- Потенциал действия

Содержание
- 2. Young JZ (1936) Structure of nerve fibres and synapses in some invertebrates. Cold Spring Harb Symp
- 3. Гигантский аксон кальмара (A) Diagram of a squid, showing the location of its giant nerve cells.
- 9. Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
- 10. Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
- 11. Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
- 12. Проблема измерения токов Разделение перечисленных токов, зависимых как от времени, так и самого потенциала, представляет собой
- 20. Этот метод оказался революционным для своего времени, поскольку предоставил ряд принципиальных возможностей для исследования возбудимости клеточных
- 21. Этот метод оказался революционным для своего времени, поскольку предоставил ряд принципиальных возможностей для исследования возбудимости клеточных
- 22. Схема усилителя с обратной связью Hodgkin AL, Huxley AF, Katz B (1952) Measurement of current-voltage relations
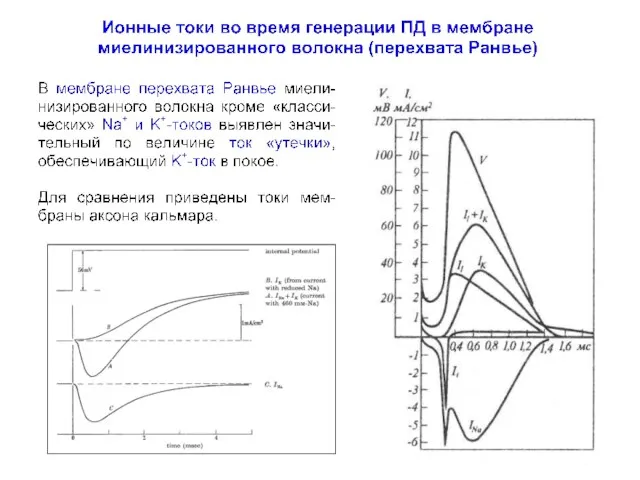
- 23. Ионные токи во время генерации ПД в аксоне кальмара Hodgkin AL, Huxley AF, Katz B (1952)
- 27. Временные динамики развития ПД (V) и определяющих его ионных проводимостей (gNa и gK), рассчитанные А. Ходжкиным
- 28. Временные динамики развития ПД (V) и определяющих его ионных проводимостей (gNa и gK), рассчитанные А. Ходжкиным
- 33. Активационные и инактивационные механизмы проводимости мембраны
- 34. Активация калиевой проводимости ПП 25 мВ HODGKIN, A. L. & HUXLEY, A. F. (1952) The components
- 35. Активация калиевой проводимости
- 36. Модель активации калиевой проводимости Зависимость величины GK от времени. Вставленный рисунок – то же в течение
- 37. Кривая активации калиевой проводимости Зависимость величины n∞ от потенциала деполяризация гиперполяризация ПП
- 38. Кривая активации калиевой проводимости Зависимость величины n∞ от потенциала гиперполяризация деполяризация ПП
- 39. Активация калииевой проводимости мембраны Для K+-проводимости А. Ходжкин и А. Хаксли ввели переменную n, которая отражает
- 40. Активация натриевой проводимости gNa = INa (V - VNa)
- 41. Инактивация натриевой проводимости gNa = INa (V - VNa)
- 42. Активация и инактивация натриевой проводимости
- 43. Кривая активации натриевой проводимости Зависимость величины m∞ от потенциала деполяризация гиперполяризация ПП
- 44. Кривая активации натриевой проводимости Зависимость величины m∞ от потенциала гиперполяризация деполяризация ПП
- 45. Кривая инактивации натриевой проводимости Зависимость величины h∞ от потенциала деполяризация гиперполяризация ПП
- 46. Кривая инактивации натриевой проводимости Зависимость величины h∞ от потенциала гиперполяризация деполяризация ПП
- 47. Активация Инактивация
- 48. Активация и инактивация натриевой проводимости мембраны Для Na+-проводимости А. Ходжкин и А. Хаксли ввели переменные m
- 49. Сначала они исследовали зависимость величин m и h от потенциала, зафиксированного на мембране в течение долгого
- 50. Затем они описали зависимость величин m и h от времени. В клетке в течение долгого времени
- 51. Инактивация натриевой проводимости мембраны (продолжение) А. Ходжкин и А. Хаксли предложили простую умозрительную физическую модель, которая
- 52. Инактивация натриевой проводимости мембраны (продолжение) Для отдельного канала переменная m отражает вероятность того, что одни m-ворота
- 53. Схема чередования различных возможных состояний m и h-ворот во время разных фаз генерации ПД (цикл А.
- 54. Фазы генерации ПД. Числами отмечены фазы ПД: 1 – ПП, 2 – восходящая, 3 нисходящая фазы
- 55. Схематическое отображение цикла А. Ходжкина. Состояние активационных (для Na+ и K+) и инактивационных (для Na+) ворот
- 56. Схема чередования различных возможных состояний m, h и n-ворот во время разных фаз генерации ПД (цикл
- 57. Зависимость состояния m- и h-ворот потенциал-зависимых Na+-каналов от мембранного потенциала (МП, мВ). В цикле А. Ходжкина
- 58. Воротные механизмы потенциал-зависимых Na+-каналов А. Ходжкин и А. Хаксли предположили, что активация Na+-каналов связана с открытием
- 59. + TTX Воротный и Na+ токи
- 64. Фазы рефрактерности потенциала действия Протеолитические ферменты (например, проназа или папаин), введенные внутриклеточно, ослабляют или даже устраняют
- 65. Фазы рефрактерности потенциала действия Воздействие деполяризации мембраны на процессы активации и инактивации Na+-проводимости имеет важное физиологическое
- 66. а – быстрое нарастание раздражающего стимула; б – медленное нарастание раздражающего стимула; v – потенциал; u
- 67. Механизм аккомодации Accommodation of the nerve membrane. Five superimposed traces show the membrane response to linearly
- 68. В мембране перехвата Ранвье !!! у млекопитающих (А) К+ токи не регистрируются у земноводных (В) К+
- 71. Генерация ПД в нейронах Аксоны представляют собой относительно простое мембранное образование, обеспечивающее проведение ПД по нервному
- 72. Разнообразие форм ПД a – схематичное представление ПД с его фазами развития. b – ПД гигантского
- 73. Метод внутриклеточного диализа Метод разработан в Институте физиологии АН УССР (г. Киев) в начале 1960-х г.г.
- 76. Метод внутриклеточного диализа a. Baker PF, Hodgkin AL, Shaw TI (1962) Replacement of the axoplasm of
- 77. Модификация метода внутриклеточного диализа Strickholm A (1962) Excitation currents and impedance of a small electrically isolated
- 79. Мультиионная природа токов в мембранах нейронов моллюсков Neher E, Lux HD (1969) Voltage clamp on Helix
- 80. Мультиионная природа токов в мембранах нейронов моллюсков Neher E, Lux HD (1969) Voltage clamp on Helix
- 81. Герасимов В.Д., Костюк П.Г., Майский В.А. Возбудимость гигантских нервных клеток различных представителей легочных моллюсков в растворах,
- 85. Скачать презентацию
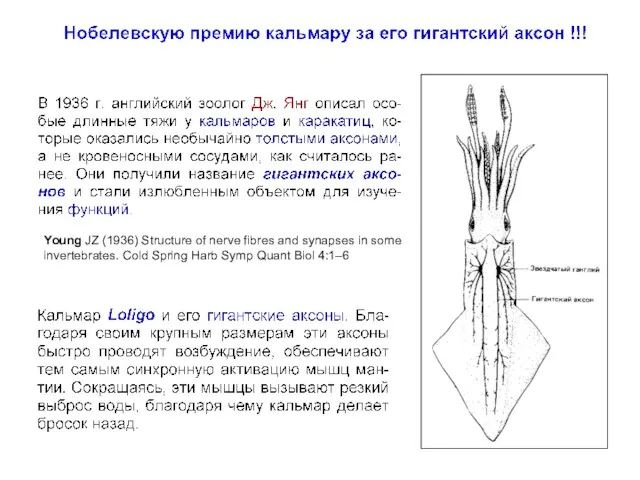
Young JZ (1936) Structure of nerve fibres and synapses in some
Young JZ (1936) Structure of nerve fibres and synapses in some
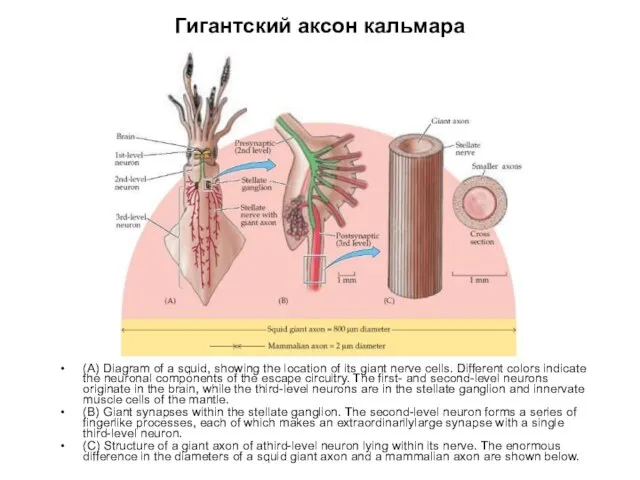
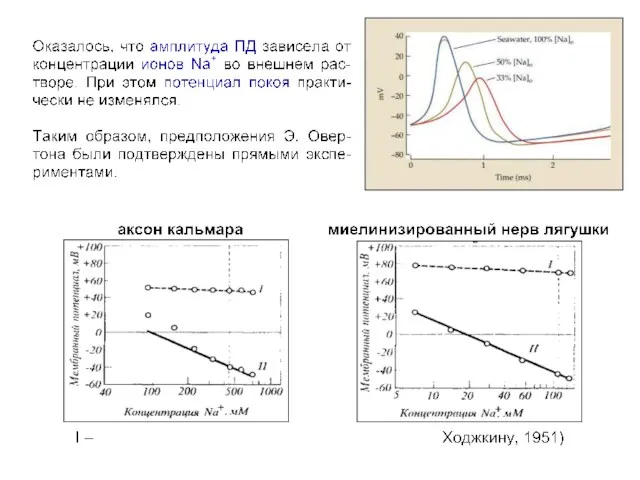
Гигантский аксон кальмара
(A) Diagram of a squid, showing the location of
Гигантский аксон кальмара
(A) Diagram of a squid, showing the location of
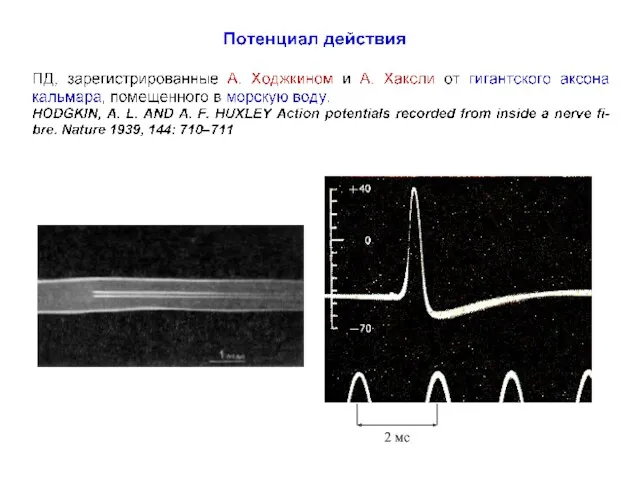
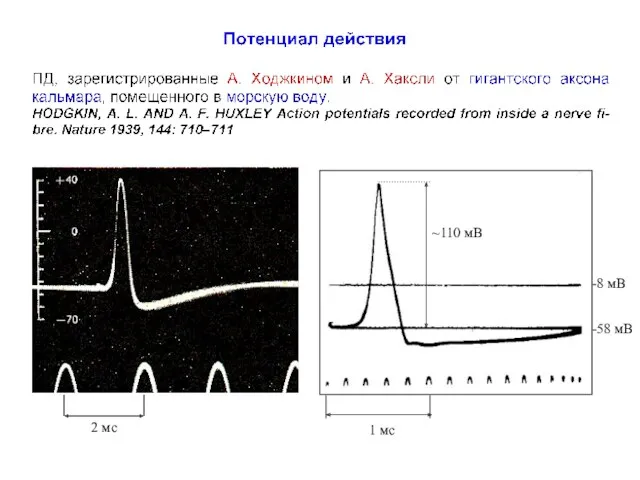


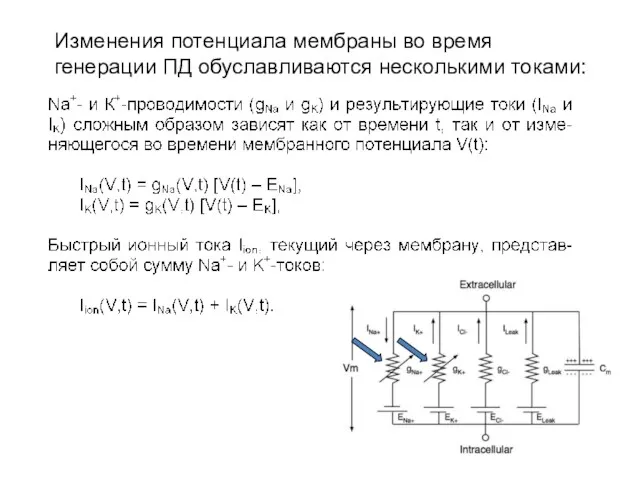
Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:

Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
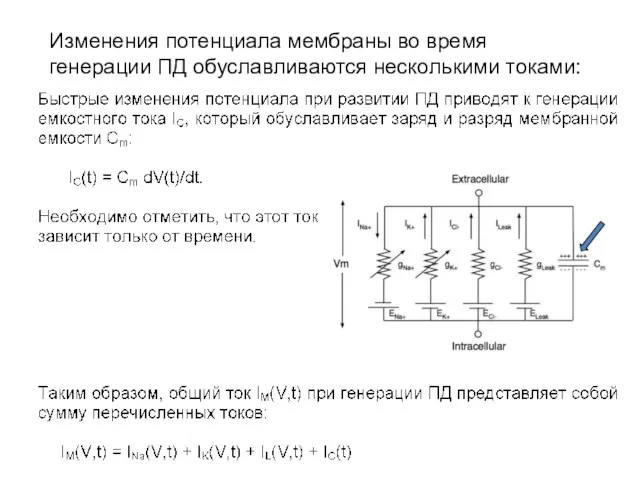
Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:
Изменения потенциала мембраны во время генерации ПД обуславливаются несколькими токами:

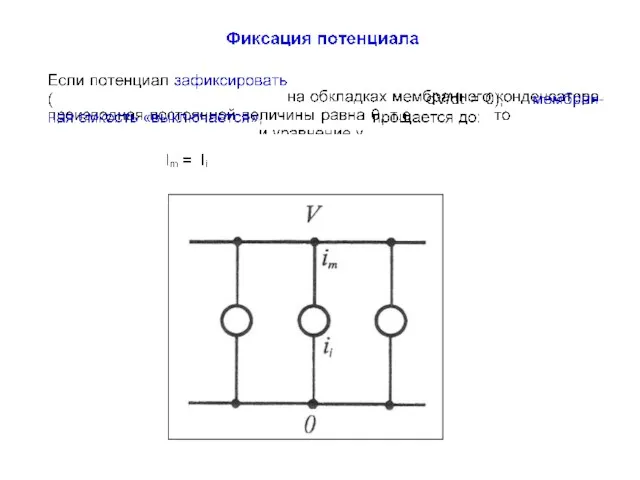
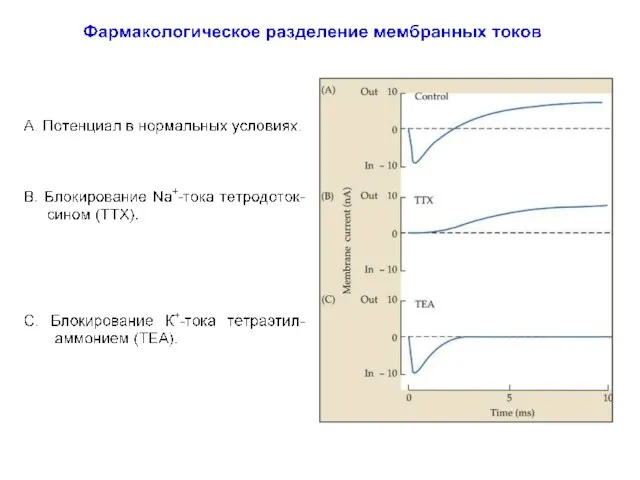
Проблема измерения токов
Разделение перечисленных токов, зависимых как от времени, так и
Проблема измерения токов
Разделение перечисленных токов, зависимых как от времени, так и
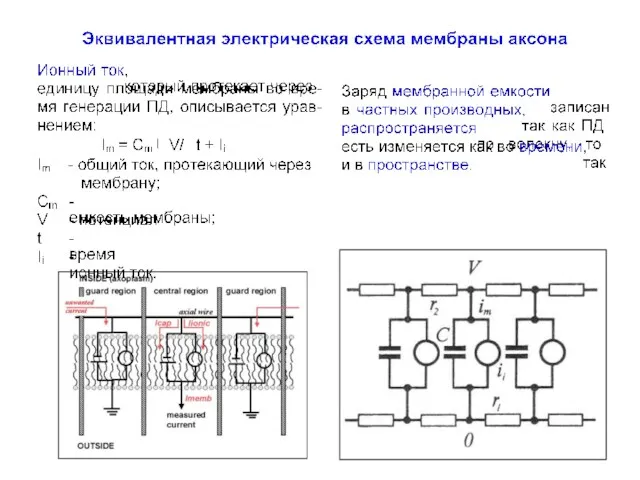
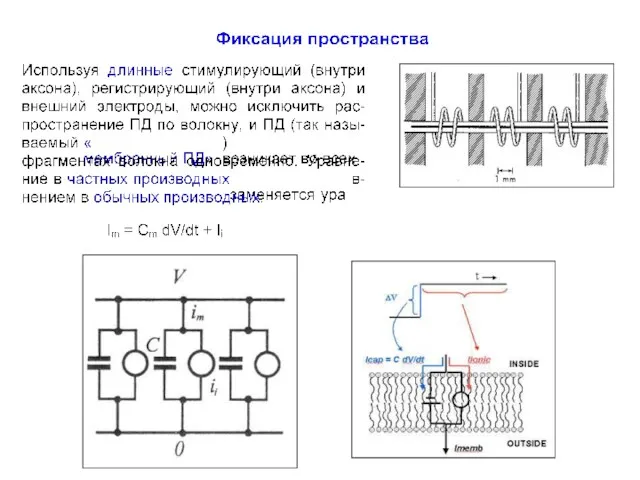
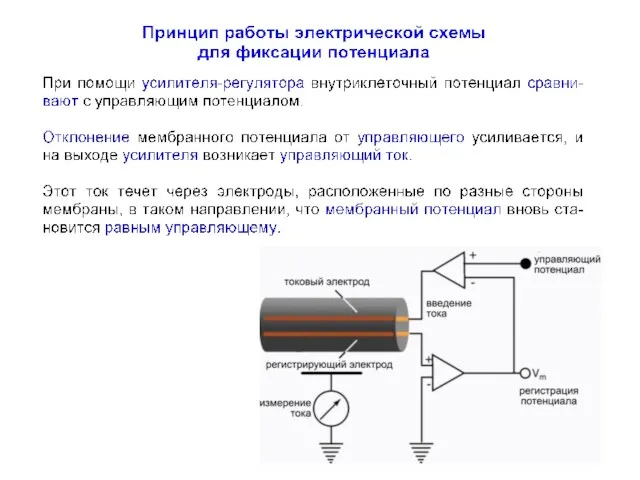
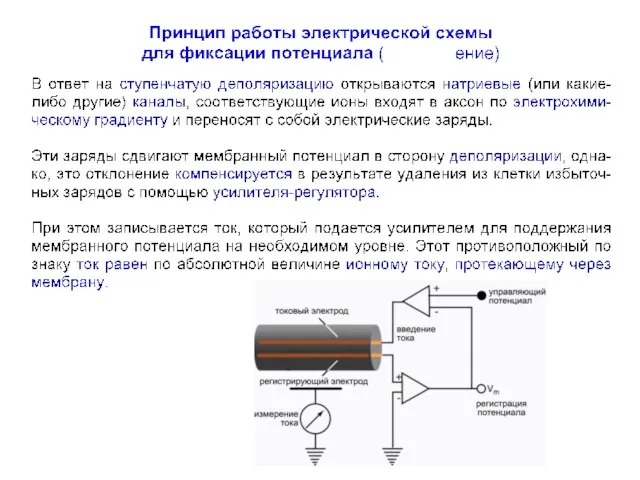

Этот метод оказался революционным для своего времени, поскольку предоставил ряд принципиальных
Этот метод оказался революционным для своего времени, поскольку предоставил ряд принципиальных

Этот метод оказался революционным для своего времени, поскольку предоставил ряд принципиальных
Этот метод оказался революционным для своего времени, поскольку предоставил ряд принципиальных
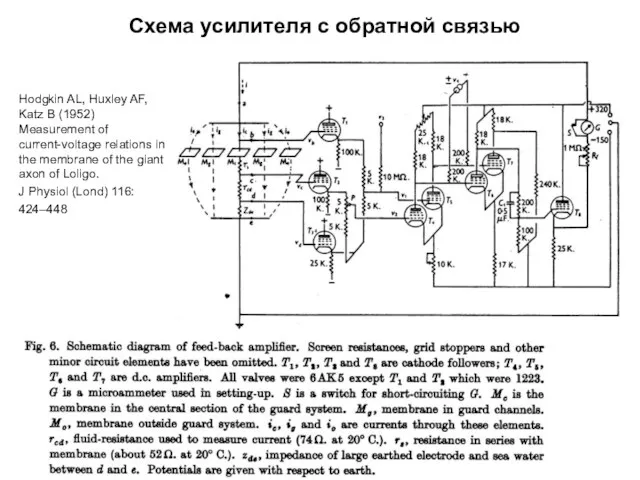
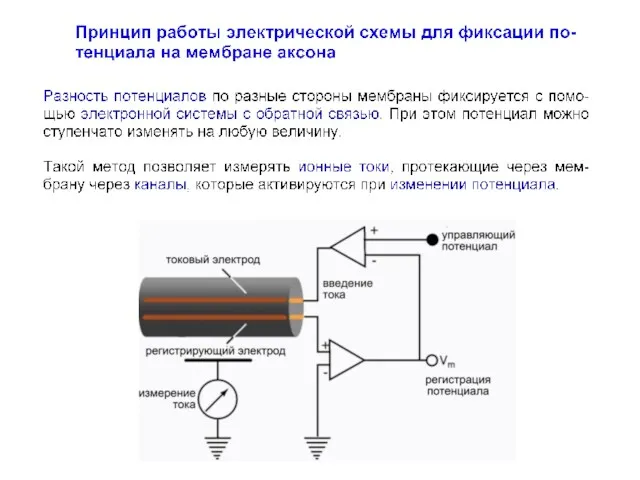
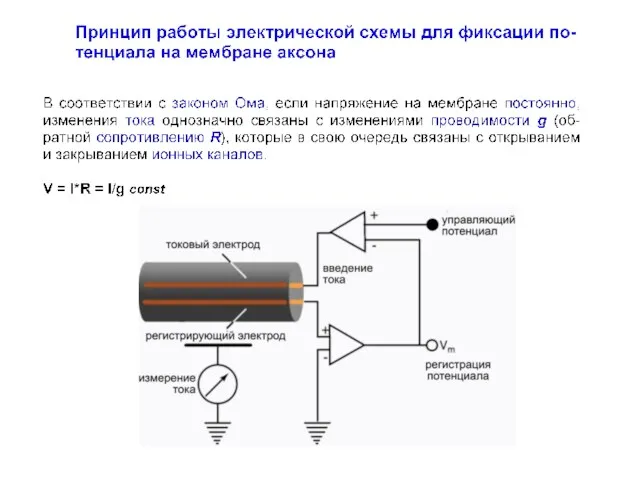
Схема усилителя с обратной связью
Hodgkin AL, Huxley AF, Katz B (1952)
Схема усилителя с обратной связью
Hodgkin AL, Huxley AF, Katz B (1952)
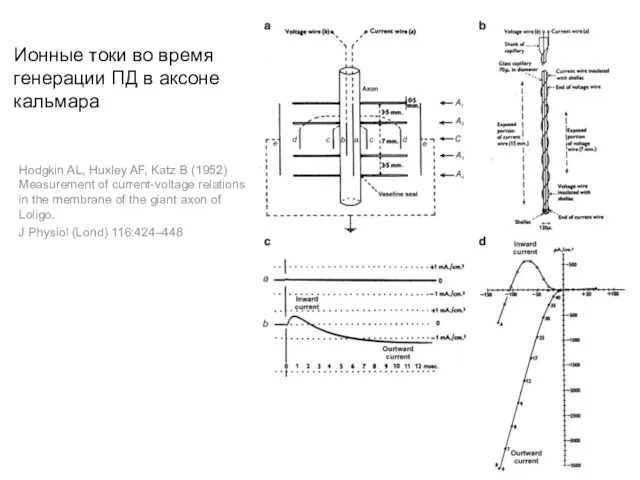
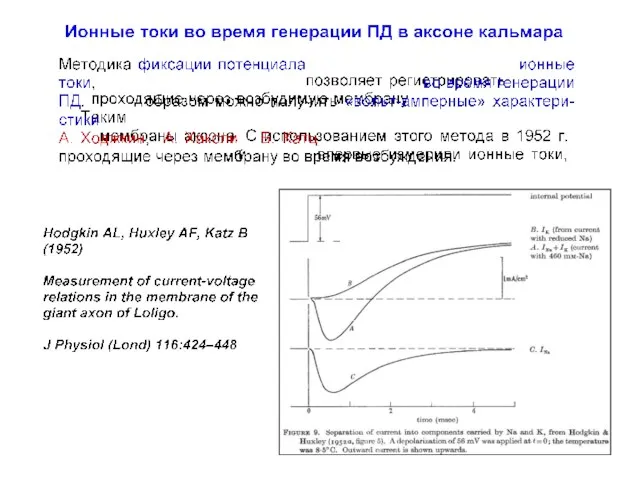
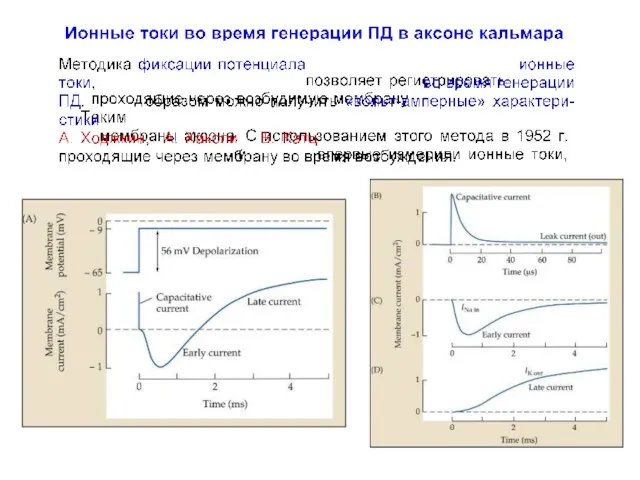
Ионные токи во время генерации ПД в аксоне кальмара
Hodgkin AL, Huxley
Ионные токи во время генерации ПД в аксоне кальмара
Hodgkin AL, Huxley
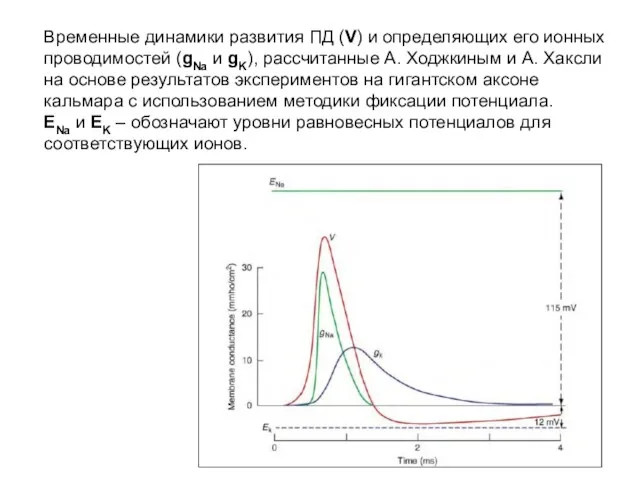
Временные динамики развития ПД (V) и определяющих его ионных проводимостей (gNa
Временные динамики развития ПД (V) и определяющих его ионных проводимостей (gNa
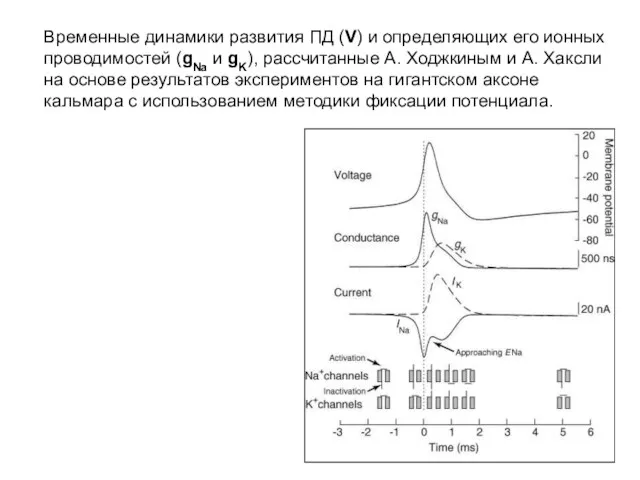
Временные динамики развития ПД (V) и определяющих его ионных проводимостей (gNa
Временные динамики развития ПД (V) и определяющих его ионных проводимостей (gNa

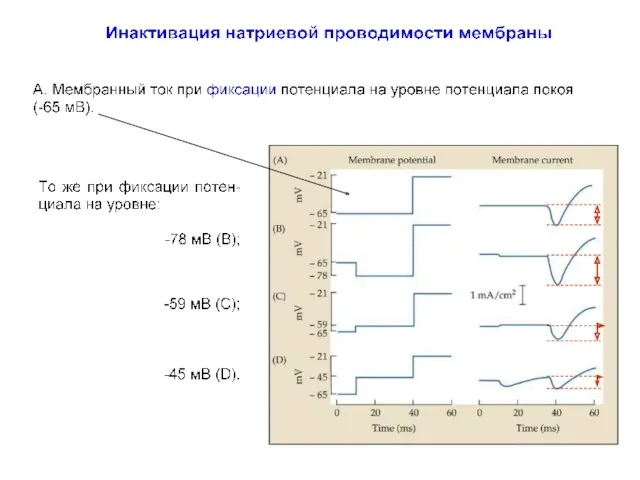
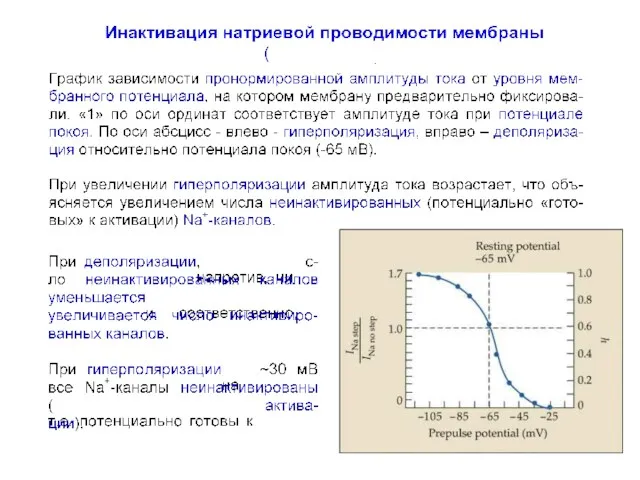
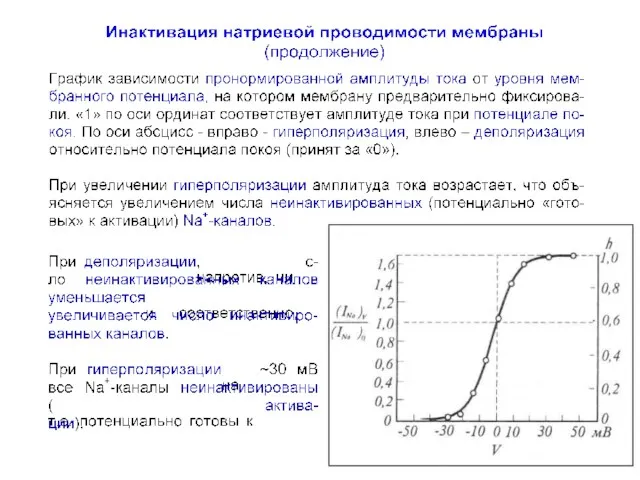

Активационные и инактивационные механизмы проводимости мембраны
Активационные и инактивационные механизмы проводимости мембраны
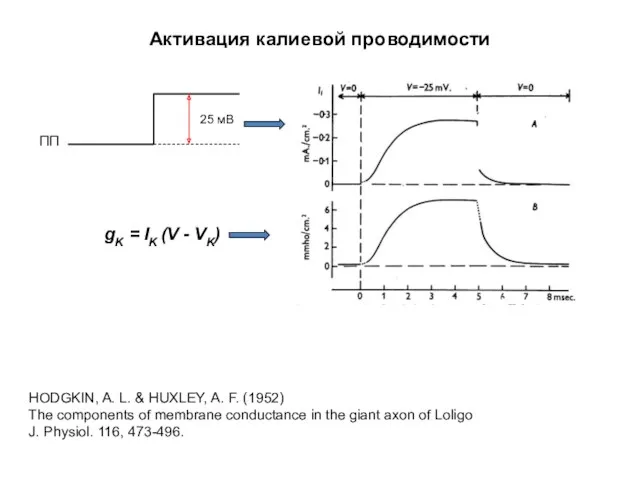
Активация калиевой проводимости
ПП
25 мВ
HODGKIN, A. L. & HUXLEY, A. F. (1952)
The
Активация калиевой проводимости
ПП
25 мВ
HODGKIN, A. L. & HUXLEY, A. F. (1952)
The
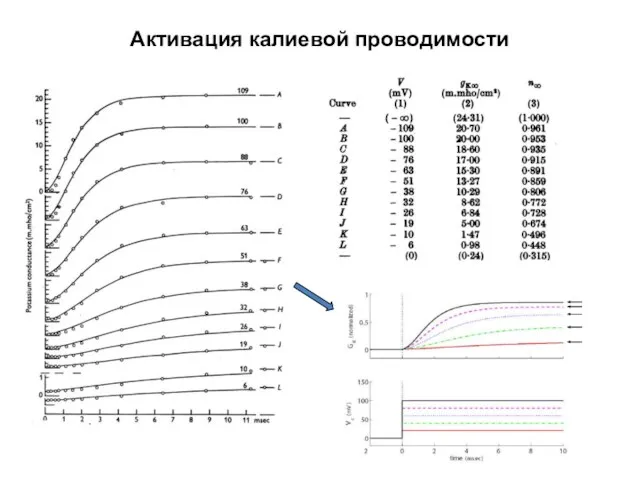
Активация калиевой проводимости
Активация калиевой проводимости
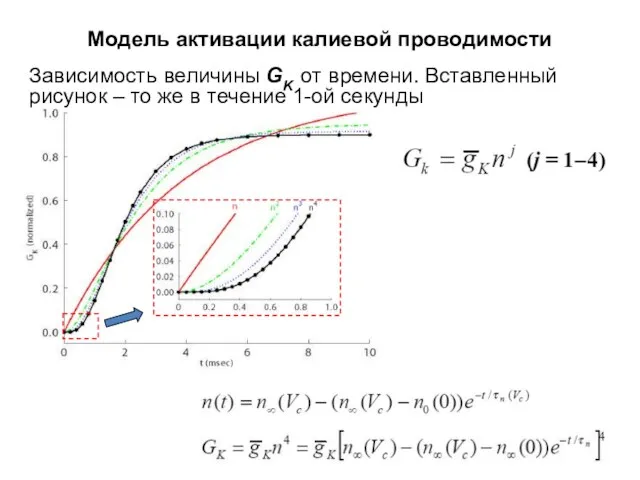
Модель активации калиевой проводимости
Зависимость величины GK от времени. Вставленный рисунок –
Модель активации калиевой проводимости
Зависимость величины GK от времени. Вставленный рисунок –
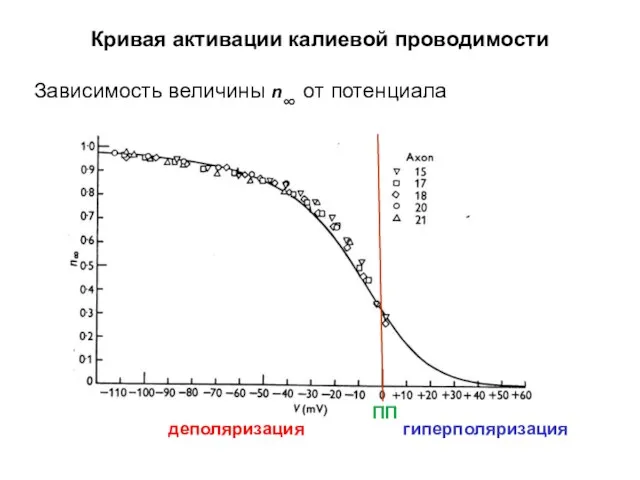
Кривая активации калиевой проводимости
Зависимость величины n∞ от потенциала
деполяризация гиперполяризация
ПП
Кривая активации калиевой проводимости
Зависимость величины n∞ от потенциала
деполяризация гиперполяризация
ПП
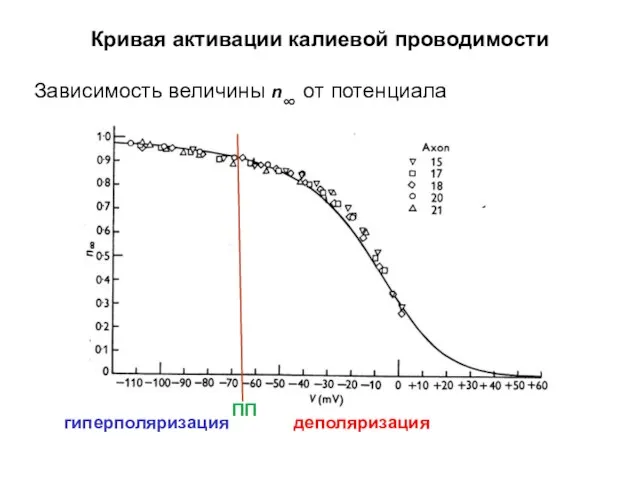
Кривая активации калиевой проводимости
Зависимость величины n∞ от потенциала
гиперполяризация деполяризация
ПП
Кривая активации калиевой проводимости
Зависимость величины n∞ от потенциала
гиперполяризация деполяризация
ПП

Активация калииевой проводимости мембраны
Для K+-проводимости А. Ходжкин и А. Хаксли ввели
Активация калииевой проводимости мембраны
Для K+-проводимости А. Ходжкин и А. Хаксли ввели
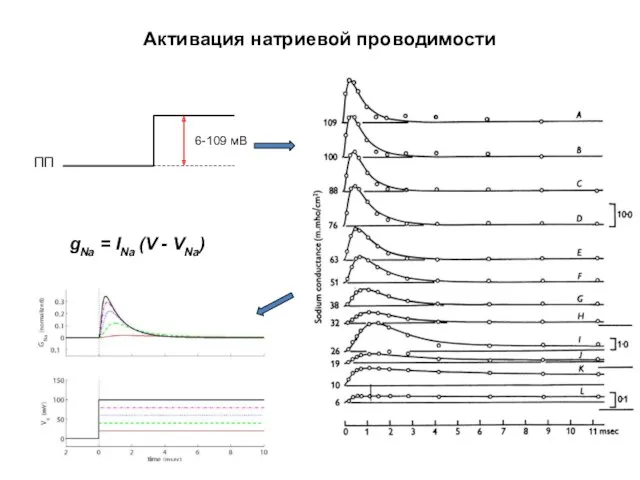
Активация натриевой проводимости
gNa = INa (V - VNa)
Активация натриевой проводимости
gNa = INa (V - VNa)
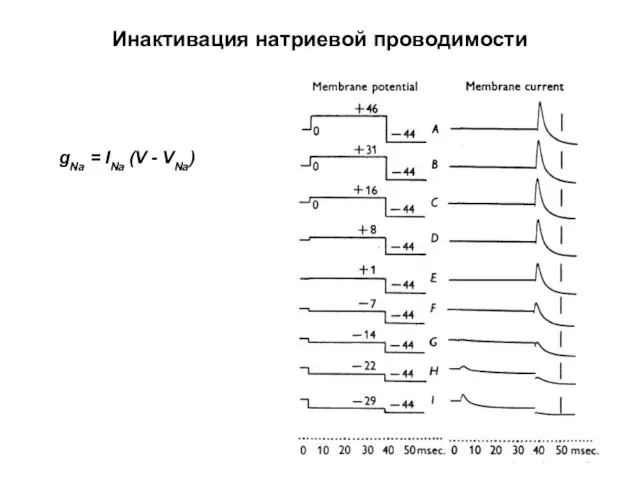
Инактивация натриевой проводимости
gNa = INa (V - VNa)
Инактивация натриевой проводимости
gNa = INa (V - VNa)

Активация и инактивация натриевой проводимости
Активация и инактивация натриевой проводимости
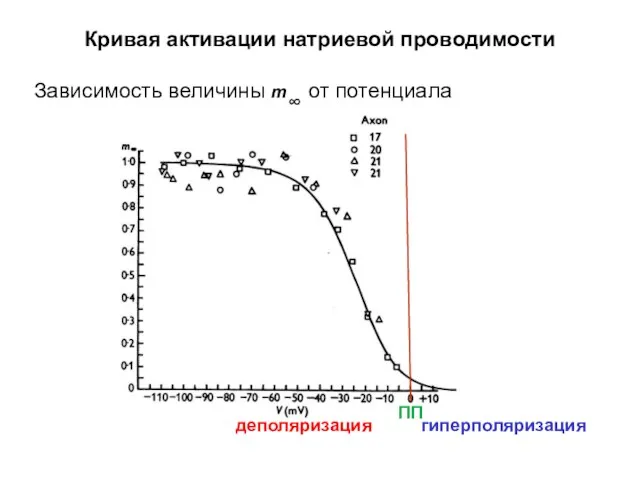
Кривая активации натриевой проводимости
Зависимость величины m∞ от потенциала
деполяризация гиперполяризация
ПП
Кривая активации натриевой проводимости
Зависимость величины m∞ от потенциала
деполяризация гиперполяризация
ПП
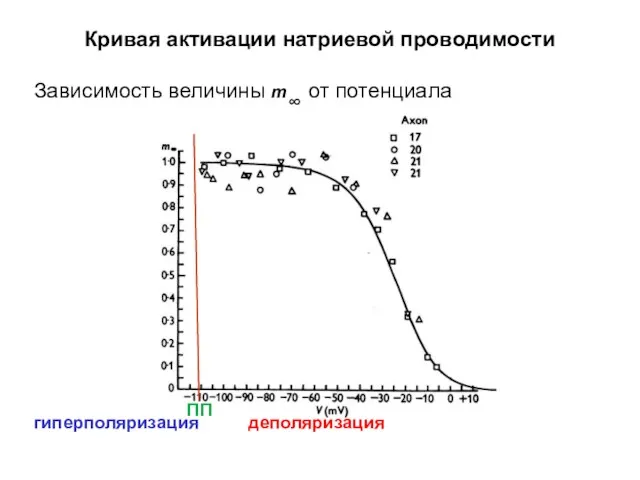
Кривая активации натриевой проводимости
Зависимость величины m∞ от потенциала
гиперполяризация деполяризация
ПП
Кривая активации натриевой проводимости
Зависимость величины m∞ от потенциала
гиперполяризация деполяризация
ПП
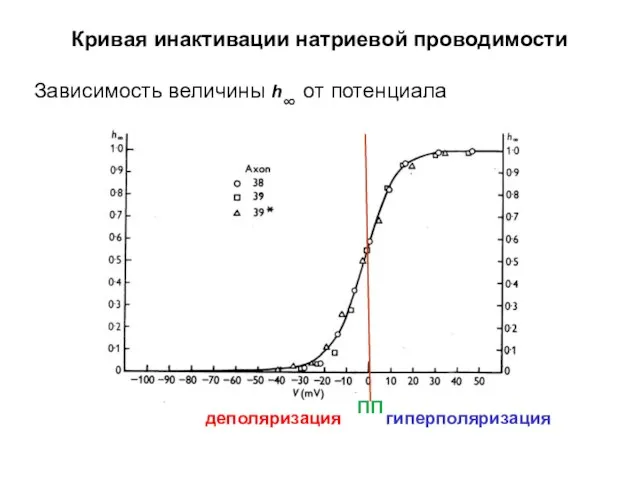
Кривая инактивации натриевой проводимости
Зависимость величины h∞ от потенциала
деполяризация гиперполяризация
ПП
Кривая инактивации натриевой проводимости
Зависимость величины h∞ от потенциала
деполяризация гиперполяризация
ПП
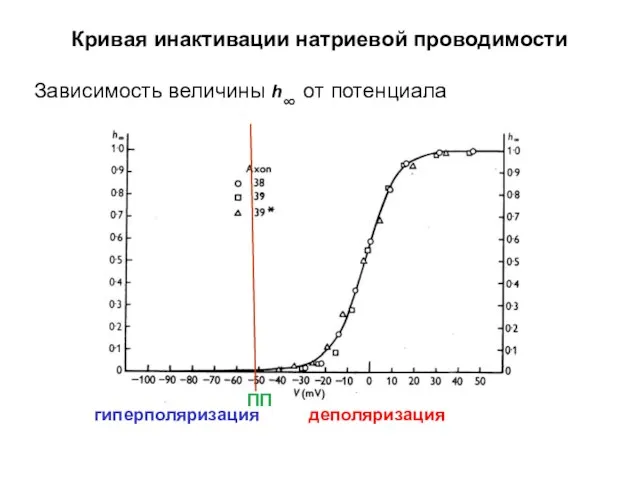
Кривая инактивации натриевой проводимости
Зависимость величины h∞ от потенциала
гиперполяризация деполяризация
ПП
Кривая инактивации натриевой проводимости
Зависимость величины h∞ от потенциала
гиперполяризация деполяризация
ПП
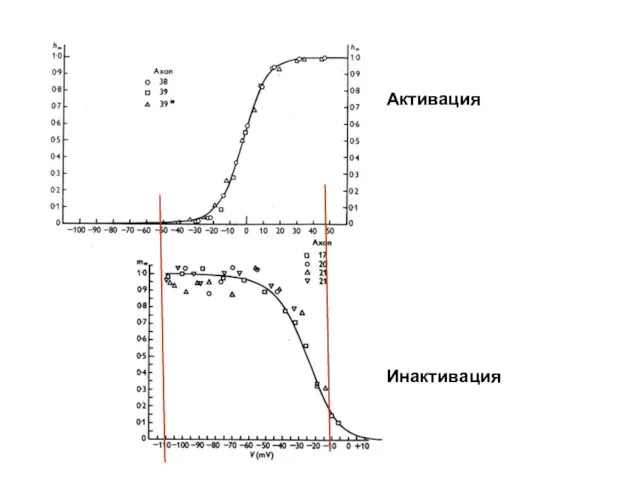
Активация
Инактивация
Активация
Инактивация

Активация и инактивация натриевой проводимости мембраны
Для Na+-проводимости А. Ходжкин и А.
Активация и инактивация натриевой проводимости мембраны
Для Na+-проводимости А. Ходжкин и А.
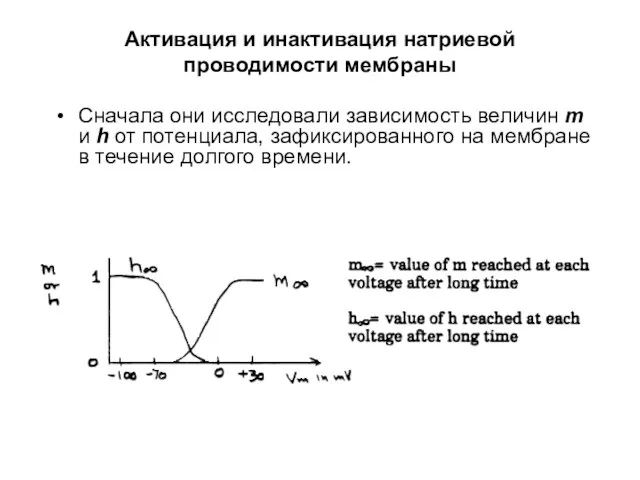
Сначала они исследовали зависимость величин m и h от потенциала, зафиксированного
Сначала они исследовали зависимость величин m и h от потенциала, зафиксированного

Затем они описали зависимость величин m и h от времени.
В клетке
Затем они описали зависимость величин m и h от времени.
В клетке

Инактивация натриевой проводимости мембраны (продолжение)
А. Ходжкин и А. Хаксли предложили простую
Инактивация натриевой проводимости мембраны (продолжение)
А. Ходжкин и А. Хаксли предложили простую

Инактивация натриевой проводимости мембраны (продолжение)
Для отдельного канала переменная m отражает вероятность
Инактивация натриевой проводимости мембраны (продолжение)
Для отдельного канала переменная m отражает вероятность
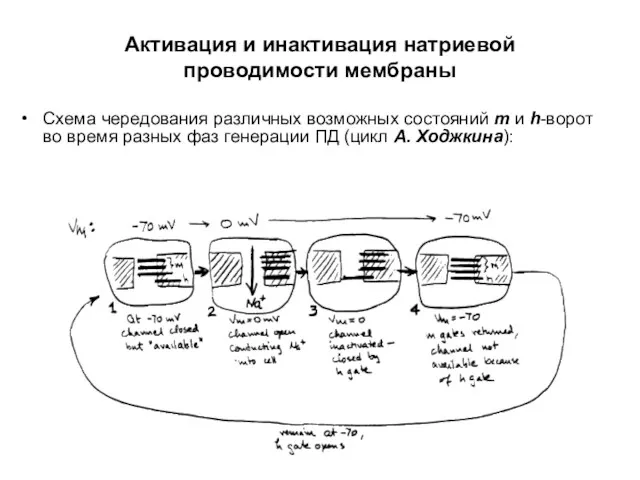
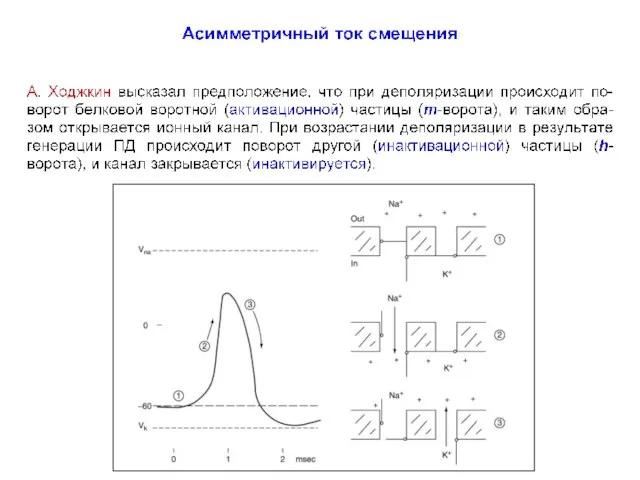
Схема чередования различных возможных состояний m и h-ворот во время разных
Схема чередования различных возможных состояний m и h-ворот во время разных
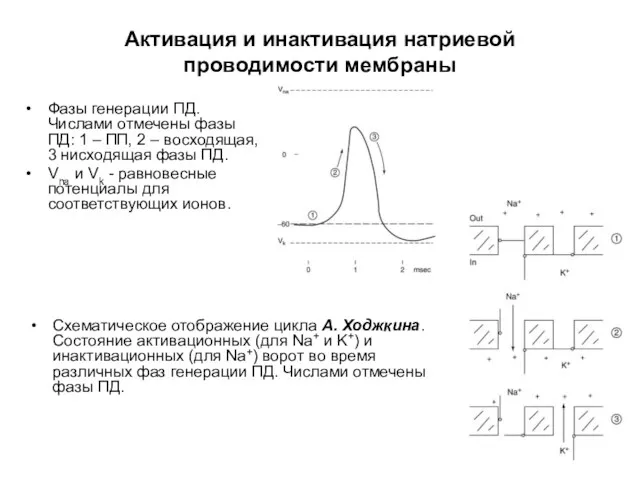
Фазы генерации ПД. Числами отмечены фазы ПД: 1 – ПП, 2
Фазы генерации ПД. Числами отмечены фазы ПД: 1 – ПП, 2
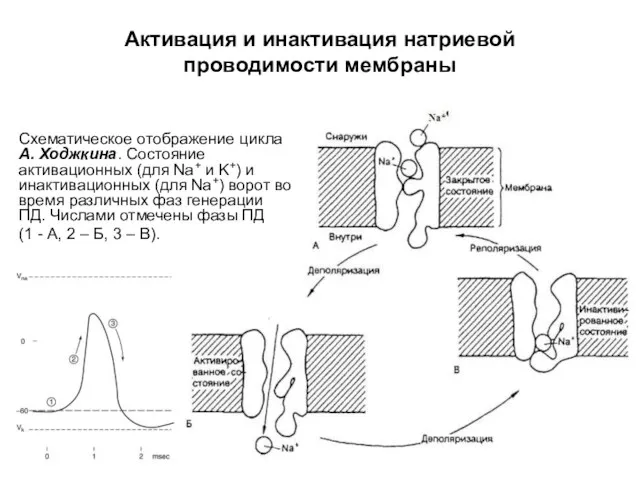
Схематическое отображение цикла А. Ходжкина. Состояние активационных (для Na+ и K+)
Схематическое отображение цикла А. Ходжкина. Состояние активационных (для Na+ и K+)
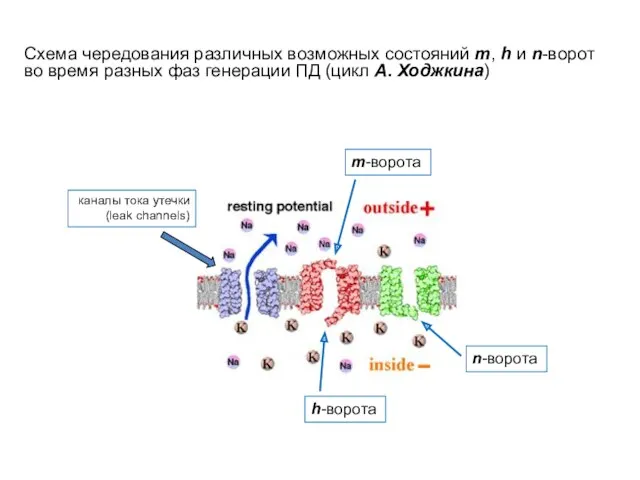
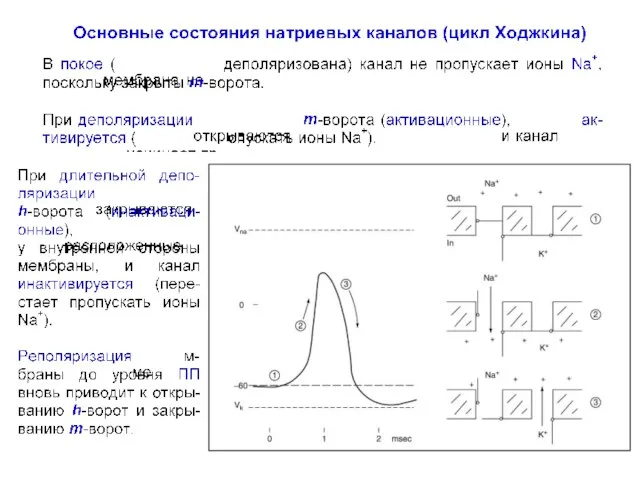
Схема чередования различных возможных состояний m, h и n-ворот во время
Схема чередования различных возможных состояний m, h и n-ворот во время
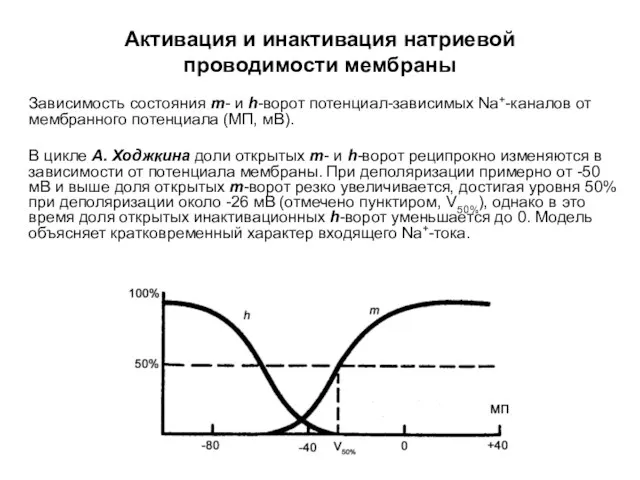
Зависимость состояния m- и h-ворот потенциал-зависимых Na+-каналов от мембранного потенциала (МП,
Зависимость состояния m- и h-ворот потенциал-зависимых Na+-каналов от мембранного потенциала (МП,
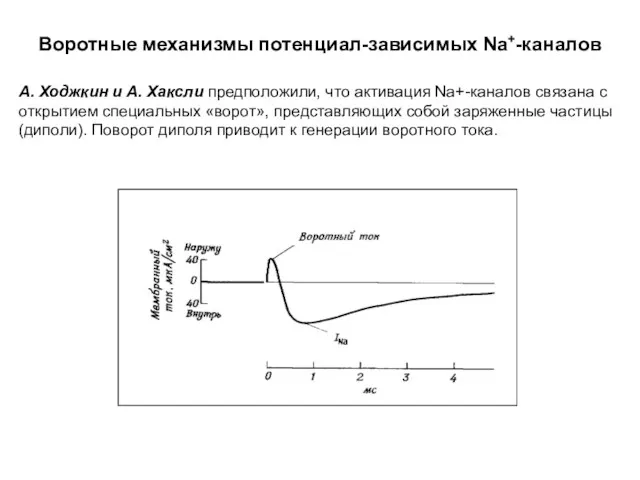
Воротные механизмы потенциал-зависимых Na+-каналов
А. Ходжкин и А. Хаксли предположили, что активация
Воротные механизмы потенциал-зависимых Na+-каналов
А. Ходжкин и А. Хаксли предположили, что активация
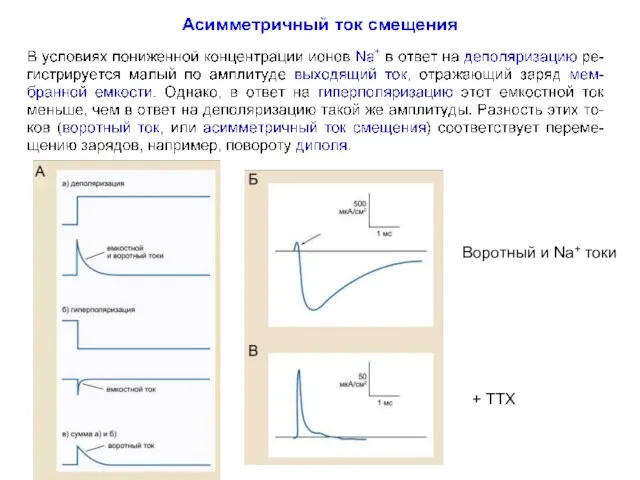
+ TTX
Воротный и Na+ токи
+ TTX
Воротный и Na+ токи

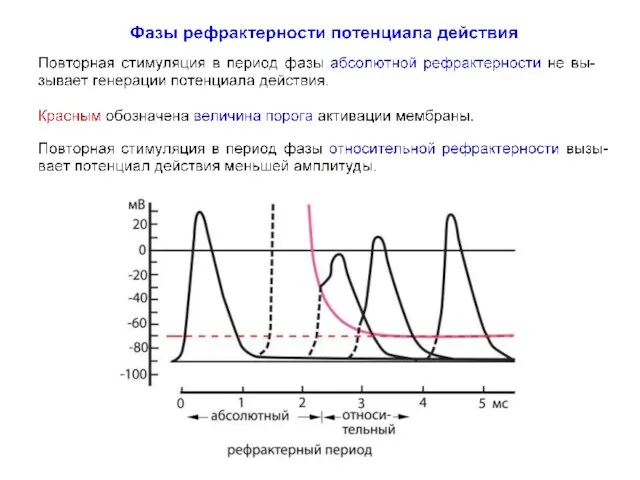
Фазы рефрактерности потенциала действия
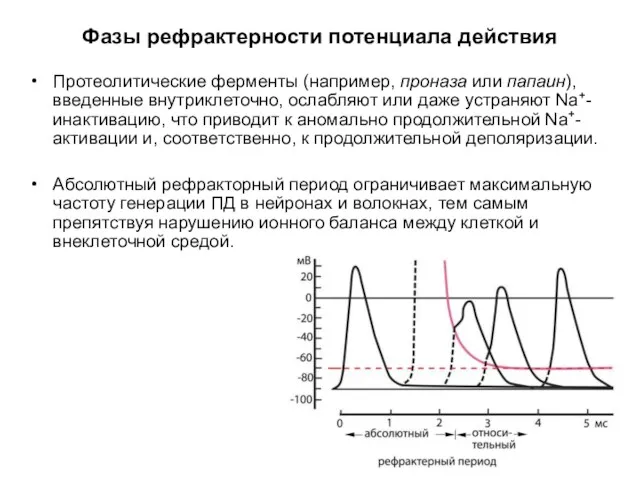
Протеолитические ферменты (например, проназа или папаин), введенные внутриклеточно,
Фазы рефрактерности потенциала действия
Протеолитические ферменты (например, проназа или папаин), введенные внутриклеточно,

Фазы рефрактерности потенциала действия
Воздействие деполяризации мембраны на процессы активации и инактивации
Фазы рефрактерности потенциала действия
Воздействие деполяризации мембраны на процессы активации и инактивации
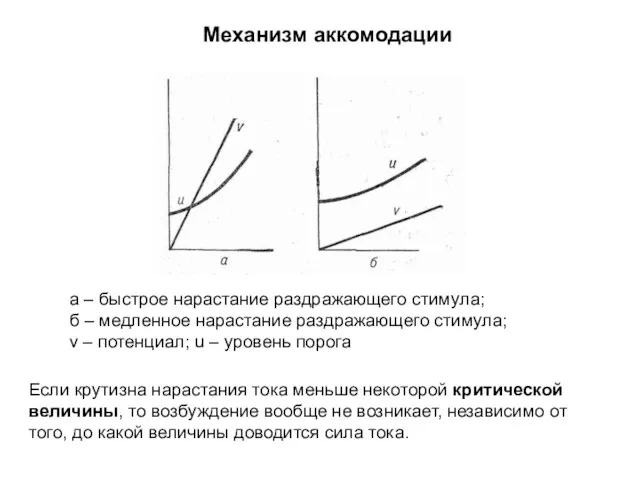
а – быстрое нарастание раздражающего стимула;
б – медленное нарастание раздражающего стимула;
v
а – быстрое нарастание раздражающего стимула;
б – медленное нарастание раздражающего стимула;
v
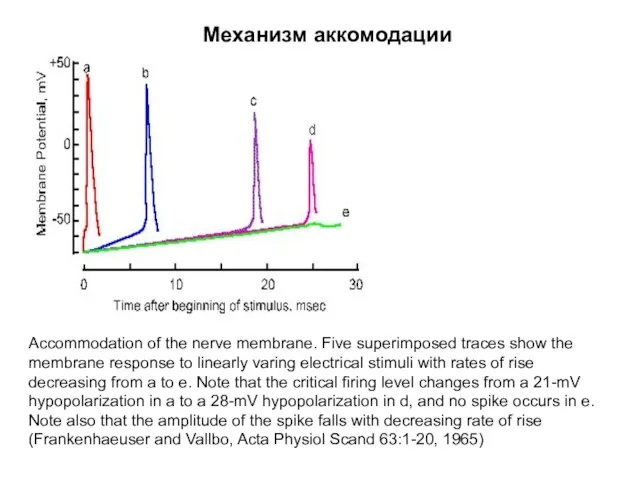
Механизм аккомодации
Accommodation of the nerve membrane. Five superimposed traces show the
Механизм аккомодации
Accommodation of the nerve membrane. Five superimposed traces show the
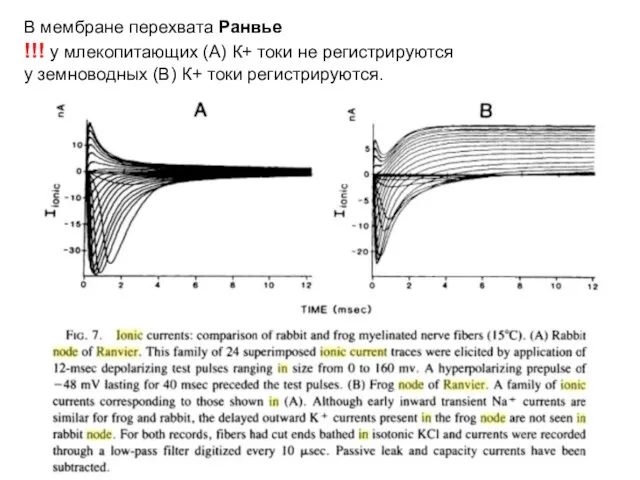
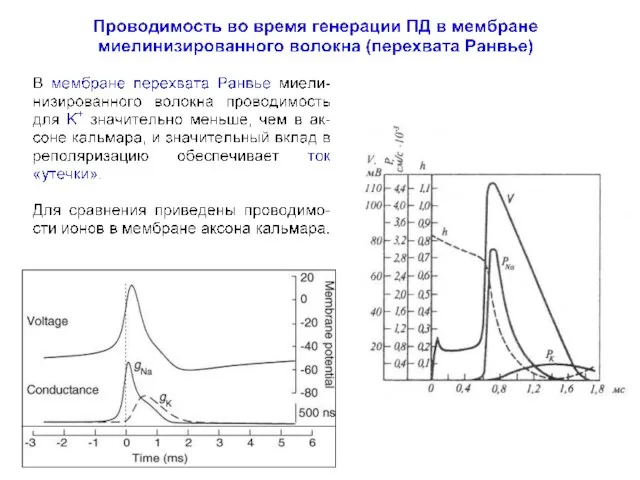
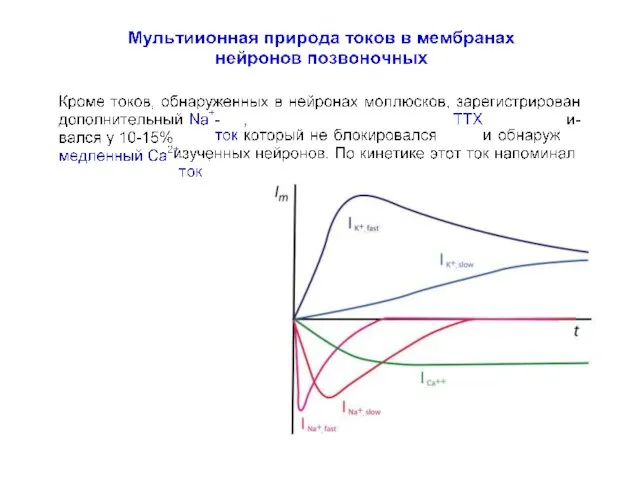
В мембране перехвата Ранвье
!!! у млекопитающих (А) К+ токи не регистрируются
у
В мембране перехвата Ранвье
!!! у млекопитающих (А) К+ токи не регистрируются
у

Генерация ПД в нейронах
Аксоны представляют собой относительно простое мембранное образование, обеспечивающее
Генерация ПД в нейронах
Аксоны представляют собой относительно простое мембранное образование, обеспечивающее
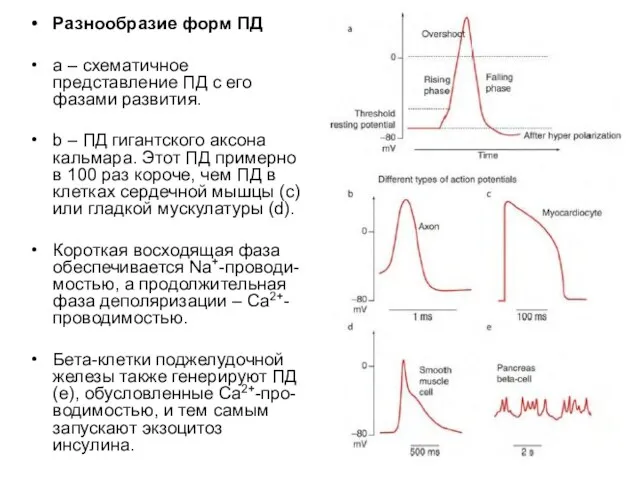
Разнообразие форм ПД
a – схематичное представление ПД с его фазами развития.
Разнообразие форм ПД
a – схематичное представление ПД с его фазами развития.

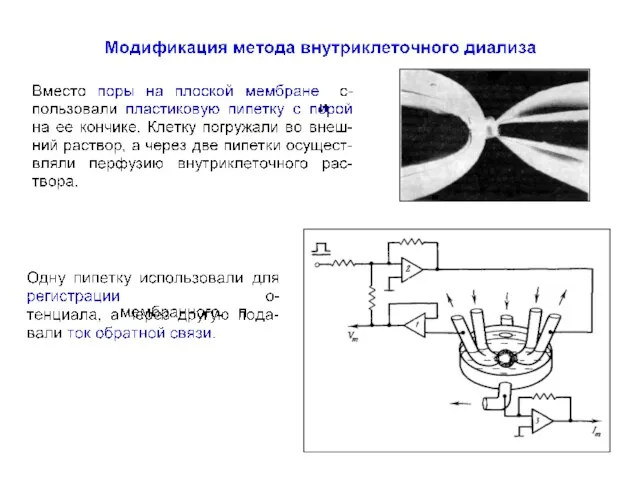
Метод внутриклеточного диализа
Метод разработан в Институте физиологии АН УССР (г. Киев)
Метод внутриклеточного диализа
Метод разработан в Институте физиологии АН УССР (г. Киев)
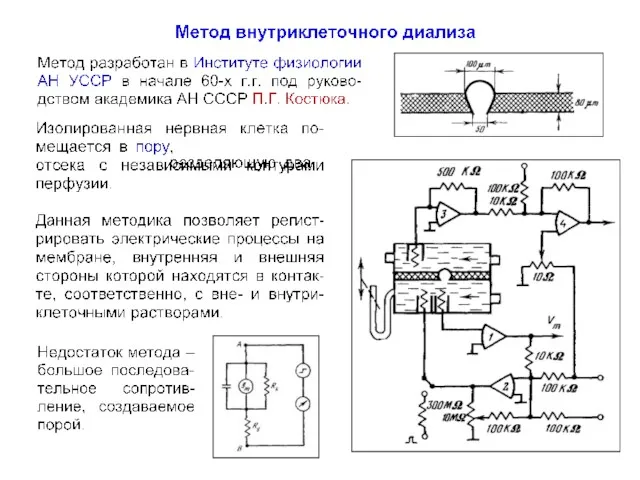
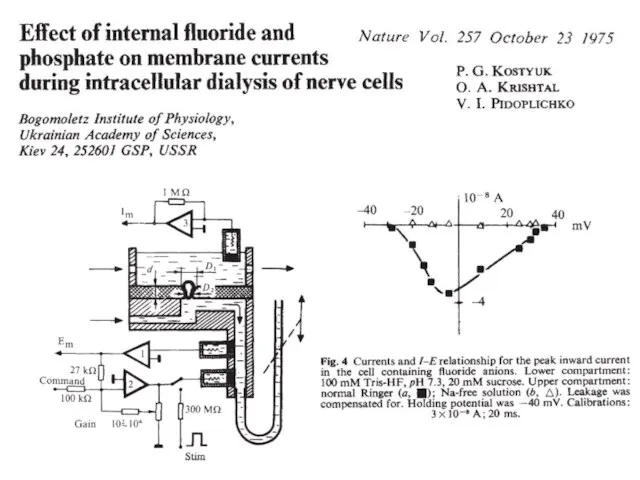
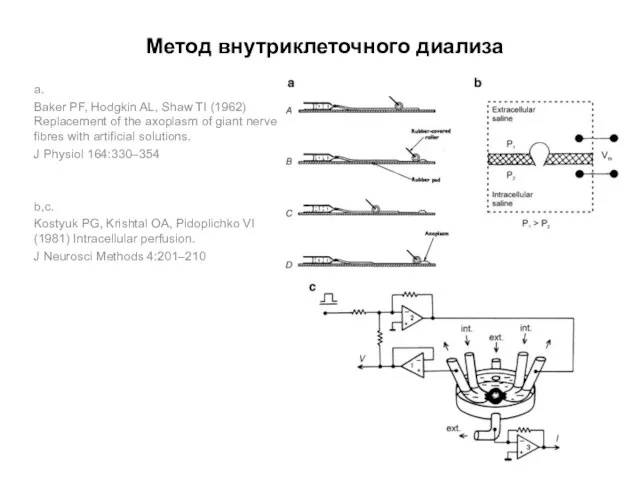
Метод внутриклеточного диализа
a.
Baker PF, Hodgkin AL, Shaw TI (1962) Replacement of
Метод внутриклеточного диализа
a.
Baker PF, Hodgkin AL, Shaw TI (1962) Replacement of
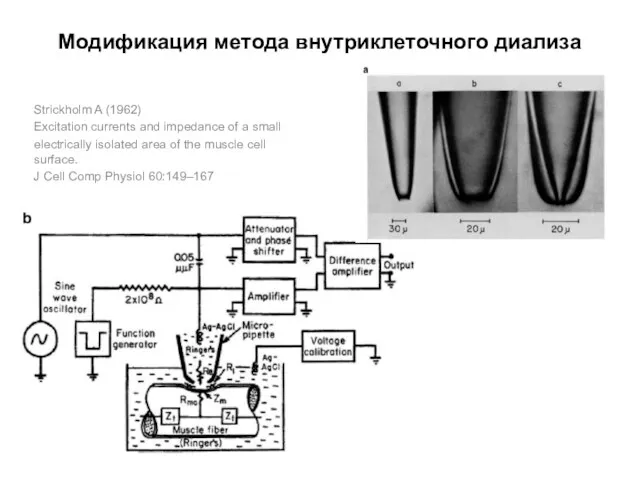
Модификация метода внутриклеточного диализа
Strickholm A (1962)
Excitation currents and impedance of a
Модификация метода внутриклеточного диализа
Strickholm A (1962)
Excitation currents and impedance of a
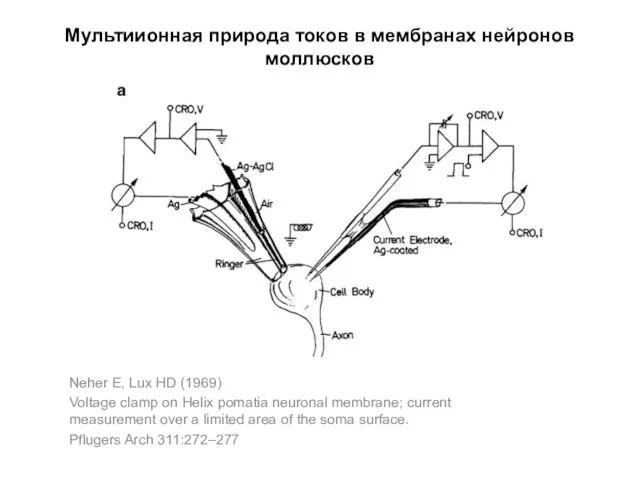
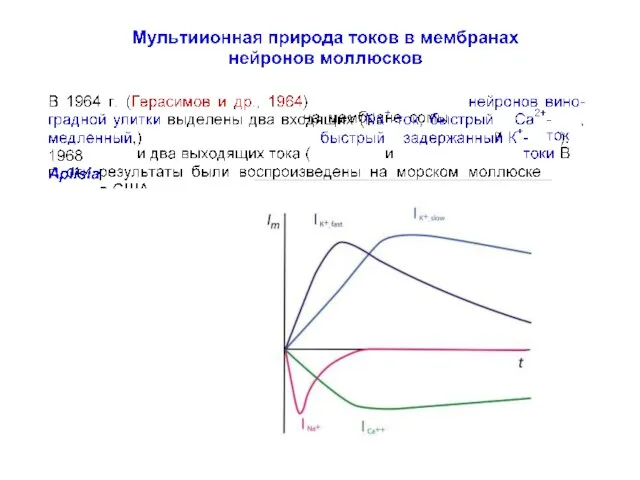
Мультиионная природа токов в мембранах нейронов моллюсков
Neher E, Lux HD (1969)
Voltage
Мультиионная природа токов в мембранах нейронов моллюсков
Neher E, Lux HD (1969)
Voltage

Мультиионная природа токов в мембранах нейронов моллюсков
Neher E, Lux HD (1969)
Voltage
Мультиионная природа токов в мембранах нейронов моллюсков
Neher E, Lux HD (1969)
Voltage

Герасимов В.Д., Костюк П.Г., Майский В.А. Возбудимость гигантских нервных клеток различных
Герасимов В.Д., Костюк П.Г., Майский В.А. Возбудимость гигантских нервных клеток различных
 Презентации к урокам 9 - 11 класс
Презентации к урокам 9 - 11 класс Значение дыхания. Органы дыхания
Значение дыхания. Органы дыхания Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс
Дополнительная презентация Что такое фитодизайн?к элективному курсу Основы фитодизайна, 9 класс Проводящие ткани. Проводящие пучки
Проводящие ткани. Проводящие пучки Система кровообращения. Большой и малый круги кровообращения. Физиология и основы анатомии сердца
Система кровообращения. Большой и малый круги кровообращения. Физиология и основы анатомии сердца Белки. Качественный состав белков
Белки. Качественный состав белков Видоизменения побегов растения
Видоизменения побегов растения Селекция растений
Селекция растений презентация о результатах педагогической деятельности учителя биологии Анохиной Е.И.
презентация о результатах педагогической деятельности учителя биологии Анохиной Е.И. Вид. Структура и критерии вида
Вид. Структура и критерии вида Роль лишайников в почвообразовании
Роль лишайников в почвообразовании Тип плоские черви
Тип плоские черви Видообразование
Видообразование Морфология бактерий
Морфология бактерий Высшие психические функции. Память
Высшие психические функции. Память Лекарственные растения Воронежской области
Лекарственные растения Воронежской области Вода и ее свойства
Вода и ее свойства Хвостатые земноводные
Хвостатые земноводные Комнатные растения в интерьере
Комнатные растения в интерьере Экология. Экосистемы
Экология. Экосистемы Рост и развитие растений
Рост и развитие растений Введение (Биология – Медицина – Человек). Молекулярно- клеточный уровень организации живого. Лекция №1
Введение (Биология – Медицина – Человек). Молекулярно- клеточный уровень организации живого. Лекция №1 Удивительное существо кошка
Удивительное существо кошка Строение и функции кожи человека
Строение и функции кожи человека Собственно- соединительные ткани
Собственно- соединительные ткани Анатомия вегетативных органов
Анатомия вегетативных органов Сердечно – сосудистая система
Сердечно – сосудистая система African - plains
African - plains