- Организация древесины на клеточном уровне. Лекция 4

Содержание
- 2. Для чего изучать клеточное строение ?
- 3. При изучении строения древесины на клеточном уровне — микроскопического строения, тонкие срезы древесины (толщиной 15-20 мкм)
- 4. Знание основ клеточного строения древесины крайне важно во многих областях древесиноведения. Микроскопическое строение является базой для
- 5. 1 Особенности клеточного строения древесины
- 6. Как и все растения, древесные растения имеют клеточное строение. Любая клетка растения состоит из оболочки и
- 7. У древесных растений различают первичную и вторичную ксилему. Первичная ксилема образуется дифференциацией клеток специализированной меристемы —
- 8. 2 Камбий и его функции в растущем дереве
- 9. Образование всех новых клеток древесины происходит из камбиальных клеток. Поскольку камбиальные клетки в древесине расположены непрерывным
- 10. Камбиальные клетки, составляющие образовательную ткань древесного растения, располагаются непрерывным слоем толщиной в одну клетку (камбиальную инициаль)
- 11. Тонкостенные камбиальные клетки остаются живыми на протяжении всей жизни дерева (следовательно, не исключено, что отдельные клетки
- 12. Камбиальные клетки могут быть двух типов: веретеновидные инициали — вытянутые камбиальные клетки, образующие основную массу клеток
- 13. Размеры инициалей могут значительно различаться у разных древесных пород. так, у Pinus strobus длина веретеновидных инициалей
- 14. Размер камбиальных инициалей даже одного типа не остается постоянным в течение жизни дерева (табл. 25). В
- 15. Схема строения камбиальных слоев: (А) А — инициальная веретеновидная клетка камбия; пунктиром показана плоскость следующего деления;
- 16. Схема строения камбиальных слоев: (Б) Б — схематичное изображение последовательности образования новых клеток флоэмы (в, е)
- 17. Схема строения камбиальных слоев: (В, Г) В — неярусный камбий (грецкий орех, тангенциальный разрез); Г —
- 18. Взаимное расположение камбиальных инициалей также может быть разным — ярусным или неярусным (рис. 26 В, Г),
- 19. 3 Этапы образования и роста новых клеток древесины
- 20. 1 Этап
- 21. Образование новых клеток в древесине сводится к нескольким этапам. Первым этапом является деление материнской клетки камбия.
- 22. По мере увеличения длины окружности ствола в процессе роста дерева, происходят мультипликативные (антиклинальные) деления (в тангенциальном
- 23. Образование новых лучевых инициалей, клеток древесной паренхимы и клеток, образующих вертикальные смоляные ходы, происходит в результате
- 24. Основной характеристикой клеточных делений является продолжительность клеточного цикла, равная промежутку времени между двумя последовательными делениями камбиальной
- 25. На количество образовавшихся клеток кроме продолжительности клеточного цикла большое влияние оказывает продолжительность периода делений в течение
- 26. 2 Этап
- 27. второй этап (фаза) образования клеток — рост растяжением. В начале этой стадии клетки обладают протопластом и
- 28. Растяжение каждой клетки обычно наиболее выражено в каком-то одном направлении. Производные лучевых инициалей претерпевают сравнительно небольшие
- 29. Клетки, развивающиеся в членики крупных сосудов, в продольном направлении могут не увеличиваться вовсе или увеличиваются незначительно.
- 30. В некоторых сосудах отдельные членики, наоборот, сильно вытянуты при относительно небольших поперечных размерах; членики таких сосудов
- 31. Размеры трахеид хвойных, например, в тангенциальном направлении почти не изменяются, вдоль волокон увеличиваются на 20% (небольшое
- 32. У лиственных пород трахеиды больше увеличиваются в поперечных направлениях, а волокна либриформа — в продольном. Окончательный
- 33. К концу этого этапа роста клетка приобретает окончательные размеры, но все еще обладает тонкой (первичной) оболочкой.
- 34. 3 Этап
- 35. третий этап развития клеток — фаза вторичного утолщения. Основная масса древесинного вещества откладывается в фазу вторичного
- 36. У некоторых пород вторичные оболочки образуются и в паренхимных клетках. Однако такие клетки не отмирают сразу
- 37. Окончательная толщина клеточной оболочки определяется не скоростью утолщения, а продолжительностью этого периода. У хвойных эта стадия
- 38. Лигнификация начинается в первичной оболочке, охватывает срединную пластинку и затем распространяется на вторичную оболочку. В результате
- 39. Заканчивается эта фаза для большинства клеток древесины отмиранием протопласта и образованием из его остатков бородавчатого слоя,
- 40. Апоптоз в трахеидах определяется экспрессией генов, контролирующих синтез гидролаз. Активность гидролаз приводит к распаду тонопласта (вакуолярной
- 41. Одновременно с завершением формирования утолщения тонопласт теряет целостность, остатки цитоплазмы и нелигнифицированные участки первичной оболочки разрушаются
- 42. 4 Основные типы клеток древесины
- 43. По типу все клетки древесины делятся на две большие группы — паренхимные и прозенхимные.
- 44. 4.1 Паренхимные клетки
- 45. Паренхимные клетки — это большей частью тонкостенные клетки, имеющие в различных направлениях примерно равную длину —
- 46. Однако известны породы, древесина которых содержит гораздо больше паренхимных клеток или даже в основном состоит из
- 47. В растущем дереве паренхимные клетки выполняют множественную функцию, участвуя тем или иным образом во всех физиологических
- 48. Паренхимные клетки являются физиологически активными клетками, обеспечивают иммунитет живой древесины (защитная функция) и определяют ее биостойкость
- 49. Паренхимным клеткам присуща и проводящая функция, поскольку в их оболочках имеются поры. Однако они проводят не
- 50. Паренхимные клетки, безусловно, выполняют и механическую функцию, особенно после формирования у них вторичной оболочки. Наиболее ярко
- 51. 4.2 Прозенхимные клетки
- 52. Второй тип клеток — прозенхимные клетки, или волокна, сильно вытянуты в одном направлении и могут достигать
- 53. Рис.27 Основные клетки древесины и луба хвойных пород, производные веретеновидных инициалей Поздняя трахеида Ранняя трахеида Тяжевая
- 54. Рис.27 Основные клетки древесины и луба лиственных пород, производные веретеновидных инициалей волокно либриформа волокнистая трахеида сосудистая
- 55. 5 Живые и мертвые клетки древесины
- 56. Непосредственно после деления камбиальных клеток все молодые вновь образованные клетки древесины (трахеиды, волокна, сосуды) являются живыми
- 57. Как уже отмечалось, при отмирании живые клетки древесины утрачивают протопласт, а их полости заполняются водой, и
- 58. В физиологически активной части ствола — в древесине заболони, среди подавляющей массы мертвых клеток имеются относительно
- 59. Распределение живых клеток по радиусу ствола у различных древесных пород разное: у заболонных пород их количество
- 60. Рис.28 Характер распределения живых паренхимных клеток по радиусу ствола
- 61. Рис. 28. Характер распределения живых паренхимных клеток по радиусу ствола
- 62. Живые паренхимные клетки, соединяясь друг с другом, образуют единую сложную сеть, распространяющуюся среди жестких каркасов оболочек
- 63. Живые паренхимные клетки древесины — это особые клетки. Они заключены в жесткие мертвые ткани основной массы
- 64. Содержимое живых клеток — клеточный сок, обычно имеет кислую реакцию (рН = 4-5); величина рН клеточного
- 65. После рубки дерева камбий и паренхимные клетки древесины и луба отмирают не сразу. Пока сохраняется достаточное
- 66. 6 Способы сообщения между полостями соседних клеток древесины
- 67. Оболочки всех клеток древесины плотно «подогнаны» друг к другу и крепко соединены межклеточным веществом. Но размеры
- 68. рис. 29. Схематичное изображение типов пор: А — пара окаймленных пор в трахеидах хвойных пород: 1
- 69. Существуют различные типы отверстий в клеточных оболочках. Наиболее распространенным типом отверстий в клеточных оболочках являются поры,
- 70. Различают простые (всегда открытые) и окаймленные (сложные, закрывающиеся) поры, которые могут иметь округлые и щелевидные отверстия
- 71. рис. 30. Окаймленная пора хвойных: А — вид на радиальной стороне оболочки; В — в разрезе:
- 72. Мембрана простых пор может иметь отверстия или вовсе отсутствовать. Само отверстие поры может быть различной формы;
- 74. Скачать презентацию

Для чего изучать клеточное строение ?
Для чего изучать клеточное строение ?

При изучении строения древесины на клеточном уровне
— микроскопического строения, тонкие срезы
При изучении строения древесины на клеточном уровне
— микроскопического строения, тонкие срезы

Знание основ клеточного строения древесины
крайне важно во многих областях древесиноведения.
Микроскопическое
Знание основ клеточного строения древесины
крайне важно во многих областях древесиноведения.
Микроскопическое

1 Особенности клеточного строения древесины
1 Особенности клеточного строения древесины

Как и все растения,
древесные растения имеют клеточное строение.
Любая клетка растения
Как и все растения,
древесные растения имеют клеточное строение.
Любая клетка растения

У древесных растений различают первичную и вторичную ксилему.
Первичная ксилема образуется дифференциацией
У древесных растений различают первичную и вторичную ксилему.
Первичная ксилема образуется дифференциацией

2 Камбий и его функции в растущем дереве
2 Камбий и его функции в растущем дереве

Образование всех новых клеток древесины
происходит из камбиальных клеток.
Поскольку камбиальные клетки
Образование всех новых клеток древесины
происходит из камбиальных клеток.
Поскольку камбиальные клетки

Камбиальные клетки,
составляющие образовательную ткань древесного растения, располагаются непрерывным слоем толщиной в
Камбиальные клетки,
составляющие образовательную ткань древесного растения, располагаются непрерывным слоем толщиной в

Тонкостенные камбиальные клетки
остаются живыми на протяжении всей жизни дерева
(следовательно, не
Тонкостенные камбиальные клетки
остаются живыми на протяжении всей жизни дерева
(следовательно, не

Камбиальные клетки могут быть двух типов:
веретеновидные инициали — вытянутые камбиальные клетки,
Камбиальные клетки могут быть двух типов:
веретеновидные инициали — вытянутые камбиальные клетки,

Размеры инициалей могут значительно различаться
у разных древесных пород. так, у Pinus
Размеры инициалей могут значительно различаться
у разных древесных пород. так, у Pinus

Размер камбиальных инициалей
даже одного типа не остается постоянным в течение жизни
Размер камбиальных инициалей
даже одного типа не остается постоянным в течение жизни

Схема строения камбиальных слоев: (А)
А — инициальная веретеновидная клетка камбия; пунктиром
Схема строения камбиальных слоев: (А)
А — инициальная веретеновидная клетка камбия; пунктиром

Схема строения камбиальных слоев: (Б)
Б — схематичное изображение последовательности образования новых
Схема строения камбиальных слоев: (Б)
Б — схематичное изображение последовательности образования новых

Схема строения камбиальных слоев: (В, Г)
В — неярусный камбий (грецкий орех,
Схема строения камбиальных слоев: (В, Г)
В — неярусный камбий (грецкий орех,

Взаимное расположение камбиальных инициалей
также может быть разным — ярусным или неярусным
Взаимное расположение камбиальных инициалей
также может быть разным — ярусным или неярусным

3 Этапы образования и роста новых клеток древесины
3 Этапы образования и роста новых клеток древесины

1 Этап
1 Этап

Образование новых клеток в древесине сводится к нескольким этапам.
Первым этапом
Образование новых клеток в древесине сводится к нескольким этапам.
Первым этапом

По мере увеличения длины окружности ствола
в процессе роста дерева, происходят мультипликативные
По мере увеличения длины окружности ствола
в процессе роста дерева, происходят мультипликативные

Образование новых лучевых инициалей,
клеток древесной паренхимы и клеток, образующих вертикальные смоляные
Образование новых лучевых инициалей,
клеток древесной паренхимы и клеток, образующих вертикальные смоляные

Основной характеристикой клеточных делений
является продолжительность клеточного цикла, равная промежутку времени между
Основной характеристикой клеточных делений
является продолжительность клеточного цикла, равная промежутку времени между

На количество образовавшихся клеток
кроме продолжительности клеточного цикла большое влияние оказывает продолжительность
На количество образовавшихся клеток
кроме продолжительности клеточного цикла большое влияние оказывает продолжительность

2 Этап
2 Этап

второй этап (фаза) образования клеток
— рост растяжением.
В начале этой стадии
второй этап (фаза) образования клеток
— рост растяжением.
В начале этой стадии

Растяжение каждой клетки обычно наиболее выражено в каком-то одном направлении.
Производные лучевых
Растяжение каждой клетки обычно наиболее выражено в каком-то одном направлении.
Производные лучевых

Клетки, развивающиеся в членики крупных сосудов,
в продольном направлении могут не увеличиваться
Клетки, развивающиеся в членики крупных сосудов,
в продольном направлении могут не увеличиваться

В некоторых сосудах отдельные членики,
наоборот, сильно вытянуты при относительно небольших поперечных
В некоторых сосудах отдельные членики,
наоборот, сильно вытянуты при относительно небольших поперечных

Размеры трахеид хвойных,
например, в тангенциальном направлении почти не изменяются,
вдоль волокон
Размеры трахеид хвойных,
например, в тангенциальном направлении почти не изменяются,
вдоль волокон

У лиственных пород
трахеиды больше увеличиваются в поперечных направлениях, а волокна либриформа
У лиственных пород
трахеиды больше увеличиваются в поперечных направлениях, а волокна либриформа

К концу этого этапа роста
клетка приобретает окончательные размеры,
но все еще
К концу этого этапа роста
клетка приобретает окончательные размеры,
но все еще

3 Этап
3 Этап

третий этап развития клеток
— фаза вторичного утолщения.
Основная масса древесинного вещества
третий этап развития клеток
— фаза вторичного утолщения.
Основная масса древесинного вещества

У некоторых пород вторичные оболочки образуются
и в паренхимных клетках.
Однако такие
У некоторых пород вторичные оболочки образуются
и в паренхимных клетках.
Однако такие

Окончательная толщина клеточной оболочки
определяется не скоростью утолщения, а продолжительностью этого периода.
Окончательная толщина клеточной оболочки
определяется не скоростью утолщения, а продолжительностью этого периода.

Лигнификация начинается в первичной оболочке,
охватывает срединную пластинку и затем распространяется на
Лигнификация начинается в первичной оболочке,
охватывает срединную пластинку и затем распространяется на

Заканчивается эта фаза для большинства клеток древесины
отмиранием протопласта и образованием из
Заканчивается эта фаза для большинства клеток древесины
отмиранием протопласта и образованием из

Апоптоз в трахеидах определяется экспрессией генов,
контролирующих синтез гидролаз.
Активность гидролаз приводит
Апоптоз в трахеидах определяется экспрессией генов,
контролирующих синтез гидролаз.
Активность гидролаз приводит

Одновременно с завершением формирования утолщения
тонопласт теряет целостность, остатки цитоплазмы и нелигнифицированные
Одновременно с завершением формирования утолщения
тонопласт теряет целостность, остатки цитоплазмы и нелигнифицированные

4 Основные типы клеток древесины
4 Основные типы клеток древесины

По типу
все клетки древесины делятся на две большие группы
— паренхимные
По типу
все клетки древесины делятся на две большие группы
— паренхимные

4.1 Паренхимные клетки
4.1 Паренхимные клетки

Паренхимные клетки
— это большей частью тонкостенные клетки, имеющие в различных направлениях
Паренхимные клетки
— это большей частью тонкостенные клетки, имеющие в различных направлениях

Однако известны породы,
древесина которых содержит гораздо больше паренхимных клеток или даже
Однако известны породы,
древесина которых содержит гораздо больше паренхимных клеток или даже

В растущем дереве паренхимные клетки выполняют множественную функцию,
участвуя тем или иным
В растущем дереве паренхимные клетки выполняют множественную функцию,
участвуя тем или иным

Паренхимные клетки являются физиологически активными клетками,
обеспечивают иммунитет живой древесины (защитная функция)
Паренхимные клетки являются физиологически активными клетками,
обеспечивают иммунитет живой древесины (защитная функция)

Паренхимным клеткам присуща и проводящая функция,
поскольку в их оболочках имеются поры.
Паренхимным клеткам присуща и проводящая функция,
поскольку в их оболочках имеются поры.

Паренхимные клетки, безусловно, выполняют и механическую функцию,
особенно после формирования у них
Паренхимные клетки, безусловно, выполняют и механическую функцию,
особенно после формирования у них

4.2 Прозенхимные клетки
4.2 Прозенхимные клетки

Второй тип клеток — прозенхимные клетки, или волокна,
сильно вытянуты в одном
Второй тип клеток — прозенхимные клетки, или волокна,
сильно вытянуты в одном
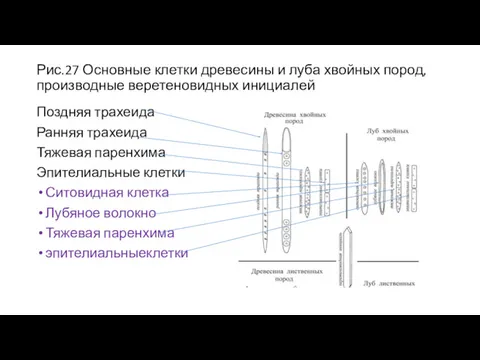
Рис.27 Основные клетки древесины и луба хвойных пород, производные веретеновидных инициалей
Поздняя
Рис.27 Основные клетки древесины и луба хвойных пород, производные веретеновидных инициалей
Поздняя
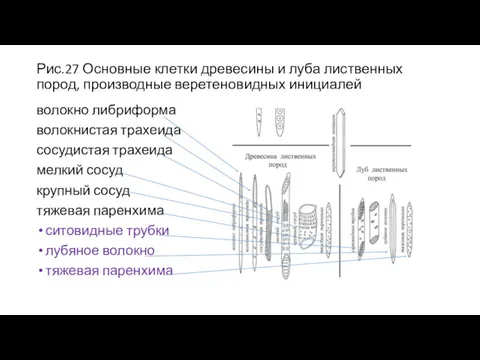
Рис.27 Основные клетки древесины и луба лиственных пород, производные веретеновидных инициалей
волокно
Рис.27 Основные клетки древесины и луба лиственных пород, производные веретеновидных инициалей
волокно

5 Живые и мертвые клетки древесины
5 Живые и мертвые клетки древесины

Непосредственно после деления камбиальных клеток
все молодые вновь образованные клетки древесины (трахеиды,
Непосредственно после деления камбиальных клеток
все молодые вновь образованные клетки древесины (трахеиды,

Как уже отмечалось, при отмирании живые клетки древесины
утрачивают протопласт, а их
Как уже отмечалось, при отмирании живые клетки древесины
утрачивают протопласт, а их

В физиологически активной части ствола
— в древесине заболони, среди подавляющей массы
В физиологически активной части ствола
— в древесине заболони, среди подавляющей массы

Распределение живых клеток по радиусу ствола
у различных древесных пород разное:
у
Распределение живых клеток по радиусу ствола
у различных древесных пород разное:
у
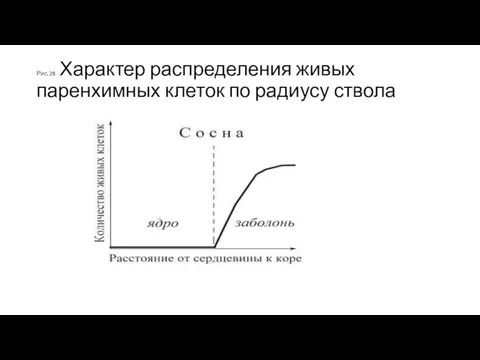
Рис.28 Характер распределения живых паренхимных клеток по радиусу ствола
Рис.28 Характер распределения живых паренхимных клеток по радиусу ствола
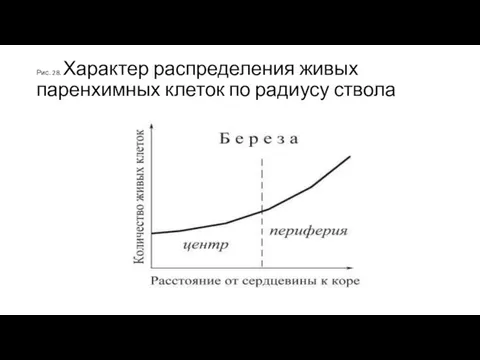
Рис. 28. Характер распределения живых паренхимных клеток по радиусу ствола
Рис. 28. Характер распределения живых паренхимных клеток по радиусу ствола

Живые паренхимные клетки,
соединяясь друг с другом, образуют единую сложную сеть, распространяющуюся
Живые паренхимные клетки,
соединяясь друг с другом, образуют единую сложную сеть, распространяющуюся

Живые паренхимные клетки древесины
— это особые клетки.
Они заключены в жесткие
Живые паренхимные клетки древесины
— это особые клетки.
Они заключены в жесткие

Содержимое живых клеток
— клеточный сок, обычно имеет кислую реакцию (рН =
Содержимое живых клеток
— клеточный сок, обычно имеет кислую реакцию (рН =

После рубки дерева камбий и паренхимные клетки древесины и луба отмирают
После рубки дерева камбий и паренхимные клетки древесины и луба отмирают

6 Способы сообщения между полостями соседних клеток древесины
6 Способы сообщения между полостями соседних клеток древесины

Оболочки всех клеток древесины
плотно «подогнаны» друг к другу и крепко соединены
Оболочки всех клеток древесины
плотно «подогнаны» друг к другу и крепко соединены

рис. 29. Схематичное изображение типов пор:
А — пара окаймленных пор в
рис. 29. Схематичное изображение типов пор: А — пара окаймленных пор в

Существуют различные типы отверстий в клеточных оболочках.
Наиболее распространенным типом отверстий в
Существуют различные типы отверстий в клеточных оболочках.
Наиболее распространенным типом отверстий в

Различают
простые (всегда открытые)
и окаймленные (сложные, закрывающиеся) поры,
которые могут иметь
Различают
простые (всегда открытые)
и окаймленные (сложные, закрывающиеся) поры,
которые могут иметь
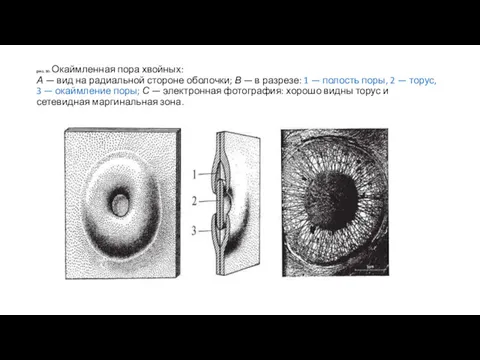
рис. 30. Окаймленная пора хвойных:
А — вид на радиальной стороне оболочки;
рис. 30. Окаймленная пора хвойных: А — вид на радиальной стороне оболочки;

Мембрана простых пор может иметь отверстия или вовсе отсутствовать.
Само отверстие поры
Мембрана простых пор может иметь отверстия или вовсе отсутствовать.
Само отверстие поры
 Николай Федорович Гамалея
Николай Федорович Гамалея Cells
Cells Строение синапсов. Потенциал действия. Механизм проведения потенциала действия. Физиология боли, медиаторы боли
Строение синапсов. Потенциал действия. Механизм проведения потенциала действия. Физиология боли, медиаторы боли Всасывание, пищеварение
Всасывание, пищеварение Процесс кровообращения. Сердце, анатомия и физиология
Процесс кровообращения. Сердце, анатомия и физиология презентация к уроку биологии 7 класс
презентация к уроку биологии 7 класс Рост и развитие животных
Рост и развитие животных Строение эукариотической клетки
Строение эукариотической клетки Урок биологии в 9 классе по теме Антропогенез
Урок биологии в 9 классе по теме Антропогенез Домашние любимцы. Кошки и собаки
Домашние любимцы. Кошки и собаки Ознаки захворювання яблук
Ознаки захворювання яблук Пищевая микробиология. Молочные продукты
Пищевая микробиология. Молочные продукты Элективный курс Репродуктивное здоровье
Элективный курс Репродуктивное здоровье Отдел голосеменные
Отдел голосеменные Рыбы Байкала
Рыбы Байкала Тип Інфузорії
Тип Інфузорії Растения Дальнего Востока
Растения Дальнего Востока Полезный мед - натуральный мед
Полезный мед - натуральный мед Цитологические основы наследования признаков при моногибридном скрещивании
Цитологические основы наследования признаков при моногибридном скрещивании Моллюски осьминоги
Моллюски осьминоги Nervous sistem
Nervous sistem Домашние питомцы
Домашние питомцы Роль генетически модифицированных организмов на организм человека
Роль генетически модифицированных организмов на организм человека Молекулярно-биологические методы диагностики
Молекулярно-биологические методы диагностики Птицы
Птицы Витамины в составе фруктов и овощей
Витамины в составе фруктов и овощей Голосеменные растения
Голосеменные растения Возникновение жизни
Возникновение жизни