- Организация хромосом и биологические функции ДНК

Содержание
- 2. Организация эукариотических хромосом Эукариотическая хромосома содержит длинные, линейные молекулы ДНК, которые должны разделяться (сегрегации), размножаться и
- 3. У эукариот ген, как правило, дискретен, то есть, разделен на участки интронами. Одна хромосома, как правило,
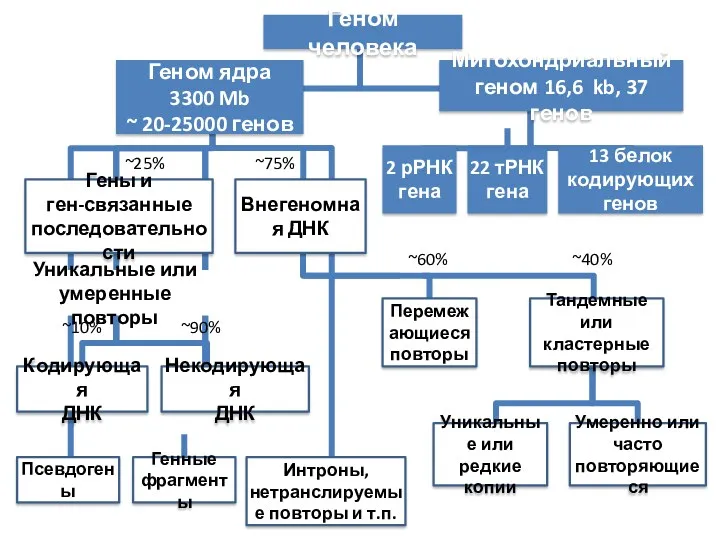
- 4. Уникальные (неповторяющиеся последовательности – в геноме встречаются один или несколько раз. Содержат структурные гены, а также
- 5. Биологическая роль ДНК повторов ДНК повторы обеспечивают разнообразие архитектоники генома и хромосом как основных участников морфогенеза.
- 6. Терминология Транскрипто́м (англ. Transcriptome). Полный специфический набор транскриптов (молекул РНК) в данном организме, представленный в клетках
- 7. Некодирующая функциональная РНК – функциональные молекулы РНК, не транслируемые в белки, например: рРНК, тРНК, piРНК, микроРНК.
- 8. Промоторы способствуют транскрипции конкретного гена и обычно располагаются вверху (в 5’-области гена) по направлению кодирующего региона.
- 9. Мобильные генетические элементы (МГЭ, англ. Mobile genetic elements, MGE) – последовательности ДНК, перемещающиеся внутри генома. Псевдогены
- 10. Ретротранспозонны - повторяющиеся длинные последовательности, включают диспергированные повторы (LINEs) и короткие диспергированные повторы (SINEs), составляют большую
- 11. Alu-повтор, Alu-элемент – короткая последовательность ДНК, которая была открыта при обработке ДНК человека рестриктазой Alu. Alu-повтор
- 12. Последовательности эндогенных ретровирусов – продукты обратной транскрипции геномов ретровирусов и их включения в геном клеток зародышевой
- 13. Гены Ортологи Паралоги филогенетически родственных организмов, разошедшихся в процессе видообразования; гены, которые у различных видов произошли
- 14. Псевдогенизация – адаптивное эволюцинное приспособление. У человека примерами псевдогенизации являются: 1) У коренных жителей Западной Африки
- 15. Большая часть генома представлена нетранскрибируемой (некодирующей) белок частью (80%), представленной, в основном, повторами различных регуляторных РНК.
- 16. Естественный отбор не может просто поддерживать некий уровень «избыточности» участков генома, степень избыточности должна постоянно увеличиваться!
- 17. По-видимому, видообразование связано не с количеством генов, а с развитостью механизмов их чтения. Свободноживущая нематода Caenorhabditis
- 18. Упаковка хроматина эукариот Если растянуть все 46 хромосом человека в одну нить, то набор хромосом одной
- 19. Нуклеосомы Повторяющиеся структурные элементы эукариотического хроматина – нуклеосомы. Они состоят из двухцепочечной ДНК, обернутой вокруг октамеров
- 20. Общая структура нуклеосом напоминает «бусины на нитке». Эта структура уменьшает длину ДНК примерно в семь раз
- 21. Участвуют в организации и компактизации хромосом Гистоновый октамер Гистон H1 Линкерная ДНК Негистоновые белки Нуклеосома показана
- 22. Нуклеосомы соединяются, образуя 30 нм волокна Нуклеосомы связываются друг с другом и формируют более компактную структуру
- 23. Регулярная, спиральная конфигурация, содержащая шесть нуклеосом Нерегулярная конфигурация нуклеосом, располагающихся друг напротив друга Соленоидная модель укладки
- 24. Дальнейшее уплотнение хромосомы В результате двух предшествующих этапов, нить ДНК уменьшилась почти в 50 раз. Третий
- 25. Третий этап компактизации ДНК предполагает формирование радиальной петли доменов Матрично- вложенные области (MAR) MARs крепятся к
- 26. Плотность разных участков интерфазных хромосом не одинакова. Эухроматин образует менее сжатые участки хромосом. Это транскрипционно активные
- 27. Существует два типа гетерохроматина: Конститутивный гетерохроматин – области, которые всегда гетерохромны и постоянно транскрипционно неактивны. Факультативный
- 28. Двойная спираль ДНК ДНК, накрученная на октамер Гистон H1 Гистоны Нуклеосома Нуклеосома «бусины на нитке» Формирование
- 29. Плотность компактизации в эухроматин Плотность компактизации в гетерохроматин Во время интерфазы большинство хромосомных областей эухроматичны Петлевой
- 30. Метафазные хромосомы В метафазных хромосомах хроматин уплотнен сверхплотно. В формировании и организации метафазных хромосом участвуют два
- 31. Конденсации метафазной хромосомы с участием конденсина Количество петель не изменилось, но диаметр каждой петли уменьшился Конденсин
- 32. Выравнивание сестринских хроматид с помощью когезина Когезин размещается вдоль ручек хромосомы
- 34. СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА 1.Триплетность кода. Как отмечалось выше, кодон состоит из последовательности, состоящей из трех нуклеотидов,
- 35. 4.Неперекрываемость кода. В соответствии с этим свойством следующие за инициирующим триплетом кодоны читаются друг за другом,
- 36. Биологические функции ДНК Хранилище генетической информации (генетическое копирование): генетическая информация кодируется трехмерным расположением нуклеотидных оснований. Репликация:
- 37. Биологические функции ДНК Мутации – изменения в последовательности азотистых оснований в связи с добавлением, удалением или
- 38. Биологические функции ДНК Развитие: ДНК контролирует развитие организма посредством работы внутренние генетические часы с или без
- 39. Что на современном этапе представляет собой структурная часть генома, кодирующая белки? Структура гена.
- 40. Учебник: ген – единица наследственной информации, занимающая определенное положение в геноме или хромосоме и контролирующая выполнение
- 41. ген – это подпрограмма в операционной системе генома [Gerstein M. B., Bruce C., Rozowsky J. S.,
- 42. Структура гена Относительно короткие кодирующие последовательности оснований (экзоны) чередуются с длинными некодирующими последовательностями – интронами, которые
- 45. Классификация генов По функциям: 1. Структурные 2. Регуляторные: а) модификаторы б) супрессоры По влиянию на жизнеспособность:
- 46. Регуляторная часть гена Промотор – на 5’ конце (инициация транскрипции) Оператор – сайт связи с белком
- 47. Кодирующая часть гена 1. нетранслируемая 5’ область (НТО) 2. кодирующая область (состоит из кодонов) 3. нетранслируемая
- 48. Строение гена Кодирующая часть АТГ STOP ДНК РНК-транскрипт Промотор Терминатор Точка начала транскрипции Окончание транскрипции 5'
- 49. У прокариот регуляторные зоны «обслуживают» несколько генов. Эти гены вместе с регуляторными элементами носят название оперон.
- 50. Промотор — участок ДНК, к которому присоединяется РНК-полимераза и начинается транскрипция. Лидерная область включает или отключает
- 51. Трейлерная последовательность транскрибируется на иРНК и является сигналом для формированию полиаденилового хвостика (аденилирование 3`-конца) при «созревании»
- 52. Активатор обеспечивает присоединение РНК-полимеразы к промотору, на оператор осаждается белок регулятор, блокируя движение РНК-полимеразы через оператор.
- 53. Генотип у всех соматических клеток одинаковый (следствие равного распределения генетического материала между дочерними клетками при митозе),
- 54. ОСОБЕННОСТИ ЭУКАРИОТ Клетки эукариот имеют одинаковую ДНК, но фенотипически различаются. Это обусловлено тем, что в клетках
- 55. Экспрессия генов это совокупность процессов в клетке обеспечивающих реализацию информации гена в признак, то есть, это
- 56. Экспрессия структурных генов несущих информацию о структуре белка состоит из нескольких этапов: транскрипция → процессинг →
- 57. Схема стадий экспрессии гена
- 58. Экспрессия гена может быть измерена Количественный анализ экспрессии генов — измерение транскрипционной активности гена, с помощью
- 59. Схема структуры оперона – А, схемы негативного – Б и позитивного – В контроля экспрессии гена.
- 60. Контроль экспрессии генов осуществляется на следующих уровнях: при транскрипции (контролируется время и характер транскрипции гена); во
- 61. Регуляция экспрессии генов может осуществляться разными способами По типу индукции – поступивший в клетку субстрат вызывает
- 62. Lac-оперон Транскрипция генов S1, S2, S3 заблокирована Lac-оперон Е.соli: Контролирует синтез ферментов катаболизма лактозы по типу
- 63. 2. Лактоза выступает индуктором работы Lac-оперона и синтеза ферментов ее метаболизма Лактоза связывает белок–репрессор и Lac-оперон
- 64. ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ В основе дифференциальной активности генов на уровне транскрипции лежат обратимые (эпигеномные) изменения отдельных
- 65. Цитоплазматическая наследственность Кроме ядерных генов, локализованных в хромосомах, существуют факторы наследственности, находящиеся в цитоплазме – плазмагены
- 66. Цитоплазматическая наследственность Возможными критериями цитоплазматической наследственности являются: отсутствие количественного менделевского расщепления в потомстве; невозможность выявления сцепления;
- 67. Цитоплазматическая наследственность Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16 569 пар нуклеотидов. В состав
- 69. Скачать презентацию

Организация эукариотических хромосом
Эукариотическая хромосома содержит длинные, линейные молекулы ДНК, которые
Организация эукариотических хромосом
Эукариотическая хромосома содержит длинные, линейные молекулы ДНК, которые

У эукариот ген, как правило, дискретен,
то есть, разделен на участки
У эукариот ген, как правило, дискретен,
то есть, разделен на участки

Уникальные (неповторяющиеся последовательности – в геноме встречаются один или несколько раз.
Уникальные (неповторяющиеся последовательности – в геноме встречаются один или несколько раз.

Биологическая роль ДНК повторов
ДНК повторы обеспечивают разнообразие архитектоники генома и хромосом
Биологическая роль ДНК повторов
ДНК повторы обеспечивают разнообразие архитектоники генома и хромосом

Терминология
Транскрипто́м (англ. Transcriptome). Полный специфический набор транскриптов (молекул РНК) в данном организме, представленный
Терминология
Транскрипто́м (англ. Transcriptome). Полный специфический набор транскриптов (молекул РНК) в данном организме, представленный

Некодирующая функциональная РНК – функциональные молекулы РНК, не транслируемые в белки, например:
Некодирующая функциональная РНК – функциональные молекулы РНК, не транслируемые в белки, например:

Промоторы способствуют транскрипции конкретного гена и обычно располагаются вверху (в 5’-области гена) по
Промоторы способствуют транскрипции конкретного гена и обычно располагаются вверху (в 5’-области гена) по

Мобильные генетические элементы (МГЭ, англ. Mobile genetic elements, MGE) – последовательности ДНК, перемещающиеся внутри генома.
Псевдогены
Мобильные генетические элементы (МГЭ, англ. Mobile genetic elements, MGE) – последовательности ДНК, перемещающиеся внутри генома.
Псевдогены

Ретротранспозонны - повторяющиеся длинные последовательности, включают диспергированные повторы (LINEs) и короткие диспергированные повторы (SINEs), составляют
Ретротранспозонны - повторяющиеся длинные последовательности, включают диспергированные повторы (LINEs) и короткие диспергированные повторы (SINEs), составляют

Alu-повтор, Alu-элемент – короткая последовательность ДНК, которая была открыта при обработке ДНК
Alu-повтор, Alu-элемент – короткая последовательность ДНК, которая была открыта при обработке ДНК

Последовательности эндогенных ретровирусов – продукты обратной транскрипции геномов ретровирусов и их включения в геном клеток зародышевой
Последовательности эндогенных ретровирусов – продукты обратной транскрипции геномов ретровирусов и их включения в геном клеток зародышевой
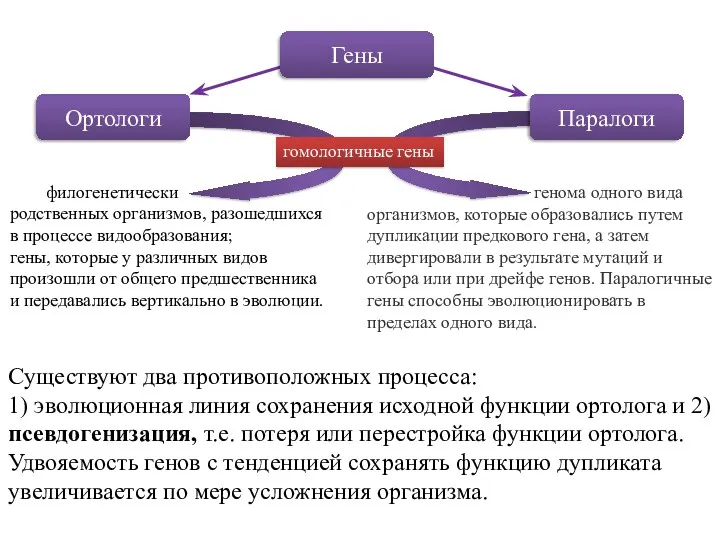
Гены
Ортологи
Паралоги
филогенетически
родственных организмов, разошедшихся в процессе видообразования;
гены, которые у
Гены
Ортологи
Паралоги
филогенетически
родственных организмов, разошедшихся в процессе видообразования;
гены, которые у

Псевдогенизация – адаптивное эволюцинное приспособление.
У человека примерами псевдогенизации являются:
1)
Псевдогенизация – адаптивное эволюцинное приспособление.
У человека примерами псевдогенизации являются:
1)
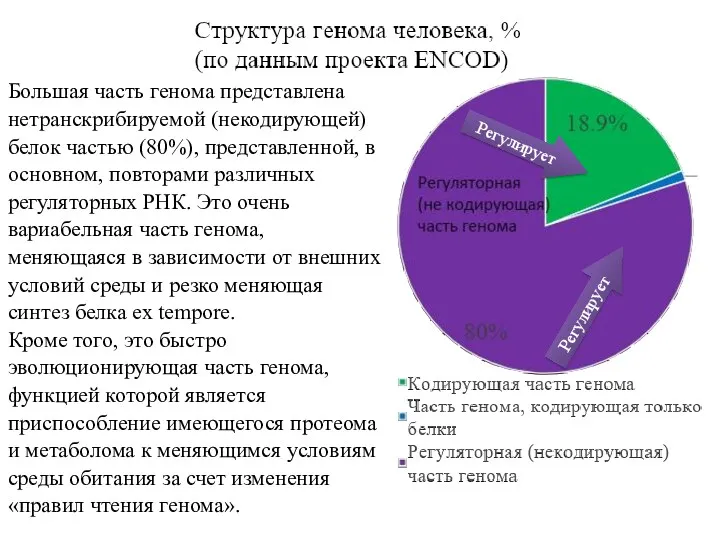
Большая часть генома представлена нетранскрибируемой (некодирующей) белок частью (80%), представленной, в
Большая часть генома представлена нетранскрибируемой (некодирующей) белок частью (80%), представленной, в

Естественный отбор не может просто поддерживать некий уровень «избыточности» участков генома,
Естественный отбор не может просто поддерживать некий уровень «избыточности» участков генома,

По-видимому, видообразование связано не с количеством генов, а с развитостью механизмов
По-видимому, видообразование связано не с количеством генов, а с развитостью механизмов

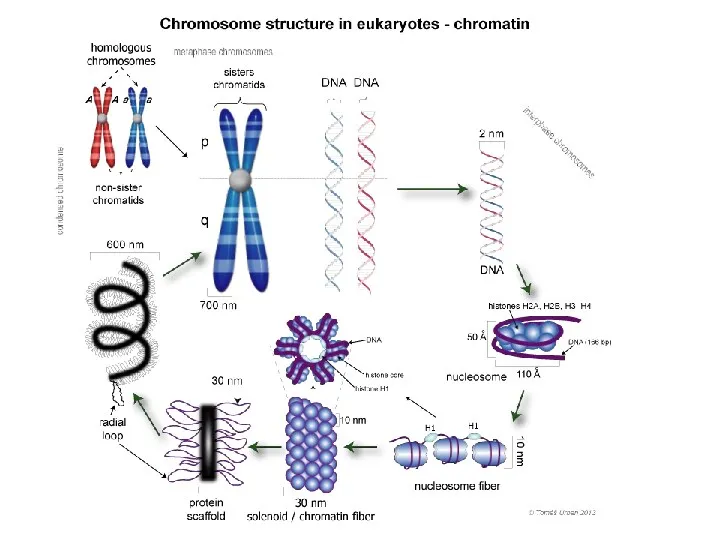
Упаковка хроматина эукариот
Если растянуть все 46 хромосом человека в одну нить,
Упаковка хроматина эукариот
Если растянуть все 46 хромосом человека в одну нить,

Нуклеосомы
Повторяющиеся структурные элементы эукариотического хроматина – нуклеосомы.
Они состоят из
Нуклеосомы
Повторяющиеся структурные элементы эукариотического хроматина – нуклеосомы.
Они состоят из
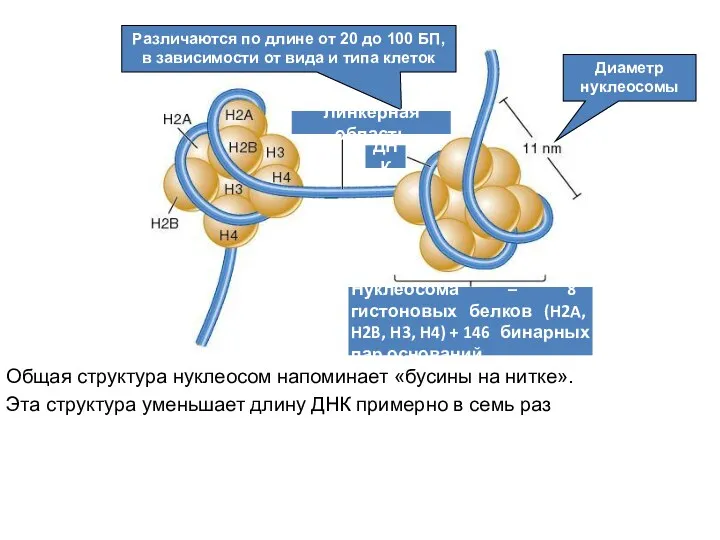
Общая структура нуклеосом напоминает «бусины на нитке».
Эта структура уменьшает длину ДНК
Общая структура нуклеосом напоминает «бусины на нитке».
Эта структура уменьшает длину ДНК
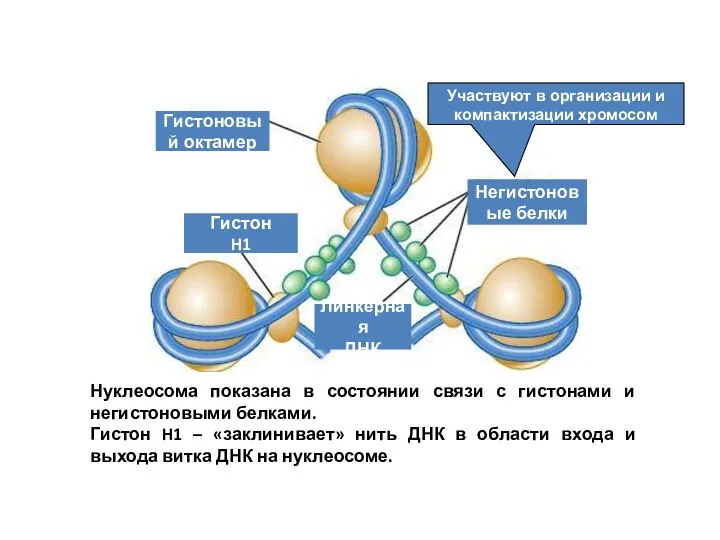
Участвуют в организации и компактизации хромосом
Гистоновый октамер
Гистон
H1
Линкерная
ДНК
Негистоновые белки
Нуклеосома показана в состоянии
Участвуют в организации и компактизации хромосом
Гистоновый октамер
Гистон
H1
Линкерная
ДНК
Негистоновые белки
Нуклеосома показана в состоянии

Нуклеосомы соединяются, образуя 30 нм волокна
Нуклеосомы связываются друг с другом и
Нуклеосомы соединяются, образуя 30 нм волокна
Нуклеосомы связываются друг с другом и
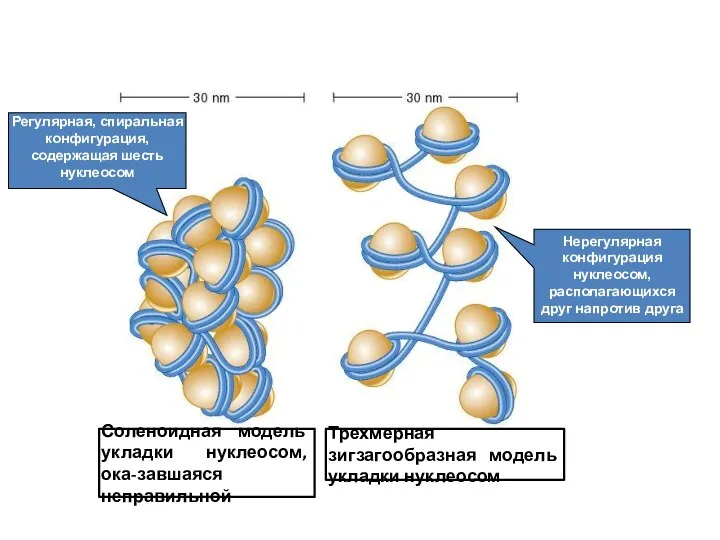
Регулярная, спиральная конфигурация, содержащая шесть нуклеосом
Нерегулярная конфигурация нуклеосом, располагающихся друг напротив
Регулярная, спиральная конфигурация, содержащая шесть нуклеосом
Нерегулярная конфигурация нуклеосом, располагающихся друг напротив

Дальнейшее уплотнение хромосомы
В результате двух предшествующих этапов, нить ДНК уменьшилась почти
Дальнейшее уплотнение хромосомы
В результате двух предшествующих этапов, нить ДНК уменьшилась почти
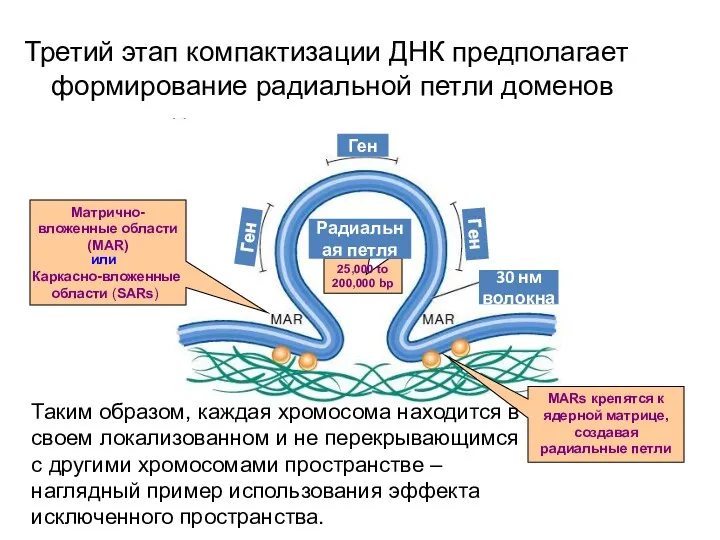
Третий этап компактизации ДНК предполагает формирование радиальной петли доменов
Матрично- вложенные области
Третий этап компактизации ДНК предполагает формирование радиальной петли доменов
Матрично- вложенные области

Плотность разных участков интерфазных хромосом не одинакова.
Эухроматин образует менее сжатые участки
Плотность разных участков интерфазных хромосом не одинакова.
Эухроматин образует менее сжатые участки
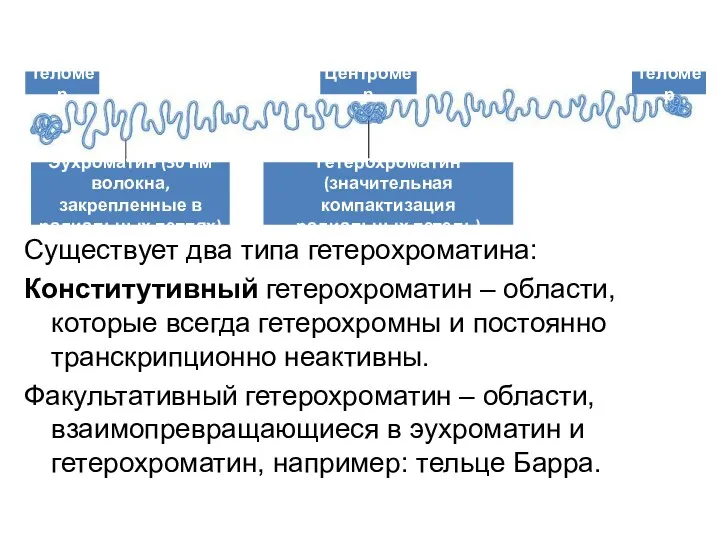
Существует два типа гетерохроматина:
Конститутивный гетерохроматин – области, которые всегда гетерохромны и
Существует два типа гетерохроматина:
Конститутивный гетерохроматин – области, которые всегда гетерохромны и
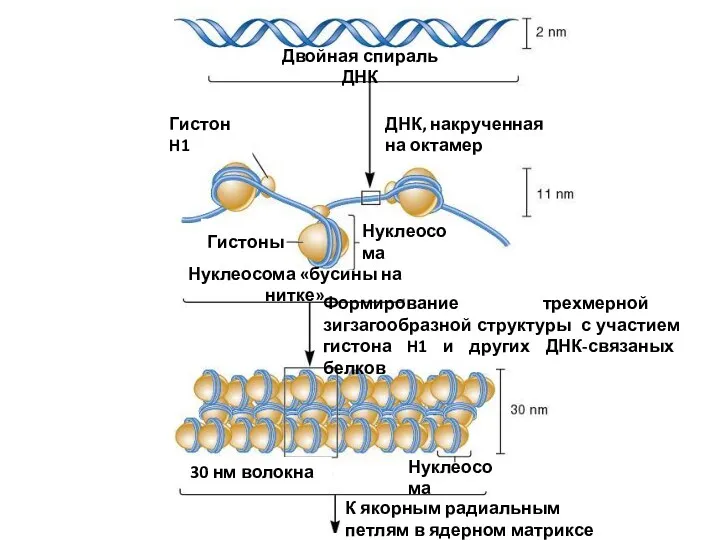
Двойная спираль ДНК
ДНК, накрученная на октамер
Гистон H1
Гистоны
Нуклеосома
Нуклеосома «бусины на нитке»
Формирование трехмерной
Двойная спираль ДНК
ДНК, накрученная на октамер
Гистон H1
Гистоны
Нуклеосома
Нуклеосома «бусины на нитке»
Формирование трехмерной
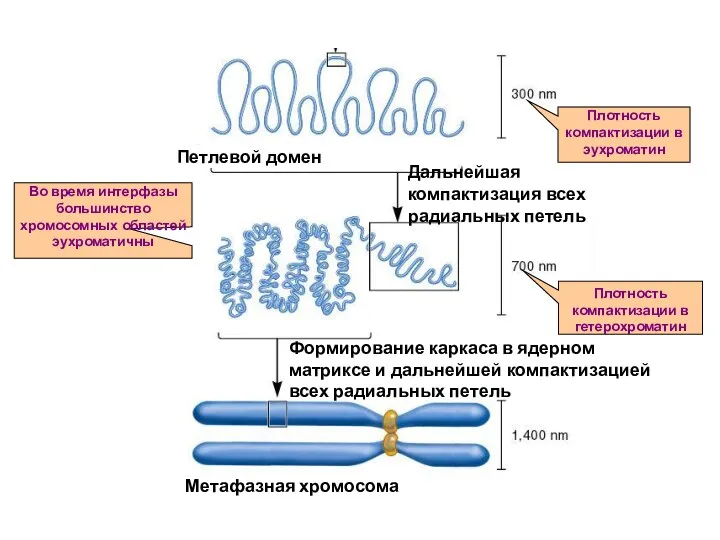
Плотность компактизации в эухроматин
Плотность компактизации в гетерохроматин
Во время интерфазы большинство
Плотность компактизации в эухроматин
Плотность компактизации в гетерохроматин
Во время интерфазы большинство

Метафазные хромосомы
В метафазных хромосомах хроматин уплотнен сверхплотно.
В формировании и организации метафазных
Метафазные хромосомы
В метафазных хромосомах хроматин уплотнен сверхплотно.
В формировании и организации метафазных
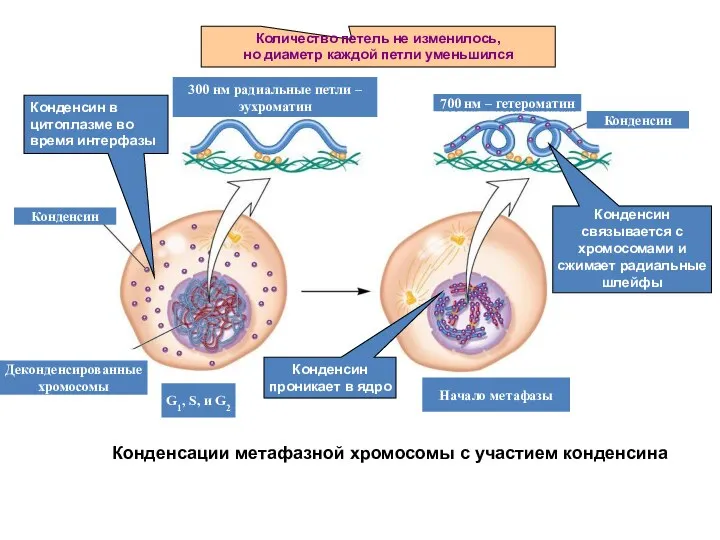
Конденсации метафазной хромосомы с участием конденсина
Количество петель не изменилось,
но диаметр каждой
Конденсации метафазной хромосомы с участием конденсина
Количество петель не изменилось,
но диаметр каждой
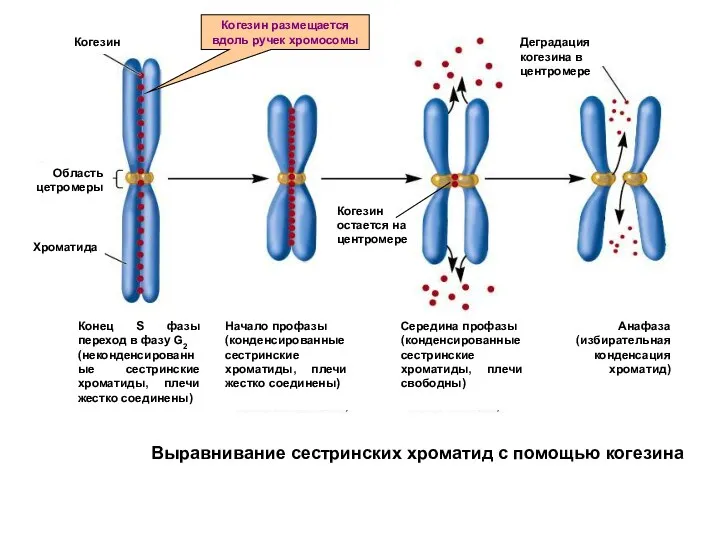
Выравнивание сестринских хроматид с помощью когезина
Когезин размещается вдоль ручек хромосомы
Выравнивание сестринских хроматид с помощью когезина
Когезин размещается вдоль ручек хромосомы

СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
1.Триплетность кода. Как отмечалось выше, кодон состоит из
СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
1.Триплетность кода. Как отмечалось выше, кодон состоит из

4.Неперекрываемость кода. В соответствии с этим свойством следующие за
4.Неперекрываемость кода. В соответствии с этим свойством следующие за

Биологические функции ДНК
Хранилище генетической информации (генетическое копирование): генетическая информация кодируется
Биологические функции ДНК
Хранилище генетической информации (генетическое копирование): генетическая информация кодируется

Биологические функции ДНК
Мутации – изменения в последовательности азотистых оснований в
Биологические функции ДНК
Мутации – изменения в последовательности азотистых оснований в

Биологические функции ДНК
Развитие: ДНК контролирует развитие организма посредством работы внутренние
Биологические функции ДНК
Развитие: ДНК контролирует развитие организма посредством работы внутренние

Что на современном этапе представляет собой структурная часть генома, кодирующая
Что на современном этапе представляет собой структурная часть генома, кодирующая

Учебник: ген – единица наследственной информации, занимающая определенное положение в геноме
Учебник: ген – единица наследственной информации, занимающая определенное положение в геноме

ген – это подпрограмма в операционной системе генома [Gerstein M.
ген – это подпрограмма в операционной системе генома [Gerstein M.
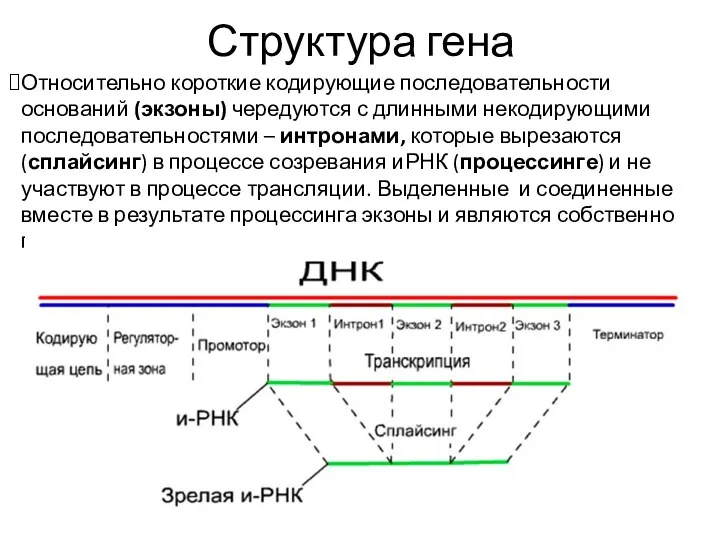
Структура гена
Относительно короткие кодирующие последовательности оснований (экзоны) чередуются с длинными некодирующими последовательностями
Структура гена
Относительно короткие кодирующие последовательности оснований (экзоны) чередуются с длинными некодирующими последовательностями
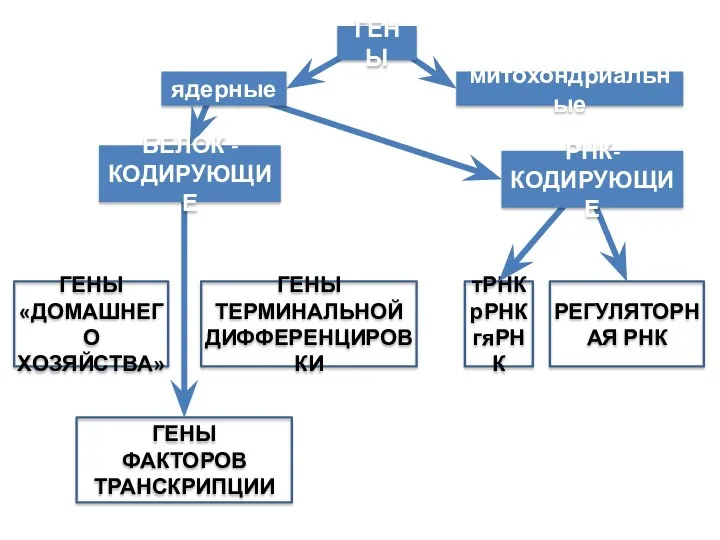

Классификация генов
По функциям:
1. Структурные
2. Регуляторные:
а) модификаторы
б) супрессоры
По влиянию на жизнеспособность:
Летальные,
полулетальные,
Классификация генов
По функциям:
1. Структурные
2. Регуляторные:
а) модификаторы
б) супрессоры
По влиянию на жизнеспособность:
Летальные,
полулетальные,

Регуляторная часть гена
Промотор – на 5’ конце (инициация транскрипции)
Оператор – сайт связи
Регуляторная часть гена
Промотор – на 5’ конце (инициация транскрипции)
Оператор – сайт связи
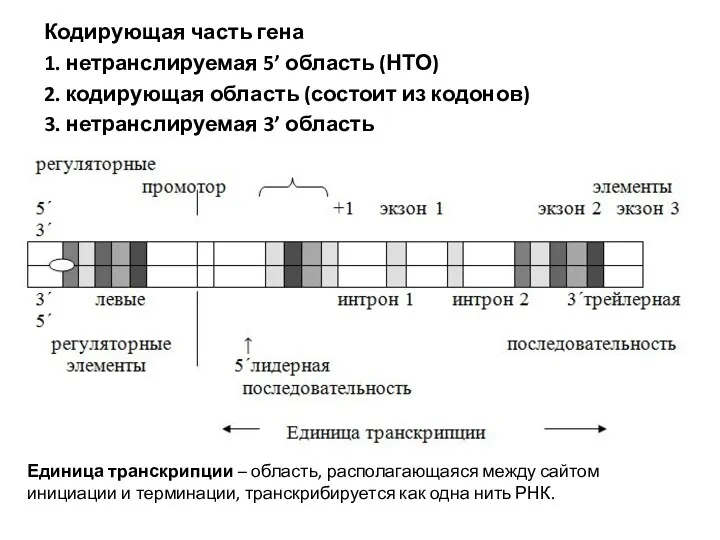
Кодирующая часть гена
1. нетранслируемая 5’ область (НТО)
2. кодирующая область (состоит из
Кодирующая часть гена
1. нетранслируемая 5’ область (НТО)
2. кодирующая область (состоит из
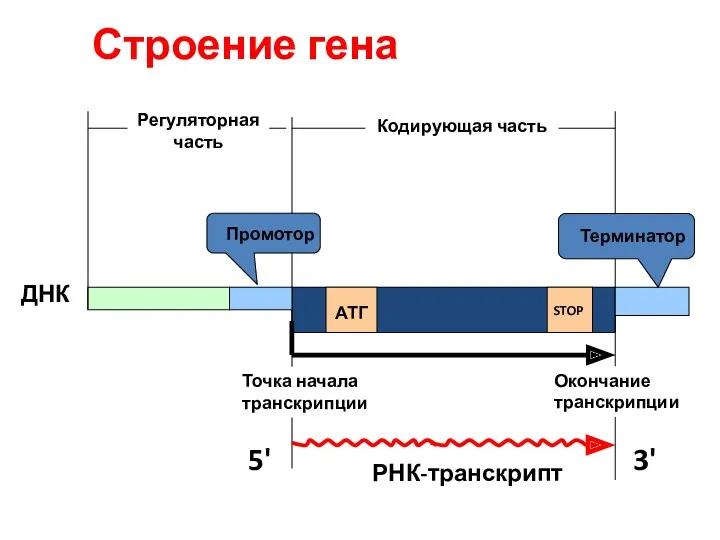
Строение гена
Кодирующая часть
АТГ
STOP
ДНК
РНК-транскрипт
Промотор
Терминатор
Точка начала транскрипции
Окончание транскрипции
5'
3'
Регуляторная часть
Строение гена
Кодирующая часть
АТГ
STOP
ДНК
РНК-транскрипт
Промотор
Терминатор
Точка начала транскрипции
Окончание транскрипции
5'
3'
Регуляторная часть

У прокариот регуляторные зоны «обслуживают» несколько генов. Эти гены вместе с
У прокариот регуляторные зоны «обслуживают» несколько генов. Эти гены вместе с

Промотор — участок ДНК, к которому присоединяется
РНК-полимераза и начинается транскрипция.
Лидерная
Промотор — участок ДНК, к которому присоединяется
РНК-полимераза и начинается транскрипция.
Лидерная

Трейлерная последовательность транскрибируется на иРНК и является сигналом для формированию полиаденилового хвостика
Трейлерная последовательность транскрибируется на иРНК и является сигналом для формированию полиаденилового хвостика

Активатор обеспечивает присоединение РНК-полимеразы к промотору, на оператор осаждается белок регулятор,
Активатор обеспечивает присоединение РНК-полимеразы к промотору, на оператор осаждается белок регулятор,

Генотип у всех соматических клеток одинаковый (следствие равного распределения генетического материала
Генотип у всех соматических клеток одинаковый (следствие равного распределения генетического материала

ОСОБЕННОСТИ ЭУКАРИОТ
Клетки эукариот имеют одинаковую ДНК, но фенотипически различаются.
Это
ОСОБЕННОСТИ ЭУКАРИОТ
Клетки эукариот имеют одинаковую ДНК, но фенотипически различаются.
Это

Экспрессия генов это совокупность процессов в клетке обеспечивающих реализацию информации
Экспрессия генов это совокупность процессов в клетке обеспечивающих реализацию информации

Экспрессия структурных генов несущих информацию о структуре белка состоит из
Экспрессия структурных генов несущих информацию о структуре белка состоит из
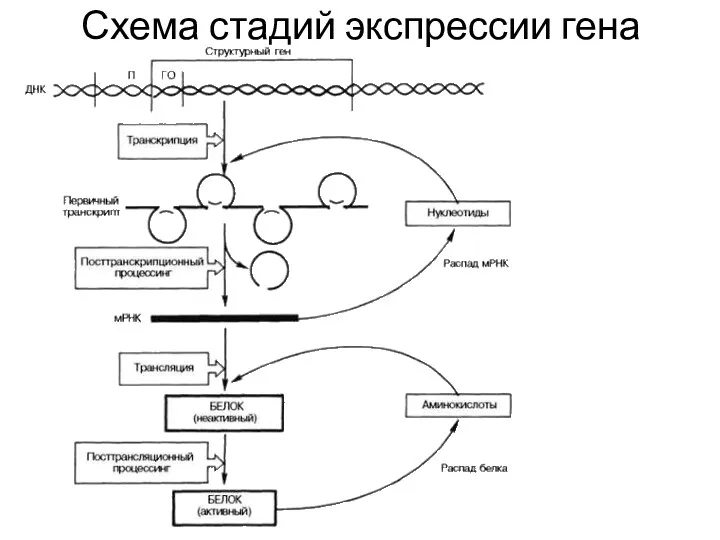
Схема стадий экспрессии гена
Схема стадий экспрессии гена

Экспрессия гена может быть измерена
Количественный анализ экспрессии генов — измерение транскрипционной активности гена,
Экспрессия гена может быть измерена
Количественный анализ экспрессии генов — измерение транскрипционной активности гена,
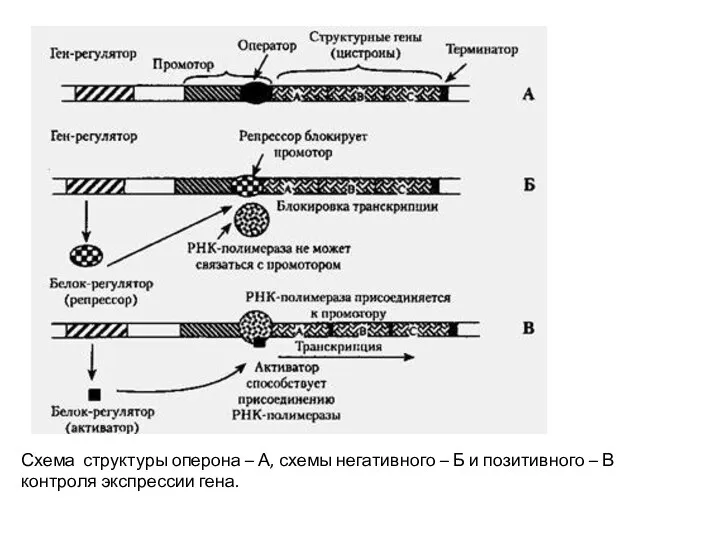
Схема структуры оперона – А, схемы негативного – Б и позитивного
Схема структуры оперона – А, схемы негативного – Б и позитивного

Контроль экспрессии генов осуществляется на следующих уровнях:
при транскрипции (контролируется время и
Контроль экспрессии генов осуществляется на следующих уровнях:
при транскрипции (контролируется время и

Регуляция экспрессии генов может осуществляться разными способами
По типу индукции – поступивший
Регуляция экспрессии генов может осуществляться разными способами
По типу индукции – поступивший
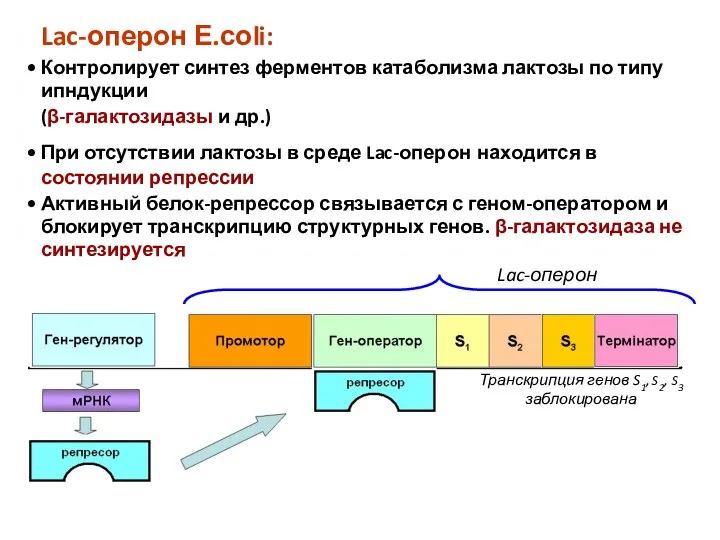
Lac-оперон
Транскрипция генов S1, S2, S3 заблокирована
Lac-оперон Е.соli:
Контролирует синтез ферментов катаболизма лактозы
Lac-оперон
Транскрипция генов S1, S2, S3 заблокирована
Lac-оперон Е.соli:
Контролирует синтез ферментов катаболизма лактозы
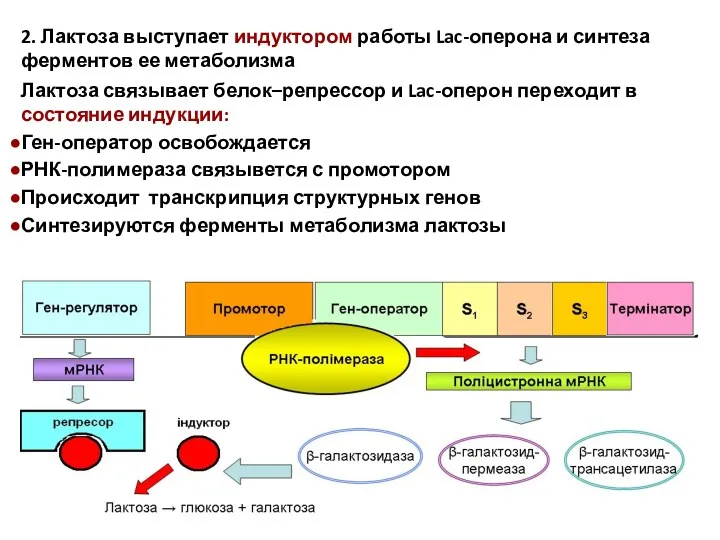
2. Лактоза выступает индуктором работы Lac-оперона и синтеза ферментов ее метаболизма
2. Лактоза выступает индуктором работы Lac-оперона и синтеза ферментов ее метаболизма

ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ
В основе дифференциальной активности генов на уровне транскрипции лежат обратимые
ДИФФЕРЕНЦИАЛЬНАЯ ЭКСПРЕССИЯ ГЕНОВ
В основе дифференциальной активности генов на уровне транскрипции лежат обратимые

Цитоплазматическая наследственность
Кроме ядерных генов, локализованных в хромосомах, существуют факторы
Цитоплазматическая наследственность
Кроме ядерных генов, локализованных в хромосомах, существуют факторы

Цитоплазматическая наследственность
Возможными критериями цитоплазматической наследственности являются:
отсутствие количественного менделевского расщепления в потомстве;
невозможность
Цитоплазматическая наследственность
Возможными критериями цитоплазматической наследственности являются:
отсутствие количественного менделевского расщепления в потомстве;
невозможность

Цитоплазматическая наследственность
Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16 569
Цитоплазматическая наследственность
Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16 569
 Мышцы головы, шеи и туловища. (Лекция 8)
Мышцы головы, шеи и туловища. (Лекция 8) В гости к весне. 4 класс
В гости к весне. 4 класс Бабочки и мотыльки
Бабочки и мотыльки Класс однодольные
Класс однодольные Ядовитые растения, произрастающие в Ленинградской области
Ядовитые растения, произрастающие в Ленинградской области Класс Головоногие моллюски
Класс Головоногие моллюски Полезные и лекарственные растения Зауралья
Полезные и лекарственные растения Зауралья Оценка видового разнообразия и экоконтролинг сезонных скоплений птиц полигона твердых бытовых отходов г. Барнаул
Оценка видового разнообразия и экоконтролинг сезонных скоплений птиц полигона твердых бытовых отходов г. Барнаул Класс Млекопитающие (Mammalia)
Класс Млекопитающие (Mammalia) Різноманітність і значення найпростіших
Різноманітність і значення найпростіших Биология – наука о жизни
Биология – наука о жизни Мимические мыщцы лица. Особенности строения и функционирования
Мимические мыщцы лица. Особенности строения и функционирования Класс млекопитающие
Класс млекопитающие Разнообразие растений
Разнообразие растений Витамины. Общая характеристика витаминов
Витамины. Общая характеристика витаминов Необычные животные и растения
Необычные животные и растения Скелет и соединения конечностей. Кости черепа человека
Скелет и соединения конечностей. Кости черепа человека Основы систематики растений. Деление покрытосеменных растений на классы и семейства
Основы систематики растений. Деление покрытосеменных растений на классы и семейства Индивидуальное развитие человека, или онтогенез
Индивидуальное развитие человека, или онтогенез Презентация к уроку биологии 8 класс на тему: Общая характеристика млекопитающих
Презентация к уроку биологии 8 класс на тему: Общая характеристика млекопитающих Биологические модели развития популяций
Биологические модели развития популяций Какие растения раньше других цвести торопятся
Какие растения раньше других цвести торопятся Черепные нервы I-VI
Черепные нервы I-VI Репарация ДНК
Репарация ДНК Отдел Покрытосеменные. Главные признаки отдела Покрытосеменные
Отдел Покрытосеменные. Главные признаки отдела Покрытосеменные Стебель. Побег
Стебель. Побег Презентация к уроку по теме Деление клетки
Презентация к уроку по теме Деление клетки Доза і основні одиниці вимірювання іонізуючого випромінювання. Основні типи дозових залежностей в радіобіології
Доза і основні одиниці вимірювання іонізуючого випромінювання. Основні типи дозових залежностей в радіобіології