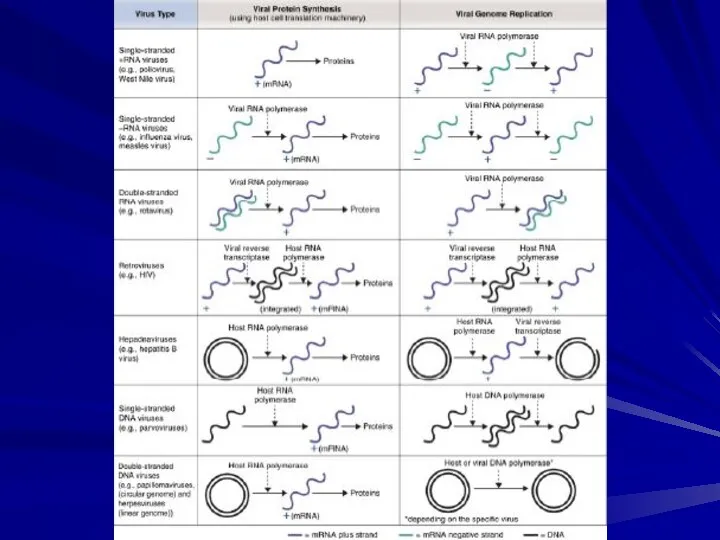
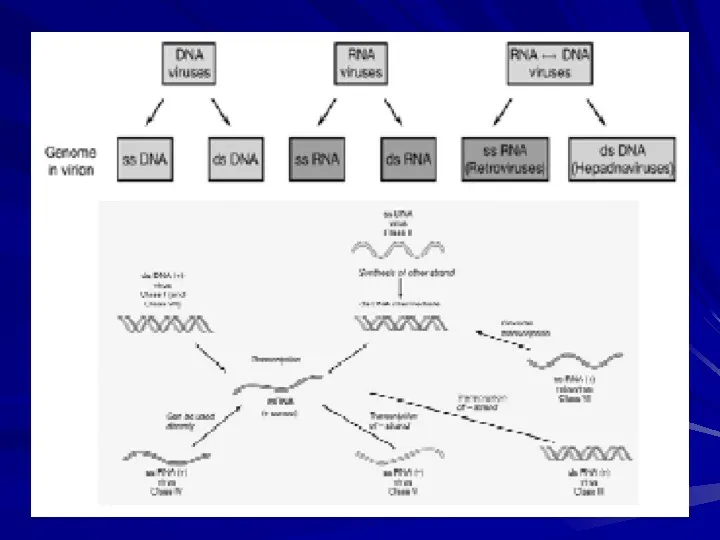
- Параретровіруси. Pararetroviruses vs. Retroviruses

Содержание
- 2. Pararetroviruses vs. Retroviruses
- 3. Вірус гепатиту В
- 4. Життєвий цикл ВГВ
- 5. Життєвий цикл ВГВ
- 6. Вірус гепатиту В ВІРУС ГЕПАТИТУ В, ВГВ (Hepatitis В virus) - збудник гепатиту В, основний представник
- 7. 7 генотипів (A – G) 9 серотипів (ayw1, ayw2, ayw3, ayw4, ayr, adw2, adw4, adrq+, adrq-)
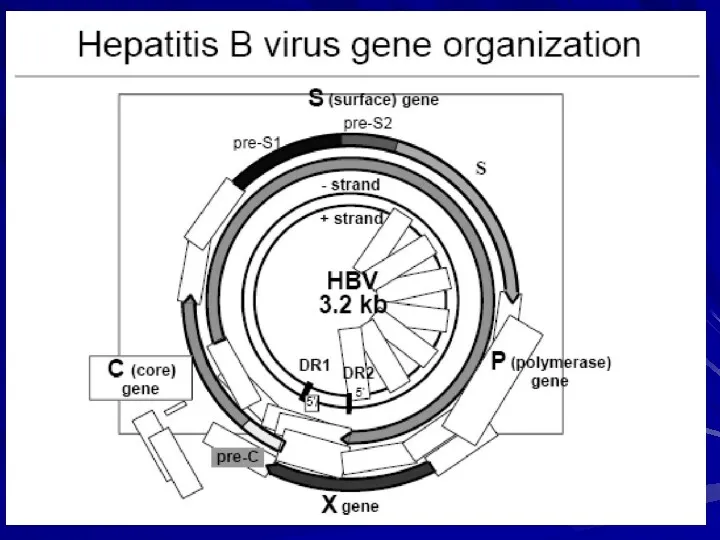
- 8. Геном ВГВ представлений дволанцюговою кільцевою молекулою ДНК — однією з найменших зі всієї нині ідентифікованої ДНК
- 10. Геном вірусу гепатиту В
- 11. Ген Р охоплює обширну зону протяжністю приблизно в 840—850 нуклеотидів, кодуючи білок з молекулярною масою 25000,
- 12. Ген С, кодує білок нуклеокапсида – HBcAg, що складається з 183-185 амінокислот. Перед геном С розташована
- 13. Precore Core X RNase H 5’ RNA Primer DR2 DR1 (+) (–) protein acer Poly EcoR1
- 14. Транскрипційно/Трансляційнаорганізація 4 major RNA species ---------- ~ 7 Proteins 3.5 kb Pre-C/C Pol Pregenomic 2.4 kb
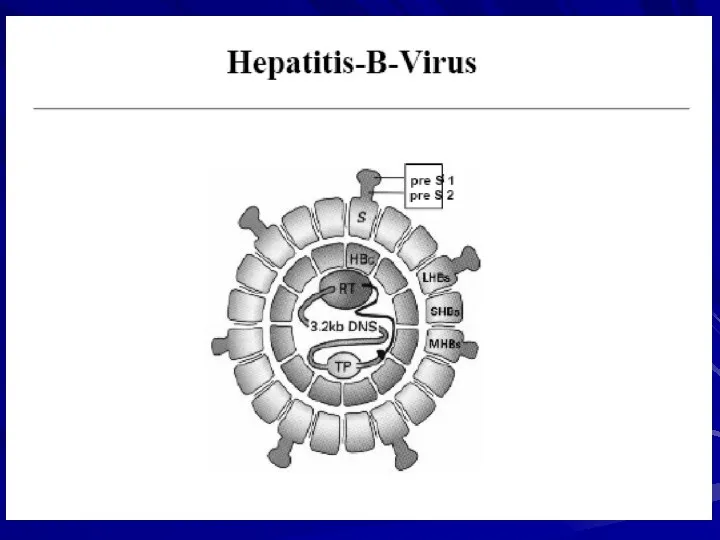
- 15. Viral Gene Products S gene: Surface, Envelope, HBsAg Envelope Small, medium and large Glycosylation C gene:
- 17. Реплікація генома вірусу гепатиту В починається з проникнення віріона в гепатоцит з руйнуванням зовнішньої оболонки частинки
- 18. Transcription of Hepatitis B virus
- 20. Регуляція синтезу білків ВГВ Синтез білків вірусу гепатиту В регулюється на рівні транскрипції і трансляції. Підсилювачі
- 21. Механізм зворотньої транскрипції ВГВ Прегеномна РНК (штрихова лінія, крок 1) кепована і поліаденільована та має довгу
- 22. Реплікація HBV та мішені дії різних терапевтичних препаратів
- 23. Мутації в геномі HBV приводять до появи резистентних штамів Observed in ADV-treated patients Terminal protein Spacer
- 24. Caulimoviridae
- 25. Зліва. Реконструкція поверхнева структура вірусу (CAMV) мозаїки цвітної капусти, що показує T = 7 симетрію. Справа.
- 26. Електронні мікрознімки тонких зрізів caulimovirus-інфікованої тканини, що показує частинки вірусу 42-46 nm і включення. (Photos courtesy
- 27. Caulimoviridae Ізометричні 50 nm частинки з T=7 або бацилярні частинки; Дл ДНК геном ~ 8кб Перший
- 28. Caulimoviridae Переважно вузьке коло господарів Переважно малозначні як патогени; виключення - Rice tungro bacilliform virus ,
- 29. Життєвий цикл Caulimovirus Вірус потрапляє до клітини, капсидний білок дисоціює длДНК мігрує в ядро; транскрибується 35S
- 30. Cauliflower mosaic virus genome structure Сім ORFs в геномі CaMV Транслюється 7 білків з 2х транскриптів
- 31. Cauliflower mosaic virus (CaMV) Кожен ланцюг нуклеїнової кислоти має розриви у специфічних місцях. Перший ланцюг –
- 32. CaMV Genome
- 33. Genome Organization of Caulimoviridae CERV CsVMV PVCV Petuvirus Caulimoviruses CaMV FMV Soymoviruses BRRV SbCMV Cavemoviruses Tungroviruses
- 34. Після потрапляння у клітину розриви в геномі «замуровуються» з подальшим формуванням суперспіралізованої ДНК, що створює мініхромосоми
- 35. Hepadnavirus & Caulimovirus vs. Retrovirus RT Replication
- 36. Hepadnaviruses and Caulimoviruses: DNA Genomes Reverse Transcribed from a +RNA Template From Flint et al. Principles
- 40. Скачать презентацию

Pararetroviruses vs. Retroviruses
Pararetroviruses vs. Retroviruses
Вірус гепатиту В
Вірус гепатиту В
Життєвий цикл ВГВ
Життєвий цикл ВГВ
Життєвий цикл ВГВ
Життєвий цикл ВГВ
Вірус гепатиту В
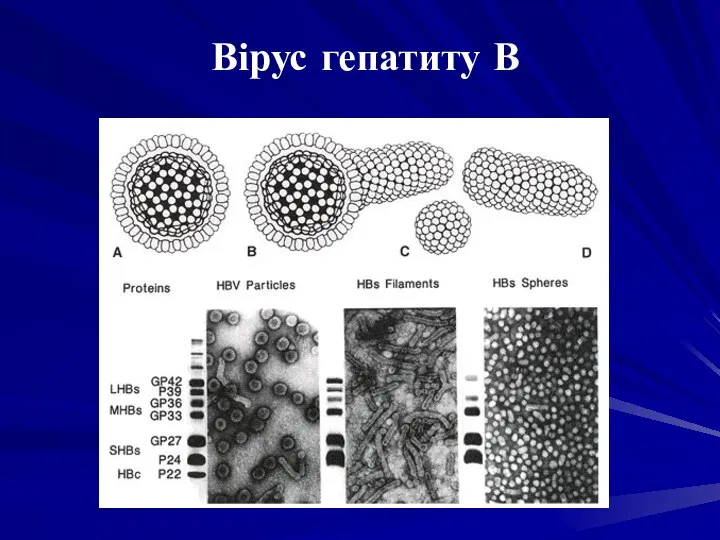
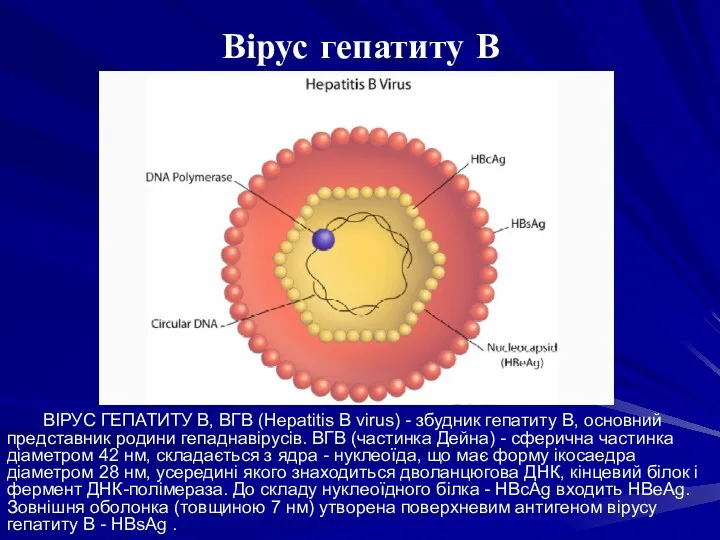
ВІРУС ГЕПАТИТУ В, ВГВ (Hepatitis В virus) - збудник
Вірус гепатиту В
ВІРУС ГЕПАТИТУ В, ВГВ (Hepatitis В virus) - збудник

7 генотипів (A – G)
9 серотипів (ayw1, ayw2, ayw3,
7 генотипів (A – G)
9 серотипів (ayw1, ayw2, ayw3,
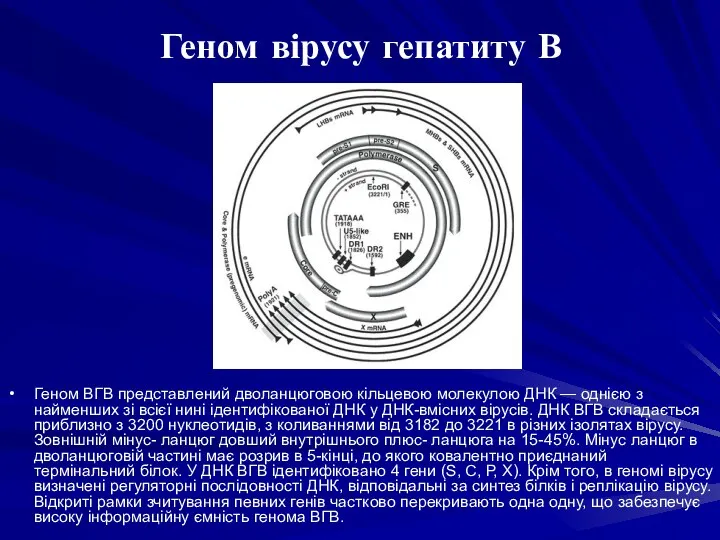
Геном ВГВ представлений дволанцюговою кільцевою молекулою ДНК — однією з найменших
Геном ВГВ представлений дволанцюговою кільцевою молекулою ДНК — однією з найменших
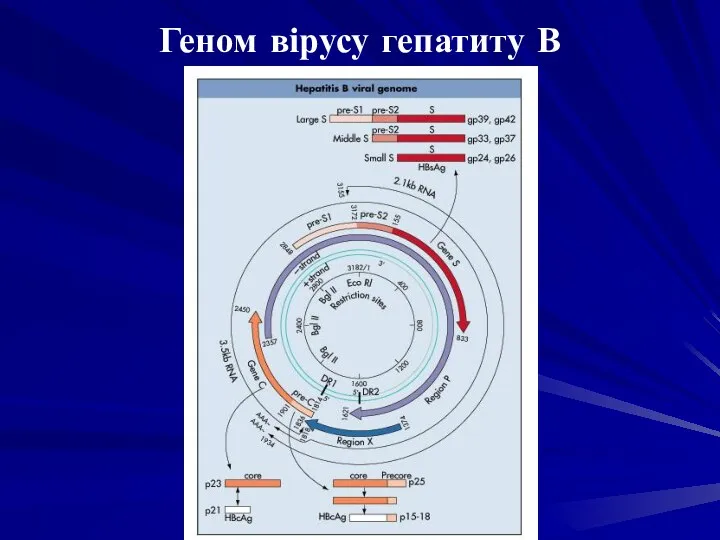
Геном вірусу гепатиту В
Геном вірусу гепатиту В

Ген Р охоплює обширну зону протяжністю приблизно в 840—850 нуклеотидів, кодуючи
Ген Р охоплює обширну зону протяжністю приблизно в 840—850 нуклеотидів, кодуючи

Ген С, кодує білок нуклеокапсида – HBcAg, що складається з 183-185
Ген С, кодує білок нуклеокапсида – HBcAg, що складається з 183-185
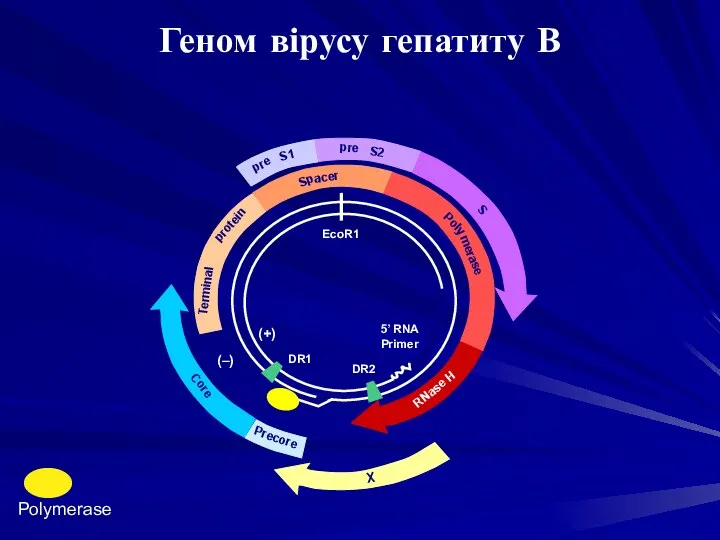
Precore
Core
X
RNase H
5’ RNA
Primer
DR2
DR1
(+)
(–)
protein
acer
Poly
EcoR1
S1
S2
S
Terminal
pre
pre
merase
Sp
Polymerase
Геном вірусу гепатиту В
Precore
Core
X
RNase H
5’ RNA
Primer
DR2
DR1
(+)
(–)
protein
acer
Poly
EcoR1
S1
S2
S
Terminal
pre
pre
merase
Sp
Polymerase
Геном вірусу гепатиту В

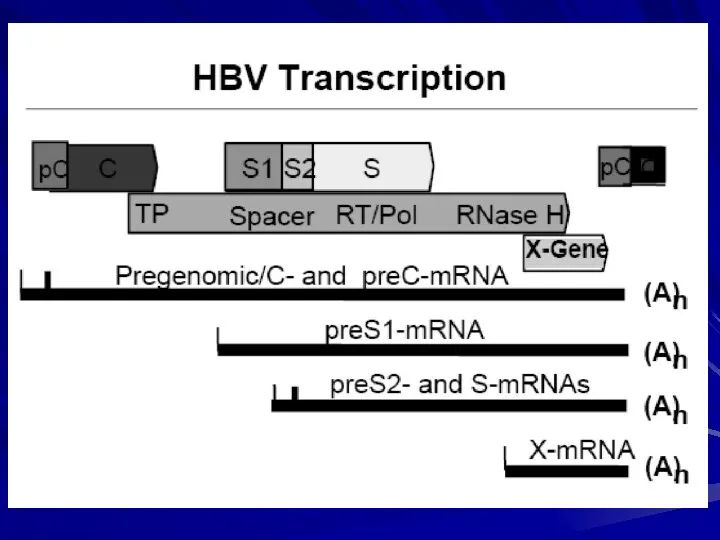
Транскрипційно/Трансляційнаорганізація
4 major RNA species ---------- ~ 7 Proteins
3.5 kb
Pre-C/C
Pol
Pregenomic
2.4 kb
Pre-S1
2.1 kb
Pre-S2/S
0.7
Транскрипційно/Трансляційнаорганізація
4 major RNA species ---------- ~ 7 Proteins
3.5 kb
Pre-C/C
Pol
Pregenomic
2.4 kb
Pre-S1
2.1 kb
Pre-S2/S
0.7

Viral Gene Products
S gene: Surface, Envelope, HBsAg
Envelope
Small, medium and large
Glycosylation
C gene:
Viral Gene Products
S gene: Surface, Envelope, HBsAg
Envelope
Small, medium and large
Glycosylation
C gene:

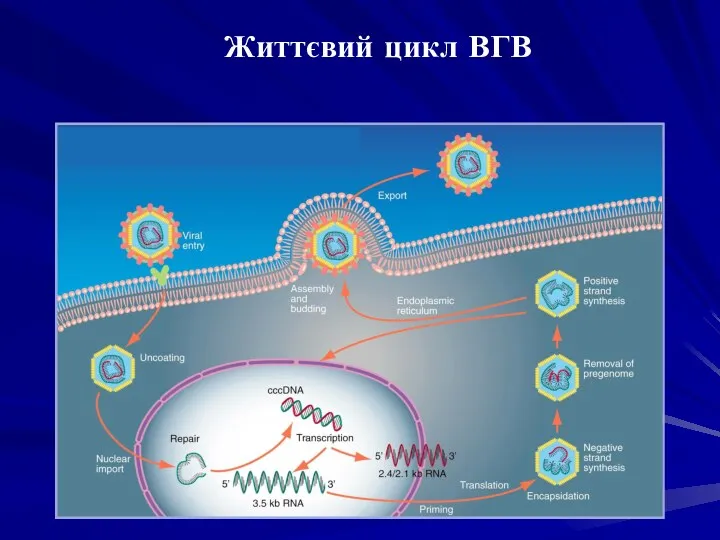
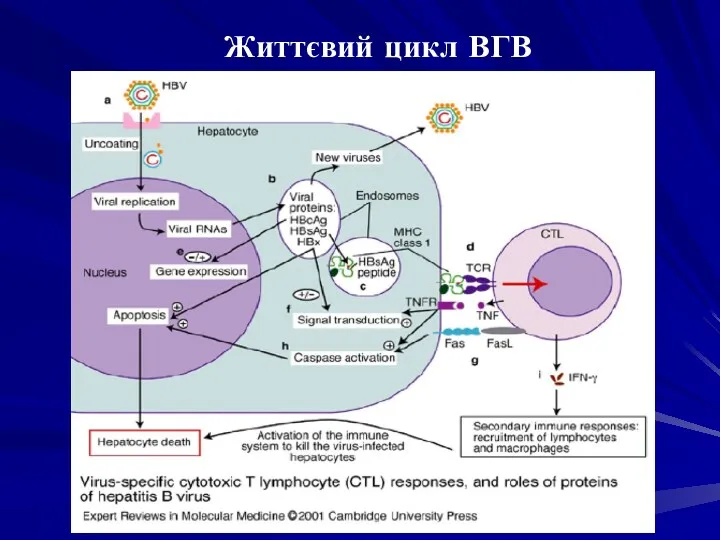
Реплікація генома вірусу гепатиту В починається з проникнення віріона в гепатоцит
Реплікація генома вірусу гепатиту В починається з проникнення віріона в гепатоцит
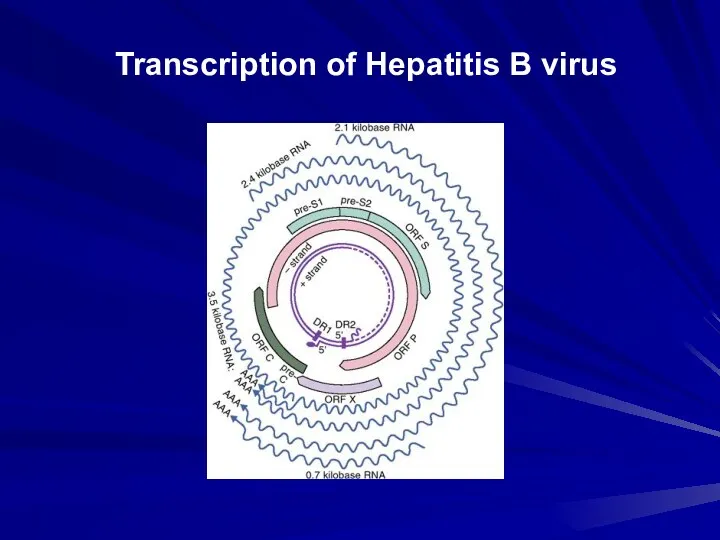
Transcription of Hepatitis B virus
Transcription of Hepatitis B virus

Регуляція синтезу білків ВГВ
Синтез білків вірусу гепатиту В регулюється на рівні
Регуляція синтезу білків ВГВ
Синтез білків вірусу гепатиту В регулюється на рівні

Механізм зворотньої транскрипції ВГВ
Прегеномна РНК (штрихова лінія, крок 1)
Механізм зворотньої транскрипції ВГВ
Прегеномна РНК (штрихова лінія, крок 1)
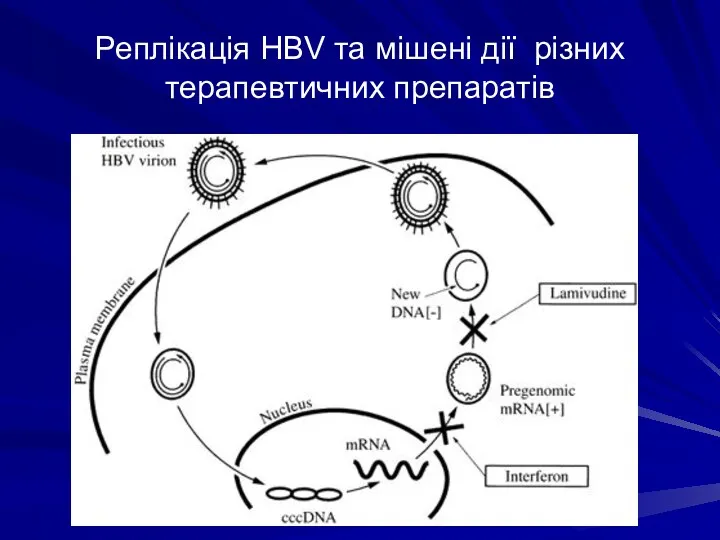
Реплікація HBV та мішені дії різних терапевтичних препаратів
Реплікація HBV та мішені дії різних терапевтичних препаратів
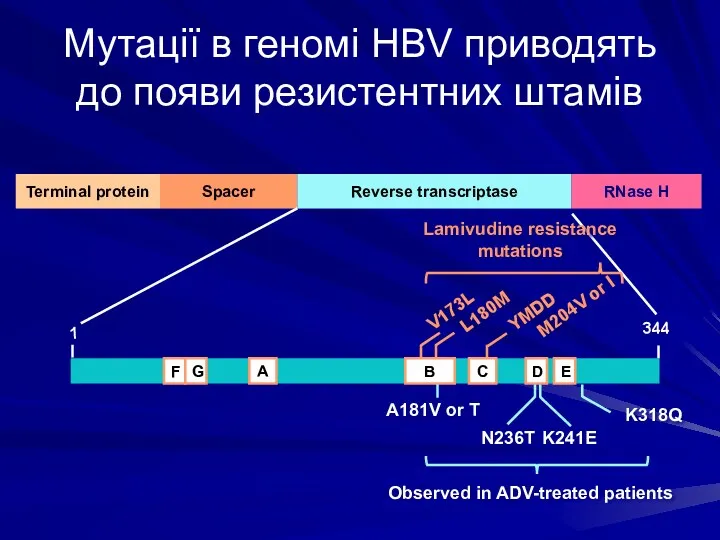
Мутації в геномі HBV приводять до появи резистентних штамів
Observed in
Мутації в геномі HBV приводять до появи резистентних штамів
Observed in

Caulimoviridae
Caulimoviridae
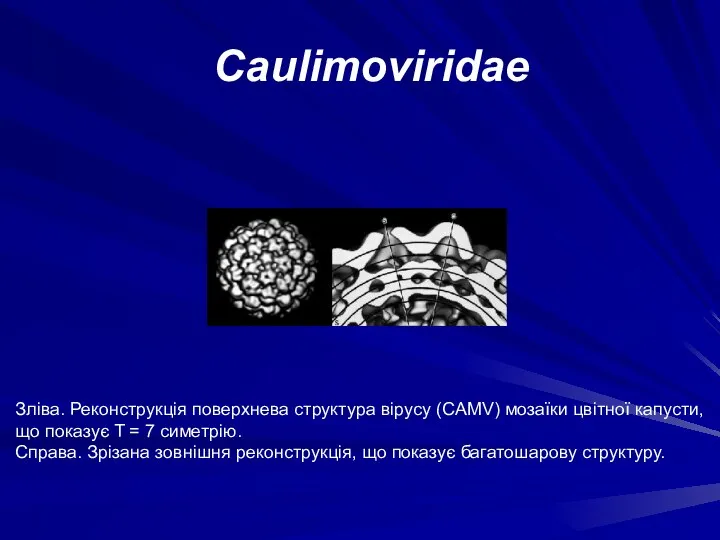
Зліва. Реконструкція поверхнева структура вірусу (CAMV) мозаїки цвітної капусти, що
Зліва. Реконструкція поверхнева структура вірусу (CAMV) мозаїки цвітної капусти, що
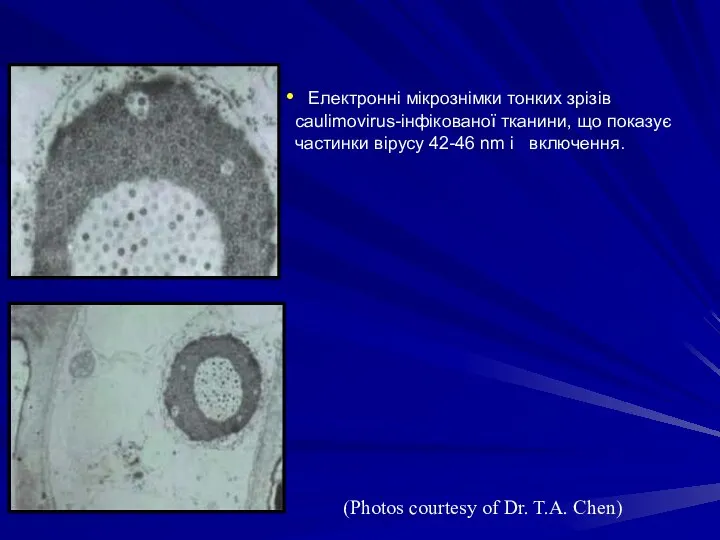
Електронні мікрознімки тонких зрізів caulimovirus-інфікованої тканини, що показує частинки вірусу
Електронні мікрознімки тонких зрізів caulimovirus-інфікованої тканини, що показує частинки вірусу

Caulimoviridae
Ізометричні 50 nm частинки з T=7 або бацилярні частинки;
Дл ДНК геном
Caulimoviridae
Ізометричні 50 nm частинки з T=7 або бацилярні частинки;
Дл ДНК геном

Caulimoviridae
Переважно вузьке коло господарів
Переважно малозначні як патогени; виключення - Rice tungro
Caulimoviridae
Переважно вузьке коло господарів
Переважно малозначні як патогени; виключення - Rice tungro
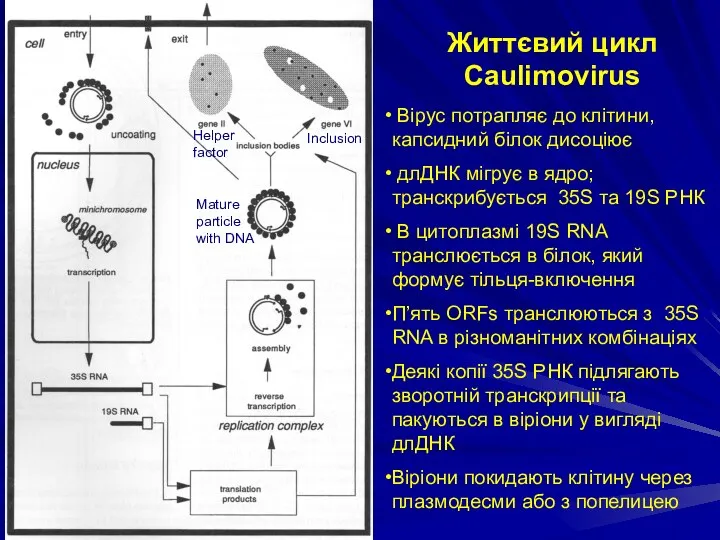
Життєвий цикл Caulimovirus
Вірус потрапляє до клітини, капсидний білок
Життєвий цикл Caulimovirus
Вірус потрапляє до клітини, капсидний білок
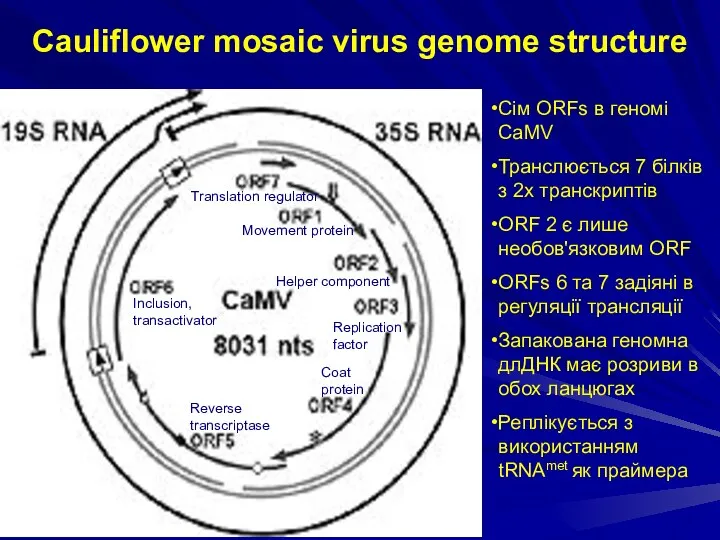
Cauliflower mosaic virus genome structure
Сім ORFs в геномі CaMV
Транслюється
Cauliflower mosaic virus genome structure
Сім ORFs в геномі CaMV
Транслюється

Cauliflower mosaic virus (CaMV)
Кожен ланцюг нуклеїнової кислоти має розриви у специфічних
Cauliflower mosaic virus (CaMV)
Кожен ланцюг нуклеїнової кислоти має розриви у специфічних
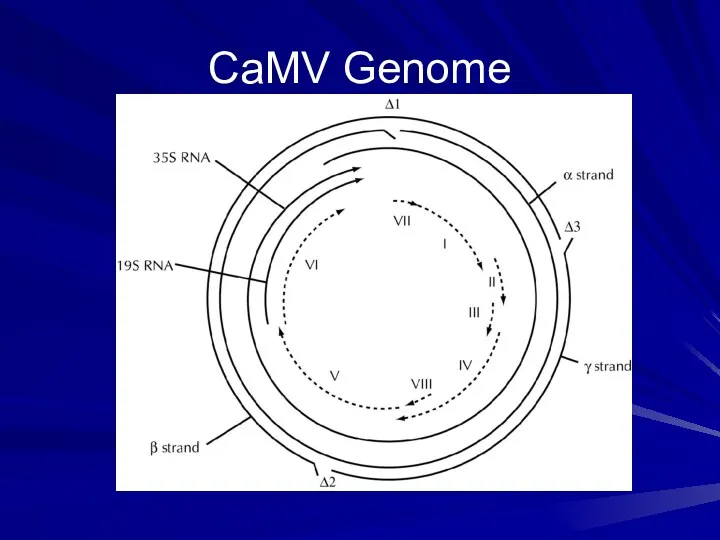
CaMV Genome
CaMV Genome

Genome Organization of Caulimoviridae
CERV
CsVMV
PVCV
Petuvirus
Caulimoviruses
CaMV
FMV
Soymoviruses
BRRV
SbCMV
Cavemoviruses
Tungroviruses
RTBV
Badnaviruses
ComYMV
I
I
I
I
II
II
II
a
II
I
III
III
III
I
IV
IV
IV
IV
III
V
V
VI
VI
b
c
II
VII
VII
V
Genome Organization of Caulimoviridae
CERV
CsVMV
PVCV
Petuvirus
Caulimoviruses
CaMV
FMV
Soymoviruses
BRRV
SbCMV
Cavemoviruses
Tungroviruses
RTBV
Badnaviruses
ComYMV
I
I
I
I
II
II
II
a
II
I
III
III
III
I
IV
IV
IV
IV
III
V
V
VI
VI
b
c
II
VII
VII
V

Після потрапляння у клітину розриви в геномі «замуровуються» з подальшим формуванням
Після потрапляння у клітину розриви в геномі «замуровуються» з подальшим формуванням
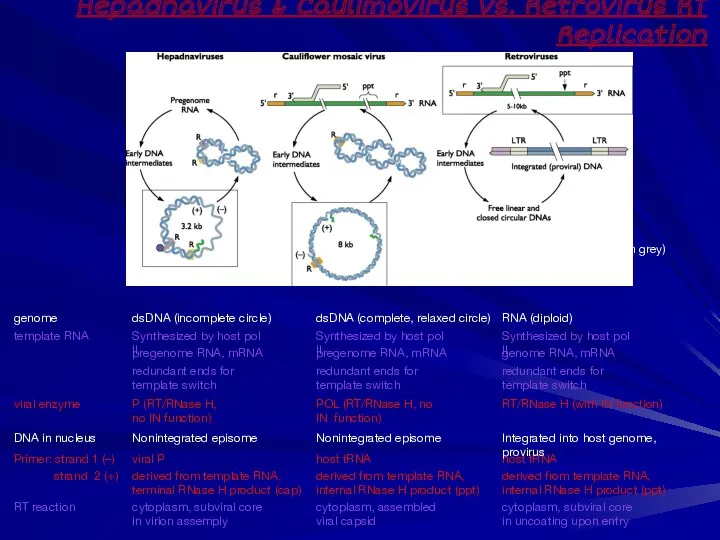
Hepadnavirus & Caulimovirus vs. Retrovirus RT Replication
Hepadnavirus & Caulimovirus vs. Retrovirus RT Replication
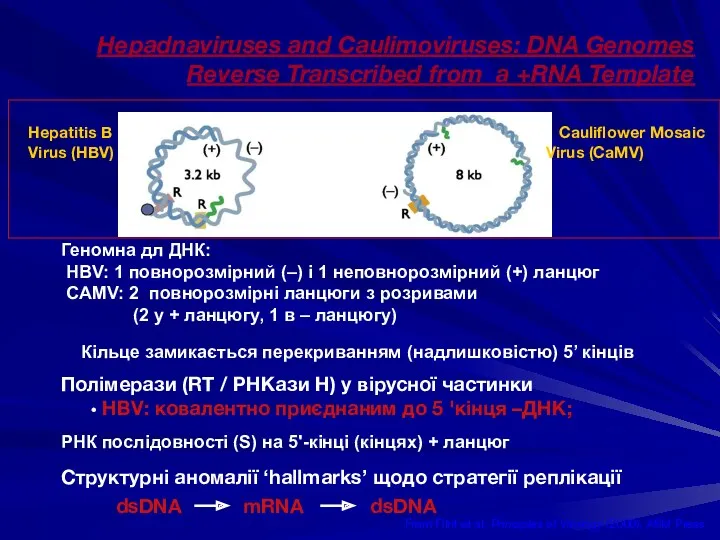
Hepadnaviruses and Caulimoviruses: DNA Genomes Reverse Transcribed from a +RNA Template
From
Hepadnaviruses and Caulimoviruses: DNA Genomes Reverse Transcribed from a +RNA Template
From
 Рост и развитие растений. (6 класс)
Рост и развитие растений. (6 класс) Мышечные цепи. Взаимосвязи миофасциальной сети
Мышечные цепи. Взаимосвязи миофасциальной сети Систематика низших растений
Систематика низших растений Органические вещества – углеводы в структуре питания
Органические вещества – углеводы в структуре питания Жануарлар биотехнологиясының жалпы биологиялық негіздері
Жануарлар биотехнологиясының жалпы биологиялық негіздері Красители. Классификация. Приготовление красителей. Артефакты. Методы микроскопии
Красители. Классификация. Приготовление красителей. Артефакты. Методы микроскопии Опорно-двигательный аппарат
Опорно-двигательный аппарат Грибы. Характеристика
Грибы. Характеристика Соединения костей туловища
Соединения костей туловища Основные перестройки, происходящие в начале культивирования растительных клеток в условиях in vitro
Основные перестройки, происходящие в начале культивирования растительных клеток в условиях in vitro Птицы
Птицы Як змінюється восени життя птахів. Перелітні та осілі птахи
Як змінюється восени життя птахів. Перелітні та осілі птахи Строение и функционирование генома
Строение и функционирование генома Строение головного мозга хордовых
Строение головного мозга хордовых Тебердинский государственный биосферный заповедник
Тебердинский государственный биосферный заповедник Филогенетическое разнообразие грибоподобных организмов. Разнообразие. Систематика. Жизненные циклы
Филогенетическое разнообразие грибоподобных организмов. Разнообразие. Систематика. Жизненные циклы презентация Пушистый доктор
презентация Пушистый доктор Красная книга Челябинской области
Красная книга Челябинской области Виды корней. Типы корневых систем растений
Виды корней. Типы корневых систем растений Цитоплазма. Одномембранные органоиды
Цитоплазма. Одномембранные органоиды Витамины. Классификация витаминов
Витамины. Классификация витаминов Ещё немного о белках
Ещё немного о белках Совообра́зные — отряд хищных птиц
Совообра́зные — отряд хищных птиц Листья и плоды деревьев и кустарников
Листья и плоды деревьев и кустарников Центры подкрепления, прилежащее ядро. Кора больших полушарий: механизмы обучения; гиппокамп
Центры подкрепления, прилежащее ядро. Кора больших полушарий: механизмы обучения; гиппокамп Кровеносная система
Кровеносная система Популяция. Функция популяции как системы
Популяция. Функция популяции как системы Ядрышко. Ядерная мембрана. Ядерно-цитоплазматический транспорт
Ядрышко. Ядерная мембрана. Ядерно-цитоплазматический транспорт