- Посттрансляционные модификации белков

Содержание
- 4. Белок-синтезирующие системы эукариотной клетки
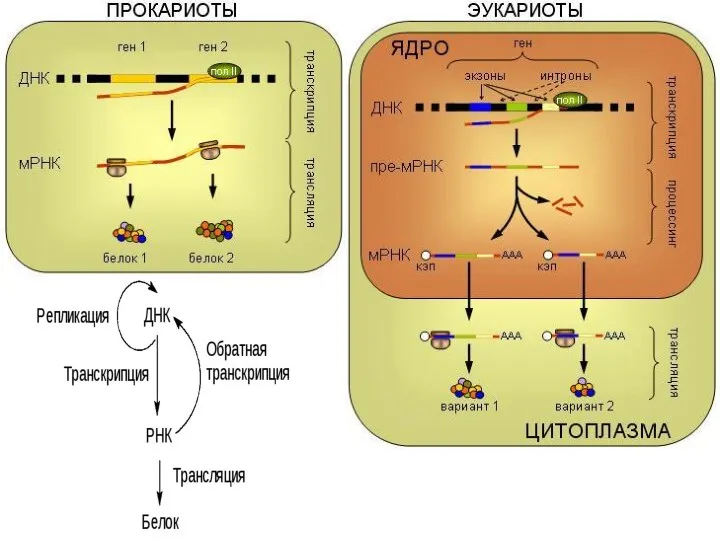
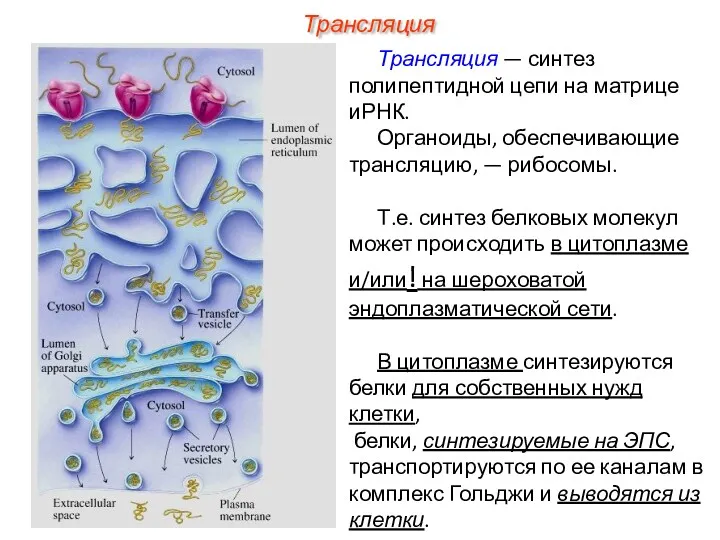
- 5. Трансляция Трансляция — синтез полипептидной цепи на матрице иРНК. Органоиды, обеспечивающие трансляцию, — рибосомы. Т.е. синтез
- 6. Для увеличения производства белка через иРНК могут одновременно проходить несколько рибосом, последовательно транслирующие один и тот
- 7. Роль посттрансляционных модификаций в многообразии белков
- 8. Посттрансляционные модификации участвуют практически во всех клеточных событиях, в том числе: Экспрессии генов Сигнальной трансдукции Белок-белковых
- 9. Посттрансляционные модификации Ковалентные (химическое присоединение дополнительных групп) НЕковалентные (фолдинг, процессинг)
- 10. Виды посттрансляционных ковалентных (химических) модификаций белков Фосфорилирование Ацетилирование Амидирование Сумоилирование Формилирование Гликозилирование Убиквитинирование Пальмитоилирование Миристоилирование Нитрование
- 13. ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот ПРИМЕР: коллаген синтезируется в виде проколлагена. Гидроксилирование остатков
- 14. ГЛИКОЗИЛИРОВАНИЕ - присоединение углеводных остатков, образование гликопротеинов Происходит в ЭПС и комплексе Гольджи Перенос олигосахаридов Укорачивание
- 15. АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ Присоединение ацильной или метильной группы ПРИМЕР: ацетилирование или метилирование гистонов, что влияет на
- 16. ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ ВЕЛИЧИНЫ ПОЛОЖИТЕЛЬНОГО ЗАРЯДА ГИСТОНОВ,
- 17. ФОСФОРИЛИРОВАНИЕ Присоединение фосфатной группы. Обратимый процесс. Фосфорилируется остаток серина (треонина) или тирозина. Несмотря на большое количество
- 18. ФОСФОРИЛИРОВАНИЕ КИНАЗА ФОСФОРИЛИРУЕТ ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ
- 20. Процессинг полипептидной цепи Регуляция трансляции. Процессинг Чаще всего в результате трансляции полипептидные цепи образуются в неактивной
- 21. Посттрансляционный процессинг
- 22. Посттрансляционный процессинг полипептидной цепи
- 23. Вопрос - каким образом белки так быстро (буквально за наносекунды) принимают необходимую третичную структуру. Так, достаточно
- 24. Фолдингом белка (укладкой белка, от англ. folding) называют процесс спонтанного сворачивания полипептидной цепи в уникальную нативную
- 25. •В фолдинге участвуют белки-шапероны. •Большинство только что синтезированных белков может сворачиваться при отсутствии шаперонов •Шапероны —
- 26. Фолдинг Участие шаперонов в фолдинге белка
- 27. Посттрансляционный процессинг Регуляция трансляции. Процессинг Роль шаперонов в фолдинге белков Синтез и фолдинг белков протекают при
- 28. •Многие шапероны являются белками теплового шока, то есть белками, экспрессия которых начинается в ответ на рост
- 29. •Фолдинг белков происходит в эндоплазматическом ретикулуме •В нём содержатся необходимые для фолдинга шапероны и ферменты •Кроме
- 31. Скачать презентацию
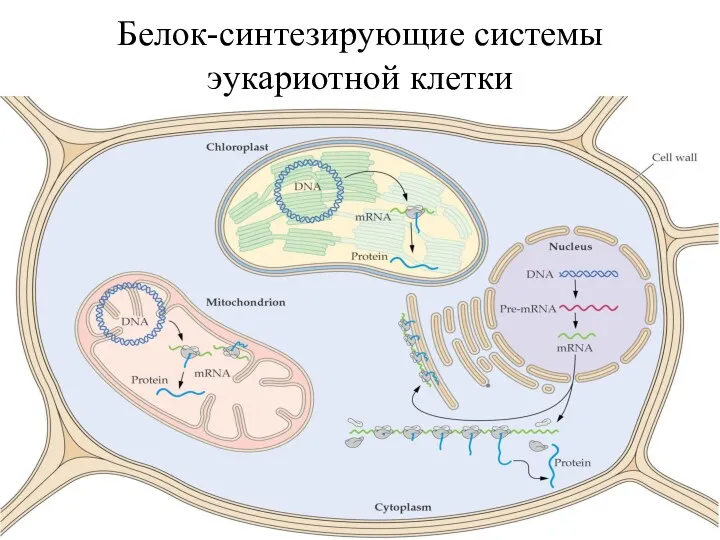
Белок-синтезирующие системы
эукариотной клетки
Белок-синтезирующие системы
эукариотной клетки
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию,
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию,
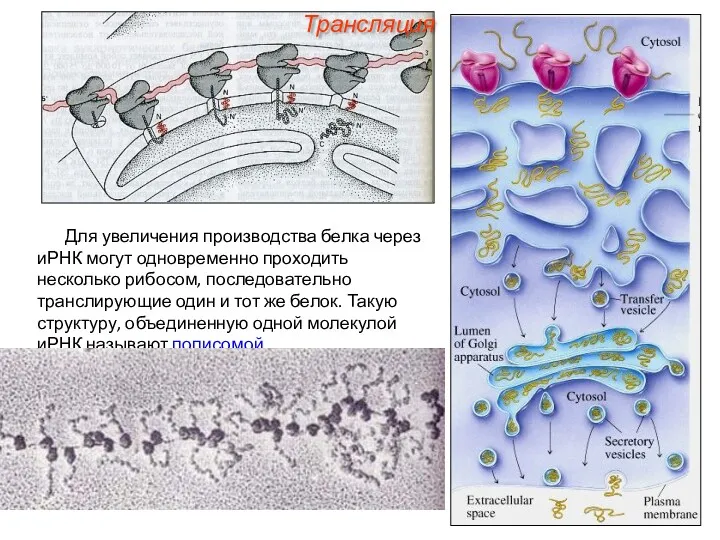
Для увеличения производства белка через иРНК могут одновременно проходить несколько рибосом,
Для увеличения производства белка через иРНК могут одновременно проходить несколько рибосом,
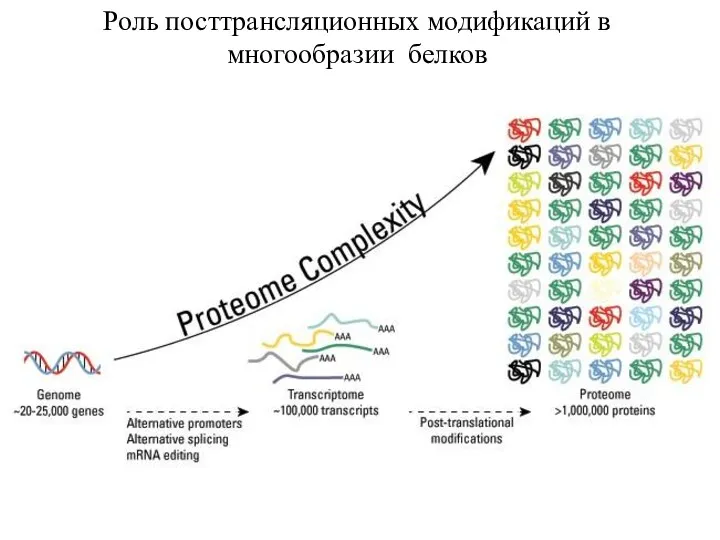
Роль посттрансляционных модификаций в многообразии белков
Роль посттрансляционных модификаций в многообразии белков

Посттрансляционные модификации участвуют практически во всех клеточных событиях, в том числе:
Экспрессии
Посттрансляционные модификации участвуют практически во всех клеточных событиях, в том числе:
Экспрессии
Посттрансляционные модификации
Ковалентные
(химическое присоединение дополнительных групп)
НЕковалентные
(фолдинг, процессинг)
Посттрансляционные модификации
Ковалентные
(химическое присоединение дополнительных групп)
НЕковалентные
(фолдинг, процессинг)
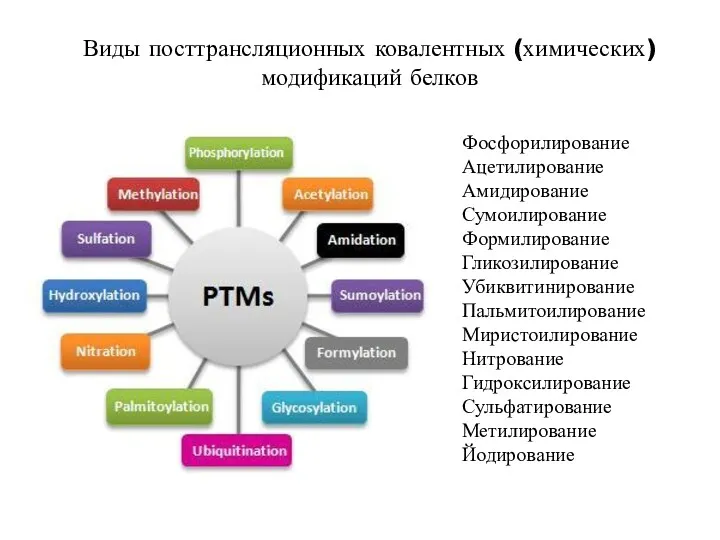
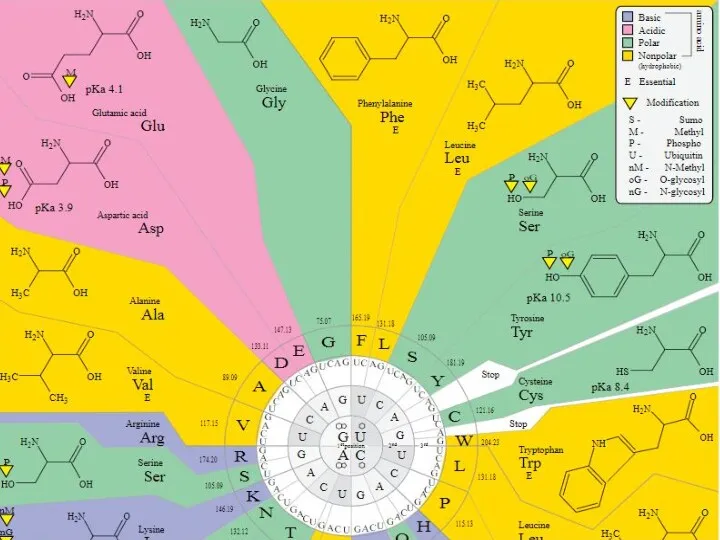
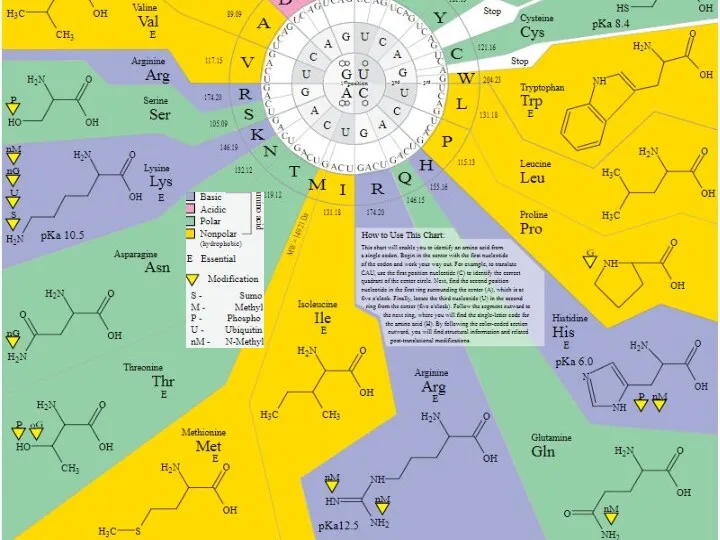
Виды посттрансляционных ковалентных (химических) модификаций белков
Фосфорилирование
Ацетилирование
Амидирование
Сумоилирование
Формилирование
Гликозилирование
Убиквитинирование
Пальмитоилирование
Миристоилирование
Нитрование
Гидроксилирование
Сульфатирование
Метилирование
Йодирование
Виды посттрансляционных ковалентных (химических) модификаций белков
Фосфорилирование
Ацетилирование
Амидирование
Сумоилирование
Формилирование
Гликозилирование
Убиквитинирование
Пальмитоилирование
Миристоилирование
Нитрование
Гидроксилирование
Сульфатирование
Метилирование
Йодирование

ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот
ПРИМЕР: коллаген синтезируется
ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот
ПРИМЕР: коллаген синтезируется
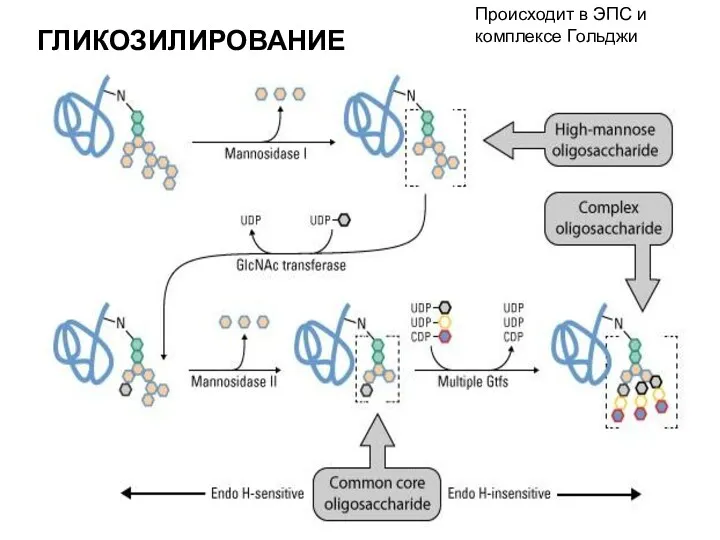
ГЛИКОЗИЛИРОВАНИЕ
- присоединение углеводных остатков, образование гликопротеинов
Происходит в ЭПС и комплексе
ГЛИКОЗИЛИРОВАНИЕ
- присоединение углеводных остатков, образование гликопротеинов
Происходит в ЭПС и комплексе
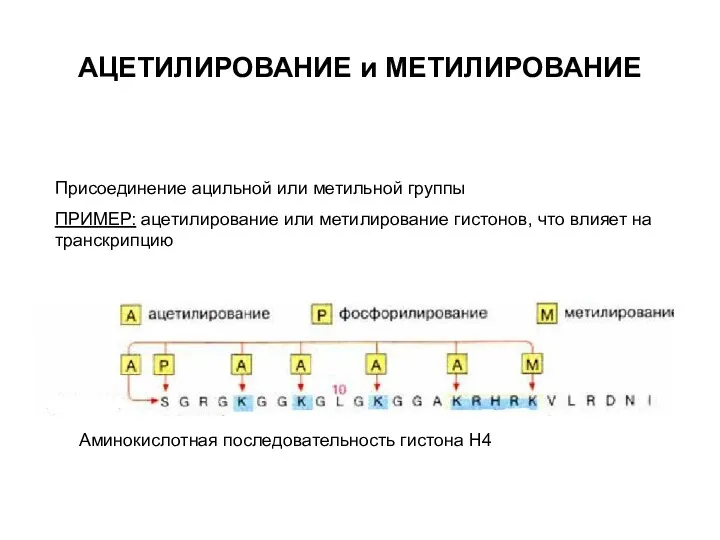
АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ
Присоединение ацильной или метильной группы
ПРИМЕР: ацетилирование или метилирование гистонов,
АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ
Присоединение ацильной или метильной группы
ПРИМЕР: ацетилирование или метилирование гистонов,
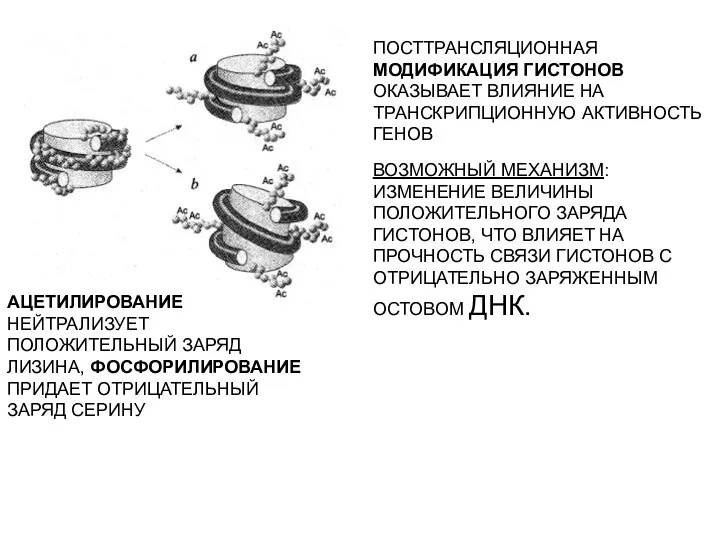
ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ
ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ
ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ
ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ

ФОСФОРИЛИРОВАНИЕ
Присоединение фосфатной группы.
Обратимый процесс.
Фосфорилируется остаток серина (треонина) или тирозина.
Несмотря на
ФОСФОРИЛИРОВАНИЕ
Присоединение фосфатной группы.
Обратимый процесс.
Фосфорилируется остаток серина (треонина) или тирозина.
Несмотря на
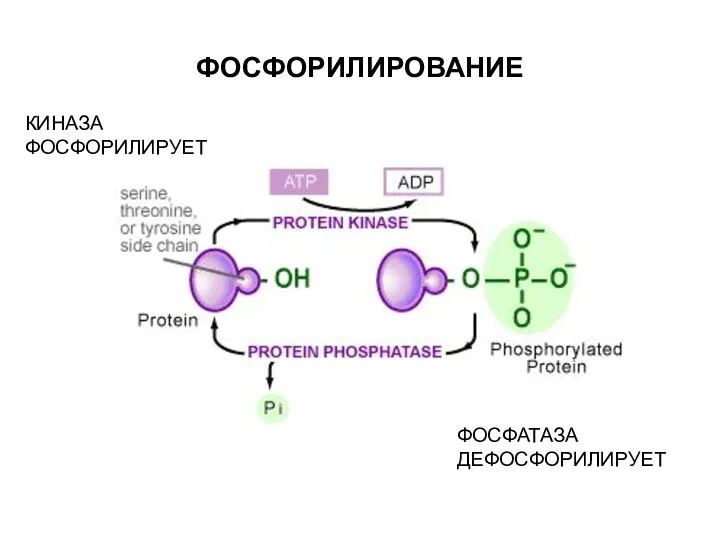
ФОСФОРИЛИРОВАНИЕ
КИНАЗА ФОСФОРИЛИРУЕТ
ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ
ФОСФОРИЛИРОВАНИЕ
КИНАЗА ФОСФОРИЛИРУЕТ
ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ

Процессинг полипептидной цепи
Регуляция трансляции. Процессинг
Чаще всего в результате трансляции полипептидные
Процессинг полипептидной цепи
Регуляция трансляции. Процессинг
Чаще всего в результате трансляции полипептидные
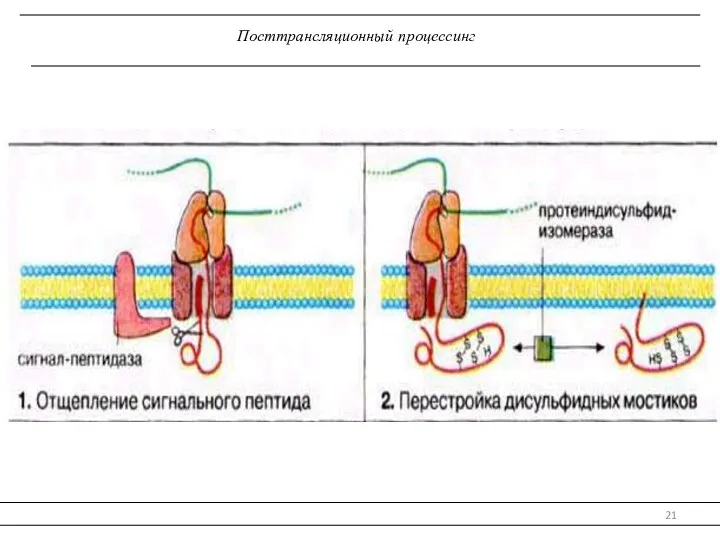
Посттрансляционный процессинг
Посттрансляционный процессинг
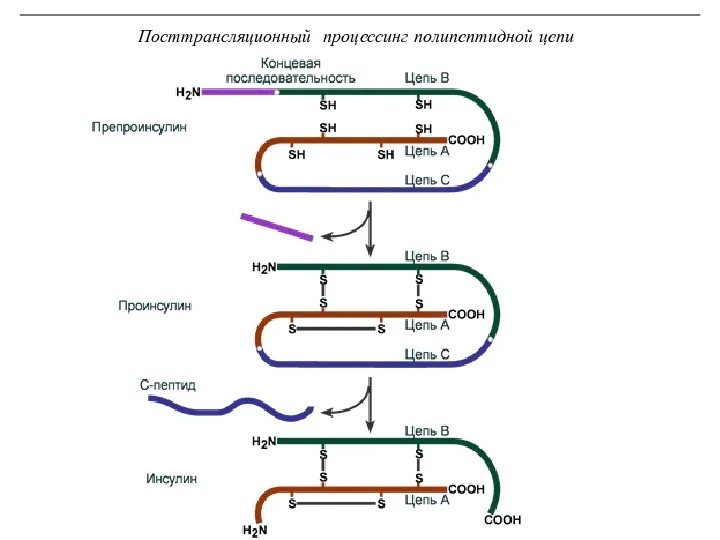
Посттрансляционный процессинг полипептидной цепи
Посттрансляционный процессинг полипептидной цепи

Вопрос - каким образом белки так быстро (буквально за наносекунды) принимают
Вопрос - каким образом белки так быстро (буквально за наносекунды) принимают
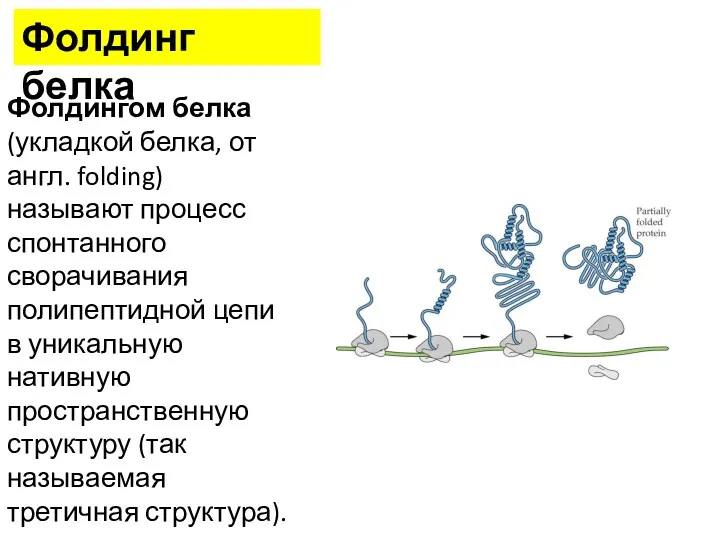
Фолдингом белка (укладкой белка, от англ. folding) называют процесс спонтанного сворачивания

•В фолдинге участвуют белки-шапероны.
•Большинство только что синтезированных белков может сворачиваться
•Большинство только что синтезированных белков может сворачиваться
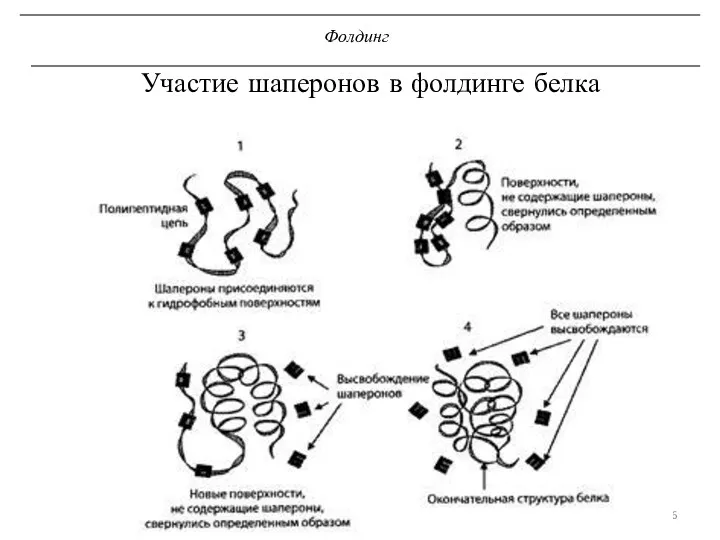
Фолдинг
Участие шаперонов в фолдинге белка
Фолдинг
Участие шаперонов в фолдинге белка
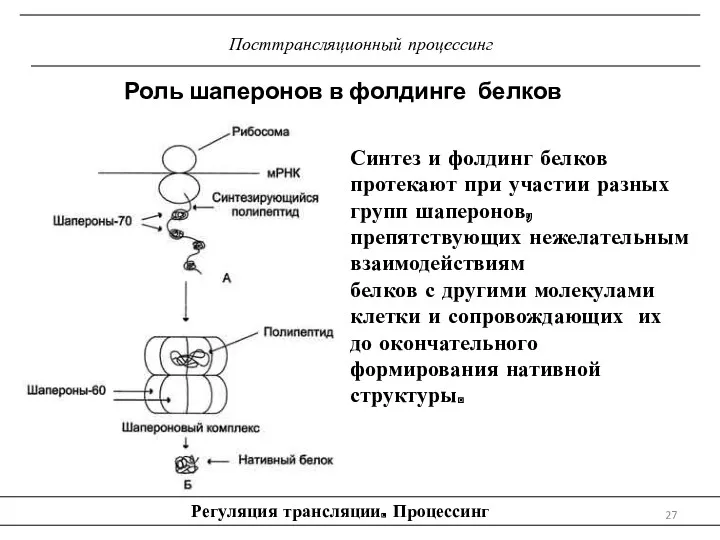
Посттрансляционный процессинг
Регуляция трансляции. Процессинг
Роль шаперонов в фолдинге белков
Синтез и фолдинг белков
Посттрансляционный процессинг
Регуляция трансляции. Процессинг
Роль шаперонов в фолдинге белков
Синтез и фолдинг белков

•Многие шапероны являются белками теплового шока, то есть белками, экспрессия которых
•Многие шапероны являются белками теплового шока, то есть белками, экспрессия которых

•Фолдинг белков происходит в эндоплазматическом ретикулуме
•В нём содержатся необходимые для
•Фолдинг белков происходит в эндоплазматическом ретикулуме
•В нём содержатся необходимые для
 Фитомелиализацияның әсерін зерттеу ауылдың жайылым топырақтарының физикалық, физикалық және химиялық қасиеттері
Фитомелиализацияның әсерін зерттеу ауылдың жайылым топырақтарының физикалық, физикалық және химиялық қасиеттері Основы учения о биосфере. Ноосфера. Техногенез
Основы учения о биосфере. Ноосфера. Техногенез Строение и функции клетки
Строение и функции клетки Обмен веществ и превращение энергии
Обмен веществ и превращение энергии СОСТАВ КРОВИ.
СОСТАВ КРОВИ. Спинной мозг
Спинной мозг Влияние стимуляторов роста на проращивание семян томатов
Влияние стимуляторов роста на проращивание семян томатов Лесные доктора
Лесные доктора Готовимся к ЕГЭ по биологии
Готовимся к ЕГЭ по биологии Өсімдіктерге төмен температураның әсері
Өсімдіктерге төмен температураның әсері Використання мутагегезу в селекції польових культур
Використання мутагегезу в селекції польових культур Заповедник Галичья гора в Липецкой области
Заповедник Галичья гора в Липецкой области Многообразие и значение земноводных
Многообразие и значение земноводных Тип Кишечнополостные
Тип Кишечнополостные В царстве цветов
В царстве цветов Исследования психологических признаков
Исследования психологических признаков Чарлз Дарвин. Путешествие на Бигле
Чарлз Дарвин. Путешествие на Бигле Мышечная система. Часть 2
Мышечная система. Часть 2 Презентация по биологии. 7 класс. Животные. Строение и деятельность внутренних органов земноводных.
Презентация по биологии. 7 класс. Животные. Строение и деятельность внутренних органов земноводных. Онтогенез. Лекция 14
Онтогенез. Лекция 14 Tiere
Tiere Прокариотическая клетка
Прокариотическая клетка Методы лесовосстановления и лесоразведения
Методы лесовосстановления и лесоразведения Жасуша, ұлпа, мүше, мүшелер жүйесі түсініктері
Жасуша, ұлпа, мүше, мүшелер жүйесі түсініктері Динозавры
Динозавры Тип Членистоногие
Тип Членистоногие История изучения клетки. Клеточная теория
История изучения клетки. Клеточная теория Тип кишечнополостные (пресноводная гидра)
Тип кишечнополостные (пресноводная гидра)