- Происхождение человека

Содержание
- 3. 1871 г. “Происхождение человека и половой отбор”
- 5. 1829-1830 г.; Пещера Анжи, Бельгия (череп ребенка). 1848 г.; Форбс Куорри (карьер Форбса), Гибралтар 1856 г.;
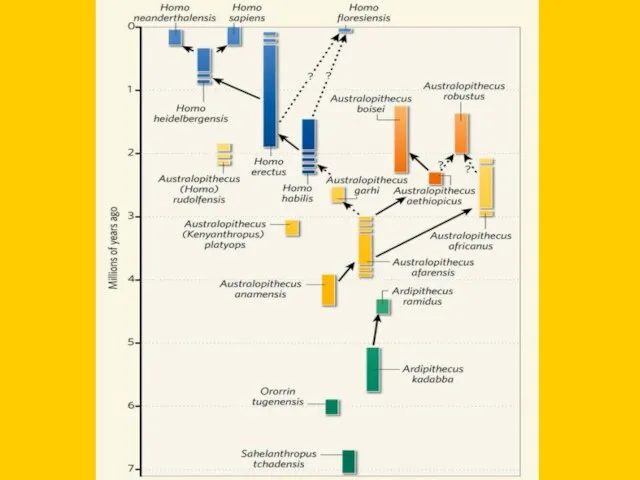
- 6. Филогенетическое древо приматов
- 7. “Линейная” эволюция гоминид по старым представлениям
- 8. A.africanus Paranthropus boisei H.habilis H.erectus H.neanderthalensis H.sapiens
- 9. Фрагмент “древа” гоминин: тупиковые ветви и “альтернативные пути”
- 10. Схема эволюции гоминин © Copyright Smithsonian Institution http://antropology.si.edu/humanorigins/ha/a_tree.html
- 11. Схема эволюции гоминин © Copyright Smithsonian Institution
- 12. Sahelanthropus tchadensis (2001 год, озеро Чад, 6-7 млн лет назад) затылочное отверстие уже было сдвинуто вперед
- 13. Orrorin tugenensis (2000 год, Кения, ок. 6 млн лет) прямохождение
- 14. Ardipithecus ramidus (1992 год, Эфиопия, ок. 4,5-4,3 млн лет) 300-370см3
- 15. Австралопитек афарский (Australopithecus afarensis) 3-4 млн лет Berger, L.R., de Ruiter, D.J., Churchill, S.E., Schmid, P.,
- 16. Paranthropus boisei 2,2-1,2 млн лет 500 - 550 см3
- 17. Australopithecus sediba 1,97-1,95 млн лет Berger, L.R., de Ruiter, D.J., Churchill, S.E., Schmid, P., Carlson, K.J.,
- 18. Homo rudolfensis 2,5 - 1,9 млн лет 526 - 700 см3
- 19. Homo habilis 1,9 - 2,4 миллиона лет назад 650см3 (500-870см3)
- 20. H. ergaster, около 1,9 млн лет 880см3
- 21. Homo georgicus 1,75 млн.лет (Homo habilis ), Дманиси (Грузия) Давид Лордкипанидзе 600-680 см3
- 22. Homo erectus 1,75 млн лет назад 900-1100 см3
- 23. Слева направо: следы австралопитека 3,6–3,7 млн лет; следы Homo erectus 1,46 и 1,53 млн лет; следы
- 24. Синантроп (Homo erectus) 1,3 млн лет назад (до 40 тыс лет)
- 25. Первый выход Homo erectus из Африки
- 26. H. floresiensis, о. Флорес, Индонезия, 95-12 тыс. лет
- 27. трапециевидная головчатая ладьевидная H. floresiensis
- 28. Homo antecessor (поздний H.erectus ?) 880 тыс лет (Испания) Eudald Carbonell et al. The first hominin
- 29. Homo heidelbergensis ~ 800 тыс.лет
- 30. Homo heidelbergensis ~ 800-200 тыс.лет 1100-1400 см3
- 32. Мозг и огонь Резкое увеличение мозга → рост энергозатрат. Чем компенсировать? Ранние эректусы: рост доли мяса
- 33. Слепки мозговой полости (эндокраны) Australopithecus Homo erectus Homo neanderthalensis Homo sapiens
- 34. Homo neanderthalensis Homo sapiens 900-2000 см3 200 000 - 28 000 лет
- 35. Homo neanderthalensis Homo sapiens
- 37. Неандертальцы были рыжими и светлокожими У неандертальцев обнаружена мутация в гене меланокортинового рецептора (MC1R), частично выводящая
- 38. Lalueza-Fox et al. A melanocortin 1 receptor allele suggests varying pigmentation among Neanderthals // Science. 2007.
- 39. «… коренное неандертальское население Европы пало под натиском ближневосточных пришельцев значительно быстрее, чем считалось. Превосходство сапиенсов
- 40. Johannes Krause, Ludovic Orlando, David Serre, Bence Viola, Kay Prüfer, Michael P. Richards, Jean-Jacques Hublin, Catherine
- 41. В Денисовой пещере (Алтай) в 2005 г. была найдена кость индивидуума, жившего 30000-48000 лет назад, где
- 42. Распределение числа нуклеотидных отличий при попарном сравнении последовательностей мтДНК Krause J., Fu Q., Good J. M.,
- 43. Цифрами показаны датировки (в тысячах лет), полученные при помощи метода «молекулярных часов». Эволюционное дерево мтДНК гоминид
- 44. Генетический обмен между древними и современными гоминидами Paabo, S. The human condition — a molecular approach.
- 45. Homo neanderthalensis Marcia S. Ponce de Leуn, Lubov Golovanova, Vladimir Doronichev, Galina Romanova, Takeru Akazawa, Osamu
- 46. шимпанзе кроманьонцы неандертальцы Рост мозга в % к его объему … ген FOXP2, связанный с речью
- 47. Реконструированная генеалогия аллелей гена microcephalin Красным показаны не-D-аллели, голубым — D-аллели Patrick D. Evans, Nitzan Mekel-Bobrov,
- 49. Черепа кроманьонцев
- 51. Карта возможного расселения Homo sapiens sapiens
- 52. Небольшая популяция в восточной Африке 160-200 тыс лет mtDNA Ychromosome
- 53. Первый выход H.sapiens из Африки ~ 135-115 тыс лет назад.
- 54. H.sapiens в Передней Азии вскоре исчезают 115-90 тыс лет
- 55. Второй выход H.sapiens из Африки 90-85 тыс лет
- 56. Расселение H.sapiens вдоль южного побережья Азии 85-75 тыс лет
- 57. ~ 74 000 лет - извержение вулкана Тоба на Суматре 10000 adults
- 58. Продолжение расселения H.sapiens, проникновение в Австралию
- 59. Приход H.sapiens в Европу, населенную неандертальцами
- 60. Rasmussen et al (2011) An Aboriginal Australian Genome Reveals Separate Human Dispersals into Asia // Science
- 61. Заселение сапиенсами Европы Колонизация Центральной и Западной Европы: 46-41 тыс. лет назад. Скорость продвижения: 400 м
- 62. Alan R. Templeton. Haplotype Trees and Modern Human Origins // Yearbook of physical anthropology. 48:33–59 (2005).
- 63. Расселяясь из Африки, наши предки теряли гены и становились более похожими Синие точки — мужские черепа,
- 64. Изменчивость размеров черепа
- 65. Снижение коэффициента изменчивости размеров черепа по мере удаления от центра происхождения
- 66. Генотипическая изменчивость
- 67. полиморфизмы Alu (повторы в 1 и 7 хромосомах)
- 69. Сравнение уровня активности 907 генов у людей, шимпанзе, орангутанов и макаков-резусов выявило 30 генов, активность которых
- 70. Эволюционная психология
- 71. Корреляция между развитием мозга (неокортекса) и размером коллектива
- 72. Размер коллективов у гоминин (оцененный на основе размеров мозга)
- 73. Esther Herrmann, Josep Call, Maria Victoria Hernandez-Lloreda, Brian Hare, Michael Tomasello. Humans Have Evolved Specialized Skills
- 74. По горизонтальной оси — средний возраст обезьян в паре, по вертикальной — количество тестов Victoria Wobbersend,
- 75. “Культурное наследование” у животных Умение колоть орехи в некоторых популяциях шимпанзе тысячелетиями передается из поколения в
- 76. Zoe R. Donaldson, Larry J. Young. Oxytocin, Vasopressin, and the Neurogenetics of Sociality // Science. 2008.
- 77. Близнецовый анализ показал, что склонность к добрым поступкам, доверчивость и благодарность имеют в значительной мере генетическую
- 78. Реципрокный (взаимный) альтруизм Стратегия: Помоги другому, и он в будущем поможет тебе. А если не поможет,
- 79. Распределение детей по “нравственным категориям” Fehr Е., Bernhard Н., Rockenbach В. Egalitarianism in young children //
- 80. Альтруизм среди “своих” и враждебность к чужакам: две стороны одной медали Дети до 3-4 лет ведут
- 81. Межгрупповые войны - причина альтруизма? Уровень межгрупповой агрессии у палеолитических охотников-собирателей был достаточно высок, чтобы обеспечить
- 82. р —доля альтруистов, w— численность популяции S. Chuang, Olivier Rivoire, Stanislas Leibler. Simpson's Paradox in a
- 83. Правило Гамильтона: «ген альтруизма» будет поддержан отбором и распространится в популяции, если: rB >C r -
- 84. Религия как адаптация и парохиальный альтруизм Выживаемость 200 замкнутых общин, возникших в Америке в XIX в
- 85. Резникова Ж.И. Социальное обучение у животных // Природа. 2009. №5. «...Учиться у взрослых быстрее и безопаснее...»
- 86. Есть мнение, что главным стимулом для развития речи у наших предков была необходимость посплетничать. Сплетни —
- 87. У людей, по-видимому, есть врожденные психологические свойства (“предрасположенности”), направленные на эффективное выявление обманщиков; “Costly punishment” -
- 88. “Комплекс гоминизации” (А.А. Зубов) высокоразвитый мозг (энцефализация); анатомические особенности руки и возможность её освобождения для использования,
- 90. Скачать презентацию


1871 г. “Происхождение человека и половой отбор”
1871 г. “Происхождение человека и половой отбор”
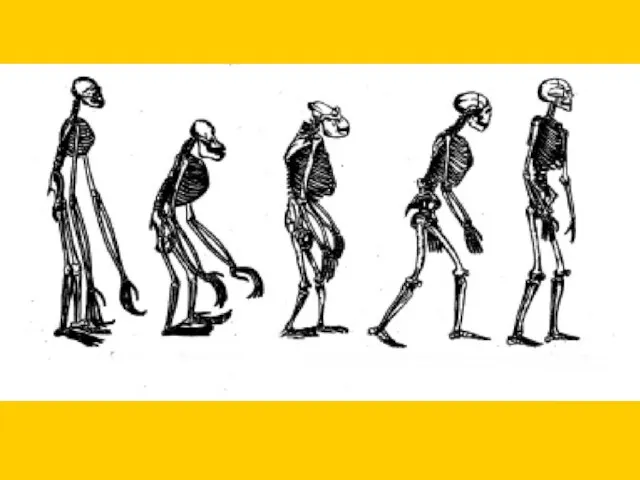

1829-1830 г.; Пещера Анжи, Бельгия (череп ребенка).
1848 г.; Форбс Куорри (карьер
1829-1830 г.; Пещера Анжи, Бельгия (череп ребенка).
1848 г.; Форбс Куорри (карьер
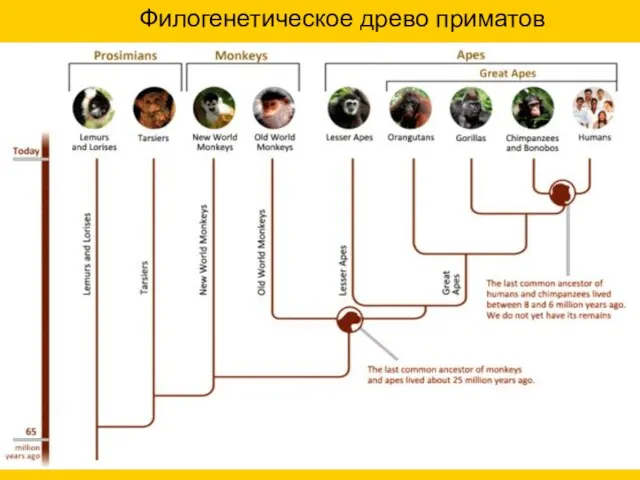
Филогенетическое древо приматов
Филогенетическое древо приматов
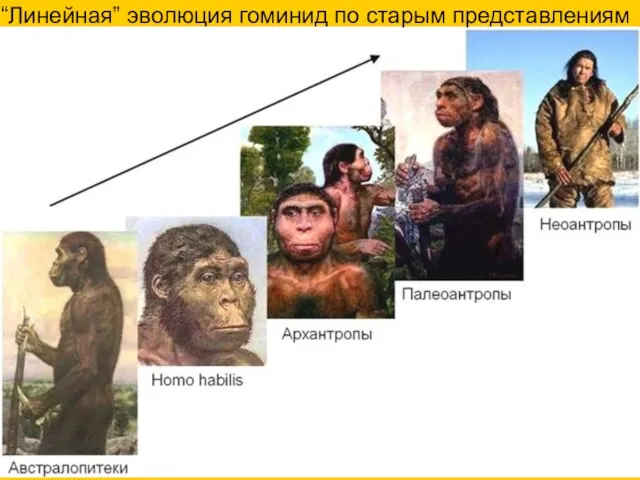
“Линейная” эволюция гоминид по старым представлениям
“Линейная” эволюция гоминид по старым представлениям
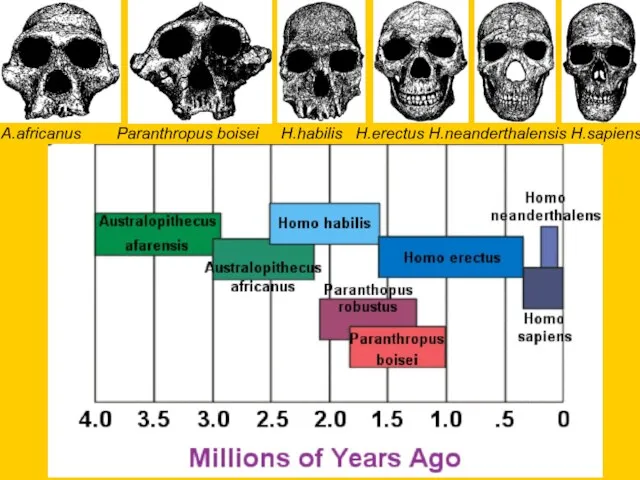
A.africanus Paranthropus boisei H.habilis H.erectus H.neanderthalensis H.sapiens
A.africanus Paranthropus boisei H.habilis H.erectus H.neanderthalensis H.sapiens
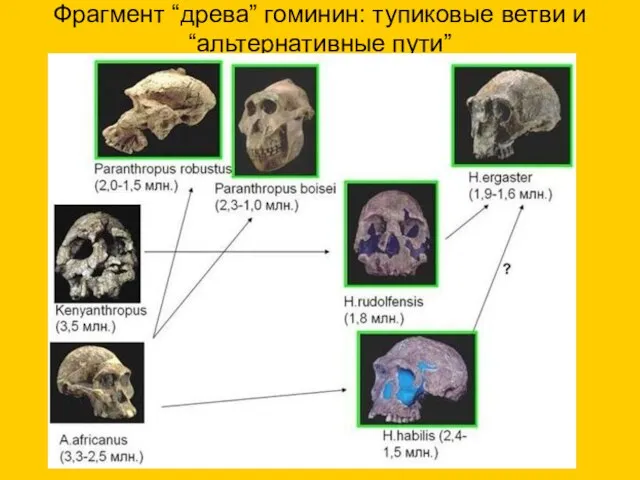
Фрагмент “древа” гоминин: тупиковые ветви и “альтернативные пути”
Фрагмент “древа” гоминин: тупиковые ветви и “альтернативные пути”
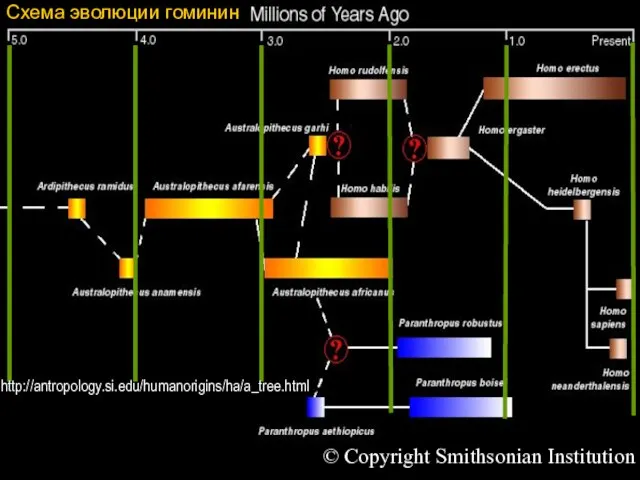
Схема эволюции гоминин
© Copyright Smithsonian Institution
http://antropology.si.edu/humanorigins/ha/a_tree.html
Схема эволюции гоминин
© Copyright Smithsonian Institution
http://antropology.si.edu/humanorigins/ha/a_tree.html
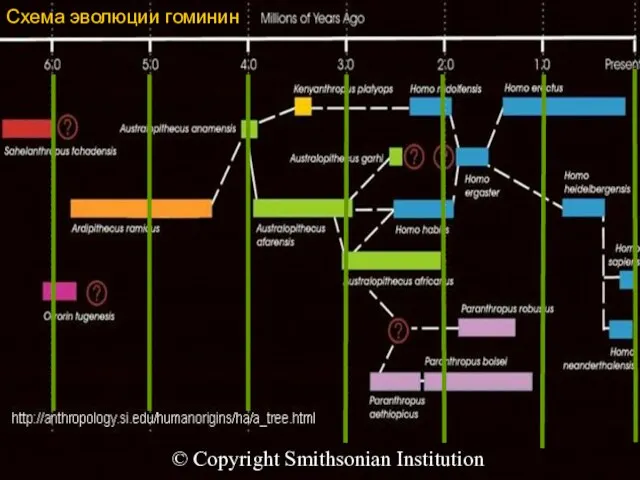
Схема эволюции гоминин
© Copyright Smithsonian Institution
Схема эволюции гоминин
© Copyright Smithsonian Institution

Sahelanthropus tchadensis (2001 год, озеро Чад, 6-7 млн лет назад)
затылочное
Sahelanthropus tchadensis (2001 год, озеро Чад, 6-7 млн лет назад)
затылочное

Orrorin tugenensis (2000 год, Кения, ок. 6 млн лет)
прямохождение
Orrorin tugenensis (2000 год, Кения, ок. 6 млн лет)
прямохождение

Ardipithecus ramidus (1992 год, Эфиопия, ок. 4,5-4,3 млн лет)
300-370см3
Ardipithecus ramidus (1992 год, Эфиопия, ок. 4,5-4,3 млн лет)
300-370см3

Австралопитек афарский (Australopithecus afarensis) 3-4 млн лет
Berger, L.R., de Ruiter, D.J.,
Австралопитек афарский (Australopithecus afarensis) 3-4 млн лет
Berger, L.R., de Ruiter, D.J.,

Paranthropus boisei
2,2-1,2 млн лет
500 - 550 см3
Paranthropus boisei
2,2-1,2 млн лет
500 - 550 см3

Australopithecus sediba
1,97-1,95 млн лет
Berger, L.R., de Ruiter, D.J., Churchill, S.E., Schmid,
Australopithecus sediba
1,97-1,95 млн лет
Berger, L.R., de Ruiter, D.J., Churchill, S.E., Schmid,

Homo rudolfensis 2,5 - 1,9 млн лет
526 - 700 см3
Homo rudolfensis 2,5 - 1,9 млн лет
526 - 700 см3

Homo habilis 1,9 - 2,4 миллиона лет назад
650см3
(500-870см3)
Homo habilis 1,9 - 2,4 миллиона лет назад
650см3
(500-870см3)

H. ergaster, около 1,9 млн лет
880см3
H. ergaster, около 1,9 млн лет
880см3

Homo georgicus 1,75 млн.лет (Homo habilis ), Дманиси (Грузия)
Давид Лордкипанидзе
600-680
Homo georgicus 1,75 млн.лет (Homo habilis ), Дманиси (Грузия)
Давид Лордкипанидзе
600-680
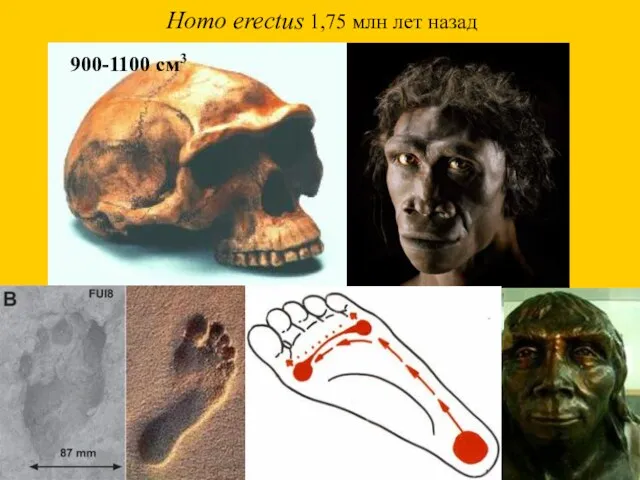
Homo erectus 1,75 млн лет назад
900-1100 см3
Homo erectus 1,75 млн лет назад
900-1100 см3
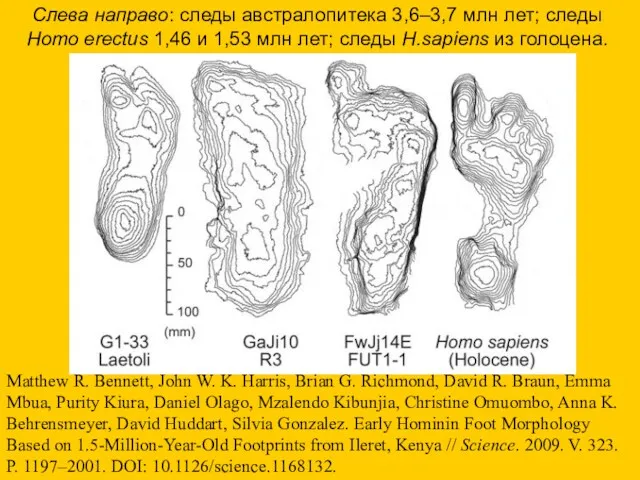
Слева направо: следы австралопитека 3,6–3,7 млн лет; следы Homo erectus 1,46
Слева направо: следы австралопитека 3,6–3,7 млн лет; следы Homo erectus 1,46

Синантроп (Homo erectus) 1,3 млн лет назад (до 40 тыс лет)
Синантроп (Homo erectus) 1,3 млн лет назад (до 40 тыс лет)
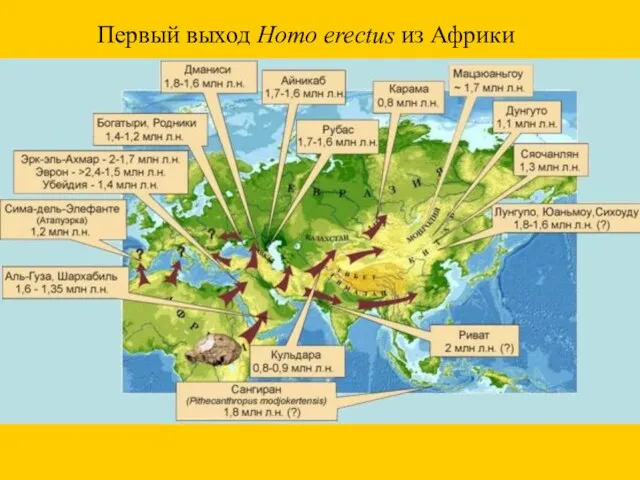
Первый выход Homo erectus из Африки
Первый выход Homo erectus из Африки

H. floresiensis, о. Флорес, Индонезия,
95-12 тыс. лет
H. floresiensis, о. Флорес, Индонезия,
95-12 тыс. лет
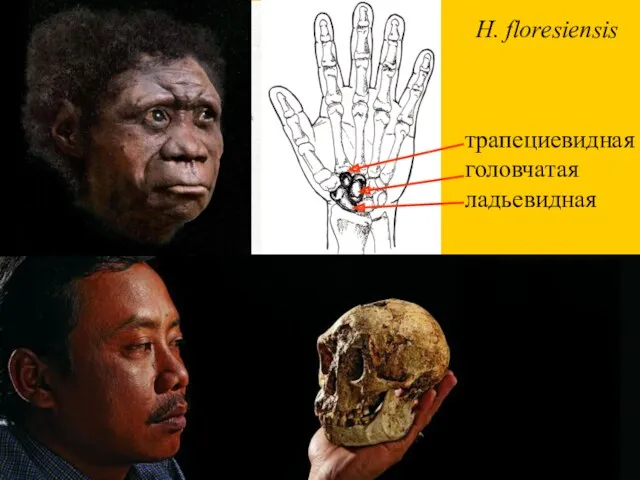
трапециевидная
головчатая
ладьевидная
H. floresiensis
трапециевидная
головчатая
ладьевидная
H. floresiensis
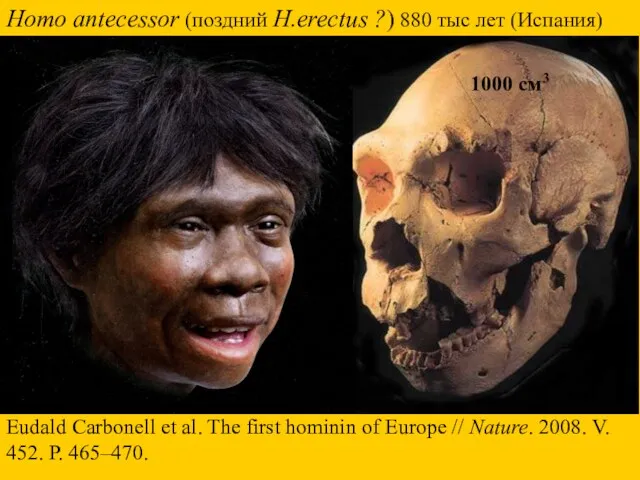
Homo antecessor (поздний H.erectus ?) 880 тыс лет (Испания)
Eudald Carbonell et
Homo antecessor (поздний H.erectus ?) 880 тыс лет (Испания)
Eudald Carbonell et

Homo heidelbergensis ~ 800 тыс.лет
Homo heidelbergensis ~ 800 тыс.лет

Homo heidelbergensis ~ 800-200 тыс.лет
1100-1400 см3
Homo heidelbergensis ~ 800-200 тыс.лет
1100-1400 см3
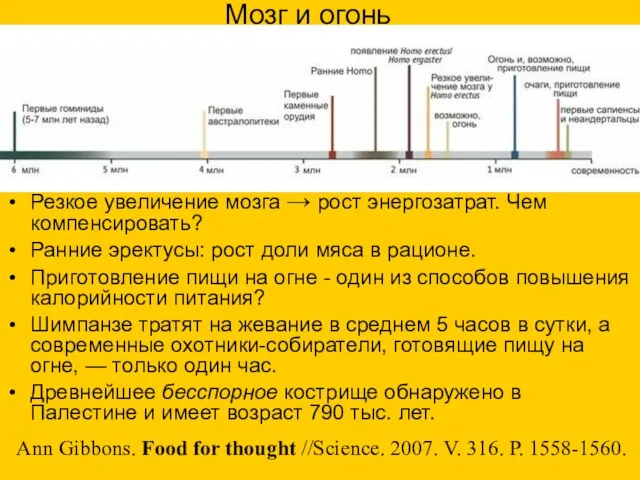
Мозг и огонь
Резкое увеличение мозга → рост энергозатрат. Чем компенсировать?
Ранние эректусы:
Мозг и огонь
Резкое увеличение мозга → рост энергозатрат. Чем компенсировать?
Ранние эректусы:

Слепки мозговой полости (эндокраны)
Australopithecus Homo erectus
Homo neanderthalensis Homo sapiens
Слепки мозговой полости (эндокраны)
Australopithecus Homo erectus
Homo neanderthalensis Homo sapiens

Homo neanderthalensis Homo sapiens
900-2000 см3
200 000 - 28 000 лет
Homo neanderthalensis Homo sapiens
900-2000 см3
200 000 - 28 000 лет

Homo neanderthalensis Homo sapiens
Homo neanderthalensis Homo sapiens


Неандертальцы были рыжими и светлокожими
У неандертальцев обнаружена мутация в гене меланокортинового
Неандертальцы были рыжими и светлокожими
У неандертальцев обнаружена мутация в гене меланокортинового

Lalueza-Fox et al. A melanocortin 1 receptor allele suggests varying pigmentation
Lalueza-Fox et al. A melanocortin 1 receptor allele suggests varying pigmentation

«… коренное неандертальское население Европы пало под натиском ближневосточных пришельцев значительно
«… коренное неандертальское население Европы пало под натиском ближневосточных пришельцев значительно
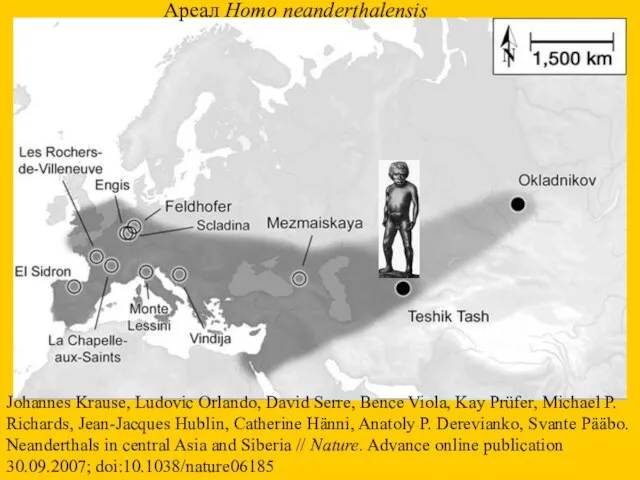
Johannes Krause, Ludovic Orlando, David Serre, Bence Viola, Kay Prüfer, Michael
Johannes Krause, Ludovic Orlando, David Serre, Bence Viola, Kay Prüfer, Michael

В Денисовой пещере (Алтай) в 2005 г. была найдена кость индивидуума,
В Денисовой пещере (Алтай) в 2005 г. была найдена кость индивидуума,
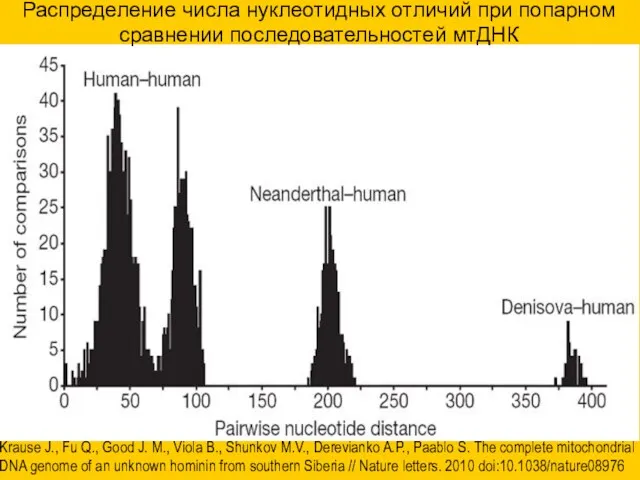
Распределение числа нуклеотидных отличий при попарном сравнении последовательностей мтДНК
Krause J., Fu
Распределение числа нуклеотидных отличий при попарном сравнении последовательностей мтДНК
Krause J., Fu
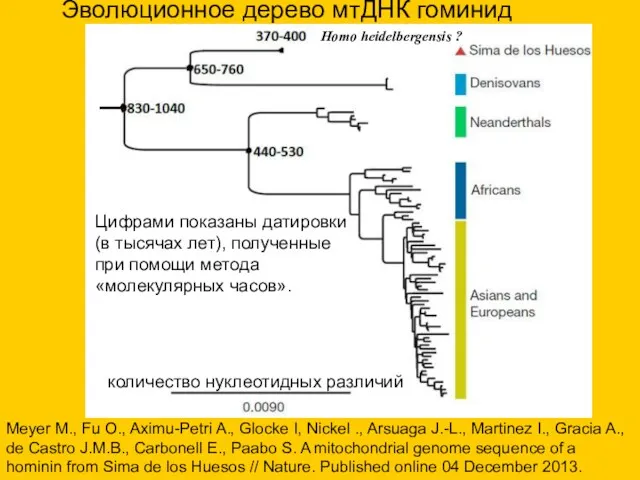
Цифрами показаны датировки (в тысячах лет), полученные при помощи метода «молекулярных
Цифрами показаны датировки (в тысячах лет), полученные при помощи метода «молекулярных
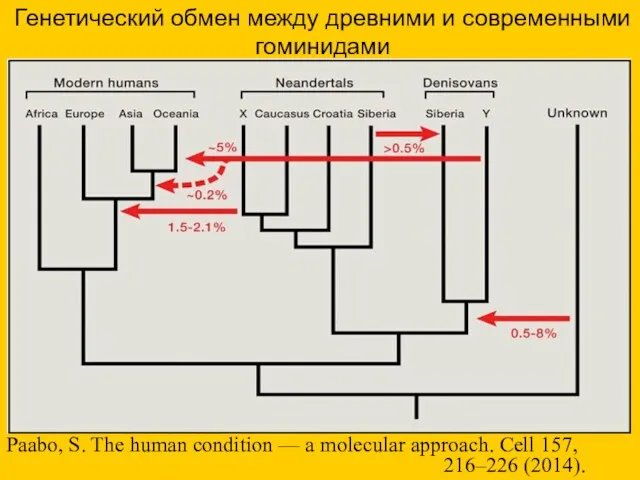
Генетический обмен между древними и современными гоминидами
Paabo, S. The human condition
Генетический обмен между древними и современными гоминидами
Paabo, S. The human condition
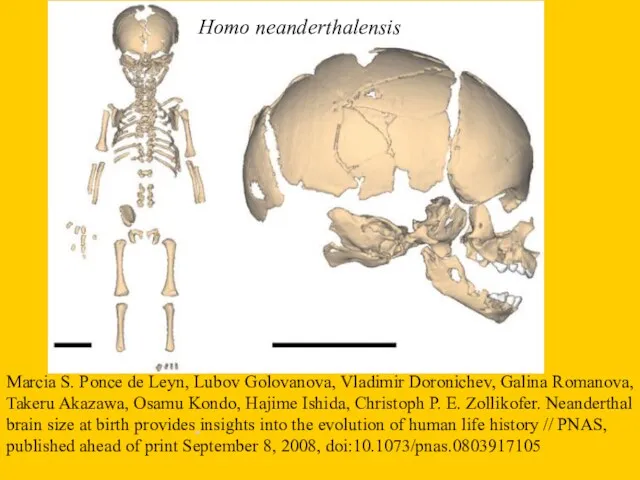
Homo neanderthalensis
Marcia S. Ponce de Leуn, Lubov Golovanova, Vladimir Doronichev,
Homo neanderthalensis
Marcia S. Ponce de Leуn, Lubov Golovanova, Vladimir Doronichev,
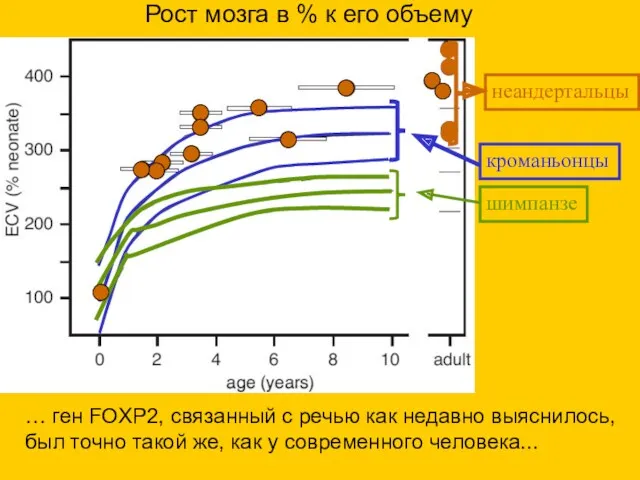
шимпанзе
кроманьонцы
неандертальцы
Рост мозга в % к его объему
… ген FOXP2, связанный с
шимпанзе
кроманьонцы
неандертальцы
Рост мозга в % к его объему
… ген FOXP2, связанный с
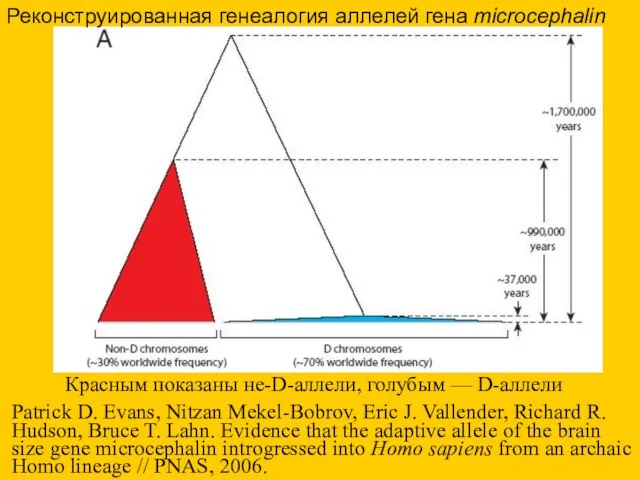
Реконструированная генеалогия аллелей гена microcephalin
Красным показаны не-D-аллели, голубым — D-аллели
Patrick D.
Реконструированная генеалогия аллелей гена microcephalin
Красным показаны не-D-аллели, голубым — D-аллели
Patrick D.

Черепа кроманьонцев
Черепа кроманьонцев
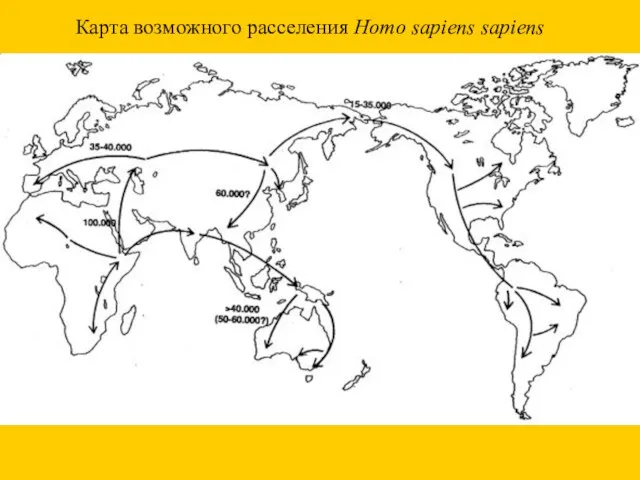
Карта возможного расселения Homo sapiens sapiens
Карта возможного расселения Homo sapiens sapiens
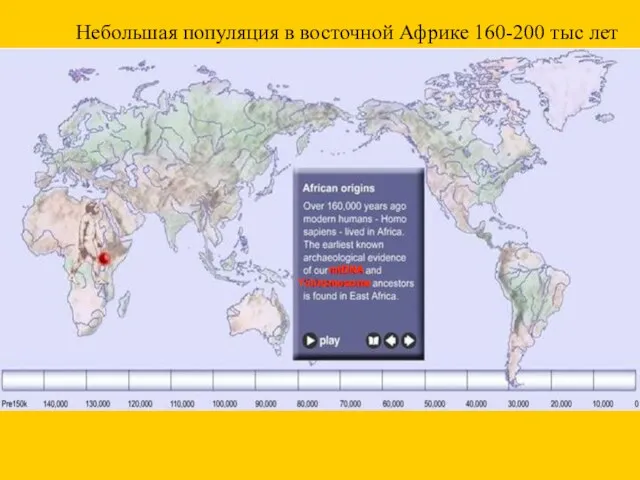
Небольшая популяция в восточной Африке 160-200 тыс лет
mtDNA
Ychromosome
Небольшая популяция в восточной Африке 160-200 тыс лет
mtDNA
Ychromosome
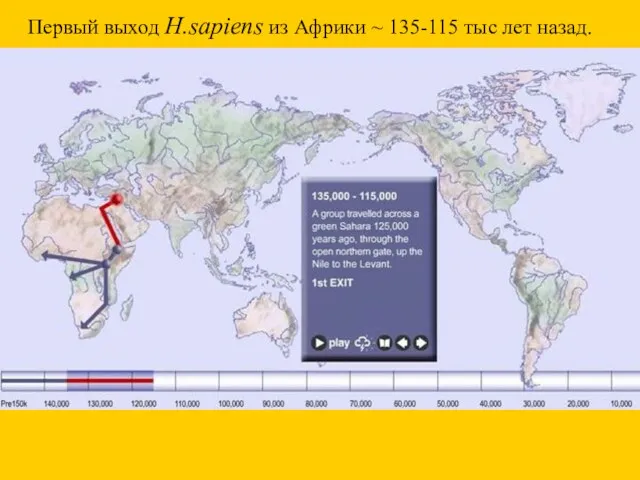
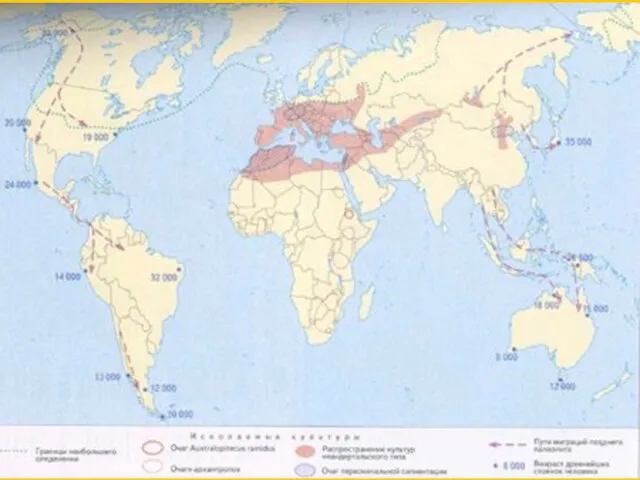
Первый выход H.sapiens из Африки ~ 135-115 тыс лет назад.
Первый выход H.sapiens из Африки ~ 135-115 тыс лет назад.
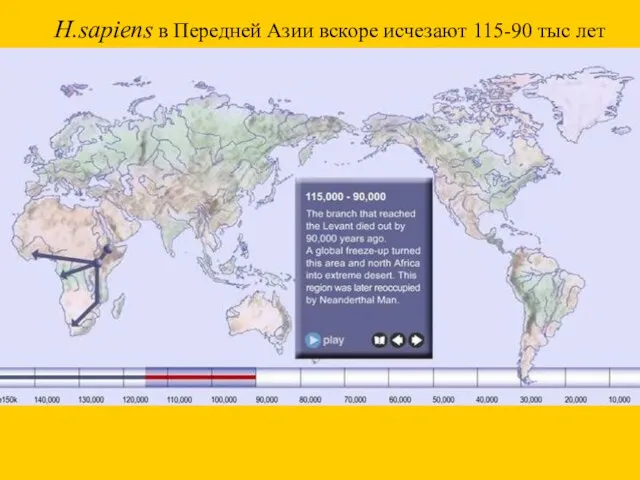
H.sapiens в Передней Азии вскоре исчезают 115-90 тыс лет
H.sapiens в Передней Азии вскоре исчезают 115-90 тыс лет
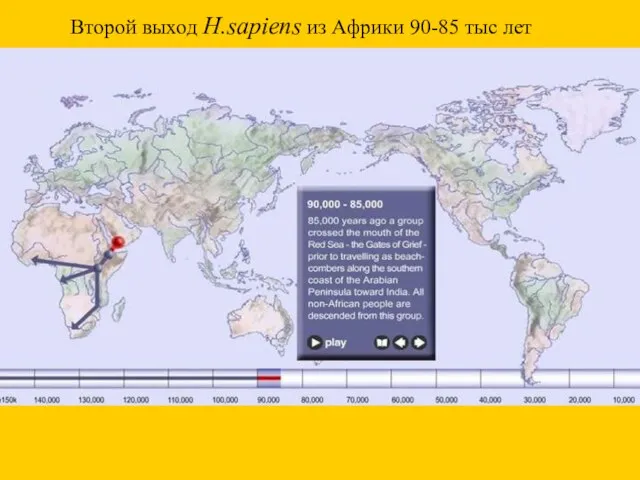
Второй выход H.sapiens из Африки 90-85 тыс лет
Второй выход H.sapiens из Африки 90-85 тыс лет
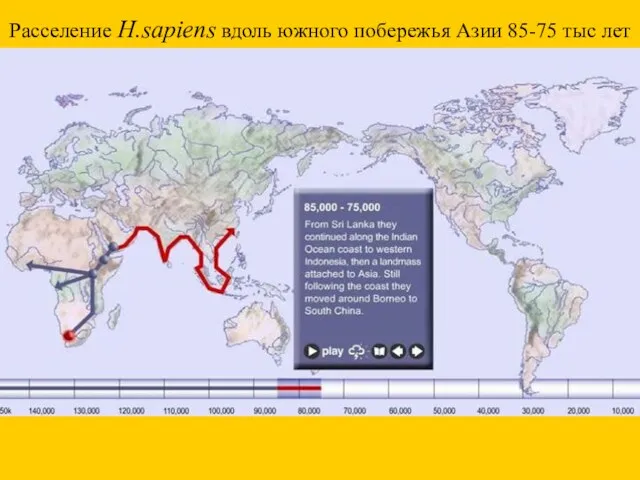
Расселение H.sapiens вдоль южного побережья Азии 85-75 тыс лет
Расселение H.sapiens вдоль южного побережья Азии 85-75 тыс лет
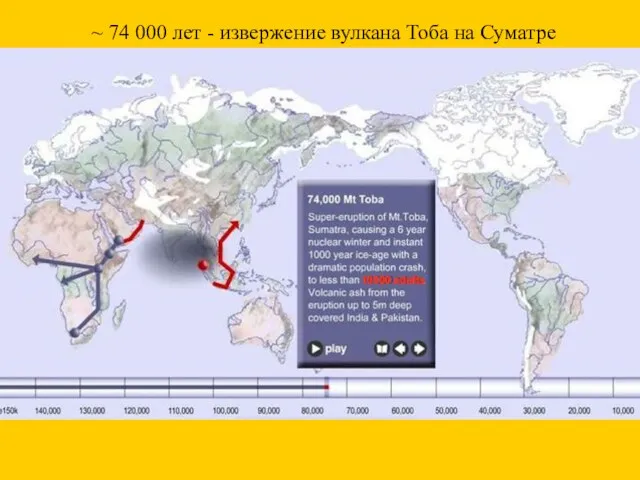
~ 74 000 лет - извержение вулкана Тоба на Суматре
10000 adults
~ 74 000 лет - извержение вулкана Тоба на Суматре
10000 adults
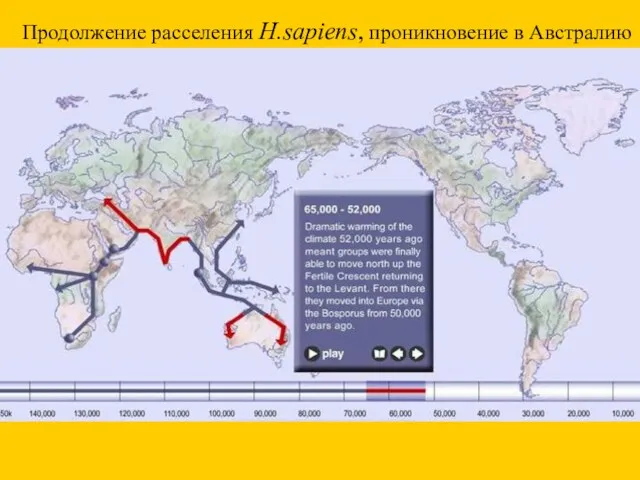
Продолжение расселения H.sapiens, проникновение в Австралию
Продолжение расселения H.sapiens, проникновение в Австралию
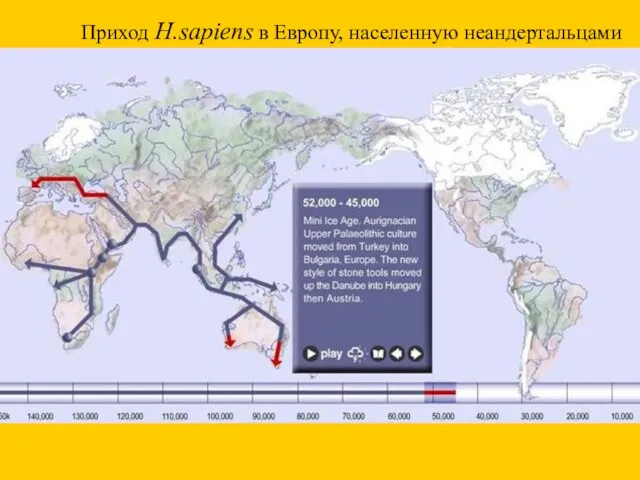
Приход H.sapiens в Европу, населенную неандертальцами
Приход H.sapiens в Европу, населенную неандертальцами
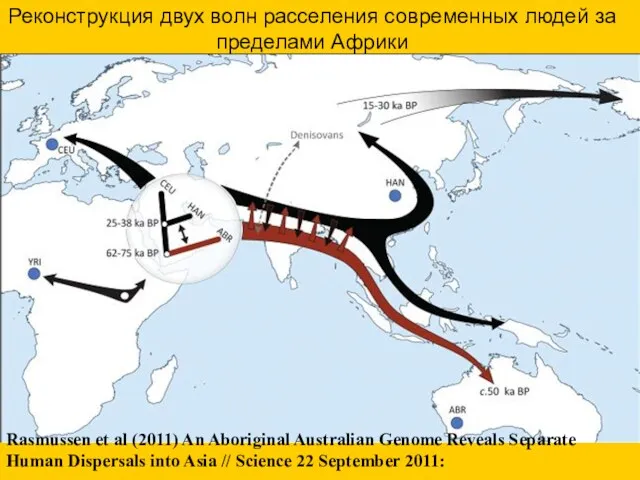
Rasmussen et al (2011) An Aboriginal Australian Genome Reveals Separate Human
Rasmussen et al (2011) An Aboriginal Australian Genome Reveals Separate Human

Заселение сапиенсами Европы
Колонизация Центральной и Западной Европы: 46-41 тыс. лет назад.
Скорость
Заселение сапиенсами Европы
Колонизация Центральной и Западной Европы: 46-41 тыс. лет назад.
Скорость
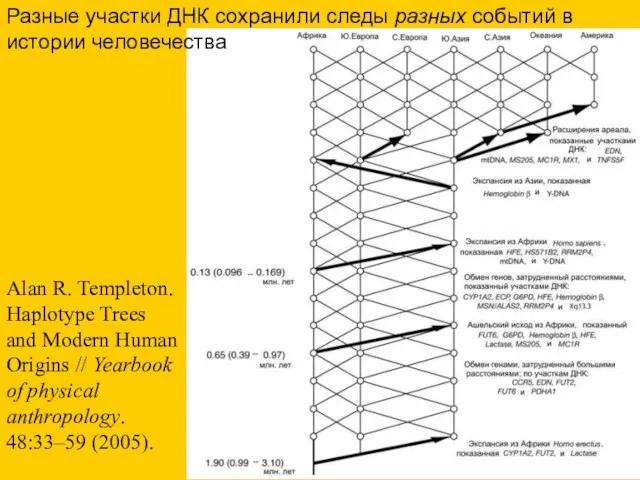
Alan R. Templeton. Haplotype Trees and Modern Human Origins // Yearbook
Alan R. Templeton. Haplotype Trees and Modern Human Origins // Yearbook
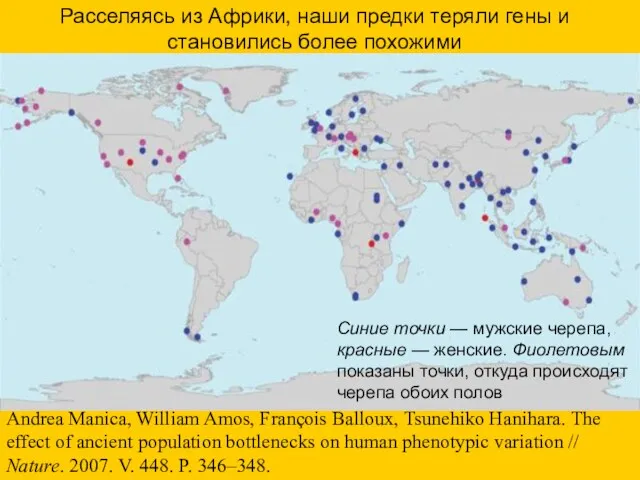
Расселяясь из Африки, наши предки теряли гены и становились более похожими
Синие
Расселяясь из Африки, наши предки теряли гены и становились более похожими
Синие
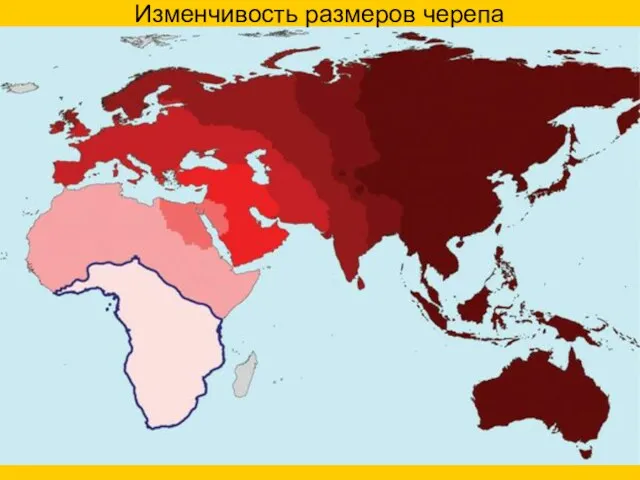
Изменчивость размеров черепа
Изменчивость размеров черепа
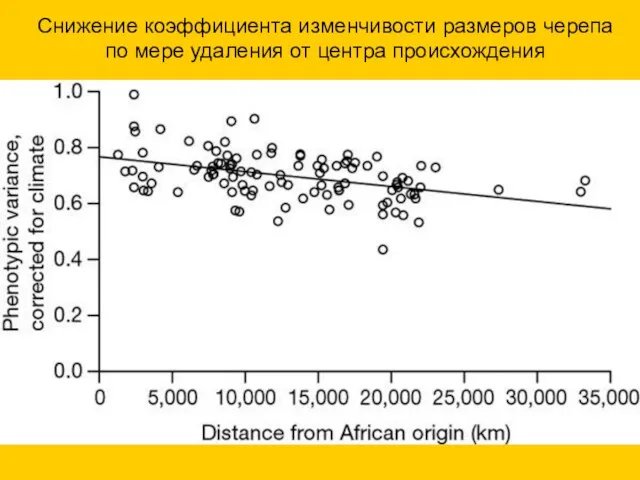
Снижение коэффициента изменчивости размеров черепа по мере удаления от центра происхождения
Снижение коэффициента изменчивости размеров черепа по мере удаления от центра происхождения
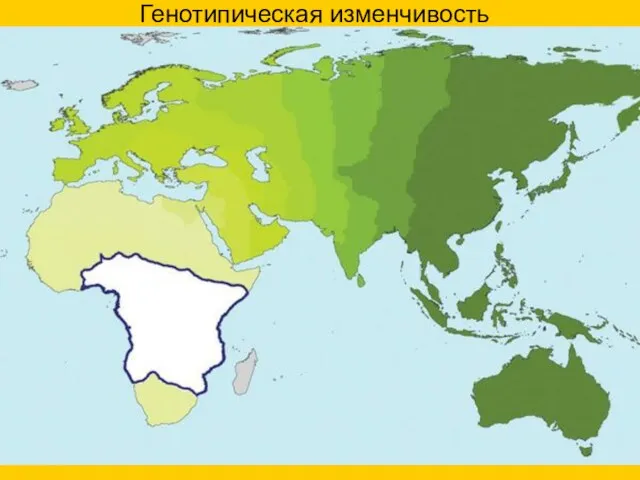
Генотипическая изменчивость
Генотипическая изменчивость
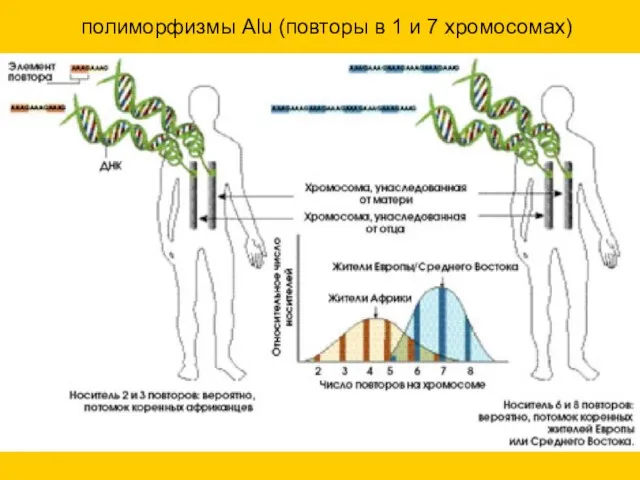
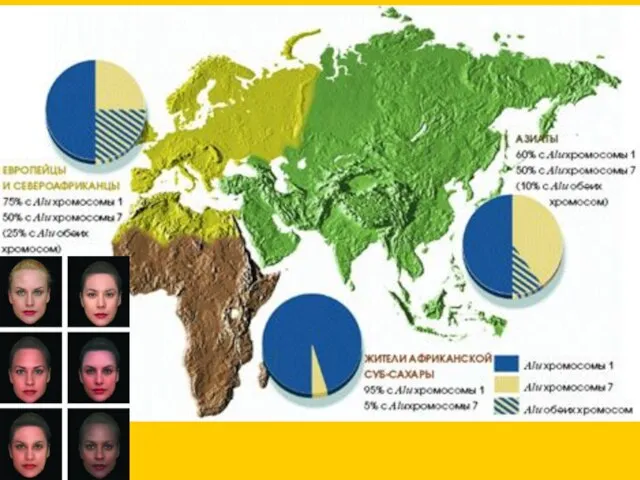
полиморфизмы Alu (повторы в 1 и 7 хромосомах)
полиморфизмы Alu (повторы в 1 и 7 хромосомах)

Сравнение уровня активности 907 генов у людей, шимпанзе, орангутанов и
Сравнение уровня активности 907 генов у людей, шимпанзе, орангутанов и

Эволюционная психология
Эволюционная психология
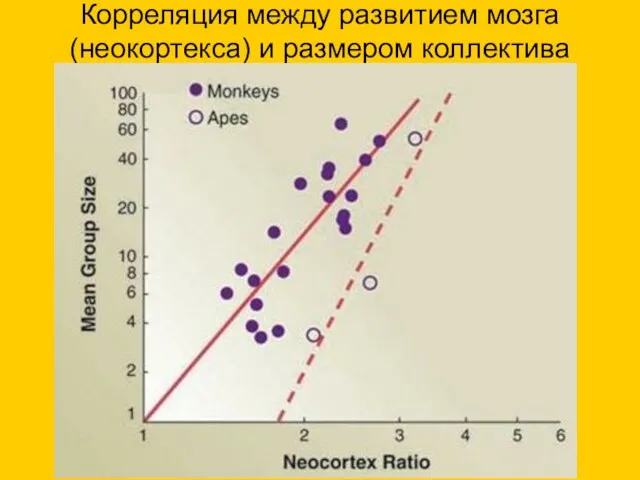
Корреляция между развитием мозга (неокортекса) и размером коллектива
Корреляция между развитием мозга (неокортекса) и размером коллектива
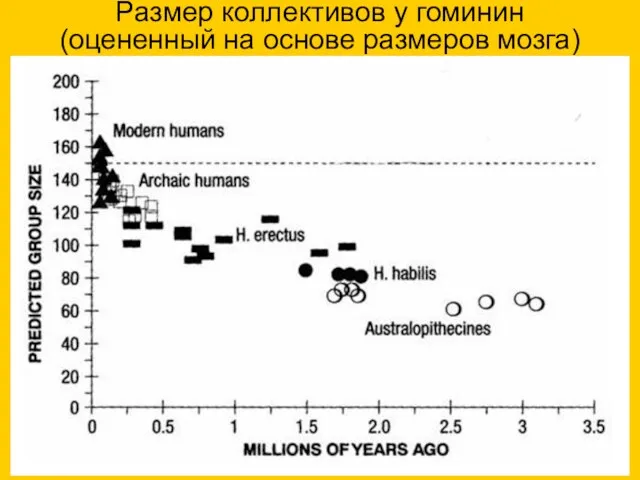
Размер коллективов у гоминин (оцененный на основе размеров мозга)
Размер коллективов у гоминин (оцененный на основе размеров мозга)
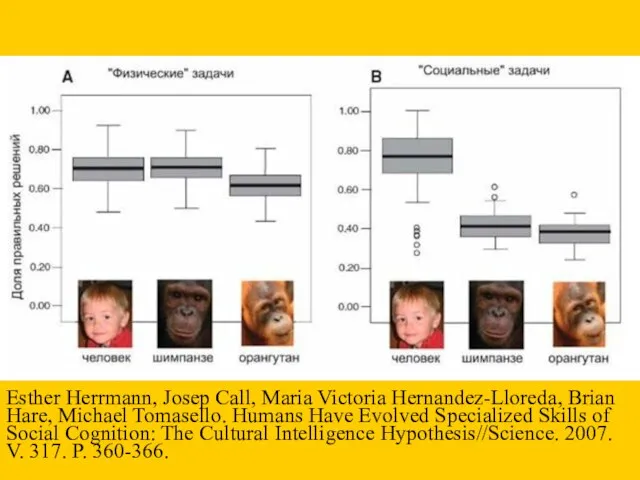
Esther Herrmann, Josep Call, Maria Victoria Hernandez-Lloreda, Brian Hare, Michael Tomasello.
Esther Herrmann, Josep Call, Maria Victoria Hernandez-Lloreda, Brian Hare, Michael Tomasello.
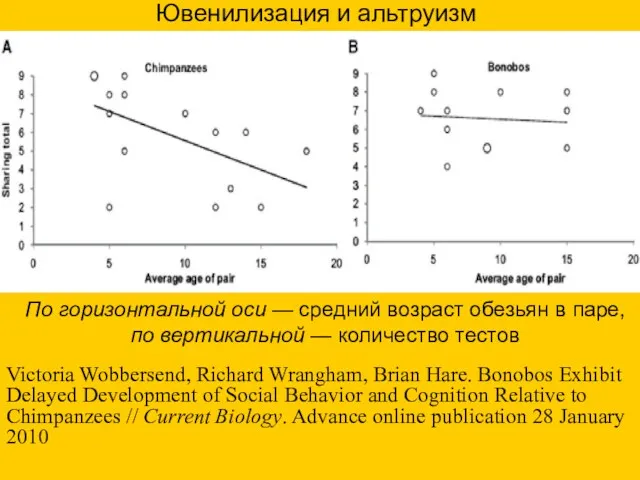
По горизонтальной оси — средний возраст обезьян в паре, по вертикальной
По горизонтальной оси — средний возраст обезьян в паре, по вертикальной

“Культурное наследование” у животных
Умение колоть орехи в некоторых популяциях шимпанзе тысячелетиями
“Культурное наследование” у животных
Умение колоть орехи в некоторых популяциях шимпанзе тысячелетиями
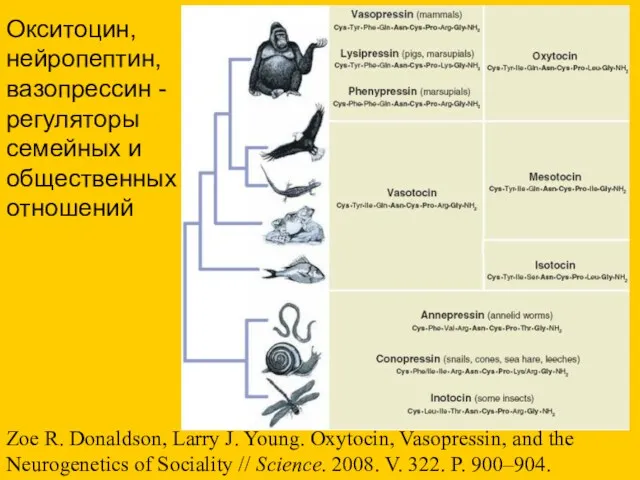
Zoe R. Donaldson, Larry J. Young. Oxytocin, Vasopressin, and the Neurogenetics
Zoe R. Donaldson, Larry J. Young. Oxytocin, Vasopressin, and the Neurogenetics

Близнецовый анализ показал, что склонность к добрым поступкам, доверчивость и благодарность
Близнецовый анализ показал, что склонность к добрым поступкам, доверчивость и благодарность

Реципрокный (взаимный) альтруизм
Стратегия:
Помоги другому, и он в будущем поможет тебе.
Реципрокный (взаимный) альтруизм
Стратегия:
Помоги другому, и он в будущем поможет тебе.
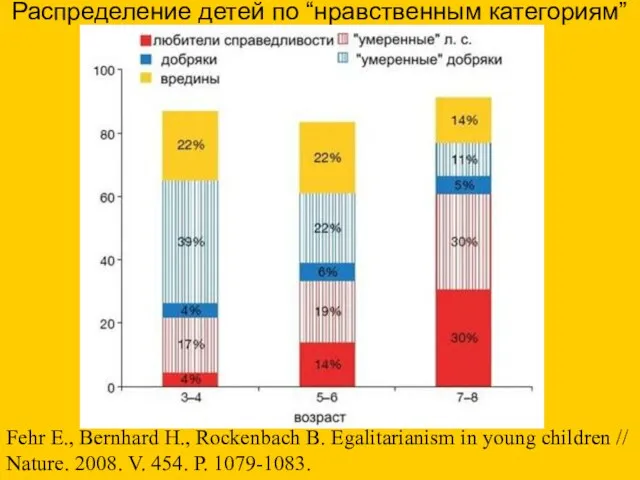
Распределение детей по “нравственным категориям”
Fehr Е., Bernhard Н., Rockenbach В. Egalitarianism
Распределение детей по “нравственным категориям”
Fehr Е., Bernhard Н., Rockenbach В. Egalitarianism

Альтруизм среди “своих” и враждебность к чужакам: две стороны одной медали
Дети
Альтруизм среди “своих” и враждебность к чужакам: две стороны одной медали
Дети

Межгрупповые войны - причина альтруизма?
Уровень межгрупповой агрессии у палеолитических охотников-собирателей был
Межгрупповые войны - причина альтруизма?
Уровень межгрупповой агрессии у палеолитических охотников-собирателей был

р —доля альтруистов, w— численность популяции
S. Chuang, Olivier Rivoire, Stanislas Leibler.
р —доля альтруистов, w— численность популяции
S. Chuang, Olivier Rivoire, Stanislas Leibler.

Правило Гамильтона:
«ген альтруизма» будет поддержан отбором и распространится в популяции,
Правило Гамильтона: «ген альтруизма» будет поддержан отбором и распространится в популяции,
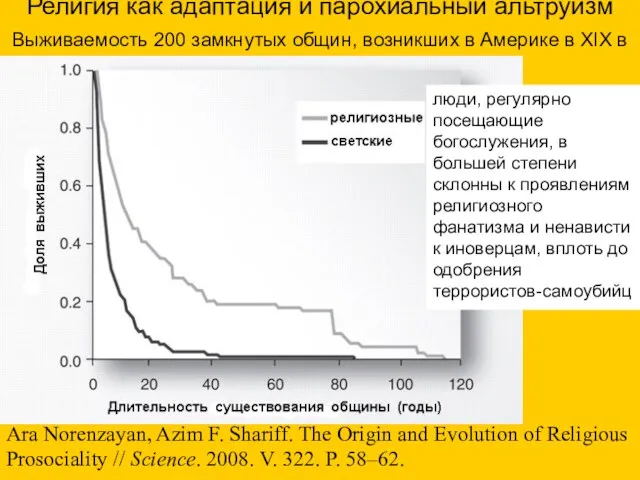
Религия как адаптация и парохиальный альтруизм
Выживаемость 200 замкнутых общин, возникших в
Религия как адаптация и парохиальный альтруизм
Выживаемость 200 замкнутых общин, возникших в

Резникова Ж.И. Социальное обучение у животных // Природа. 2009. №5.
«...Учиться у
Резникова Ж.И. Социальное обучение у животных // Природа. 2009. №5.
«...Учиться у

Есть мнение, что главным стимулом для развития речи у наших предков
Есть мнение, что главным стимулом для развития речи у наших предков

У людей, по-видимому, есть врожденные психологические свойства (“предрасположенности”), направленные на эффективное
У людей, по-видимому, есть врожденные психологические свойства (“предрасположенности”), направленные на эффективное

“Комплекс гоминизации” (А.А. Зубов)
высокоразвитый мозг (энцефализация);
анатомические особенности руки и возможность
“Комплекс гоминизации” (А.А. Зубов)
высокоразвитый мозг (энцефализация);
анатомические особенности руки и возможность
 Ткани из растений. Классификация волокон
Ткани из растений. Классификация волокон Основы здорового питания собак и кошек
Основы здорового питания собак и кошек Рефлексы
Рефлексы Презентация по биологии Класс Птицы для 7 класса.
Презентация по биологии Класс Птицы для 7 класса. Земноводні України
Земноводні України Презентация Экологический кризис планеты
Презентация Экологический кризис планеты Взаимосвязь путей обмена веществ
Взаимосвязь путей обмена веществ Теории происхождения человека
Теории происхождения человека Первая помощь при ушибах, вывихах, переломах
Первая помощь при ушибах, вывихах, переломах Естественный отбор
Естественный отбор A Leaf
A Leaf Рефлексы. Условные и безусловные рефлексы. 8 класс
Рефлексы. Условные и безусловные рефлексы. 8 класс Разнообразие птиц
Разнообразие птиц Бұлшық ет
Бұлшық ет Физико-химические основы патологии клетки. Митохондрии и апоптоз
Физико-химические основы патологии клетки. Митохондрии и апоптоз Транспорт веществ через многомембранные системы. Вторичный активный транспорт
Транспорт веществ через многомембранные системы. Вторичный активный транспорт РАЗРАБОТКА ПРЕЗЕНТАЦИИ УРОКА ПО БИОЛОГИИ В 11 КЛАССЕ НА ТЕМУ: Движущие силы антропогенеза. Ф. Энгельс о роли труда в процессе превращения обезьяны в человека. Развитие членораздельной речи, сознания и общественных отношений в становлении ч
РАЗРАБОТКА ПРЕЗЕНТАЦИИ УРОКА ПО БИОЛОГИИ В 11 КЛАССЕ НА ТЕМУ: Движущие силы антропогенеза. Ф. Энгельс о роли труда в процессе превращения обезьяны в человека. Развитие членораздельной речи, сознания и общественных отношений в становлении ч Живое клеточный уровень. Живое организменный уровень ботаника. Живое организменный уровень зоология
Живое клеточный уровень. Живое организменный уровень ботаника. Живое организменный уровень зоология Кости черепа
Кости черепа Презентации к теме Химический состав клетки (5 класс)
Презентации к теме Химический состав клетки (5 класс) Задания ОГЭ по теме Размножение организмов
Задания ОГЭ по теме Размножение организмов Маргаритки.Двухлетние цветочные растения.
Маргаритки.Двухлетние цветочные растения. Developmental Psychology. Lecture 2. The Interaction of genes and Environment in Human Development
Developmental Psychology. Lecture 2. The Interaction of genes and Environment in Human Development Nobel prize in physiology or medicine
Nobel prize in physiology or medicine Современные представления об эволюции органического мира
Современные представления об эволюции органического мира Растительные ткани
Растительные ткани Проблема ГМО-продуктів – проблема вибору
Проблема ГМО-продуктів – проблема вибору Введение в гистологию
Введение в гистологию