- основы возбудимости

Содержание
- 3. Раздражимость - способность живых систем формировать ответную реакцию на действие раздражителя в виде изменения метаболизма, функций
- 4. Все ткани организма делятся на возбудимые (нервная, мышечная и секреторная) и невозбудимые
- 5. Возбудимость - способность клеток формировать ответную реакцию на действие раздражителя в виде возбуждения. Возбудимость – частный
- 6. Возбуждение - ответная реакция клетки на действие раздражителя, проявляющаяся в специфической для нее функции (проведение возбуждения,
- 7. Возбудимость – это свойство. Возбуждение – это процесс.
- 10. Микроэлектрод - стеклянная трубочка с вытянутым концом. Диаметр кончика микроэлектрода 1-3 мкм, что позволяет проткнуть мембрану
- 11. Мембранный потенциал покоя клетки (МПП) – разность потенциалов между цитоплазмой клетки и межклеточной средой. Величина составляет
- 12. Условия формирования МП. Разность концентраций ионов вне и внутри клетки. Мембрана избирательно проницаема для различных ионов:
- 13. Механизмы формирования МП. Ионный. Насосный.
- 14. Ионный механизм формирования МП (на примере нейрона) .
- 15. внутриклеточная среда Признак созревания – появление на мембране нейрона постоянно открытых К+-каналов (определяется включением соотв. гена).
- 16. внутриклеточная среда межклеточная среда Как долго идет диффузия К+ из нейрона? Очевидный вариант «до выравнивания концентраций»)
- 17. внутриклеточная среда межклеточная среда При диффузии ионов К+ в клетке увеличивается доля отрицательного заряда, а вне
- 18. внутриклеточная среда межклеточная среда Повышающаяся разность потенциалов между внутри- и вне- клеточной средой приводит к появлению
- 19. внутриклеточная среда межклеточная среда ЭДС препятствует диффузии и в конце концов останавливает её. Возникает состояние «динамического
- 20. Вальтер Нернст (Ноб.пр. 1921) «Уравнение Нернста»: Равновесный потенциал Ер R – универсальная газовая постоянная , T-
- 21. внутриклеточная среда межклеточная среда ЭДС В реальной клетке МП находится ближе к нулю (в среднем -70
- 22. внутриклеточная среда межклеточная среда ЭДС Избыток ионов Na+ в межклеточной среде, а также их притяжение к
- 23. внутриклеточная среда межклеточная среда ЭДС Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной
- 24. Насосный механизм формирования МП.
- 25. Электрогенность Na+-K+-АТФазы (Na+-К+-насоса). внутриклеточная среда межклеточная среда
- 26. В целом МП зависит от 3-х главных факторов: - диффузии K+ из клетки; - диффузии Na+
- 27. Потенциал действия
- 28. Потенциал действия – быстрое фазовое закономерное изменение величины мембранного потенциала (МП) , вызванное раздражителем пороговой или
- 29. биоусилитель 0 -50 -70 10 мВ 15 мВ 20 мВ +30 порог запуска ПД КУД 25
- 30. 0 +30 -60 0 2 1 3 4 Время, мс Мембранный потенциал, мВ МП КУД 1
- 31. Деполяризация – уменьшение отрицательной величины МП Реполяризация – увеличение отрицательной величины МП КУД (критический уровень деполяризации)
- 32. 0 +30 -60 0 Время, мс Мембранный потенциал, мВ МП КУД 1 2 3 4 5
- 33. Восходящая фаза (деполяризация): преобладание входящих в клетки Na+токов. Нисходящая фаза (реполяризация): преобладание выходящих из клетки К+токов.
- 34. В основе этих процессов лежит открывание и закрывание каналов. Эти каналы имеют створки, реагирующие на изменение
- 35. + + + + + + ++ -45mV + + + + + +
- 36. Если заряд внутри нейрона вновь ниже -50 мВ – створка закрывается, т.к. положительные заряды, расположенные на
- 37. Открытие потенциалзависимого Na+-канала «разрешает» вход Na+ в клетку. Открытие электрочувствительного К+-канала «разрешает» выход К+ из клетки.
- 38. Na+-каналы открываются очень быстро после стимула и самопроизвольно закрываются примерно через 0.5 мс. К+-каналы открываются медленно
- 39. Именно разная скорость открытия Na+-каналов и К+-каналов позволяет возникнуть сначала восходящей, а затем – нисходящей фазе
- 40. Для закрытия Na+-каналов на пике ПД служит дополнительная (внутриклеточная, инактивационная, И-) створка – h-ворота. Вторая створка
- 41. 1 1 2 3 4 5 1 = 5 = ПП (большая h-створка открыта, ма- лая
- 42. Рефрактерность (от франц . refractaire - невосприимчивый), кратковременный период полного исчезновения или снижения возбудимости нервной и
- 43. 0 +30 -60 0 2 1 3 4 Время, мс Мембранный потенциал, мВ МП КУД Период
- 45. Лабильность (лат. labilitas лабильность, неустойчивость) функциональная подвижность понятие в физиологию ввёл в 1892 г. русский физиолог
- 46. Лабильность Этим понятием в настоящее время обозначается скорость протекания элементарных физиологических реакций (циклов возбуждения в нервной
- 47. Лабильность Мерой является наибольшая частота раздражения ткани, воспроизводимая ею без преобразования ритма.
- 48. Лабильность отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла возбуждения.
- 49. Итак, мы познакомились с общими принципами генера- ции ПД. Следующие три вопроса: [1]. Что будет, если
- 50. тетродотоксин – яд рыбы фугу (аминогруппа работает как «пробка» для Na+-канала) В результате действия токсина прекращается
- 51. ТЕА – тетраэтиламмоний: работает как «пробка» по отношению к К+-каналу. В результате восходящая фаза ПД изменяется
- 52. Этот рисунок – из предыдущей лекции. Однако он подходит для иллюстрации как вклада Na+-K+-АТФазы в поддержание
- 55. Скачать презентацию


Раздражимость -
способность живых систем
формировать ответную реакцию на действие раздражителя
Раздражимость -
способность живых систем
формировать ответную реакцию на действие раздражителя
Все ткани организма делятся на возбудимые (нервная, мышечная и секреторная) и
Все ткани организма делятся на возбудимые (нервная, мышечная и секреторная) и

Возбудимость - способность клеток формировать ответную реакцию на действие раздражителя в
Возбудимость - способность клеток формировать ответную реакцию на действие раздражителя в

Возбуждение - ответная реакция клетки на действие раздражителя,
проявляющаяся в специфической
Возбуждение - ответная реакция клетки на действие раздражителя,
проявляющаяся в специфической

Возбудимость –
это свойство.
Возбуждение –
это процесс.
Возбудимость –
это свойство.
Возбуждение –
это процесс.
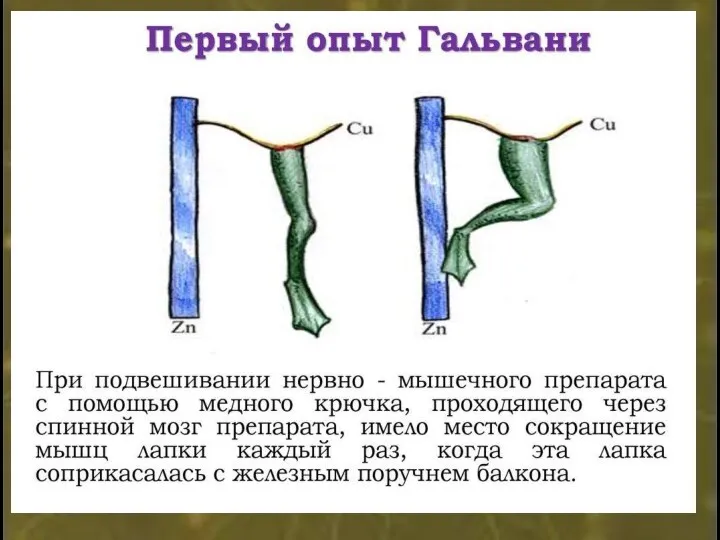
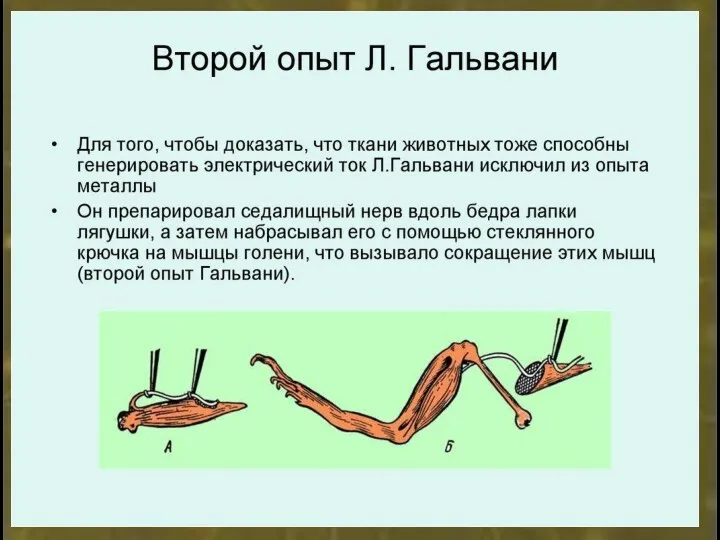
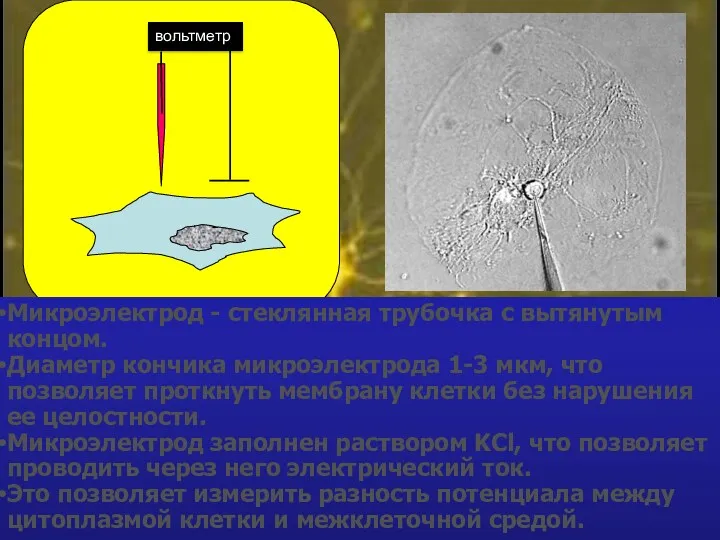
Микроэлектрод - стеклянная трубочка с вытянутым концом.
Диаметр кончика микроэлектрода 1-3
Микроэлектрод - стеклянная трубочка с вытянутым концом.
Диаметр кончика микроэлектрода 1-3
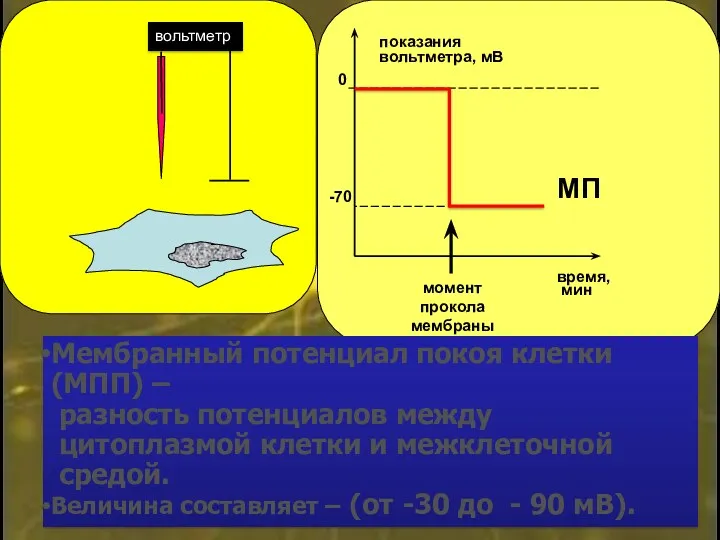
Мембранный потенциал покоя клетки (МПП) –
разность потенциалов между
цитоплазмой
Мембранный потенциал покоя клетки (МПП) –
разность потенциалов между
цитоплазмой

Условия формирования МП.
Разность концентраций ионов вне и внутри клетки.
Мембрана избирательно проницаема
Условия формирования МП.
Разность концентраций ионов вне и внутри клетки.
Мембрана избирательно проницаема

Механизмы формирования МП.
Ионный.
Насосный.
Механизмы формирования МП.
Ионный.
Насосный.
Ионный механизм формирования МП
(на примере нейрона) .
Ионный механизм формирования МП
(на примере нейрона) .
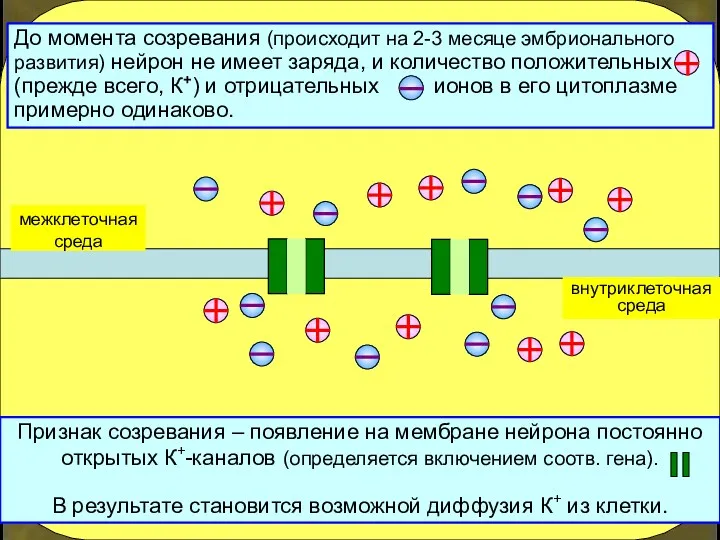
внутриклеточная
среда
Признак созревания – появление на мембране нейрона постоянно открытых К+-каналов (определяется
внутриклеточная
среда
Признак созревания – появление на мембране нейрона постоянно открытых К+-каналов (определяется
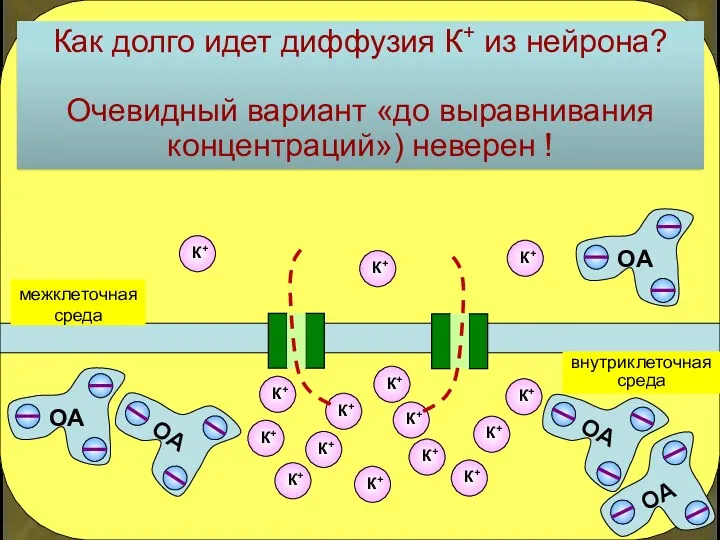
внутриклеточная
среда
межклеточная
среда
Как долго идет диффузия К+ из нейрона?
Очевидный вариант «до выравнивания концентраций»)
внутриклеточная
среда
межклеточная
среда
Как долго идет диффузия К+ из нейрона?
Очевидный вариант «до выравнивания концентраций»)
внутриклеточная
среда
межклеточная
среда
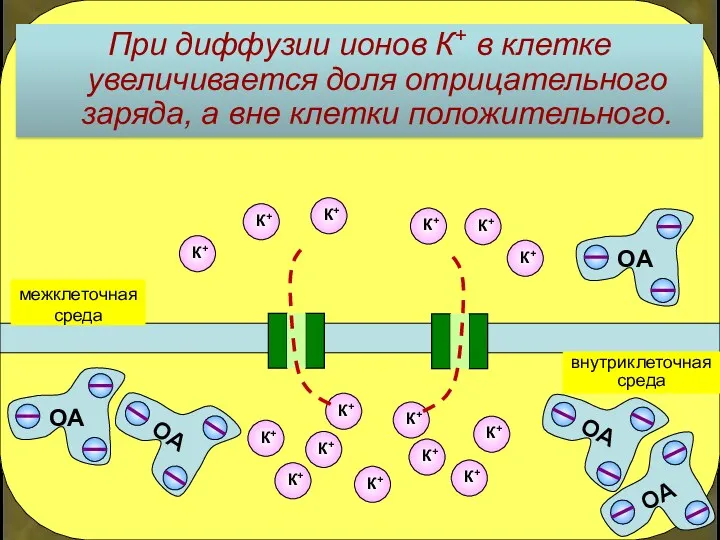
При диффузии ионов К+ в клетке увеличивается доля отрицательного заряда, а
внутриклеточная
среда
межклеточная
среда
При диффузии ионов К+ в клетке увеличивается доля отрицательного заряда, а
внутриклеточная
среда
межклеточная
среда
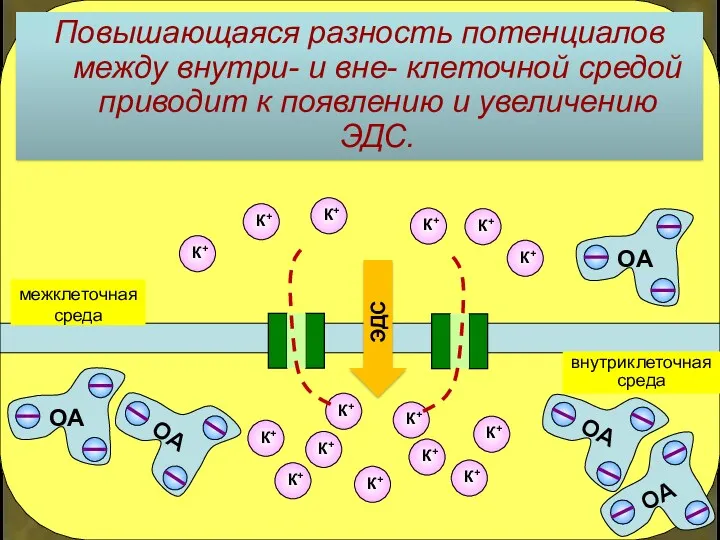
Повышающаяся разность потенциалов между внутри- и вне- клеточной средой приводит к
внутриклеточная
среда
межклеточная
среда
Повышающаяся разность потенциалов между внутри- и вне- клеточной средой приводит к
внутриклеточная
среда
межклеточная
среда
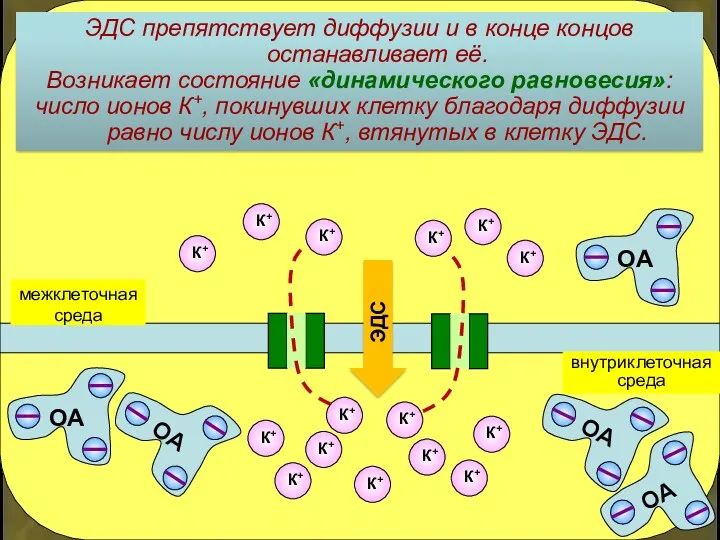
ЭДС препятствует диффузии и в конце концов останавливает её.
Возникает состояние
внутриклеточная
среда
межклеточная
среда
ЭДС препятствует диффузии и в конце концов останавливает её.
Возникает состояние

Вальтер Нернст
(Ноб.пр. 1921)
«Уравнение Нернста»:
Равновесный потенциал Ер
R – универсальная газовая
Вальтер Нернст
(Ноб.пр. 1921)
«Уравнение Нернста»:
Равновесный потенциал Ер
R – универсальная газовая
внутриклеточная
среда
межклеточная
среда
ЭДС
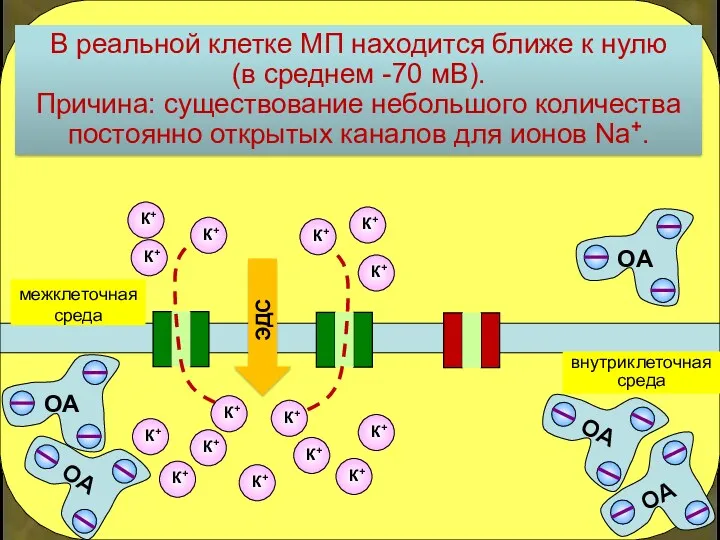
В реальной клетке МП находится ближе к нулю
(в среднем -70
внутриклеточная
среда
межклеточная
среда
ЭДС
В реальной клетке МП находится ближе к нулю
(в среднем -70
внутриклеточная
среда
межклеточная
среда
ЭДС
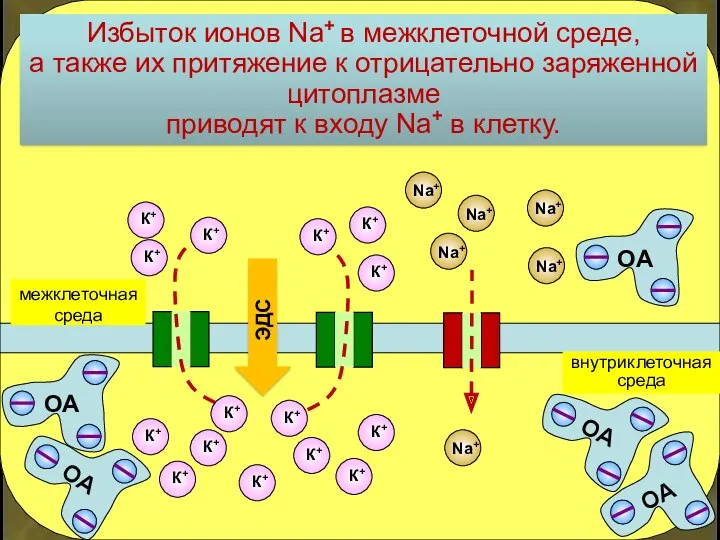
Избыток ионов Na+ в межклеточной среде,
а также их притяжение к
внутриклеточная
среда
межклеточная
среда
ЭДС
Избыток ионов Na+ в межклеточной среде,
а также их притяжение к
внутриклеточная
среда
межклеточная
среда
ЭДС
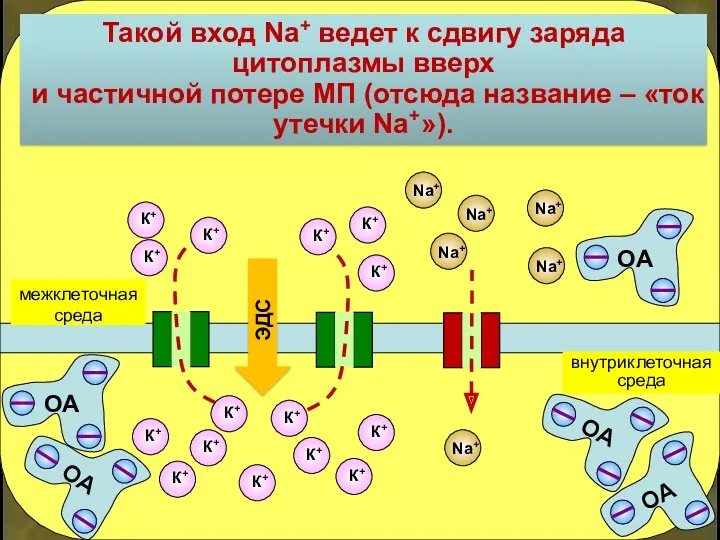
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
внутриклеточная
среда
межклеточная
среда
ЭДС
Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх
и частичной потере
Насосный механизм формирования МП.
Насосный механизм формирования МП.
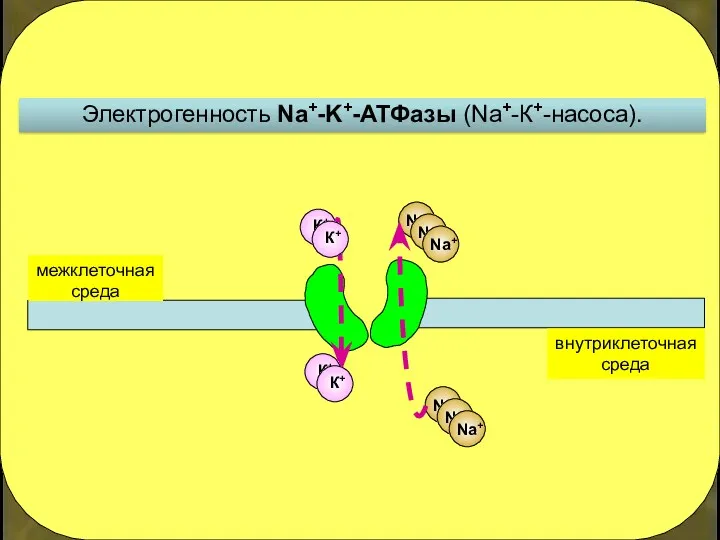
Электрогенность Na+-K+-АТФазы (Na+-К+-насоса).
внутриклеточная
среда
межклеточная
среда
Электрогенность Na+-K+-АТФазы (Na+-К+-насоса).
внутриклеточная
среда
межклеточная
среда

В целом МП зависит от 3-х главных факторов:
- диффузии K+ из
В целом МП зависит от 3-х главных факторов:
- диффузии K+ из
Потенциал действия
Потенциал действия
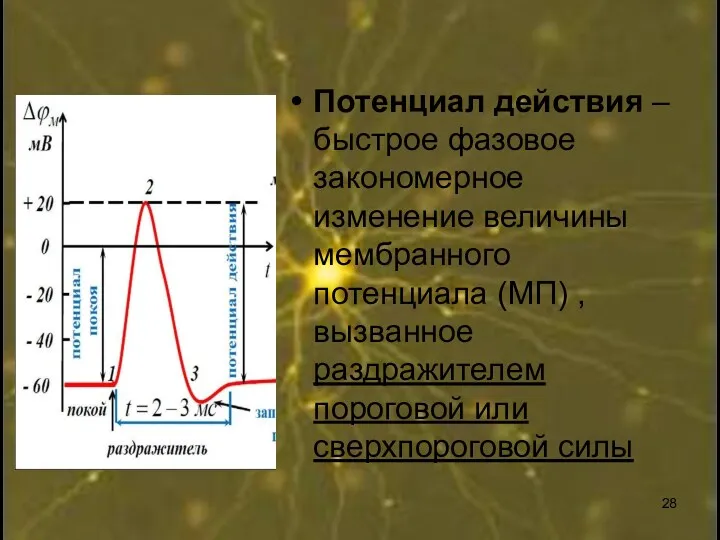
Потенциал действия – быстрое фазовое закономерное изменение величины мембранного потенциала (МП)
Потенциал действия – быстрое фазовое закономерное изменение величины мембранного потенциала (МП)
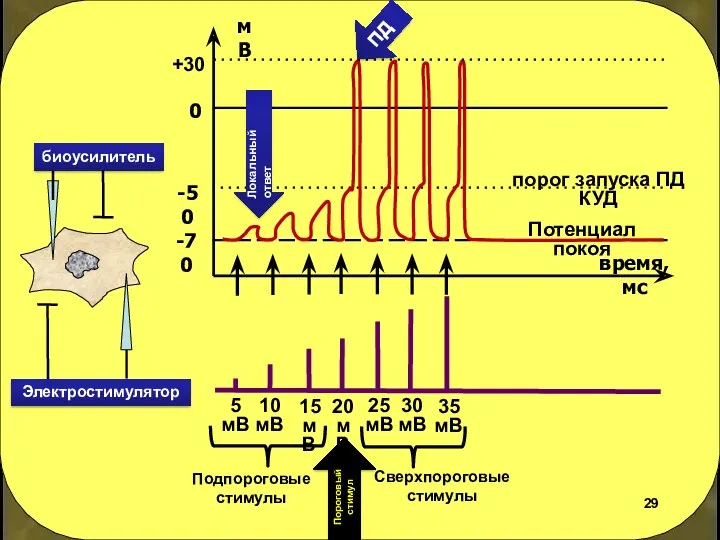
биоусилитель
0
-50
-70
10
мВ
15
мВ
20
мВ
+30
порог запуска ПД
КУД
25
мВ
5
мВ
30
мВ
35
мВ
Подпороговые стимулы
Сверхпороговые стимулы
Пороговый стимул
ПД
Локальный ответ
Потенциал покоя
Электростимулятор
биоусилитель
0
-50
-70
10
мВ
15
мВ
20
мВ
+30
порог запуска ПД
КУД
25
мВ
5
мВ
30
мВ
35
мВ
Подпороговые стимулы
Сверхпороговые стимулы
Пороговый стимул
ПД
Локальный ответ
Потенциал покоя
Электростимулятор
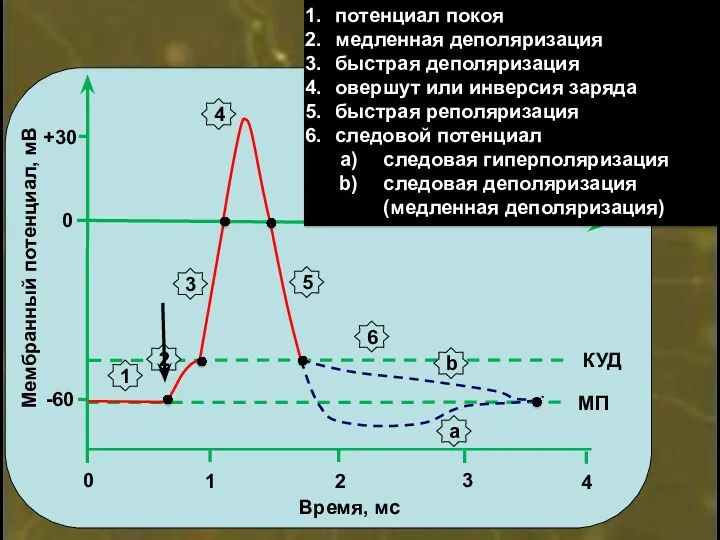
0
+30
-60
0
2
1
3
4
Время, мс
Мембранный потенциал, мВ
МП
КУД
1
2
3
4
5
6
потенциал покоя
медленная деполяризация
быстрая деполяризация
овершут
0
+30
-60
0
2
1
3
4
Время, мс
Мембранный потенциал, мВ
МП
КУД
1
2
3
4
5
6
потенциал покоя
медленная деполяризация
быстрая деполяризация
овершут

Деполяризация – уменьшение отрицательной величины МП
Реполяризация – увеличение отрицательной величины МП
Деполяризация – уменьшение отрицательной величины МП
Реполяризация – увеличение отрицательной величины МП
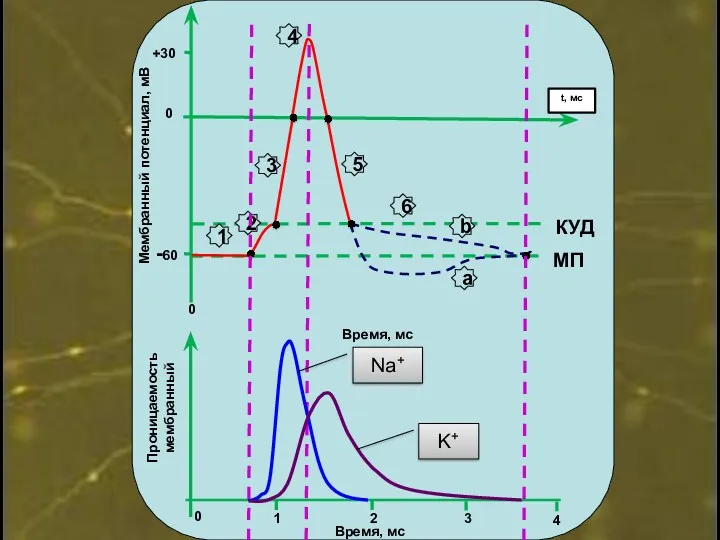
0
+30
-60
0
Время, мс
Мембранный потенциал, мВ
МП
КУД
1
2
3
4
5
6
a
b
0
2
1
3
4
Время, мс
Проницаемость
мембранный
Na+
K+
t, мс
0
+30
-60
0
Время, мс
Мембранный потенциал, мВ
МП
КУД
1
2
3
4
5
6
a
b
0
2
1
3
4
Время, мс
Проницаемость
мембранный
Na+
K+
t, мс

Восходящая фаза (деполяризация):
преобладание входящих в клетки Na+токов.
Нисходящая фаза (реполяризация):
преобладание выходящих
Восходящая фаза (деполяризация):
преобладание входящих в клетки Na+токов.
Нисходящая фаза (реполяризация):
преобладание выходящих

В основе этих процессов лежит открывание
и закрывание каналов.
Эти каналы имеют створки,
В основе этих процессов лежит открывание
и закрывание каналов.
Эти каналы имеют створки,
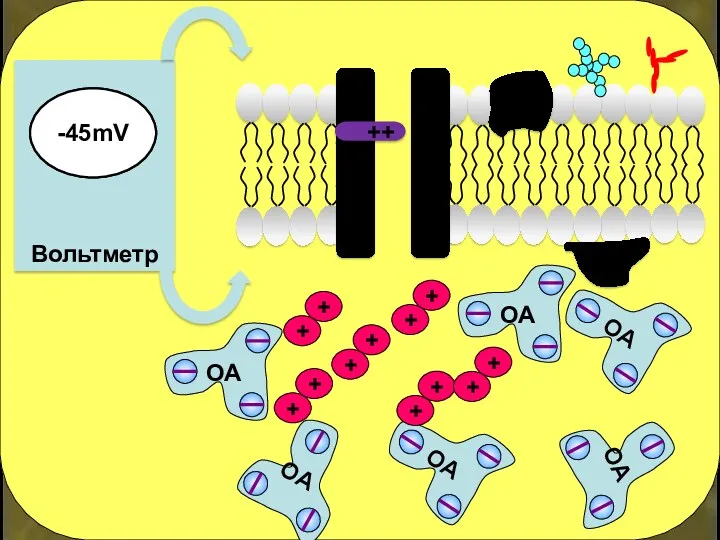
+
+
+
+
+
+
++
-45mV
+
+
+
+
+
+
+
+
+
+
+
+
++
-45mV
+
+
+
+
+
+

Если заряд внутри нейрона вновь ниже -50 мВ – створка закрывается,
Если заряд внутри нейрона вновь ниже -50 мВ – створка закрывается,

Открытие потенциалзависимого Na+-канала «разрешает» вход Na+ в клетку.
Открытие электрочувствительного
К+-канала
Открытие потенциалзависимого Na+-канала «разрешает» вход Na+ в клетку.
Открытие электрочувствительного
К+-канала

Na+-каналы открываются
очень быстро после стимула и
самопроизвольно закрываются примерно через 0.5
очень быстро после стимула и
самопроизвольно закрываются примерно через 0.5

Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем
Именно разная скорость открытия
Na+-каналов и К+-каналов позволяет
возникнуть сначала восходящей, а
затем

Для закрытия Na+-каналов на пике ПД служит дополнительная
(внутриклеточная, инактивационная, И-)
(внутриклеточная, инактивационная, И-)
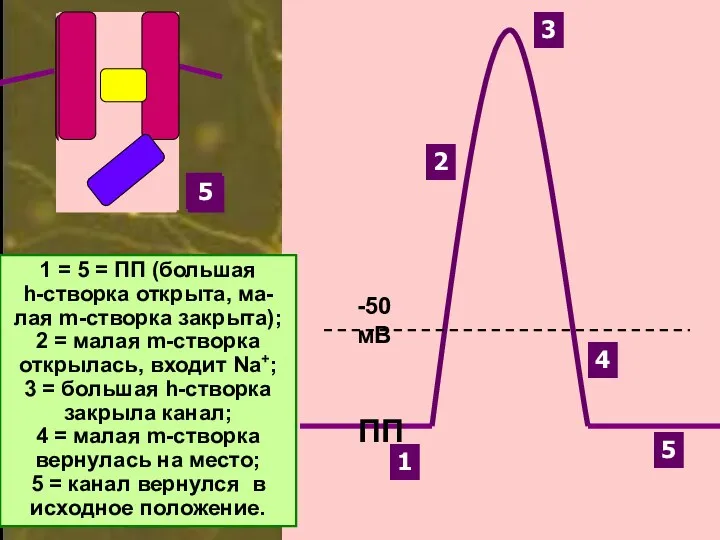
1
1
2
3
4
5
1 = 5 = ПП (большая
h-створка открыта, ма-
лая m-створка закрыта);
2
1
1
2
3
4
5
1 = 5 = ПП (большая
h-створка открыта, ма-
лая m-створка закрыта);
2

Рефрактерность
(от франц . refractaire - невосприимчивый),
кратковременный период полного исчезновения или
Рефрактерность
(от франц . refractaire - невосприимчивый),
кратковременный период полного исчезновения или
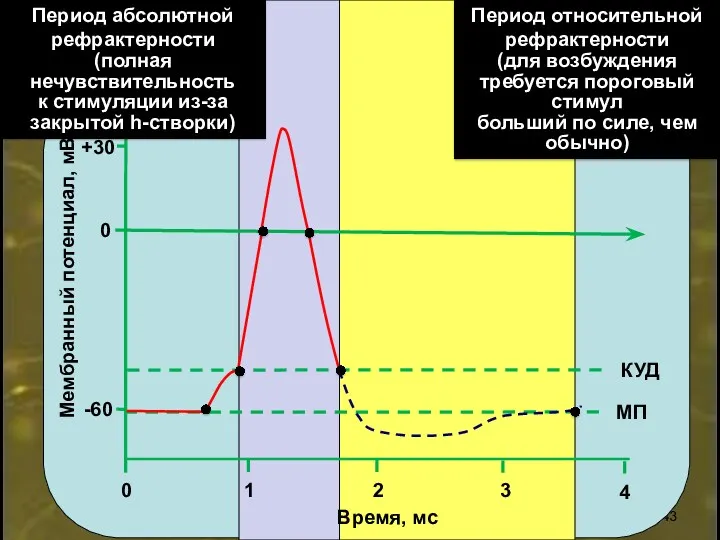
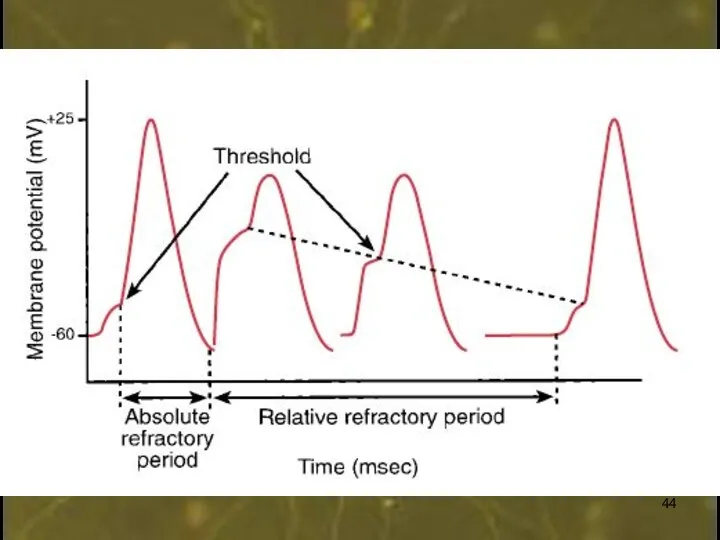
0
+30
-60
0
2
1
3
4
Время, мс
Мембранный потенциал, мВ
МП
КУД
Период абсолютной рефрактерности
(полная
нечувствительность
к стимуляции из-за
закрытой h-створки)
Период относительной
0
+30
-60
0
2
1
3
4
Время, мс
Мембранный потенциал, мВ
МП
КУД
Период абсолютной рефрактерности
(полная
нечувствительность
к стимуляции из-за
закрытой h-створки)
Период относительной

Лабильность
(лат. labilitas лабильность, неустойчивость)
функциональная подвижность
понятие в физиологию
Лабильность
(лат. labilitas лабильность, неустойчивость)
функциональная подвижность
понятие в физиологию

Лабильность
Этим понятием в настоящее время обозначается скорость протекания элементарных физиологических реакций
Лабильность
Этим понятием в настоящее время обозначается скорость протекания элементарных физиологических реакций

Лабильность
Мерой является наибольшая частота раздражения ткани, воспроизводимая ею без преобразования ритма.
Лабильность
Мерой является наибольшая частота раздражения ткани, воспроизводимая ею без преобразования ритма.

Лабильность
отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла
Лабильность
отражает время, в течение которого ткань восстанавливает работоспособность после очередного цикла

Итак, мы познакомились с общими принципами генера-
ции ПД. Следующие три вопроса:
[1].
Итак, мы познакомились с общими принципами генера-
ции ПД. Следующие три вопроса:
[1].
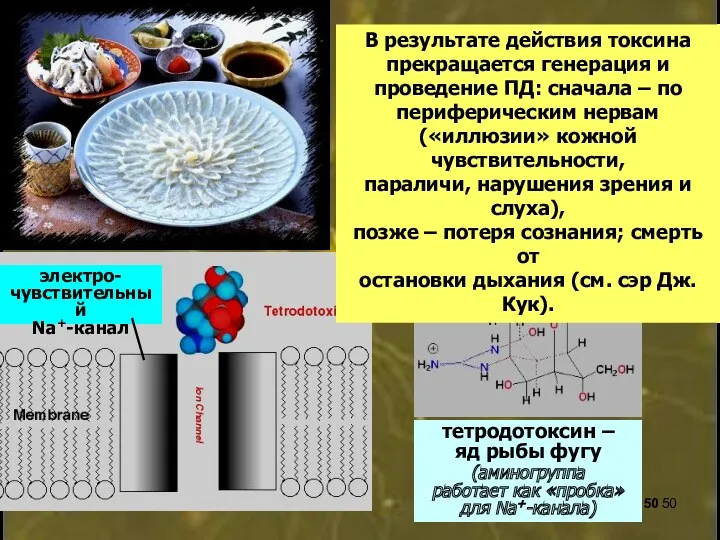
тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
В результате действия токсина прекращается
тетродотоксин –
яд рыбы фугу
(аминогруппа
работает как «пробка»
для Na+-канала)
В результате действия токсина прекращается
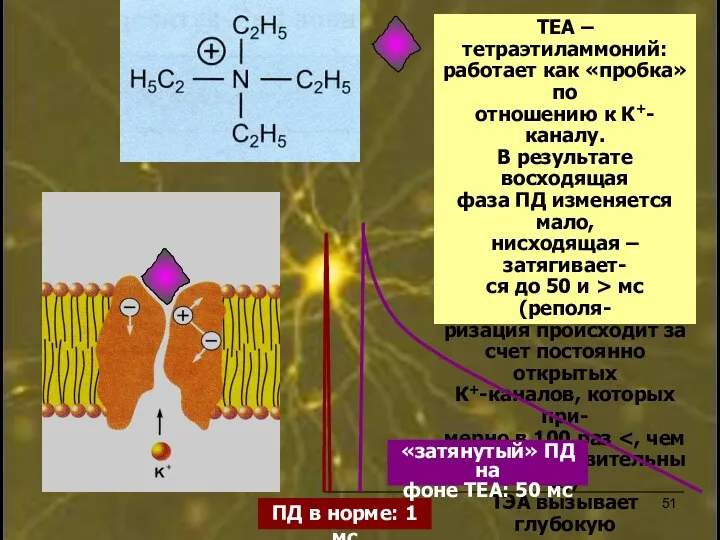
ТЕА – тетраэтиламмоний:
работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД
ТЕА – тетраэтиламмоний:
работает как «пробка» по
отношению к К+-каналу.
В результате восходящая
фаза ПД
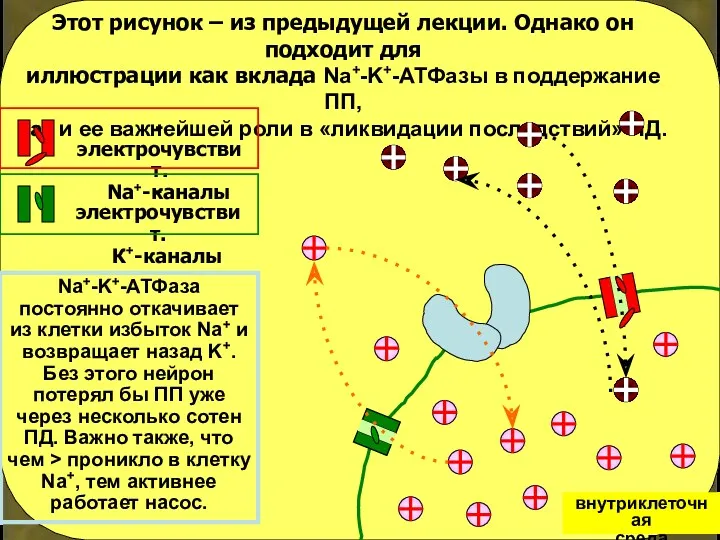
Этот рисунок – из предыдущей лекции. Однако он подходит для
иллюстрации как
Этот рисунок – из предыдущей лекции. Однако он подходит для
иллюстрации как
 Цікаві факти про метеликів: швидше, далі, яскравіше
Цікаві факти про метеликів: швидше, далі, яскравіше Движущие силы эволюции по Дарвину. Формы естественного отбора
Движущие силы эволюции по Дарвину. Формы естественного отбора История представлений о развитии жизни на Земле
История представлений о развитии жизни на Земле Генетика пола. Наследование сцепленное с полом
Генетика пола. Наследование сцепленное с полом Генетически модифицированные организмы (ГМО)
Генетически модифицированные организмы (ГМО) Многообразие природы родного края. Краснодарский край
Многообразие природы родного края. Краснодарский край Самые редкие и необычные породы кошек
Самые редкие и необычные породы кошек Суарудың гидропоника әдісі
Суарудың гидропоника әдісі Органоиды цитоплазмы: клеточный центр, органоиды движения. Включения
Органоиды цитоплазмы: клеточный центр, органоиды движения. Включения Размножение. Начало новой жизни
Размножение. Начало новой жизни Типы наследования признаков у человека
Типы наследования признаков у человека Большой практикум. Опорно-двигательная система
Большой практикум. Опорно-двигательная система Биосинтез белка
Биосинтез белка Биология 9 классы 15.04 Кочетова М И
Биология 9 классы 15.04 Кочетова М И Использование современного оборудования в образовательном процессе
Использование современного оборудования в образовательном процессе ГОСТы на посадочный материал
ГОСТы на посадочный материал Соединения костей
Соединения костей Плоды. Строение плода. Классификация плодов
Плоды. Строение плода. Классификация плодов Методы биологических исследований
Методы биологических исследований Генная инженерия растений
Генная инженерия растений Закономерности индивидуального развития опорно-двигательного аппарата
Закономерности индивидуального развития опорно-двигательного аппарата Анатомия. Мышцы
Анатомия. Мышцы Онтогенез. 10 класс
Онтогенез. 10 класс Основы сравнительной эмбриологии
Основы сравнительной эмбриологии Сколько царств живой природы?
Сколько царств живой природы? Живая и неживая природа
Живая и неживая природа Биология клетки. Лекция 1
Биология клетки. Лекция 1 Насекомые
Насекомые