- Основы генетики микробов

Содержание
- 2. НАСЛЕДСТВЕННОСТЬ И БАКТЕРИАЛЬНЫЙ ГЕНОМ СВОЙСТВА ОРГАНИЧЕСКИХ ФОРМ наследственность изменчивость ЭВОЛЮЦИОННЫЙ ПРОЦЕСС ОРГАНИЧЕСКИХ ФОРМ МИКРОБ - идеальная
- 3. Нуклеотид - это (nucleotide) - соединение, в состав которого входит сахар, фосфатная группа и азотсодержащее основание
- 4. РНК ДНК рибонуклеиновая кислота дезоксирибонуклеиновая кислота - Кодирование генетической информации - Синтез белка иРНК – информационная
- 6. МАКРОМОЛЕКУЛА ДНК ДНК имеет полярность. На одном ее конце фосфатная группа в положении 5‘, а на
- 7. ГЕНОТИП – это совокупность всех генов микробной клетки. => Изменение – ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ФЕНОТИП – это
- 8. РЕПЛИКАЦИЯ ДНК и ДЕЛЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ ЦИТОКИНЕЗ – деление бактериальной клетки. Обязательно предшествует репликация дезоксирибонуклеиновой кислоты
- 9. РЕПЛИКАЦИЯ ДНК и ДЕЛЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ Характеристики процесса репликации - МАТРИЧНЫЙ — последовательность синтезируемой цепи ДНК
- 10. Ферментативный комплекс процесса репликации ДНК-гираза – фермент инициации синтеза ДНК; Топоизомераза – раскручивание узлов, образующихся перед
- 12. Видео Основные моменты Репликации ДНК
- 13. ХАРАКТЕРИСТИКА ОСНОВНЫХ ФОРМ ИЗМЕНЧИВОСТИ ИНФОРМАЦИЯ Приобретает Бактериальный геном Теряет Измененный генотип Измененный фенотип Изменение фенотипа, без
- 14. ФЕНОТИПИЧЕСКАЯ (не наследуется) ИЗМЕНЧИВОСТЬ ГЕНТИПИЧЕСКАЯ (наследуется) Во время действия фактора После действия фактора Мутации Рекомбинации ЛАБИЛЬНЫЕ
- 15. Таким образом, на изменение внешних условий среды обитания, микробы отвечают изменением своих свойств, что способствует сохранению
- 16. ФОРМЫ ФЕНОТИПИЧЕСКОЙ (НЕНАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ МОДИФИКАЦИИ КРАТКОВРЕМЕННЫЕ ДЛИТЕЛЬНЫЕ Const. Первые генерации (1-2) Const. Ряд поколений Свойства бактерий:
- 17. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ МОДИФИКАЦИОННОЙ ИЗМЕНЧИВОСТИ 1. Носит групповой характер; 2. Зависит от окружающих условий; 3. Является определённой;
- 18. ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ МУТАЦИИ РЕКОМБИНАЦИИ СПОНТАННЫЕ ИНДУЦИРОВАННЫЕ трансформация конъюгация трансдукция лизогенная конверсия естественные экспериментальные
- 19. ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ МУТАЦИИ СПОНТАННЫЕ ИНДУЦИРОВАННЫЕ естественные экспериментальные – мутации, возникающие при нормальных условиях жизни.
- 20. ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ МУТАЦИИ ГЕННЫЕ ХРОМОСОМНЫЕ в пределах одного гена в пределах нескольких генов ТОЧКОВЫЕ
- 21. ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ МУТАГЕНЫ ФИЗИЧЕСКИЕ БИОЛОГИЧЕСКИЕ ХИМИЧЕСКИЕ Ультрафиолетовый свет Рентгеновские излучения и т.п. Инсерционные генетические
- 22. РЕПАРАЦИЯ КЛЕТОЧНОГО ГЕНОМА ФОТОРЕАКТИВАЦИЯ специальные ферменты Наличие видимого света РЕПАРИРУЮЩИЕ СИСТЕМЫ Репарация (от лат. reparatio —
- 25. ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ РЕКОМБИНАЦИИ Возникновение новых последовательностей ДНК в результате разрывов и перевоссоединений предсуществующих молекул
- 26. ТРАНСФОРМАЦИЯ Передача генетической информации из разрушенных бактерий-доноров в виде неповрежденной молекулы ДНК в клетки реципиенты ТРАНСФОРМАЦИЯ
- 27. 1928 год. Ф. Гриффит. Английский журнал «Гигиена»
- 28. ТРАНСФОРМАЦИЯ 1 этап – адсорбция выделившийся трансформирующей ДНК из клетки донора на рецепторах бактерии – реципиента.
- 29. Компетентность – это способность бактерий воспринимать (поглощать) трансформирующую ДНК. Фаза в конце лог-фазы (стадия лог-фазы III
- 30. КОНЪЮГАЦИЯ – (Ледерберг и Тейтум, 1946) Передача генетического материала от бактерии-донора к бактерии-реципиенту при непосредственном контакте.
- 34. Передача ДНК полового фактора F(+) от клетки – донора к F(-) в процессе репликации у бактерии
- 35. Образование Hfr – бактерий и процесса конъюгации между Hfr – клеткой (донором) и F (-) реципиентом
- 36. ПЛАЗМИДЫ и МИГРИРУЮЩИЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ Плазмиды – небольшие кольцевые двухцепочечные молекулы ДНК. Плазмиды содержат от 40
- 37. F-ПЛАЗМИДА F-плазмида. E.coli Обеспечивают участие в конъюгации между бактериями с помощью кодируемых ими половых пилей. -
- 38. БАКТЕРИОЦИНОГЕННАЯ ПЛАЗМИДА Большинство видов бактерий выделяют белковые антибактериальные вещества – БАКТЕРИОЦИНЫ (бактерицидная активность только в отношении
- 39. БАКТЕРИОЦИНОГЕННАЯ ПЛАЗМИДА Col-плазмида отличается от других плазмид рядом присущих только ей особенностей В отличие от других
- 40. R-ПЛАЗМИДА (resistance – устойчивость) РЕЗИСТЕНТНОСТЬ – 3 пути: мутации; рекомбинации; плазмиды с геном, кодирующим устойчивость к
- 41. R-ПЛАЗМИДА (resistance – устойчивость) RTF r-гены 2 фрагмента ДНК Содержит гены, кодирующие его репликацию и перенос
- 42. R-ПЛАЗМИДА (resistance – устойчивость)
- 44. ТРАНСПОЗИРУЕМЫЕ (МИГРИРУЮЩИЕ) ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ - Сегменты ДНК, самостоятельно перемещающиеся в пределах хромосомы бактерий, а также способны
- 45. Is - элементы - Это линейные участки ДНК, состоящие из 700-1750 пар азотистых оснований, имеющие на
- 46. ФУНКЦИИ IS-ЭЛЕМЕНТОВ Координирующая - взаимодействие транспозонов, плазмид, умеренных фагов между собой и хромосомой бактерии, обеспечивая их
- 47. - Подобен Is-элементам и транспозонам, так как может интегрировать в любой участок ДНК бактерий и обуславливать
- 48. БАКТЕРИОФАГИ Электронная микрофотография Т-четного бактериофага
- 50. Электронная микрофотография “атаки” бактериофагами клетки кишечной палочки E. coli
- 51. Электронная микрофотография фага T4, адсорбированного на E. coli Видны сокращенные чехлы отростка и “инъекция” фаговой ДНК
- 52. БАКТЕРИОФАГИ ( ОТ ЛАТ.«PHAGOS» - ПОЖИРАЮЩИЙ) – ВИРУСЫ БАКТЕРИЙ, ОБЛАДАЮЩИЕ ТЕМИ ЖЕ ХАРАКТЕРНЫМИ ОСОБЕННОСТЯМИ, ЧТО И
- 53. СТРОЕНИЕ БАКТЕРИОФАГОВ. ПОКОЯЩАЯСЯ, ВНЕКЛЕТОЧНАЯ, ФОРМА – ВИРИОН. ВНУТРИКЛЕТОЧНАЯ ФОРМА – ВЕГЕТАТИВНАЯ. ВИРИОН Головка белковый футляр (капсид)
- 54. Морфологические типы фагов I – нитевидные фаги; II – фаги без отростка; III- фаги с аналогом
- 55. НАИБОЛЕЕ СЛОЖНО УСТРОЕНЫ ФАГИ С СОКРАЩАЮЩИМСЯ ЧЕХЛОМ ОТРОСТКА, НАПРИМЕР, Т- ЧЕТНЫЕ ФАГИ (Т4) E. COLI
- 57. Электронная микрофотография ультратонких срезов E. coli, инфицированных фагом T2 на разных стадиях репродукции.
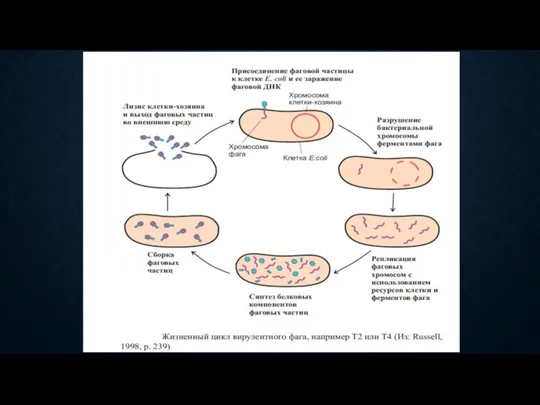
- 58. ВЗАИМОДЕЙСТВИЕ ФАГА С КЛЕТКОЙ вирулентные фаги вызывают продуктивную инфекцию, при которой происходит репродукция фагов и лизис
- 59. ЭТАПЫ ПРОДУКТИВНОЙ ИНФЕКЦИИ: 1я стадия. Адсорбция фага на чувствительной клетке. Происходит при наличии комплементарных рецепторов в
- 60. 4я стадия. Морфогенез фага. Созревание фага - разобщенный процесс. Отдельно формируются головки фага: вокруг ДНК строится
- 61. ЭТАПЫ ПРОДУКТИВНОЙ ИНФЕКЦИИ.
- 63. ИНТЕГРАТИВНАЯ ИНФЕКЦИЯ (ЛИЗОГЕНИЯ) ДНК фага включается в кольцевую хромосому бактериальной клетки. Во время деления клетки профаг
- 64. ИНТЕГРАЦИЯ ДНК ФАГА ЛЯМБДА В БАКТЕРИАЛЬНУЮ ДНК С ФОРМИРОВАНИЕМ ЛИЗОГЕННОЙ БАКТЕРИИ
- 65. ТРАНСДУКЦИЯ (ЦИНДЕР И ЛЕДЕРБЕРГ, 1952) Виды: общая (генерализированная) специфическая абортивная Вызывают умеренные, дефектные фаги – передача
- 66. Схема трансдукции: 1 – проникновение фага в клетку-донор; 2,3 – образование трансдуцирующего фага; 4 – взаимодействие
- 67. Схема неспецифической (общей) трансдукции
- 68. Схема специфической трансдукции с формированием мерозиготы
- 69. Абортивная трансдукция — перенос фагом участка ДНК клетки-донора в клетку-реципиент, которая не включается в ее геном,
- 70. ОТЛИЧИЯ ТРАНСДУКЦИИ И ФАГОВОЙ КОНВЕРСИИ Трансдукция – перенос генетической информации из клетки в клетку при помощи
- 71. Адсорбция фагов на бактериальной клетке
- 72. ПРИМЕНЕНИЕ БАКТЕРИОФАГОВ. 1. Для диагностики инфекционных заболеваний. а) для определения видовой принадлежности выделенной культуры бактерий. б)
- 73. ФАГОТИПИРОВАНИЕ Основа метода: с помощью типовых фагов дифференцируют культуры одного вида на основании их различной чувствительности
- 75. Скачать презентацию

НАСЛЕДСТВЕННОСТЬ И БАКТЕРИАЛЬНЫЙ ГЕНОМ
СВОЙСТВА ОРГАНИЧЕСКИХ ФОРМ
наследственность
изменчивость
ЭВОЛЮЦИОННЫЙ ПРОЦЕСС ОРГАНИЧЕСКИХ ФОРМ
МИКРОБ - идеальная
НАСЛЕДСТВЕННОСТЬ И БАКТЕРИАЛЬНЫЙ ГЕНОМ
СВОЙСТВА ОРГАНИЧЕСКИХ ФОРМ
наследственность
изменчивость
ЭВОЛЮЦИОННЫЙ ПРОЦЕСС ОРГАНИЧЕСКИХ ФОРМ
МИКРОБ - идеальная
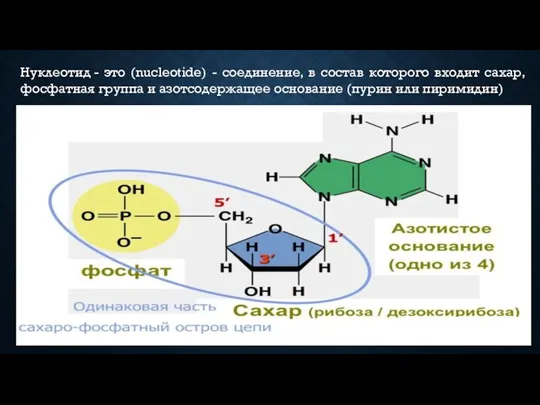
Нуклеотид - это (nucleotide) - соединение, в состав которого входит сахар, фосфатная
Нуклеотид - это (nucleotide) - соединение, в состав которого входит сахар, фосфатная

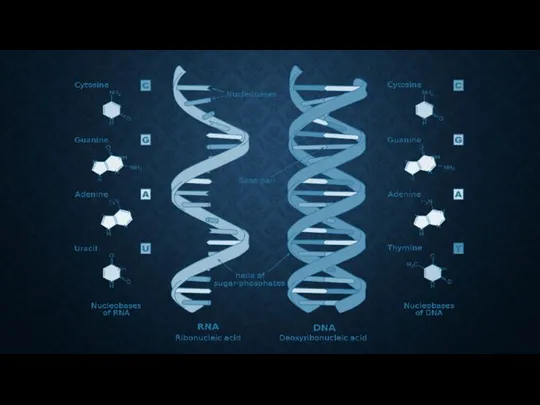
РНК
ДНК
рибонуклеиновая кислота
дезоксирибонуклеиновая кислота
- Кодирование генетической информации
- Синтез белка
иРНК – информационная РНК
тРНК
РНК
ДНК
рибонуклеиновая кислота
дезоксирибонуклеиновая кислота
- Кодирование генетической информации
- Синтез белка
иРНК – информационная РНК
тРНК

МАКРОМОЛЕКУЛА ДНК
ДНК имеет полярность. На одном ее конце фосфатная группа в
МАКРОМОЛЕКУЛА ДНК
ДНК имеет полярность. На одном ее конце фосфатная группа в

ГЕНОТИП – это совокупность всех генов микробной клетки.
=> Изменение – ГЕНОТИПИЧЕСКАЯ
ГЕНОТИП – это совокупность всех генов микробной клетки.
=> Изменение – ГЕНОТИПИЧЕСКАЯ

РЕПЛИКАЦИЯ ДНК и ДЕЛЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ
ЦИТОКИНЕЗ – деление бактериальной клетки. Обязательно
РЕПЛИКАЦИЯ ДНК и ДЕЛЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ
ЦИТОКИНЕЗ – деление бактериальной клетки. Обязательно

РЕПЛИКАЦИЯ ДНК и ДЕЛЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ
Характеристики процесса репликации
- МАТРИЧНЫЙ — последовательность синтезируемой
РЕПЛИКАЦИЯ ДНК и ДЕЛЕНИЕ БАКТЕРИАЛЬНОЙ КЛЕТКИ
Характеристики процесса репликации
- МАТРИЧНЫЙ — последовательность синтезируемой

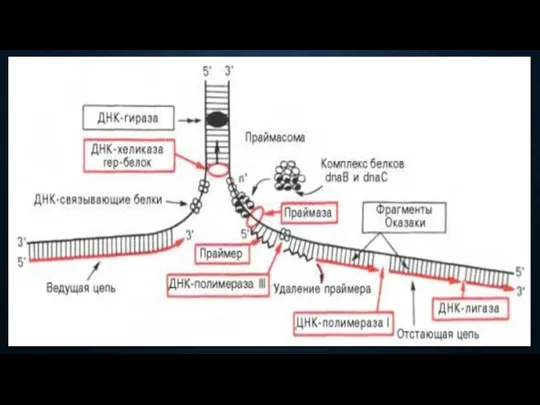
Ферментативный комплекс процесса репликации
ДНК-гираза – фермент инициации синтеза ДНК;
Топоизомераза – раскручивание
Ферментативный комплекс процесса репликации
ДНК-гираза – фермент инициации синтеза ДНК;
Топоизомераза – раскручивание

Видео
Основные моменты Репликации ДНК
Видео
Основные моменты Репликации ДНК
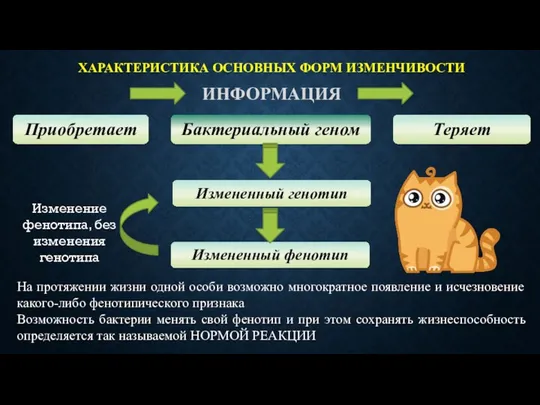
ХАРАКТЕРИСТИКА ОСНОВНЫХ ФОРМ ИЗМЕНЧИВОСТИ
ИНФОРМАЦИЯ
Приобретает
Бактериальный геном
Теряет
Измененный генотип
Измененный фенотип
Изменение фенотипа, без изменения генотипа
На
ХАРАКТЕРИСТИКА ОСНОВНЫХ ФОРМ ИЗМЕНЧИВОСТИ
ИНФОРМАЦИЯ
Приобретает
Бактериальный геном
Теряет
Измененный генотип
Измененный фенотип
Изменение фенотипа, без изменения генотипа
На
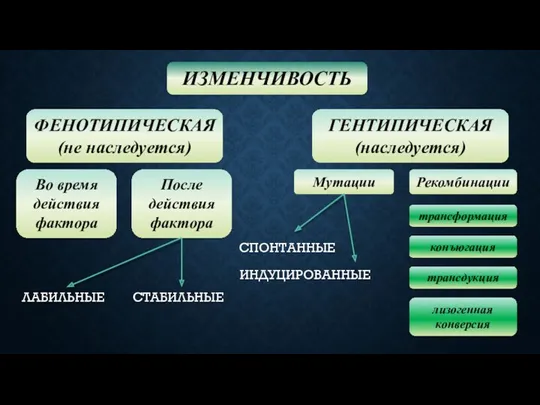
ФЕНОТИПИЧЕСКАЯ
(не наследуется)
ИЗМЕНЧИВОСТЬ
ГЕНТИПИЧЕСКАЯ
(наследуется)
Во время действия фактора
После действия фактора
Мутации
Рекомбинации
ЛАБИЛЬНЫЕ
СТАБИЛЬНЫЕ
СПОНТАННЫЕ
ИНДУЦИРОВАННЫЕ
трансформация
конъюгация
трансдукция
лизогенная конверсия
ФЕНОТИПИЧЕСКАЯ
(не наследуется)
ИЗМЕНЧИВОСТЬ
ГЕНТИПИЧЕСКАЯ
(наследуется)
Во время действия фактора
После действия фактора
Мутации
Рекомбинации
ЛАБИЛЬНЫЕ
СТАБИЛЬНЫЕ
СПОНТАННЫЕ
ИНДУЦИРОВАННЫЕ
трансформация
конъюгация
трансдукция
лизогенная конверсия

Таким образом, на изменение внешних условий среды обитания, микробы отвечают изменением
Таким образом, на изменение внешних условий среды обитания, микробы отвечают изменением
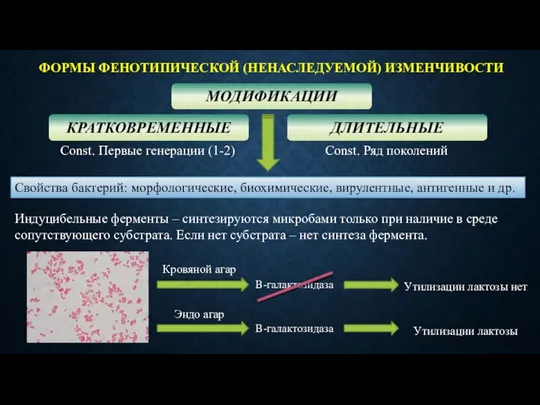
ФОРМЫ ФЕНОТИПИЧЕСКОЙ (НЕНАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МОДИФИКАЦИИ
КРАТКОВРЕМЕННЫЕ
ДЛИТЕЛЬНЫЕ
Const. Первые генерации (1-2)
Const. Ряд поколений
Свойства бактерий: морфологические,
ФОРМЫ ФЕНОТИПИЧЕСКОЙ (НЕНАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МОДИФИКАЦИИ
КРАТКОВРЕМЕННЫЕ
ДЛИТЕЛЬНЫЕ
Const. Первые генерации (1-2)
Const. Ряд поколений
Свойства бактерий: морфологические,

ОСНОВНЫЕ ХАРАКТЕРИСТИКИ МОДИФИКАЦИОННОЙ ИЗМЕНЧИВОСТИ
1. Носит групповой характер;
2. Зависит от окружающих условий;
3.
ОСНОВНЫЕ ХАРАКТЕРИСТИКИ МОДИФИКАЦИОННОЙ ИЗМЕНЧИВОСТИ
1. Носит групповой характер;
2. Зависит от окружающих условий;
3.

ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАЦИИ
РЕКОМБИНАЦИИ
СПОНТАННЫЕ
ИНДУЦИРОВАННЫЕ
трансформация
конъюгация
трансдукция
лизогенная конверсия
естественные
экспериментальные
ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАЦИИ
РЕКОМБИНАЦИИ
СПОНТАННЫЕ
ИНДУЦИРОВАННЫЕ
трансформация
конъюгация
трансдукция
лизогенная конверсия
естественные
экспериментальные

ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАЦИИ
СПОНТАННЫЕ
ИНДУЦИРОВАННЫЕ
естественные
экспериментальные
– мутации, возникающие при нормальных
условиях жизни. Спонтанный процесс
ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАЦИИ
СПОНТАННЫЕ
ИНДУЦИРОВАННЫЕ
естественные
экспериментальные
– мутации, возникающие при нормальных
условиях жизни. Спонтанный процесс
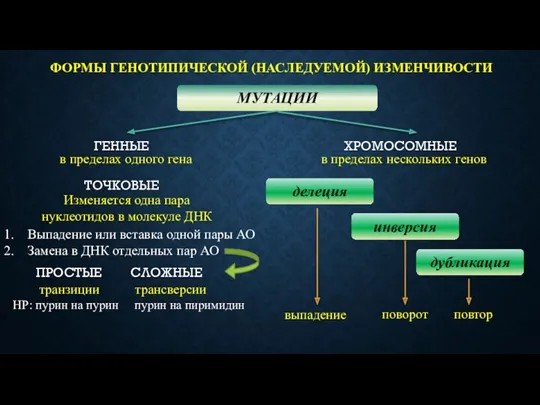
ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАЦИИ
ГЕННЫЕ
ХРОМОСОМНЫЕ
в пределах одного гена
в пределах нескольких генов
ТОЧКОВЫЕ
Изменяется одна
ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАЦИИ
ГЕННЫЕ
ХРОМОСОМНЫЕ
в пределах одного гена
в пределах нескольких генов
ТОЧКОВЫЕ
Изменяется одна

ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАГЕНЫ
ФИЗИЧЕСКИЕ
БИОЛОГИЧЕСКИЕ
ХИМИЧЕСКИЕ
Ультрафиолетовый свет
Рентгеновские излучения и т.п.
Инсерционные генетические элементы
Mu-бактериофаг –
ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
МУТАГЕНЫ
ФИЗИЧЕСКИЕ
БИОЛОГИЧЕСКИЕ
ХИМИЧЕСКИЕ
Ультрафиолетовый свет
Рентгеновские излучения и т.п.
Инсерционные генетические элементы
Mu-бактериофаг –

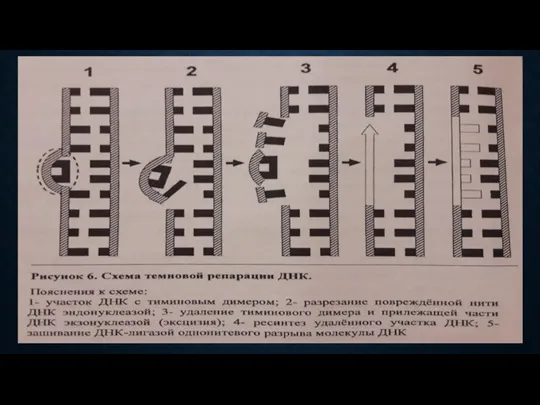
РЕПАРАЦИЯ КЛЕТОЧНОГО ГЕНОМА
ФОТОРЕАКТИВАЦИЯ
специальные ферменты
Наличие видимого света
РЕПАРИРУЮЩИЕ СИСТЕМЫ
Репарация (от лат. reparatio —
РЕПАРАЦИЯ КЛЕТОЧНОГО ГЕНОМА
ФОТОРЕАКТИВАЦИЯ
специальные ферменты
Наличие видимого света
РЕПАРИРУЮЩИЕ СИСТЕМЫ
Репарация (от лат. reparatio —


ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
РЕКОМБИНАЦИИ
Возникновение новых последовательностей ДНК в результате разрывов и
ФОРМЫ ГЕНОТИПИЧЕСКОЙ (НАСЛЕДУЕМОЙ) ИЗМЕНЧИВОСТИ
РЕКОМБИНАЦИИ
Возникновение новых последовательностей ДНК в результате разрывов и

ТРАНСФОРМАЦИЯ
Передача генетической информации из разрушенных бактерий-доноров в виде неповрежденной молекулы ДНК
ТРАНСФОРМАЦИЯ
Передача генетической информации из разрушенных бактерий-доноров в виде неповрежденной молекулы ДНК
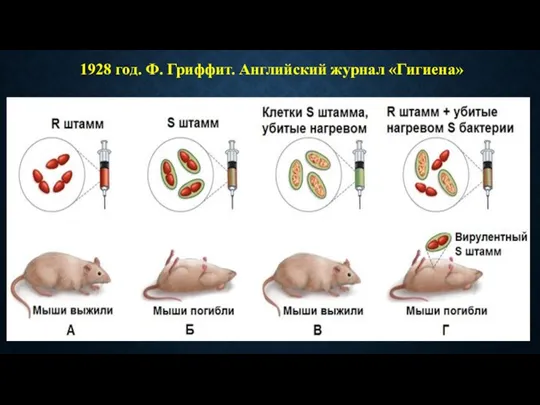
1928 год. Ф. Гриффит. Английский журнал «Гигиена»
1928 год. Ф. Гриффит. Английский журнал «Гигиена»

ТРАНСФОРМАЦИЯ
1 этап – адсорбция выделившийся трансформирующей ДНК из клетки донора на
ТРАНСФОРМАЦИЯ
1 этап – адсорбция выделившийся трансформирующей ДНК из клетки донора на
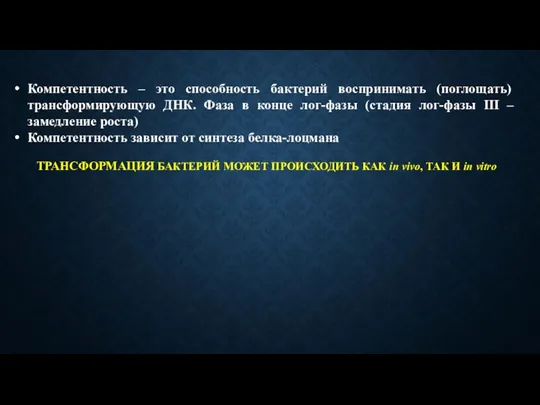
Компетентность – это способность бактерий воспринимать (поглощать) трансформирующую ДНК. Фаза в
Компетентность – это способность бактерий воспринимать (поглощать) трансформирующую ДНК. Фаза в

КОНЪЮГАЦИЯ –
(Ледерберг и Тейтум, 1946)
Передача генетического материала от бактерии-донора
КОНЪЮГАЦИЯ –
(Ледерберг и Тейтум, 1946)
Передача генетического материала от бактерии-донора
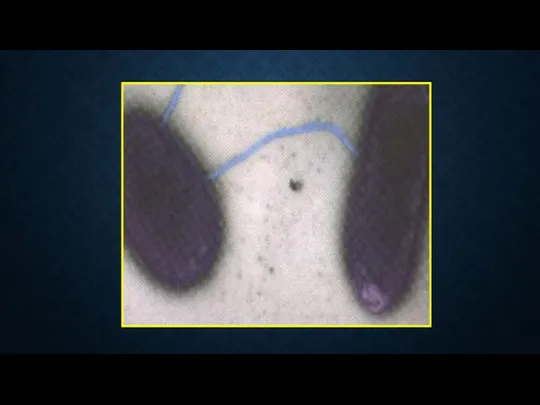

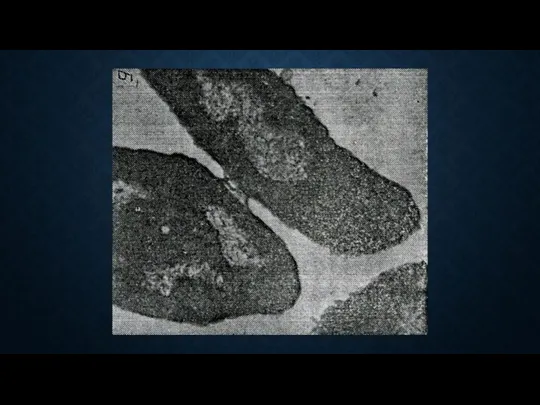
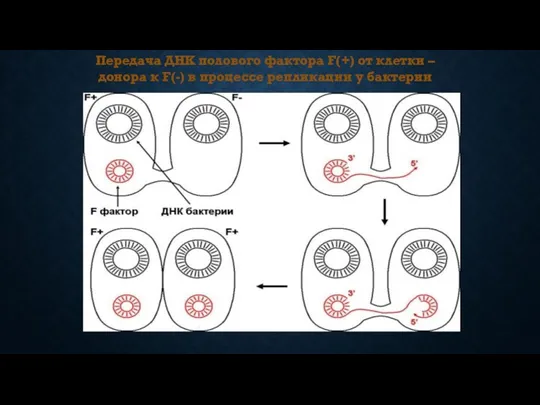
Передача ДНК полового фактора F(+) от клетки – донора к F(-)
Передача ДНК полового фактора F(+) от клетки – донора к F(-)
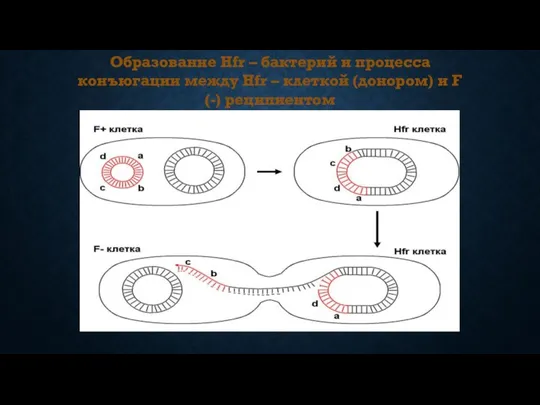
Образование Hfr – бактерий и процесса конъюгации между Hfr – клеткой
Образование Hfr – бактерий и процесса конъюгации между Hfr – клеткой

ПЛАЗМИДЫ и МИГРИРУЮЩИЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ
Плазмиды – небольшие кольцевые двухцепочечные молекулы
ПЛАЗМИДЫ и МИГРИРУЮЩИЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ
Плазмиды – небольшие кольцевые двухцепочечные молекулы

F-ПЛАЗМИДА
F-плазмида. E.coli
Обеспечивают участие в конъюгации между бактериями с помощью кодируемых ими
F-ПЛАЗМИДА
F-плазмида. E.coli
Обеспечивают участие в конъюгации между бактериями с помощью кодируемых ими

БАКТЕРИОЦИНОГЕННАЯ ПЛАЗМИДА
Большинство видов бактерий выделяют белковые антибактериальные вещества – БАКТЕРИОЦИНЫ (бактерицидная
БАКТЕРИОЦИНОГЕННАЯ ПЛАЗМИДА
Большинство видов бактерий выделяют белковые антибактериальные вещества – БАКТЕРИОЦИНЫ (бактерицидная

БАКТЕРИОЦИНОГЕННАЯ ПЛАЗМИДА
Col-плазмида отличается от других плазмид рядом присущих только ей особенностей
В отличие
БАКТЕРИОЦИНОГЕННАЯ ПЛАЗМИДА
Col-плазмида отличается от других плазмид рядом присущих только ей особенностей
В отличие

R-ПЛАЗМИДА (resistance – устойчивость)
РЕЗИСТЕНТНОСТЬ – 3 пути: мутации; рекомбинации; плазмиды с
R-ПЛАЗМИДА (resistance – устойчивость)
РЕЗИСТЕНТНОСТЬ – 3 пути: мутации; рекомбинации; плазмиды с

R-ПЛАЗМИДА (resistance – устойчивость)
RTF
r-гены
2 фрагмента ДНК
Содержит гены, кодирующие его репликацию и
R-ПЛАЗМИДА (resistance – устойчивость)
RTF
r-гены
2 фрагмента ДНК
Содержит гены, кодирующие его репликацию и

R-ПЛАЗМИДА (resistance – устойчивость)
R-ПЛАЗМИДА (resistance – устойчивость)
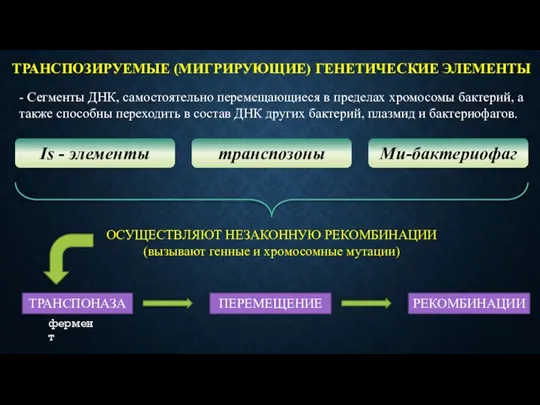
ТРАНСПОЗИРУЕМЫЕ (МИГРИРУЮЩИЕ) ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ
- Сегменты ДНК, самостоятельно перемещающиеся в пределах хромосомы
ТРАНСПОЗИРУЕМЫЕ (МИГРИРУЮЩИЕ) ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ
- Сегменты ДНК, самостоятельно перемещающиеся в пределах хромосомы

Is - элементы
- Это линейные участки ДНК, состоящие из 700-1750 пар
Is - элементы
- Это линейные участки ДНК, состоящие из 700-1750 пар

ФУНКЦИИ IS-ЭЛЕМЕНТОВ
Координирующая - взаимодействие транспозонов, плазмид, умеренных фагов между собой и
ФУНКЦИИ IS-ЭЛЕМЕНТОВ
Координирующая - взаимодействие транспозонов, плазмид, умеренных фагов между собой и

- Подобен Is-элементам и транспозонам, так как может интегрировать в любой
- Подобен Is-элементам и транспозонам, так как может интегрировать в любой
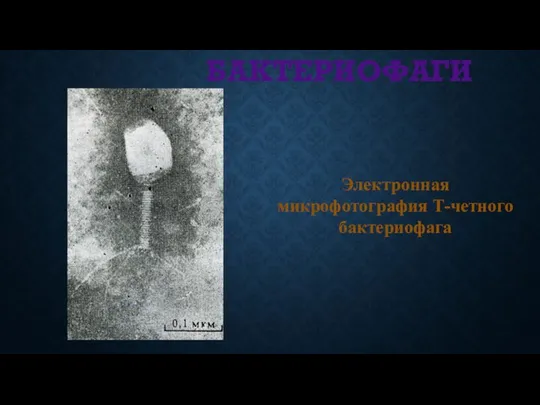
БАКТЕРИОФАГИ
Электронная микрофотография Т-четного бактериофага
БАКТЕРИОФАГИ
Электронная микрофотография Т-четного бактериофага
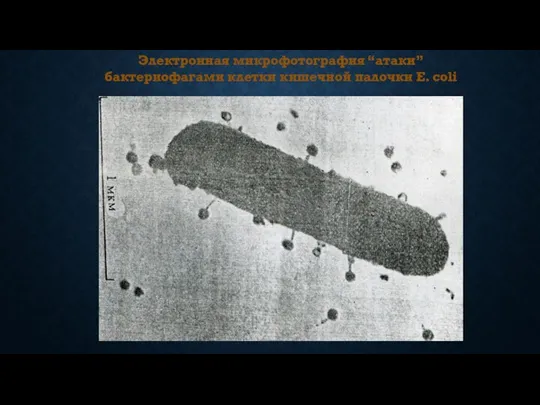
Электронная микрофотография “атаки” бактериофагами клетки кишечной палочки E. coli
Электронная микрофотография “атаки” бактериофагами клетки кишечной палочки E. coli
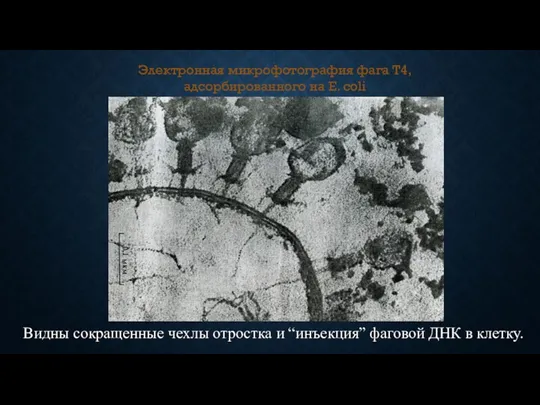
Электронная микрофотография фага T4, адсорбированного на E. coli
Видны сокращенные чехлы отростка
Электронная микрофотография фага T4, адсорбированного на E. coli
Видны сокращенные чехлы отростка

БАКТЕРИОФАГИ ( ОТ ЛАТ.«PHAGOS» - ПОЖИРАЮЩИЙ) – ВИРУСЫ БАКТЕРИЙ, ОБЛАДАЮЩИЕ ТЕМИ
БАКТЕРИОФАГИ ( ОТ ЛАТ.«PHAGOS» - ПОЖИРАЮЩИЙ) – ВИРУСЫ БАКТЕРИЙ, ОБЛАДАЮЩИЕ ТЕМИ

СТРОЕНИЕ БАКТЕРИОФАГОВ.
ПОКОЯЩАЯСЯ, ВНЕКЛЕТОЧНАЯ, ФОРМА – ВИРИОН.
ВНУТРИКЛЕТОЧНАЯ ФОРМА – ВЕГЕТАТИВНАЯ.
ВИРИОН
Головка
белковый
СТРОЕНИЕ БАКТЕРИОФАГОВ.
ПОКОЯЩАЯСЯ, ВНЕКЛЕТОЧНАЯ, ФОРМА – ВИРИОН.
ВНУТРИКЛЕТОЧНАЯ ФОРМА – ВЕГЕТАТИВНАЯ.
ВИРИОН
Головка
белковый

Морфологические типы фагов
I – нитевидные фаги; II – фаги без отростка;
Морфологические типы фагов
I – нитевидные фаги; II – фаги без отростка;
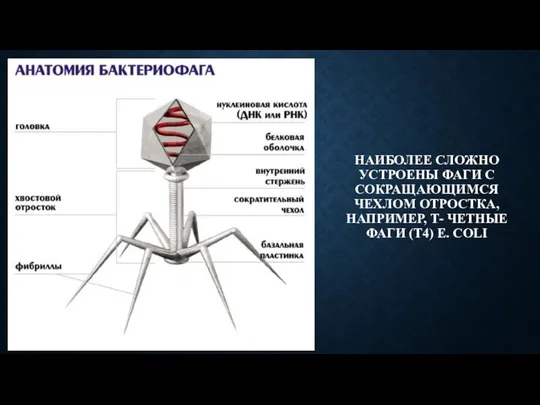
НАИБОЛЕЕ СЛОЖНО УСТРОЕНЫ ФАГИ С СОКРАЩАЮЩИМСЯ ЧЕХЛОМ ОТРОСТКА, НАПРИМЕР, Т- ЧЕТНЫЕ
НАИБОЛЕЕ СЛОЖНО УСТРОЕНЫ ФАГИ С СОКРАЩАЮЩИМСЯ ЧЕХЛОМ ОТРОСТКА, НАПРИМЕР, Т- ЧЕТНЫЕ
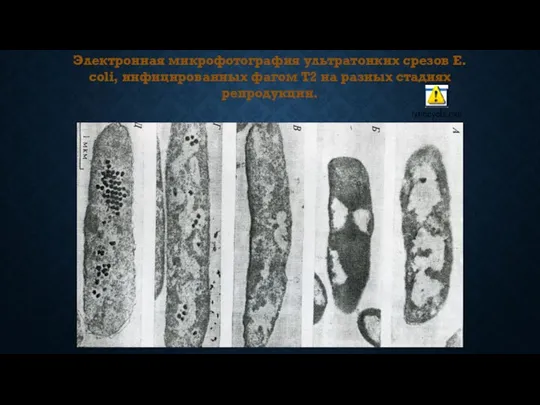
Электронная микрофотография ультратонких срезов E. coli, инфицированных фагом T2 на разных
Электронная микрофотография ультратонких срезов E. coli, инфицированных фагом T2 на разных

ВЗАИМОДЕЙСТВИЕ ФАГА С КЛЕТКОЙ
вирулентные фаги
вызывают продуктивную инфекцию, при которой происходит репродукция
ВЗАИМОДЕЙСТВИЕ ФАГА С КЛЕТКОЙ
вирулентные фаги
вызывают продуктивную инфекцию, при которой происходит репродукция

ЭТАПЫ ПРОДУКТИВНОЙ ИНФЕКЦИИ:
1я стадия. Адсорбция фага на чувствительной клетке. Происходит при
ЭТАПЫ ПРОДУКТИВНОЙ ИНФЕКЦИИ:
1я стадия. Адсорбция фага на чувствительной клетке. Происходит при

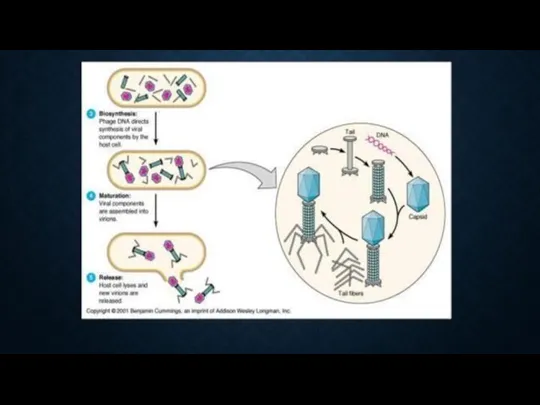
4я стадия. Морфогенез фага. Созревание фага - разобщенный процесс. Отдельно формируются
4я стадия. Морфогенез фага. Созревание фага - разобщенный процесс. Отдельно формируются
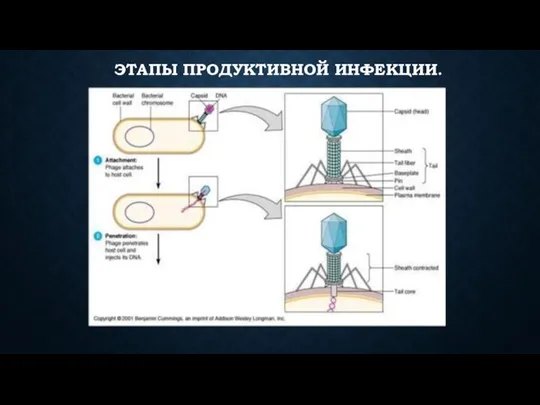
ЭТАПЫ ПРОДУКТИВНОЙ ИНФЕКЦИИ.
ЭТАПЫ ПРОДУКТИВНОЙ ИНФЕКЦИИ.

ИНТЕГРАТИВНАЯ ИНФЕКЦИЯ (ЛИЗОГЕНИЯ)
ДНК фага включается в кольцевую хромосому бактериальной клетки.
ИНТЕГРАТИВНАЯ ИНФЕКЦИЯ (ЛИЗОГЕНИЯ)
ДНК фага включается в кольцевую хромосому бактериальной клетки.
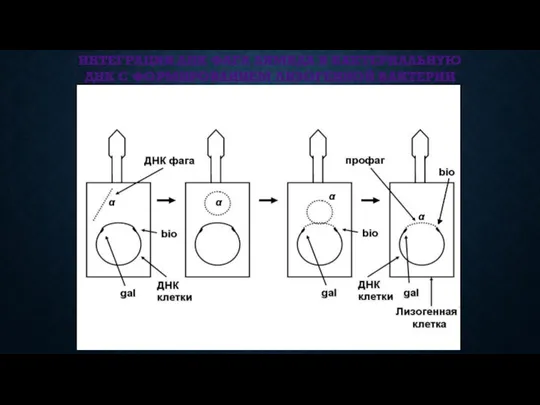
ИНТЕГРАЦИЯ ДНК ФАГА ЛЯМБДА В БАКТЕРИАЛЬНУЮ ДНК С ФОРМИРОВАНИЕМ ЛИЗОГЕННОЙ БАКТЕРИИ
ИНТЕГРАЦИЯ ДНК ФАГА ЛЯМБДА В БАКТЕРИАЛЬНУЮ ДНК С ФОРМИРОВАНИЕМ ЛИЗОГЕННОЙ БАКТЕРИИ

ТРАНСДУКЦИЯ
(ЦИНДЕР И ЛЕДЕРБЕРГ, 1952)
Виды:
общая (генерализированная)
специфическая
абортивная
Вызывают умеренные,
ТРАНСДУКЦИЯ
(ЦИНДЕР И ЛЕДЕРБЕРГ, 1952)
Виды:
общая (генерализированная)
специфическая
абортивная
Вызывают умеренные,
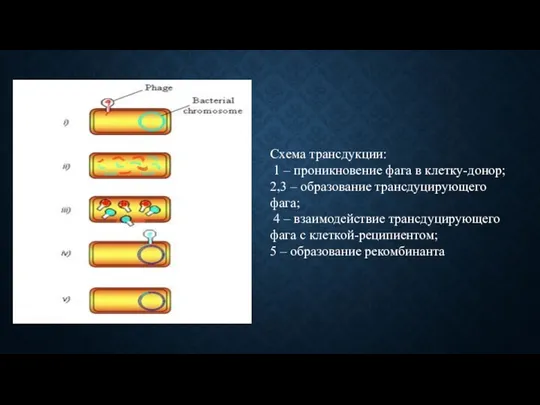
Схема трансдукции:
1 – проникновение фага в клетку-донор;
2,3 – образование
Схема трансдукции:
1 – проникновение фага в клетку-донор;
2,3 – образование
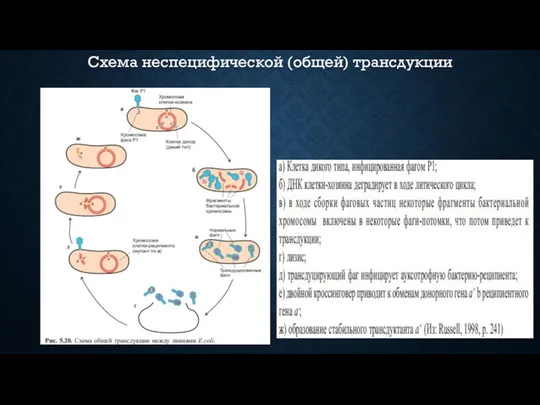
Схема неспецифической (общей) трансдукции
Схема неспецифической (общей) трансдукции
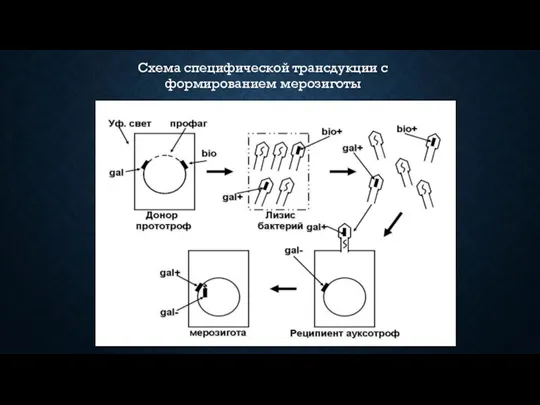
Схема специфической трансдукции с формированием мерозиготы
Схема специфической трансдукции с формированием мерозиготы
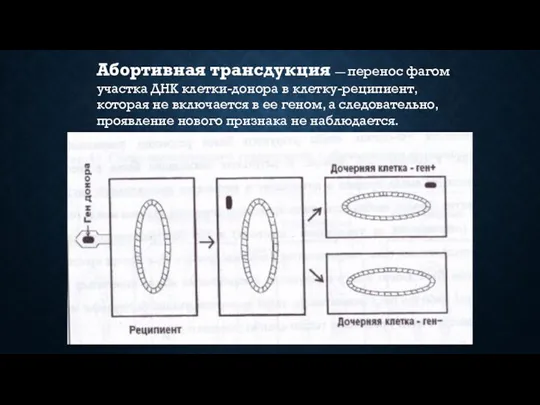
Абортивная трансдукция — перенос фагом участка ДНК клетки-донора в клетку-реципиент, которая
Абортивная трансдукция — перенос фагом участка ДНК клетки-донора в клетку-реципиент, которая

ОТЛИЧИЯ ТРАНСДУКЦИИ И ФАГОВОЙ КОНВЕРСИИ
Трансдукция – перенос генетической информации из клетки
ОТЛИЧИЯ ТРАНСДУКЦИИ И ФАГОВОЙ КОНВЕРСИИ
Трансдукция – перенос генетической информации из клетки
Адсорбция фагов на бактериальной клетке
Адсорбция фагов на бактериальной клетке

ПРИМЕНЕНИЕ БАКТЕРИОФАГОВ.
1. Для диагностики инфекционных заболеваний.
а) для определения видовой принадлежности
ПРИМЕНЕНИЕ БАКТЕРИОФАГОВ.
1. Для диагностики инфекционных заболеваний.
а) для определения видовой принадлежности

ФАГОТИПИРОВАНИЕ
Основа метода: с помощью типовых фагов дифференцируют культуры одного вида
ФАГОТИПИРОВАНИЕ
Основа метода: с помощью типовых фагов дифференцируют культуры одного вида
 Антропогенез. Расы. Расизм. Часть 4
Антропогенез. Расы. Расизм. Часть 4 Краснокнижные виды растений и животных РТ
Краснокнижные виды растений и животных РТ Циркумбореальная область
Циркумбореальная область Фенологические сезоны года
Фенологические сезоны года Почему мы плачем
Почему мы плачем Вегетативне розмноження рослин
Вегетативне розмноження рослин Селекция. Новые открытия селекционеров
Селекция. Новые открытия селекционеров Красные водоросли
Красные водоросли Бездомные животные – проблема каждого из нас
Бездомные животные – проблема каждого из нас Обмен веществ и энергии. Биологическое окисление. Окислительное фосфорилирование и его регуляция. Цикл трикарбоновых кислот
Обмен веществ и энергии. Биологическое окисление. Окислительное фосфорилирование и его регуляция. Цикл трикарбоновых кислот Насекомые, развивающиеся с неполным превращением
Насекомые, развивающиеся с неполным превращением Голосеменные растения
Голосеменные растения Классификация микроорганизмов. Типы взаимоотношений микро- и макроорганизмов
Классификация микроорганизмов. Типы взаимоотношений микро- и макроорганизмов Интерактивный итоговый тест по биологии за 1 полугодие
Интерактивный итоговый тест по биологии за 1 полугодие Сосуды малого и большого кругов кровообращения
Сосуды малого и большого кругов кровообращения מאפייני חיים
מאפייני חיים Гельминтология. Общие черты организации гельминтов
Гельминтология. Общие черты организации гельминтов Двойное оплодотворение
Двойное оплодотворение Всё о кошках
Всё о кошках Кровообращение, лимфообращение
Кровообращение, лимфообращение Урок Трофические взаимоотношения организмов
Урок Трофические взаимоотношения организмов Одноклітинні еукаріоти. Колоніальні організми
Одноклітинні еукаріоти. Колоніальні організми Фотосинтез и дыхание. Презентация 6 класс
Фотосинтез и дыхание. Презентация 6 класс Ферменты. Регуляция активности. Активаторы и ингибиторы
Ферменты. Регуляция активности. Активаторы и ингибиторы Фотосинтез як характерна особливість рослин
Фотосинтез як характерна особливість рослин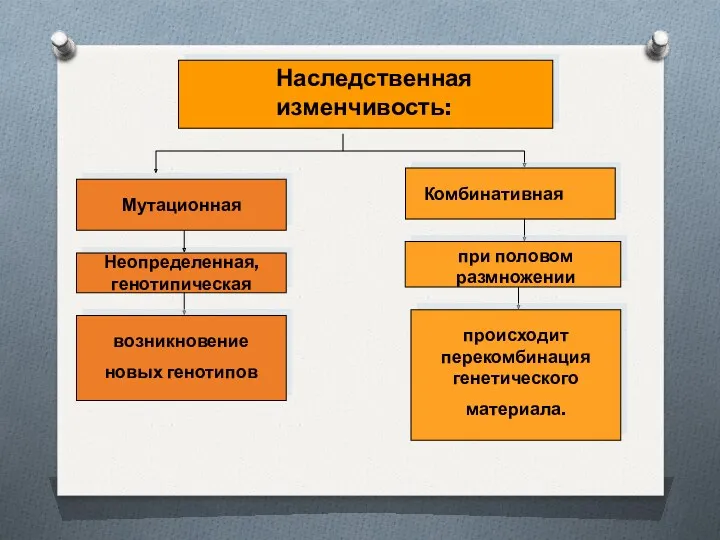 Наследственная изменчивость
Наследственная изменчивость Матричные биосинтезы: блокаторы матричных синтезов, мутации, репарация днк
Матричные биосинтезы: блокаторы матричных синтезов, мутации, репарация днк Углеводы. Лекция 4
Углеводы. Лекция 4