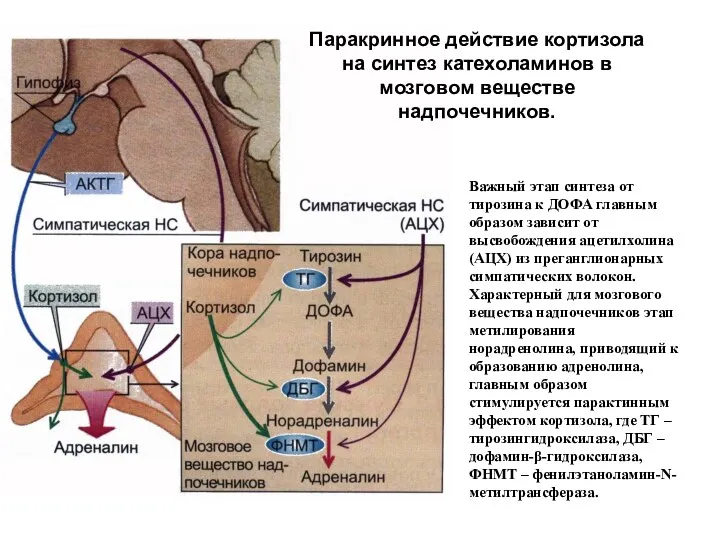
Паракринное действие кортизола на синтез катехоламинов в мозговом веществе надпочечников.
Важный этап
Паракринное действие кортизола на синтез катехоламинов в мозговом веществе надпочечников.
Важный этап
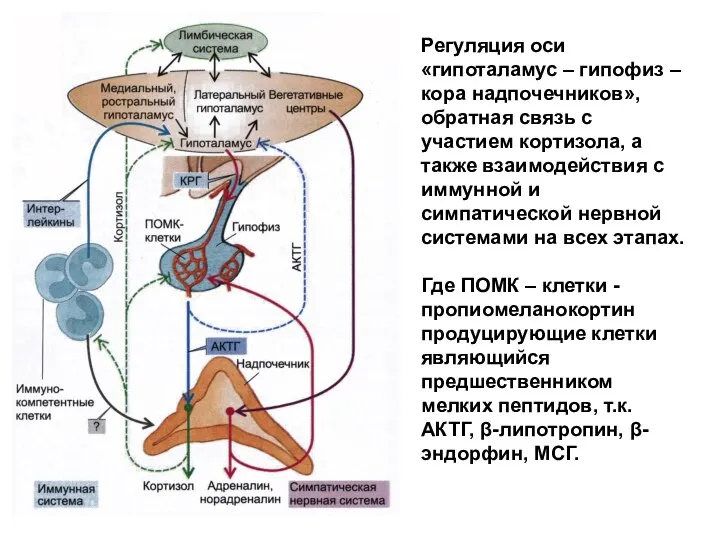
Регуляция оси «гипоталамус – гипофиз – кора надпочечников», обратная связь с
Регуляция оси «гипоталамус – гипофиз – кора надпочечников», обратная связь с
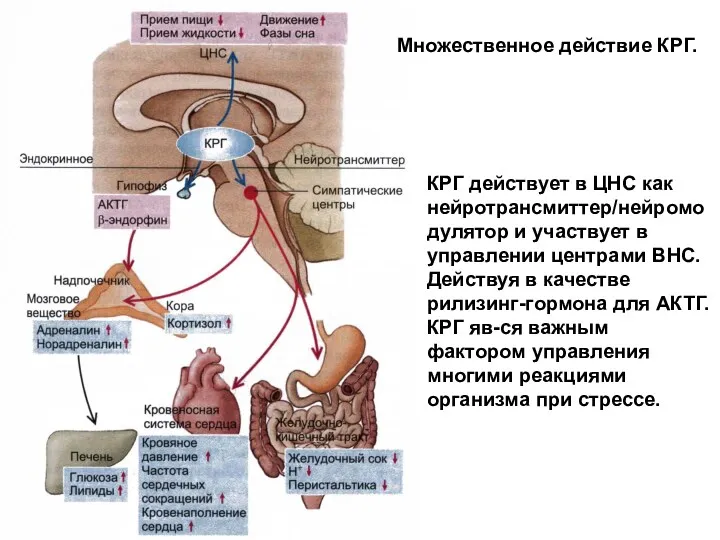
Множественное действие КРГ.
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует
Множественное действие КРГ.
КРГ действует в ЦНС как нейротрансмиттер/нейромодулятор и участвует

МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА
ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ
ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ
КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ
МОРФОЛОГИЧЕСКАЯ ТРИАДА СТРЕССА
ГИПЕРТРОФИЯ КОРЫ НАДПОЧЕЧНИКОВ
ИНВОЛЮЦИЯ ТИМУСА И ЛИМФАТИЧЕСКИХ УЗЛОВ
КРОВОИЗЛИЯНИЯ И ИЗЪЯЗВЛЕНИЕ
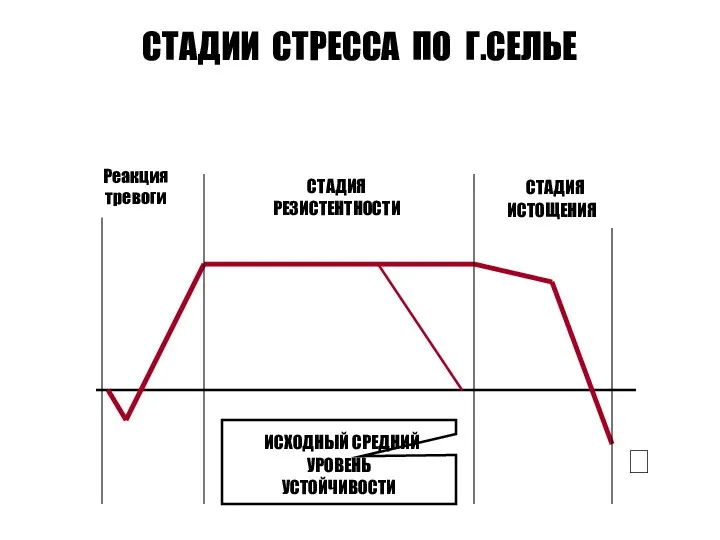
СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ
Реакция
тревоги
СТАДИЯ
РЕЗИСТЕНТНОСТИ
СТАДИЯ
ИСТОЩЕНИЯ
ИСХОДНЫЙ СРЕДНИЙ
УРОВЕНЬ
УСТОЙЧИВОСТИ
СТАДИИ СТРЕССА ПО Г.СЕЛЬЕ
Реакция
тревоги
СТАДИЯ
РЕЗИСТЕНТНОСТИ
СТАДИЯ
ИСТОЩЕНИЯ
ИСХОДНЫЙ СРЕДНИЙ
УРОВЕНЬ
УСТОЙЧИВОСТИ

ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
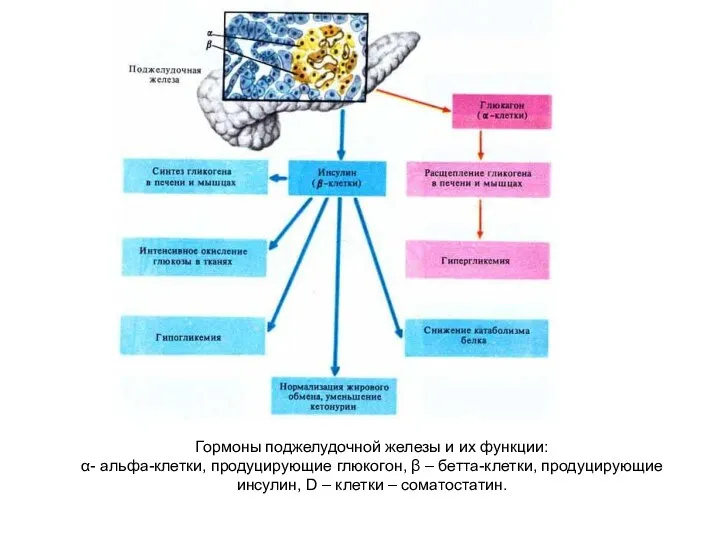
Гормоны поджелудочной железы и их функции:
α- альфа-клетки, продуцирующие глюкогон, β –
Гормоны поджелудочной железы и их функции: α- альфа-клетки, продуцирующие глюкогон, β –
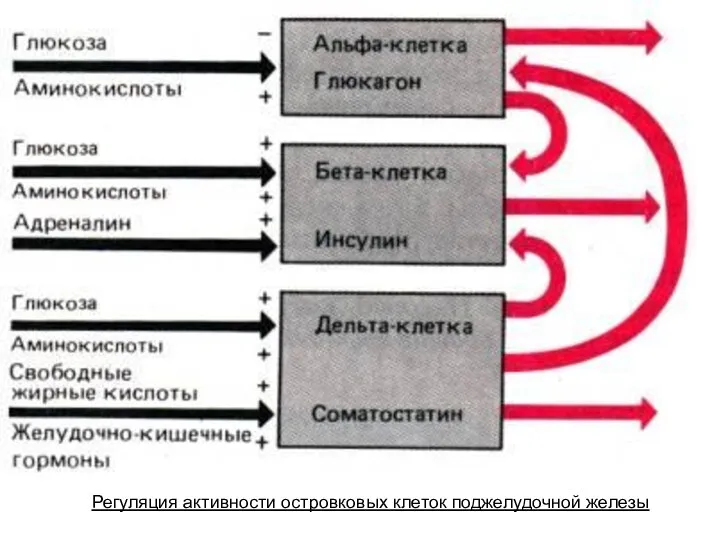
Регуляция активности островковых клеток поджелудочной железы
Регуляция активности островковых клеток поджелудочной железы

Спасибо
за ваше
внимание
Спасибо
за ваше
внимание
 Учёные, внёсшие вклад в развитие биологии с античности до наших дней
Учёные, внёсшие вклад в развитие биологии с античности до наших дней Отработка заданий части В и С ОГЭ. Биология 9 класс
Отработка заданий части В и С ОГЭ. Биология 9 класс Перелетные птицы весной. II младшая группа
Перелетные птицы весной. II младшая группа Скелет головы и туловища
Скелет головы и туловища Филэмбриогенез. Эволюционные преобразования
Филэмбриогенез. Эволюционные преобразования Клеточная теория
Клеточная теория Тип молюски. Клас двостулкові
Тип молюски. Клас двостулкові Природоохрана и защита живой природы
Природоохрана и защита живой природы Урок биологии 9 класс Звёздный час – Человек
Урок биологии 9 класс Звёздный час – Человек Задания ЕГЭ по биологии теме Формы достижения биологического прогресса
Задания ЕГЭ по биологии теме Формы достижения биологического прогресса Азотное питание растений
Азотное питание растений Разнообразие животных
Разнообразие животных Мужской ликбез. Все об уходе за внешностью
Мужской ликбез. Все об уходе за внешностью Пищеварительные железы
Пищеварительные железы Заочная экологическая викторина Чудо в перьях, 1 класс
Заочная экологическая викторина Чудо в перьях, 1 класс Класс Птицы
Класс Птицы Отруйнi гриби
Отруйнi гриби Движение крови по сосудам. Большой и малый круги кровообращения. 8 класс
Движение крови по сосудам. Большой и малый круги кровообращения. 8 класс Ткани растений
Ткани растений Химический состав клетки - технологическая карта урока.
Химический состав клетки - технологическая карта урока. Пропорции выражают характер. Птицы с разным характером пропорций
Пропорции выражают характер. Птицы с разным характером пропорций Предмет і завдання мікробіології. Основні етапи розвитку. Систематика номенклатура і класифікація мікроорганізмів
Предмет і завдання мікробіології. Основні етапи розвитку. Систематика номенклатура і класифікація мікроорганізмів Сезонные изменения в жизни организмов
Сезонные изменения в жизни организмов Анатомия глаза. Веки
Анатомия глаза. Веки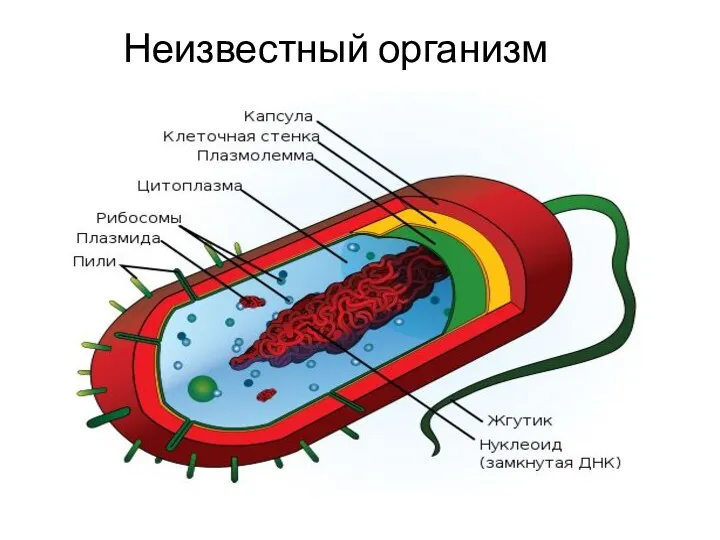 Бактерии. Строение бактериальной клетки. Формы бактерий. Распространение и условия обитания
Бактерии. Строение бактериальной клетки. Формы бактерий. Распространение и условия обитания Протерозойская эра
Протерозойская эра Клеточный сигналинг
Клеточный сигналинг Отряд Непарнокопытные
Отряд Непарнокопытные